
Distribution of Nereilinum murmanicum (Annelida, Siboglinidae) in the Barents Sea in the Context of Its Oil and Gas Potential

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
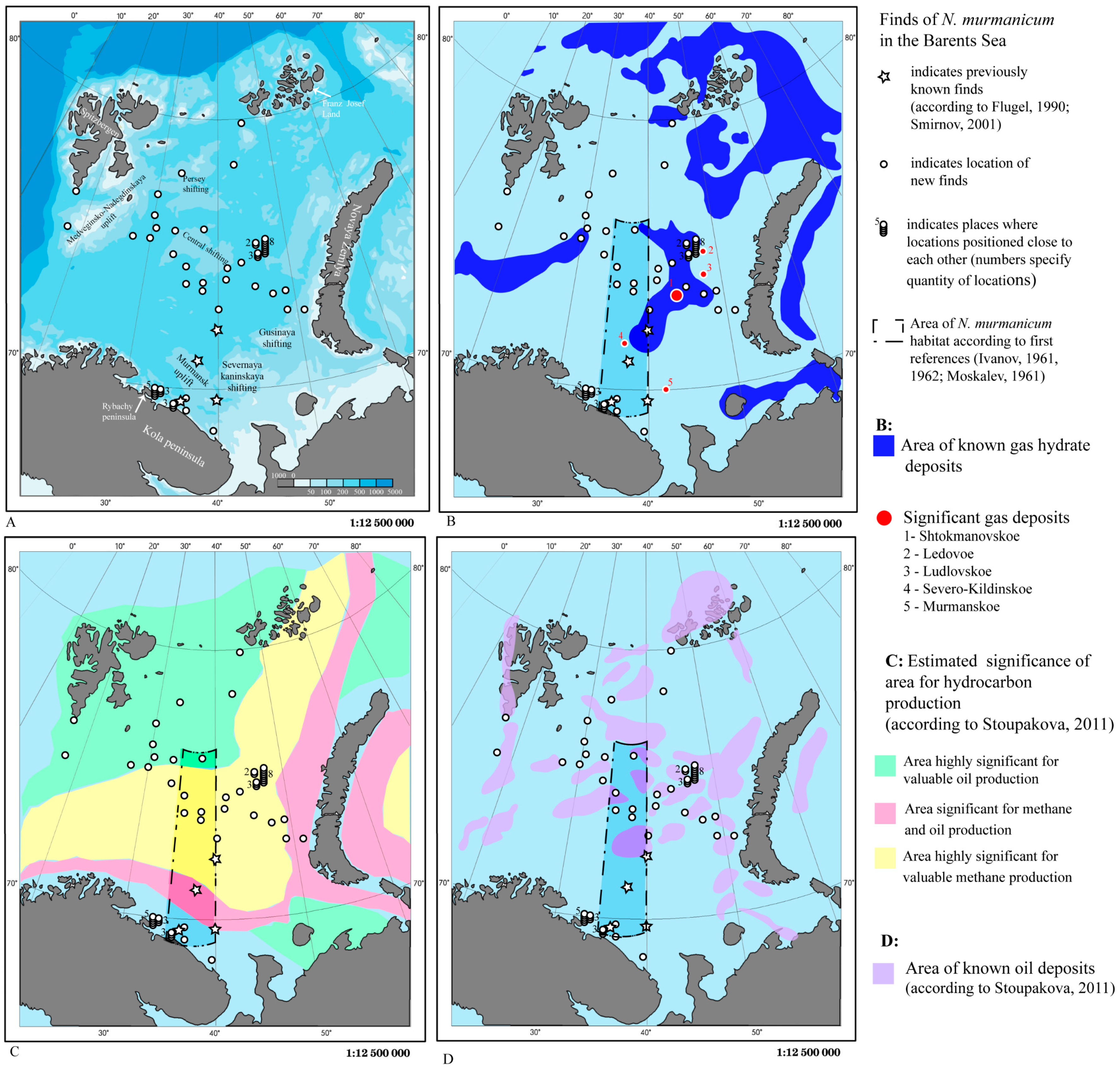
3. Results
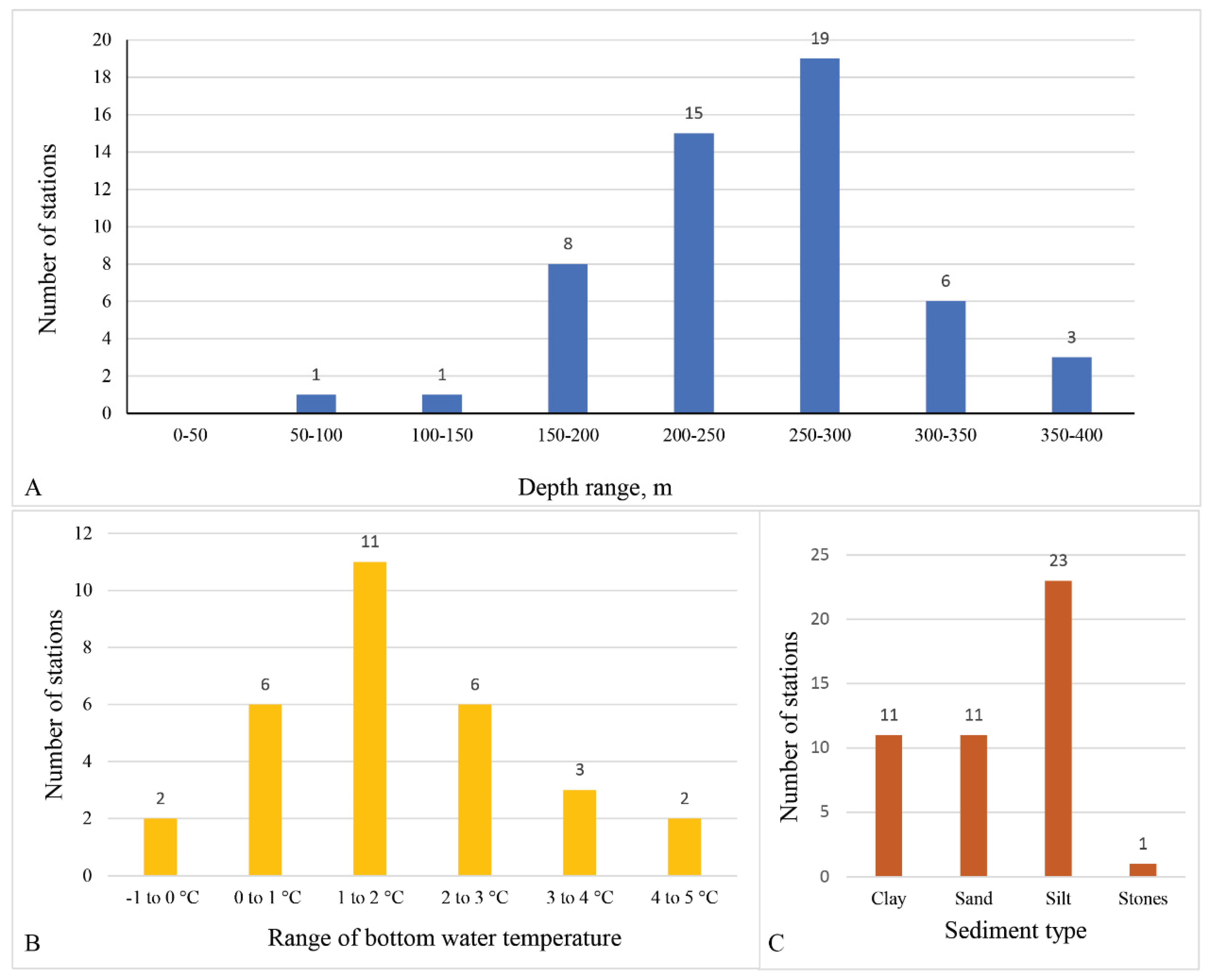
3.1. Geographical and Bathymetric Distribution
3.2. Relation of Distribution to Abiotic Factors
4. Discussion
4.1. Geographical and Bathymetric Distribution
4.2. Environment
4.3. General Connection of Siboglinids Findings to Hydrocarbons
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Southward, E.C. Bacterial symbionts in Pogonophora. J. Mar. Biol. Assoc. U. K. 1982, 62, 889–906. [Google Scholar] [CrossRef]
- Fisher, C., Jr.; Childress, J. Substrate oxidation by trophosome tissue from Riftia pachyptila Jones (Phylum pogonophora). Mar. Biol. Lett. 1984, 5, 171–183. [Google Scholar]
- Katz, S.; Klepal, W.; Bright, M. The Osedax Trophosome: Organization and Ultrastructure. Biol. Bull. 2011, 220, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Halanych, K.M.; Feldman, R.A.; Vrijenhoek, R.C. Molecular Evidence that Sclerolinum brattstromi Is Closely Related to Vestimentiferans, not to Frenulate Pogonophorans (Siboglinidae, Annelida). Biol. Bull. 2001, 201, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Rousset, V.; Rouse, G.W.; Siddall, M.E.; Tillier, A.; Pleijel, F. The phylogenetic position of Siboglinidae (Annelida) inferred from 18S rRNA, 28S rRNA and morphological data. Cladistics 2004, 20, 518–533. [Google Scholar] [CrossRef]
- Hilário, A.; Capa, M.; Dahlgren, T.G.; Halanych, K.M.; Little, C.T.S.; Thornhill, D.J.; Verna, C.; Glover, A.G. New Perspectives on the Ecology and Evolution of Siboglinid Tubeworms. PLoS ONE 2011, 6, e16309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaseva, N.; Rimskaya-Korsakova, N.N.; Galkin, S.V.; Malakhov, V.V. Taxonomy, geographical and bathymetric distribution of vestimentiferan tubeworms (Annelida, Siboglinidae). Biol. Bull. 2016, 43, 937–969. [Google Scholar] [CrossRef]
- Li, Y.; Kocot, K.M.; Whelan, N.V.; Santos, S.R.; Waits, D.; Thornhill, D.J.; Halanych, K.M. Phylogenomics of tubeworms (Siboglinidae, Annelida) and comparative performance of different reconstruction methods. Zool. Scr. 2016, 46, 200–213. [Google Scholar] [CrossRef]
- Rouse, G.W.; Goffredi, S.K.; Vrijenhoek, R.C. Osedax: Bone-Eating Marine Worms with Dwarf Males. Science 2004, 305, 668–671. [Google Scholar] [CrossRef] [Green Version]
- Goffredi, S.K.; Orphan, V.; Rouse, G.; Jahnke, L.; Embaye, T.; Turk, K.; Lee, R.; Vrijenhoek, R.C. Evolutionary innovation: A bone-eating marine symbiosis. Environ. Microbiol. 2005, 7, 1369–1378. [Google Scholar] [CrossRef]
- Goffredi, S.K.; Johnson, S.B.; Vrijenhoek, R.C. Genetic Diversity and Potential Function of Microbial Symbionts Associated with Newly Discovered Species of Osedax Polychaete Worms. Appl. Environ. Microbiol. 2007, 73, 2314–2323. [Google Scholar] [CrossRef] [Green Version]
- Schmaljohann, R.; Faber, E.; Whiticar, M.; Dando, P. Co-existence of methane- and sulphur-based endosymbioses between bacteria and invertebrates at a site in the Skagerrak. Mar. Ecol. Prog. Ser. 1990, 61, 119–124. [Google Scholar] [CrossRef]
- Pimenov, N.V.; Savvichev, A.S.; Rusanov, I.I.; Lein, A.Y.; Ivanov, M.V. Microbiological Processes of the Carbon and Sulfur Cycles at Cold Methane Seeps of the North Atlantic. Microbiology 2000, 69, 709–720. [Google Scholar] [CrossRef]
- Kimura, H.; Sato, M.; Sasayama, Y.; Naganuma, T. Molecular Characterization and In Situ Localization of Endosymbiotic 16S Ribosomal RNA and RuBisCO Genes in the Pogonophoran Tissue. Mar. Biotechnol. 2003, 5, 261–269. [Google Scholar] [CrossRef]
- Naganuma, T.; Elsaied, H.E.; Hoshii, D.; Kimura, H. Bacterial Endosymbioses of Gutless Tube-Dwelling Worms in Nonhydrothermal Vent Habitats. Mar. Biotechnol. 2005, 7, 416–428. [Google Scholar] [CrossRef]
- Lösekann, T.; Robador, A.; Niemann, H.; Knittel, K.; Boetius, A.; Dubilier, N. Endosymbioses between bacteria and deep-sea siboglinid tubeworms from an Arctic Cold Seep (Haakon Mosby Mud Volcano, Barents Sea). Environ. Microbiol. 2008, 10, 3237–3254. [Google Scholar] [CrossRef]
- Felbeck, H. Chemoautotrophic Potential of the Hydrothermal Vent Tube Worm, Riftia pachyptila Jones (Vestimentifera). Science 1981, 213, 336–338. [Google Scholar] [CrossRef]
