
Mackerel (Scomber australasicus) Reproduction in Northeastern Taiwan


Abstract
:1. Introduction
2. Materials and Methods
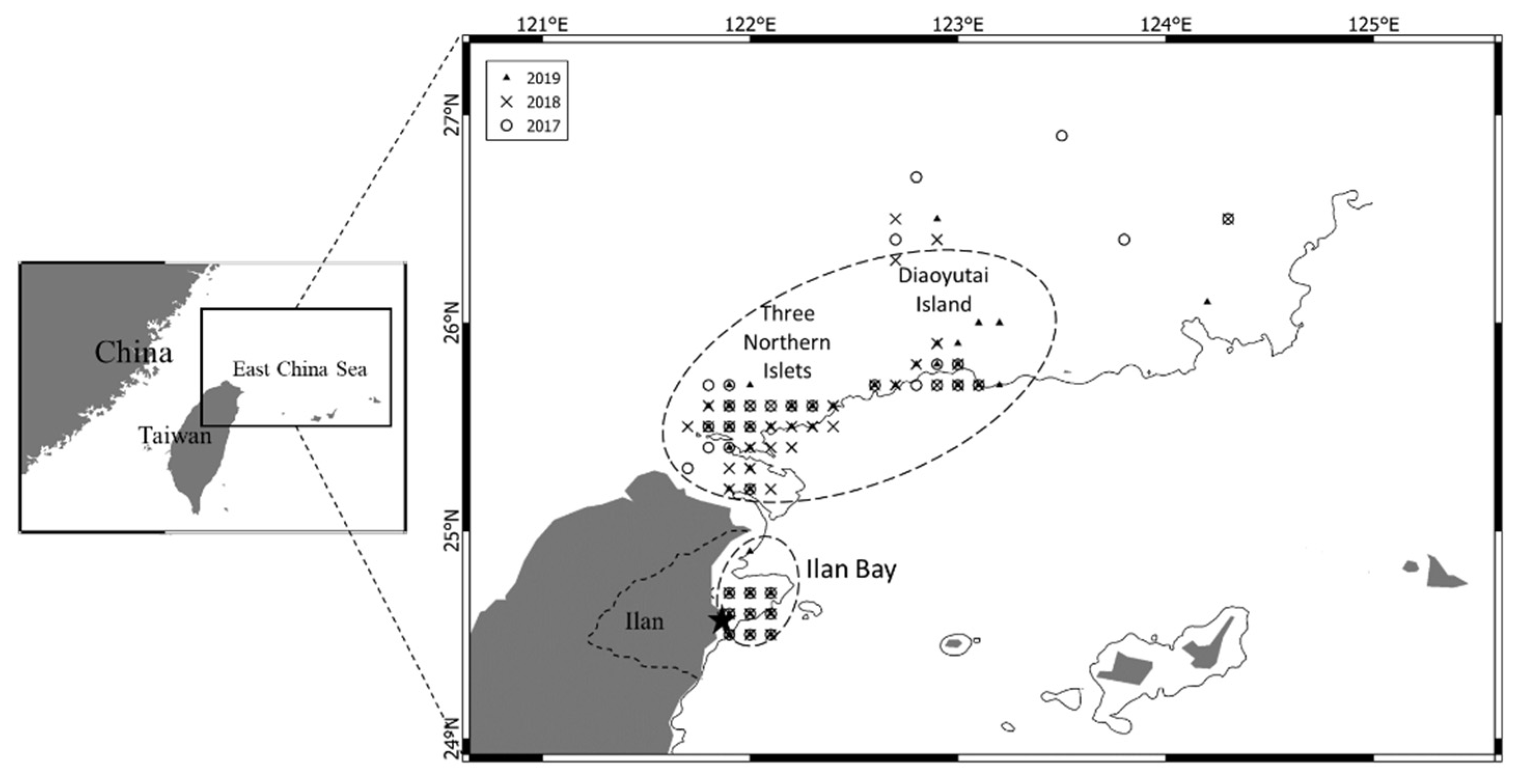
2.1. Sample Collection and Basic Measurements