- Distel, D.; Lane, D.J.; Olsen, G.J.; Giovannoni, S.J.; Pace, B.; Pace, N.R.; Stahl, D.A.; Felbeck, H. Sulfur-oxidizing bacterial endosymbionts: Analysis of phylogeny and specificity by 16S rRNA sequences. J. Bacteriol. 1988, 170, 2506–2510. [Google Scholar] [CrossRef] [Green Version]
- Robidart, J.C.; Bench, S.R.; Feldman, R.A.; Novoradovsky, A.; Podell, S.B.; Gaasterland, T.; Allen, E.E.; Felbeck, H. Metabolic versatility of the Riftia pachyptila endosymbiont revealed through metagenomics. Environ. Microbiol. 2008, 10, 727–737. [Google Scholar] [CrossRef]
- Reveillaud, J.; Anderson, R.; Reves-Sohn, S.; Cavanaugh, C.; Huber, J.A. Metagenomic investigation of vestimentiferan tubeworm endosymbionts from Mid-Cayman Rise reveals new insights into metabolism and diversity. Microbiome 2018, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Webb, M. A new bitentagulate pogonophoran from hardangerfjorden, norway. Sarsia 1964, 15, 49–56. [Google Scholar] [CrossRef]
- Webb, M. Additional notes on Sclerolinum brattstromi (Pogonophora) and the establishment of a new family, Sclerolinidae. Sarsia 1964, 16, 47–58. [Google Scholar] [CrossRef]
- Southward, E.C. On some Pogonophora from the Caribbean and the Gulf of Mexico. Bull. Mar. Sci. 1972, 22, 739–776. [Google Scholar]
- Smirnov, R.V. Two new species of Pogonophora from the arctic mud volcano off northwestern Norway. Sarsia 2000, 85, 141–150. [Google Scholar] [CrossRef]
- Smirnov, R.V. Morphological characters and classification of the subclass Monilifera (Pogonophora) and the problem of evolution of the bridle in pogonophorans. Russ. J. Mar. Biol. 2008, 34, 359–368. [Google Scholar] [CrossRef]
- Southward, A.; Southward, E.C.; Dando, P.; Barrett, R.; Ling, R. Chemoautotrophic function of bacterial symbionts in small Pogonophora. J. Mar. Biol. Assoc. U. K. 1986, 66, 415–437. [Google Scholar] [CrossRef]
- Dando, P.; Southward, A.; Southward, E.; Barrett, R. Possible energy sources for chemoautotrophic prokaryotes symbiotic with invertebrates from a Norwegian fjord. Ophelia 1986, 26, 135–150. [Google Scholar] [CrossRef]
- Schmaljohann, R.; Flügel, H.J. Methane-oxidizing bacteria in Pogonophora. Sarsia 1987, 72, 91–98. [Google Scholar] [CrossRef]
- Kubota, N.; Kanemori, M.; Sasayama, Y.; Aida, M.; Fukumori, Y. Identification of Endosymbionts in Oligobrachia mashikoi (Siboglinidae, Annelida). Microbes Environ. 2007, 22, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Aida, M.; Kanemori, M.; Kubota, N.; Matada, M.; Sasayama, Y.; Fukumori, Y. Distribution and Population of Free-Living Cells Related to Endosymbiont a Harbored in Oligobrachia mashikoi (a Siboglinid Polychaete) Inhabiting Tsukumo Bay. Microbes Environ. 2008, 23, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Savvichev, A.S.; Kadnikov, V.V.; Kravchishina, M.; Galkin, S.V.; Novigatskii, A.N.; Sigalevich, P.A.; Merkel, A.Y.; Ravin, N.V.; Pimenov, N.V.; Flint, M.V. Methane as an Organic Matter Source and the Trophic Basis of a Laptev Sea Cold Seep Microbial Community. Geomicrobiol. J. 2018, 35, 411–423. [Google Scholar] [CrossRef]
- Aharon, P.; Fu, B. Sulfur and oxygen isotopes of coeval sulfate–sulfide in pore fluids of cold seep sediments with sharp redox gradients. Chem. Geol. 2003, 195, 201–218. [Google Scholar] [CrossRef]
- Boetius, A.; Ravenschlag, K.; Schubert, C.J.; Rickert, D.; Widdel, F.; Gieseke, A.; Amann, R.; Jorgensen, B.B.; Witte, U.; Pfannkuche, O. A marine microbial consortium apparently mediating anaerobic oxidation of methane. Nature 2000, 407, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Joye, S.; Boetius, A.; Orcutt, B.; Montoya, J.; Schulz-Vogt, H.; Erickson, M.J.; Lugo, S.K. The anaerobic oxidation of methane and sulfate reduction in sediments from Gulf of Mexico cold seeps. Chem. Geol. 2004, 205, 219–238. [Google Scholar] [CrossRef]
- Levin, L.A. Ecology of Cold Seep Sediments: Interactions of Fauna with Flow, Chemistry and Microbes. In Oceanography and Marine Biology; CRC Press: Beijing, China, 2005; pp. 11–56. [Google Scholar] [CrossRef]
- Dattagupta, S.; Miles, L.L.; Barnabei, M.S.; Fisher, C.R. The hydrocarbon seep tubeworm Lamellibrachia luymesi primarily eliminates sulfate and hydrogen ions across its roots to conserve energy and ensure sulfide supply. J. Exp. Biol. 2006, 209, 3795–3805. [Google Scholar] [CrossRef] [Green Version]
- Gautier, D.L.; Bird, K.J.; Charpentier, R.R.; Grantz, A.; Houseknecht, D.W.; Klett, T.R.; Moore, T.E.; Pitman, J.K.; Schenk, C.J.; Schuenemeyer, J.H.; et al. Assessment of Undiscovered Oil and Gas in the Arctic. Science 2009, 324, 1175–1179. [Google Scholar] [CrossRef] [Green Version]
- Gautier, D.L.; Bird, K.J.; Charpentier, R.R.; Grantz, A.; Houseknecht, D.W.; Klett, T.R.; Moore, T.E.; Pitman, J.K.; Schenk, C.J.; Schuenemeyer, J.H.; et al. Chapter 9 Oil and gas resource potential north of the Arctic Circle. Geol. Soc. Lond. Mem. 2011, 35, 151–161. [Google Scholar] [CrossRef]
- Spencer, A.M.; Embry, A.F.; Gautier, D.L.; Stoupakova, A.V.; Sørensen, K. Chapter 1 An overview of the petroleum geology of the Arctic. Geol. Soc. Lond. Mem. 2011, 35, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Max, M.D.; Johnson, A.H.; Dillon, W.P. Natural Gas Hydrate-Arctic Ocean Deepwater Resource Potential; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Giustiniani, M.; Tinivella, U.; Jakobsson, M.; Rebesco, M. Arctic Ocean Gas Hydrate Stability in a Changing Climate. J. Geol. Res. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.V. Deux genres nouveaux de Pogonophores diplobrachiaux Nereilinum et Siboglinoides. Cah. Biol. Mar. 1961, 2, 381–397. [Google Scholar]
- Georgieva, M.N.; Wiklund, H.; Bell, J.B.; Eilertsen, M.H.; Mills, R.A.; Little, C.T.S.; Glover, A.G. A chemosynthetic weed: The tubeworm Sclerolinum contortum is a bipolar, cosmopolitan species. BMC Evol. Biol. 2015, 15, 280. [Google Scholar] [CrossRef] [Green Version]
- Moskalev, L.I. Pogonophora from the Barents sea. Dokl. Akad. Nauk AN USSR 1961, 137, 730–731. [Google Scholar]
- Ivanov, A.V. Pogonophora; Academic Press: London, UK, 1963; pp. 1–479. [Google Scholar]
- Smirnov, R.V. Systematics and Morphology of the Pogonophores of the Arctic and Southern Oceans; Zoological Institute of Russian Academy of Sciences: Saint Petersburg, Russia, 2001. [Google Scholar]
- Flügel, H.J. A new species ofsiboglinum(pogonophora) from the north atlantic and notes onnereilinum murmanicumivanov. Sarsia 1990, 75, 233–241. [Google Scholar] [CrossRef]
- Dando, P.; Southward, A.; Southward, E.; Lamont, P.; Harvey, R. Interactions between sediment chemistry and frenulate pogonophores (Annelida) in the north-east Atlantic. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2008, 55, 966–996. [Google Scholar] [CrossRef]
- Southward, E.C.; Schulze, A.; Gardiner, S.L. Pogonophora (Annelida): Form and function. Hydrobiologia 2005, 535, 227–251. [Google Scholar] [CrossRef]
- Southward, E.C. Pogonophora. In Microscopic Anatomy of Invertebrates, Vol. 12: Onychophora, Chilopoda, and Lesser Protostomata; Frederick, W., Harrison, M.E.R., Eds.; Wiley-Liss: New York, NY, USA, 1993; pp. 327–369. [Google Scholar]
- Webb, M. The Morphology and Formation of the Pogonophoran Tube and its Value in Systematics. J. Zool. Syst. Evol. Res. 1971, 9, 169–181. [Google Scholar] [CrossRef]
- Smirnov, R.; Zaitseva, O.; Vedenin, A. A remarkable pogonophoran from a desalted shallow near the mouth of the Yenisey River in the Kara Sea, with the description of a new species of the genus Galathealinum (Annelida: Pogonophora: Frenulata). Zoosystematica Ross. 2020, 29, 138–154. [Google Scholar] [CrossRef]
- Karaseva, N.P.; Rimskaya-Korsakova, N.N.; Ekimova, I.A.; Gantsevich, M.M.; Kokarev, V.N.; Kremnyov, S.V.; Simakov, M.I.; Udalov, A.A.; Vedenin, A.A.; Malakhov, V.V. A new genus of frenulates (Annelida: Siboglinidae) from shallow waters of the Yenisey River estuary, Kara Sea. Invertebr. Syst. 2021, 35, 857–875. [Google Scholar] [CrossRef]
- Dolgopolova, E.N. Regularities in the motion of water and sediments at the mouth of a river of estuarine-deltaic type: Case study of the Yenisei, R. Water Resour. 2015, 42, 198–207. [Google Scholar] [CrossRef]
- Macdonald, I.R.; Boland, G.S.; Baker, J.S.; Brooks, J.M.; Ii, M.C.K.; Bidigare, R.R. Gulf of Mexico hydrocarbon seep communities. Mar. Biol. 1989, 101, 235–247. [Google Scholar] [CrossRef]
- Fisher, C.R.; Urcuyo, I.A.; Simpkins, M.A.; Nix, E. Life in the Slow Lane: Growth and Longevity of Cold-seep Vestimentiferans. Mar. Ecol. 1997, 18, 83–94. [Google Scholar] [CrossRef]
- Bergquist, D.C.; Ward, T.; Cordes, E.; McNelis, T.; Howlett, S.; Kosoff, R.; Hourdez, S.; Carney, R.; Fisher, C.R. Community structure of vestimentiferan-generated habitat islands from Gulf of Mexico cold seeps. J. Exp. Mar. Biol. Ecol. 2003, 289, 197–222. [Google Scholar] [CrossRef]
- Bergquist, D.; Urcuyo, I.; Fisher, C. Establishment and persistence of seep vestimentiferan aggregations on the upper Louisiana slope of the Gulf of Mexico. Mar. Ecol. Prog. Ser. 2002, 241, 89–98. [Google Scholar] [CrossRef]
- Cordes, E.E.; Arthur, M.A.; Shea, K.; Arvidson, R.; Fisher, C.R. Modeling the Mutualistic Interactions between Tubeworms and Microbial Consortia. PLoS Biol. 2005, 3, e77. [Google Scholar] [CrossRef] [Green Version]
- Cordes, E.E.; Bergquist, D.C.; Predmore, B.L.; Jones, C.; Deines, P.; Telesnicki, G.; Fisher, C.R. Alternate unstable states: Convergent paths of succession in hydrocarbon-seep tubeworm-associated communities. J. Exp. Mar. Biol. Ecol. 2006, 339, 159–176. [Google Scholar] [CrossRef]
- Cordes, E.E.; Carney, S.L.; Hourdez, S.; Carney, R.S.; Brooks, J.M.; Fisher, C.R. Cold seeps of the deep Gulf of Mexico: Community structure and biogeographic comparisons to Atlantic equatorial belt seep communities. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 637–653. [Google Scholar] [CrossRef]
- Cordes, E.E.; Bergquist, D.C.; Fisher, C.R. Macro-Ecology of Gulf of Mexico Cold Seeps. Annu. Rev. Mar. Sci. 2009, 1, 143–168. [Google Scholar] [CrossRef] [Green Version]
- Baker, M.C.; Ramirez-Llodra, E.Z.; Tyler, P.A.; German, C.R.; Boetius, A.; Cordes, E.E.; Dubilier, N.; Fisher, C.R.; Levin, L.A.; Metaxas, A.; et al. Biogeography, Ecology, and Vulnerability of Chemosynthetic Ecosystems in the Deep Sea. In Life in the World’s Oceans: Diversity, Distribution, and Abundance; McIntyre, A., Ed.; Wiley-Blackwell Publishing: Oxford, UK, 2010; pp. 161–182. [Google Scholar] [CrossRef]
- Flügel, H.J.; Callsen-Cencic, P. New observations on the biology of Siboglinum poseidoni Flügel & Langhof (Pogonophora) from the Skagerrak. Sarsia 1992, 77, 287–290. [Google Scholar] [CrossRef]
- Gebruk, A.V.; Krylova, E.M.; Lein, A.Y.; Vinogradov, G.M.; Anderson, E.; Pimenov, N.V.; Cherkashev, G.A.; Crane, K. Methane seep community of the Håkon Mosby mud volcano (the Norwegian Sea): Composition and trophic aspects. Sarsia 2003, 88, 394–403. [Google Scholar] [CrossRef]
- Sahling, H.; Wallmann, K.J.G.; Dählmann, A.; Schmaljohann, R.; Petersen, S. The physicochemical habitat of Sclerolinum sp. at Hook Ridge hydrothermal vent, Bransfield Strait, Antarctica. Limnol. Oceanogr. 2005, 50, 598–606. [Google Scholar] [CrossRef] [Green Version]
- Niemann, H.; Loesekann, T.; de Beer, D.; Elvert, M.; Nadalig, T.; Knittel, K.; Amann, R.; Sauter, E.; Schlueter, M.; Klages, M.; et al. Novel microbial communities of the Haakon Mosby mud volcano and their role as a methane sink. Nature 2006, 443, 854–858. [Google Scholar] [CrossRef] [PubMed]
- Sauter, E.J.; Muyakshin, S.I.; Charlou, J.-L.; Schlueter, M.; Boetius, A.; Jerosch, K.; Damm, E.; Foucher, J.-P.; Klages, M. Methane discharge from a deep-sea submarine mud volcano into the upper water column by gas hydrate-coated methane bubbles. Earth Planet. Sci. Lett. 2006, 243, 354–365. [Google Scholar] [CrossRef] [Green Version]
- Dubilier, N.; Bergin, C.; Lott, C. Symbiotic diversity in marine animals: The art of harnessing chemosynthesis. Nat. Rev. Genet. 2008, 6, 725–740. [Google Scholar] [CrossRef]
- Sommer, S.; Linke, P.; Pfannkuche, O.; Schleicher, T.; Reitz, A.; Haeckel, M.; Flögel, S.; Hensen, C. Seabed methane emissions and the habitat of frenulate tubeworms on the Captain Arutyunov mud volcano (Gulf of Cadiz). Mar. Ecol. Prog. Ser. 2009, 382, 69–86. [Google Scholar] [CrossRef]
- Southward, E.C.; Andersen, A.C.; Hourdez, S. Lamellibrachia anaximandrin sp., a new vestimentiferan tubeworm (Annelida) from the Mediterranean, with notes on frenulate tubeworms from the same habitat. Zoosystema 2011, 33, 245–279. [Google Scholar] [CrossRef]
- Aquilina, A.; Connelly, U.P.; Copley, J.T.; Green, D.R.H.; Hawkes, J.A.; Hepburn, L.E.; Huvenne, V.A.I.; Marsh, L.; Mills, R.A.; Tyler, P.A. Geochemical and Visual Indicators of Hydrothermal Fluid Flow through a Sediment-Hosted Volcanic Ridge in the Central Bransfield Basin (Antarctica). PLoS ONE 2013, 8, e54686. [Google Scholar] [CrossRef] [Green Version]
- Åström, E.K.L.; Carroll, M.L.; Ambrose, W.G.; Sen, A.; Silyakova, A.; Carroll, J. Methane cold seeps as biological oases in the high-Arctic deep sea. Limnol. Oceanogr. 2017, 63, S209–S231. [Google Scholar] [CrossRef]
- Rimskaya-Korsakova, N.N.; Karaseva, N.; Kokarev, V.N.; Simakov, M.I.; Gantsevich, M.M.; Malakhov, V.V. First Discovery of Pogonophora (Annelida, Siboglinidae) in the Kara Sea Coincide with the Area of High Methane Concentration. Dokl. Biol. Sci. 2020, 490, 25–27. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Didriksen, A.; Hourdez, S.; Svenning, M.M.; Rasmussen, T.L. Frenulate siboglinids at high Arctic methane seeps and insight into high latitude frenulate distribution. Ecol. Evol. 2020, 10, 1339–1351. [Google Scholar] [CrossRef]
- Baranov, B.; Galkin, S.; Vedenin, A.; Dozorova, K.; Gebruk, A.; Flint, M. Methane seeps on the outer shelf of the Laptev Sea: Characteristic features, structural control, and benthic fauna. Geo-Mar. Lett. 2020, 40, 541–557. [Google Scholar] [CrossRef]
- Vedenin, A.A.; Kokarev, V.N.; Chikina, M.V.; Basin, A.B.; Galkin, S.V.; Gebruk, A.V. Fauna associated with shallow-water methane seeps in the Laptev Sea. PeerJ 2020, 8, e9018. [Google Scholar] [CrossRef]
- Huntington, P.H. Arctic Oil and Gas 2007; Arctic Monitoring and Assessment Programme (AMAP): Oslo, Norway, 2007. [Google Scholar]
- Ostisty, B.; Fedorovsky, Y. Main results of oil and gas prospecting in the Barents and Kara Sea inspire optimism. In Norwegian Petroleum Society Special Publications; Elsevier: Amsterdam, The Netherlands, 1993; Volume 2, pp. 243–252. [Google Scholar] [CrossRef]
- Ostisty, B.; Cheredeev, S.; Dore, A. Main factors controlling regional oil and gas potential in the west Arctic, former USSR. In Basin Modelling: Advances and Applications, NPF (Norwegian Petroleum Society) Special Publication 3; Doré, A., Augustson, J., Hermanrud, C., Stewart, D., Sylta, Ø., Eds.; Elsevier Science, Norwegian Petroleum Society (NPF) Special Publication: Amsterdam, The Netherlands, 1993; Volume 3, pp. 591–597. [Google Scholar]
- Johansen, S.; Ostisty, B.; Fedorovsky, Y.; Martirosjan, V.; Christensen, O.B.; Cheredeev, S.; Ignatenko, E.; Margulis, L. Hydrocarbon potential in the Barents Sea region: Play distribution and potential. In Norwegian Petroleum Society Special Publications; Elsevier: Amsterdam, The Netherlands, 1993; Volume 2, pp. 273–320. [Google Scholar] [CrossRef]
- Doré, A. Barents Sea Geology, Petroleum Resources and Commercial Potential. ARCTIC 1995, 48, 207–221. [Google Scholar] [CrossRef]
- Borisov, A.V.; Tanygin, I.A.; Vinnikovsky, V.S.; Borisova, B.A. The Shtokmanovsko-Luninskii structural barrier of the Barents Sea shelf is a new large oil and gas bearing region of Russia. Geol. Nefti Gasa 1995, 7, 10. (In Russian) [Google Scholar]
- Gramberg, I.S.E.N.K.; Suprunenko, O.I. Catagenetic zonation of the sedimentary cover on the Barents shelf in connection with petroleum potential. Geol. Geofiz. 2001, 42, 1808–1820. (In Russian) [Google Scholar]
- Margulis, E.A. Evolution of the Barents Sea region and its hydrocarbon systems. Neftegazov. Geologia. Teor. Pract. 2009, 4, 8. [Google Scholar]
- Drachev, S.S.; Malyshev, N.A.; Nikishin, A.M. Tectonic history and petroleum geology of the Russian Arctic Shelves: An overview. Geol. Soc. Lond. Pet. Geol. Conf. Ser. 2010, 7, 591–619. [Google Scholar] [CrossRef]
- Henriksen, E.; Ryseth, A.E.; Larssen, G.B.; Heide, T.; Rønning, K.; Sollid, K.; Stoupakova, A.V. Chapter 10 Tectonostratigraphy of the greater Barents Sea: Implications for petroleum systems. Geol. Soc. Lond. Mem. 2011, 35, 163–195. [Google Scholar] [CrossRef]
- Norina, D.A.; Stoupkova, A.V.; Kiryuhina, T.A. Usloviya osadkonakoplenia b neftegazomaterinskij potentsial triasovyh otlogenij Barentsevomorskogo basseina [Sedimentation conditions and oil and gas source potential of Triassic sediments in the Barents Sea basin]. Mosc. Univ. Bull. Ser. 4. Geol. 2014, 1, 6–16. (In Russian) [Google Scholar]
- Andreassen, K.; Hubbard, A.; Winsborrow, M.; Patton, H.; Vadakkepuliyambatta, S.; Plaza-Faverola, A.; Gudlaugsson, E.; Serov, P.; Deryabin, A.; Mattingsdal, R.; et al. Massive blow-out craters formed by hydrate-controlled methane expulsion from the Arctic seafloor. Science 2017, 356, 948–953. [Google Scholar] [CrossRef] [Green Version]
- Stoupakova, A.V.; Bolshakova, M.A.; Suslova, A.A.; Mordasova, A.V.; Osipov, K.O.; Kovalevskaya, S.O.; Kolesnikova, T.O.; Shevchenko, G.A.; Masterkov, I.A.; Tsygankova, A.A.; et al. Generation potential, distribution area and maturity of the Barents-Kara Sea source rocks. Georesursy 2021, 23, 6–25. [Google Scholar] [CrossRef]
- Suslova, A.A.; Stoupakova, A.V.; Bolshakova, M.A.; Mordasova, A.V.; Sautkin, R.S.; Krasnova, E.A.; Osipov, K.O.; Kolesnikova, T.O.; Kovalevskaya, S.O.; Ghilmillina, A.A.; et al. Characteristics of the oil and gas mother strata of the Barents-Kara region the basis of basin analysis and resource forecast. Neftegaz. RU 2021, 2, 64–71. (In Russian) [Google Scholar]
- Stoupakova, A.V. Struktura i neftegazonosnost’ Barentsevo-Karskogo shelfa i prilegayushih territorij. [Structure and oil and gas content of the Barents-Kara shelf and adjacent territories]. Geol. Oil Gas 2011, 6, 99–115. (In Russian) [Google Scholar]
- Ivanov, A. Pogonofory (Pogonophora); Fauna SSSR. 75; Pavovsky, E.N., Ed.; ANSSSR: Moscow, Leningrad, Russia, 1960; p. 271. [Google Scholar]
- Southward, E.C. A New Species of Galathealinum (Pogonophora) from the Canadian Arctic. Can. J. Zool. 1962, 40, 385–389. [Google Scholar] [CrossRef]
- Smirnov, R.V. New Species of the Genus Polarsternium (Pogonophora) from the Scotia Sea and Adjacent Waters of the Antarctic. Russ. J. Mar. Biol. 2005, 31, 146–154. [Google Scholar] [CrossRef]
- Smirnov, R.V. A revision of the Oligobrachiidae (Annelida: Pogonophora), with notes on the morphology and distribution of Oligobrachia haakonmosbiensis Smirnov. Mar. Biol. Res. 2014, 10, 972–982. [Google Scholar] [CrossRef]
- Karaseva, N.P.; Rimskaya-Korsakova, N.N.; Ekimova, I.A.; Kokarev, V.N.; Simakov, M.I.; Gantsevich, M.M.; Malakhov, V.V. The first discovery of pogonophores (Annelida, Siboglinidae) in the East Siberian Sea coincides with the areas of methane seeps. Doklady RAS 2021, 501, 23–27. [Google Scholar]
- Lee, Y.M.; Noh, H.-J.; Lee, D.-H.; Kim, J.-H.; Jin, Y.K.; Paull, C. Bacterial endosymbiont of Oligobrachia sp. (Frenulata) from an active mud volcano in the Canadian Beaufort Sea. Polar Biol. 2019, 42, 2305–2312. [Google Scholar] [CrossRef]
- Smirnov, R.V. A new genus and two new species of pogonophora from the Arctic Ocean. Russ. J. Mar. Biol. 1999, 25, 312–319. [Google Scholar]
- Flint, M.V.; Poyarkov, S.G.; Rimsky-Korsakov, N.A. Ecosystems of the Russian Arctic-2015 (63rd Cruise of the research vessel Akademik Mstislav Keldysh). Oceanology 2016, 56, 459–461. [Google Scholar] [CrossRef]
- Shakhova, N.E.; Semiletov, I.P.; Belcheva, N.N. Great Siberian rivers as sources of methane on the Arctic shelf. Dokl. Akad. Nauk. 2007, 144, 683–685. [Google Scholar] [CrossRef]
- Guo, L.; Semiletov, I.; Gustafsson, O.; Ingri, J.; Andersson, P.; Dudarev, O.; White, D. Characterization of Siberian Arctic coastal sediments: Implications for terrestrial organic carbon export. Glob. Biogeochem. Cycles 2004, 18. [Google Scholar] [CrossRef]
- Dando, P.; Bussmann, I.; Nlven, S.; O’Hara, S.; Schmaljohann, R.; Taylor, L. A methane seep area in the Skagerrak, the habitat of the pogonophore Siboglinum poseidoni and the bivalve mollusc Thyasira sarsi. Mar. Ecol. Prog. Ser. 1994, 107, 157–167. [Google Scholar] [CrossRef]
- Ivanov, A.V. Pogonophorans of the noth-western part of the Pacific ocean. Tr. Probl. Temat. Soveshaniy ZIN 1956, 6, 20–21. [Google Scholar]
- Ivanov, A.V. Neue Pogonophora aus dem nordwestlichen Teil des Stillen Ozeans. Zoologische Jahrbücher. Abteilung für Systematik. Okol. Geogr. Tiere 1957, 85, 431–500. [Google Scholar]
- Ivanov, A.V. Pogonophorans and their geographical distribution. Dostijeniya Okeanol. 1959, 1, 258–284. [Google Scholar]
- Karaseva, N.; Gantsevich, M.M.; Obzhirov, A.I.; Shakirov, R.B.; Starovoytov, A.V.; Smirnov, R.V.; Malakhov, V.V. Siboglinids (Annelida, Siboglinidae) as Possible Hydrocarbon Indicators as Exemplified by the Sea of Okhotsk. Dokl. Biol. Sci. 2019, 486, 72–75. [Google Scholar] [CrossRef]
- Karaseva, N.; Gantsevich, M.; Obzhirov, A.; Shakirov, R.; Starovoitov, A.; Smirnov, R.; Malakhov, V. Correlation of the siboglinid (Annelida: Siboglinidae) distribution to higher concentrations of hydrocarbons in the Sea of Okhotsk. Mar. Pollut. Bull. 2020, 158, 111448. [Google Scholar] [CrossRef]
- Caullery, M. Sur les Siboglinidae, type nouveau d’invertebres recueillis par l’expedition du Siboga. Bull. Société Zool. Fr. 1914, 39, 1–204. [Google Scholar]
- Ivanov, A.V.; Southward, E.C. New Pogonophora from the Atlantic and Pacific Oceans. J. Zool. 1971, 164, 271–304. [Google Scholar] [CrossRef]
- Kirkegaard, J.B. Pogonophora. In Galathea Report; Danish Science Press: Copenhagen, Denmark, 1956; Volume 2, pp. 79–83. [Google Scholar]
- Southward, E.C. New Records of Pogonophora from Central American Seas. Bull. Mar. Sci. 1966, 16, 643–647. [Google Scholar]
- Southward, E.C. New Pogonophora from the northeast Pacific Ocean. Can. J. Zool. 1969, 47, 395–403. [Google Scholar] [CrossRef]
- Southward, E.C. Pogonophora of the northwest Atlantic: Nova Scotia to Florida. Smithson. Contrib. Zool. 1971, 1–29. [Google Scholar] [CrossRef]
- Southward, E.C. Description of a New Species of Oligobrachia (Pogonophora) from the North Atlantic, With a Survey of the Oligobrachiidae. J. Mar. Biol. Assoc. U. K. 1978, 58, 357–365. [Google Scholar] [CrossRef]
- Southward, E.C. Two New Species of Pogonophora from Hawaii. Pac. Sci. 1980, 34, 371–378. [Google Scholar]
- Webb, M. Siboglinum fiordicum sp. nov. (pogonophora) from the raunefjord, Western Norway. Sarsia 1963, 13, 33–44. [Google Scholar] [CrossRef]
- Adegoke, O.S. Pogonophora from the northeastern Pacific: First records from the Gulf of Tehuantepec, Mexico. Pac. Sci. 1967, 21, 188–192. [Google Scholar]
- Southward, E.C.; Brattegard, T. Pogonophora of the Northwest Atlantic: North Carolina Region. Bull. Mar. Sci. 1968, 18, 836–875. [Google Scholar]
- Cavanaugh, C.M. Symbiosis of chemoautotrophic bacteria and marine invertebrates. Biol. Bull. Mar. Biol. Lab. 1980, 159, 457. [Google Scholar]
- Cavanaugh, C.M.; Gardiner, S.L.; Jones, M.L.; Jannasch, H.W.; Waterbury, J.B. Prokaryotic Cells in the Hydrothermal Vent Tube Worm Riftia pachyptila Jones: Possible Chemoautotrophic Symbionts. Science 1981, 213, 340–342. [Google Scholar] [CrossRef]
- Cavanaugh, C.M. Symbiotic chemoautotrophic bacteria in marine invertebrates from sulphide-rich habitats. Nature 1983, 302, 58–61. [Google Scholar] [CrossRef]
- Uschakov, P. Eine neue form aus der Familie Sabellidae (Polychaeta). Zool. Anz 1933, 104, 205–208. [Google Scholar]
- Zenkevich, L.; Filatova, Z. Obshchaya kratkaya kharakteristika kachestvennogo sostava i kolichestvennogo raspredeleniya donnoi fauny dal’nevostochnykh morei i severo-zapadnoi chasti Tikhogo okeana (General Brief Description of the Qualitative Composition and the Quantitative Distribution of Benthic Fauna of the Far Eastern Seas and the Northwestern Pacific). Tr. IOAN 1958, 27, 154–160. [Google Scholar]
- Ivanov, A.V. Pogonofory severo-zapadnoi chasti Tihogookeana [Pogonophorans of the north-western part of the Pacific Ocean]. Trudy problem i tematicheskih Soveshaniy ZIN 1956, 6, 20–21. (In Russian) [Google Scholar]
- Ivanov, A.V. Pogonofory i ih geograficheskoe rasprostranenie. [Pogonophorans and Their Geographical Distribution]. Dostijeniya Okeanologii. AN SSSR 1959, 1, 1–10. (In Russian) [Google Scholar]



{kind=link}
{kind=link}
| R/V | Stations | NN | EE | Date | Depth, m | Temp., °C | Salinity | Sediment | Material |
|---|---|---|---|---|---|---|---|---|---|
| R/V Vilnius | 37 | 69.05 | 39.71 | 8 August 2007 | 226.7 | Silt | A, T | ||
| R/V Dalnie Zelentcy | 36 | 69.21 | 35.96 | 26 June 1993 | 195 | Sand | A | ||
| R/V Dalnie Zelentcy | 34 | 69.43 | 37.21 | 24 June 1993 | 185 | A | |||
| R/V Professor Boyko | 2011-5 | 69.48 | 35.21 | 8 July 2011 | 175.5 | Sand | T | ||
| R/V Professor Boyko | 38 | 69.51 | 36.00 | 27 June 1993 | 200 | A | |||
| R/V Romuald Muklevich | 23-2 | 69.53 | 32.88 | 12 August 2003 | 254 | 3.08 | 34.489 | Silt | A |
| R/V Professor Boyko | C-1 | 69.55 | 32.59 | 5 July 2011 | 204 | Sand | A, J, L | ||
| R/V Romuald Muklevich | 22-1 | 69.55 | 32.87 | 11 August 2003 | 224.8 | 2.9 | 34.451 | Silt | A, E, L, T |
| R/V Romuald Muklevich | 19-1 | 69.55 | 32.59 | 11 August2003 | 204 | 4.76 | A, T | ||
| R/V Romuald Muklevich | 20-4 | 69.56 | 32.65 | 11 August 2003 | 241.8 | 2.76 | 34.418 | Silt | A, L, T |
| R/V Romuald Muklevich | 27-2 | 69.58 | 33.10 | 13 August 2003 | 225.6 | 3.12 | 34.501 | Silt | A |
| R/V Romuald Muklevich | 28-1 | 69.59 | 33.15 | 13 August 2003 | 232.6 | 2.94 | 34.451 | Silt | A |
| R/V Professor Boyko | 15-1 | 69.59 | 36.34 | 23 November 2008 | 190.3 | 3.49 | 34.599 | Silt | A, L |
| R/V Dalnie Zelentcy | 32 | 69.67 | 40.00 | 24 June 1993 | 170 | Sand | A | ||
| R/V Fridtjof Nansen | 43-2 | 72.30 | 40.00 | 28 September 2006 | 341 | 0.15 | 34.99 | Silt | A, E, T |
| R/V Smolensk | 23-5 | 72.30 | 51.01 | 11 September2006 | 75.2 | 0.11 | 34.73 | Stones | A |
| R/V Smolensk | 19-3 | 72.30 | 48.60 | 7 September2006 | 175.2 | 2.98 | 34.49 | Silt | A, T |
| R/V Fridtjof Nansen | 44-2 | 72.99 | 38.01 | 28 June 2006 | 237 | 0.63 | 35.03 | Sand | A, E, L, T |
| R/V Smolensk | 21-1 | 73.00 | 47.02 | 10 September2006 | 312.6 | −0.12 | 34.96 | Clay | A, T |
| R/V Smolensk | 22-3 | 73.00 | 49.02 | 10 September2006 | 243.7 | 1.93 | 34.99 | Clay | A, T |
| R/V Smolensk | 33 | 73.30 | 35.59 | 25 Septembe 2006 | 261 | Silt | T | ||
| R/V Smolensk | 25-3 | 73.30 | 45.10 | 12 September2006 | 321.5 | −0.31 | 34.96 | Clay | A, E, T |
| R/V Smolensk | 34-2 | 73.30 | 38.03 | 25 September 2006 | 263.5 | Clay | A, T | ||
| R/V Professor Levanidov | D2 | 73.58 | 41.33 | 7 August 2020 | 275 | Clay | A, T | ||
| R/V Vilnius | 51 | 73.97 | 41.54 | 24 October2018 | 274 | 0.15 | 34.964 | Silt | A |
| R/V Smolensk | 32-3 | 74.00 | 35.60 | 24 September2006 | 228.9 | Silt | A, T | ||
| R/V Vilnius | Lud15 | 74.36 | 45.99 | 10 July 2019 | 287 | A, L, T | |||
| R/V Professor Levanidov | Lud15 | 74.22 | 43.38 | 30 July 2020 | 289 | Clay | A, T | ||
| R/V Vilnius | 6 | 74.36 | 46.01 | 17 October 2018 | 286 | 1.62 | 34.964 | Silt | A, L, T |
| R/V Professor Levanidov | Lud6 | 74.44 | 45.59 | 1 August 2020 | 292 | Clay | A, E, T | ||
| R/V Vilnius | Lud10 | 74.48 | 45.64 | 9 July 2019 | 306 | A | |||
| R/V Professor Levanidov | Lud7 | 74.49 | 46.38 | 2 August 2020 | 221 | Clay | A, E, L, T | ||
| R/V Fridtjof Nansen | 12-1 | 74.50 | 33.48 | 2 September 2006 | 257.6 | 0.08 | 35.03 | Silt | A, J, T |
| R/V Professor Levanidov | Lud2 | 74.52 | 45.35 | 2 August 2020 | 291 | Clay | A | ||
| R/V Professor Levanidov | Lud3 | 74.58 | 46.16 | 2 August 2020 | 270 | Clay | A, E, T | ||
| R/V Vilnius | Lud6 | 74.72 | 45.95 | 9 July 2019 | 297 | A, T | |||
| R/V Vilnius | 17 | 74.73 | 45.98 | 18 October 2018 | 296 | 1.55 | 34.956 | Silt | A, T |
| R/V Vilnius | Lud7 | 74.82 | 46.65 | 8 July 2019 | 222 | T | |||
| R/V Vilnius | 18 | 74.83 | 46.66 | 18 October 2018 | 221 | 1.45 | 34.883 | Silt | A, L, T |
| R/V Fridtjof Nansen | 6-4 | 74.83 | 17.44 | 29 August 2006 | 296.4 | 4.43 | 35.065 | Silt | T |
| R/V Vilnius | 22 | 74.97 | 45.26 | 18 October 2018 | 269 | 1.45 | 34.908 | Silt | A, L, T |
| R/V Vilnius | 3 | 74.97 | 46.25 | 8 July 2019 | 279 | A, L, T | |||
| R/V Fridtjof Nansen | 16-5 | 75.00 | 29.90 | 4 September 2006 | 375.3 | 1.35 | 35.06 | Sand | A |
| R/V Fridtjof Nansen | 60-5 | 75.01 | 27.48 | 5 October 2005 | 311.4 | 1.58 | 35.05 | Silt | T |
| R/V Fridtjof Nansen | 15-2 | 75.50 | 30.48 | 4 September 2006 | 367.1 | 1.27 | 35.06 | Silt | T |
| R/V Fridtjof Nansen | 14-1 | 75.50 | 33.49 | 3 September2006 | 223.1 | 1.30 | 35.07 | Sand | A, E |
| R/V Smolensk | 27 | 75.60 | 38.01 | 23 September2006 | 248.1 | Clay | T | ||
| R/V Fridtjof Nansen | 18-4 | 75.86 | 29.93 | 6 September2006 | 303.2 | 1.88 | 35.07 | Sand | A, T |
| R/V Fridtjof Nansen | 5-1 | 76.06 | 16.25 | 28 August 2006 | 364.6 | 2.61 | 35.01 | Silt | A |
| R/V Fridtjof Nansen | 21-2 | 76.61 | 30.00 | 8 September 2006 | 282.6 | 2.05 | 35.07 | Sand | A, T |
| R/V Fridtjof Nansen | 24-4 | 77.51 | 33.54 | 8 September2006 | 150.9 | 1.81 | 34.94 | Sand | A |
| R/V Fridtjof Nansen | 35-3 | 78.03 | 43.00 | 15 September2006 | 267.6 | 0.05 | 34.99 | Sand | A, L, T |
| R/V Academik Treshnikov | 39B | 79.63 | 44.72 | 6 May 2019 | 126 | A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karaseva, N.; Kanafina, M.; Gantsevich, M.; Rimskaya-Korsakova, N.; Zakharov, D.; Golikov, A.; Smirnov, R.; Malakhov, V. Distribution of Nereilinum murmanicum (Annelida, Siboglinidae) in the Barents Sea in the Context of Its Oil and Gas Potential. J. Mar. Sci. Eng. 2021, 9, 1339. https://doi.org/10.3390/jmse9121339
Karaseva N, Kanafina M, Gantsevich M, Rimskaya-Korsakova N, Zakharov D, Golikov A, Smirnov R, Malakhov V. Distribution of Nereilinum murmanicum (Annelida, Siboglinidae) in the Barents Sea in the Context of Its Oil and Gas Potential. Journal of Marine Science and Engineering. 2021; 9(12):1339. https://doi.org/10.3390/jmse9121339
Chicago/Turabian StyleKaraseva, Nadezda, Madina Kanafina, Mikhail Gantsevich, Nadezhda Rimskaya-Korsakova, Denis Zakharov, Alexey Golikov, Roman Smirnov, and Vladimir Malakhov. 2021. "Distribution of Nereilinum murmanicum (Annelida, Siboglinidae) in the Barents Sea in the Context of Its Oil and Gas Potential" Journal of Marine Science and Engineering 9, no. 12: 1339. https://doi.org/10.3390/jmse9121339
APA StyleKaraseva, N., Kanafina, M., Gantsevich, M., Rimskaya-Korsakova, N., Zakharov, D., Golikov, A., Smirnov, R., & Malakhov, V. (2021). Distribution of Nereilinum murmanicum (Annelida, Siboglinidae) in the Barents Sea in the Context of Its Oil and Gas Potential. Journal of Marine Science and Engineering, 9(12), 1339. https://doi.org/10.3390/jmse9121339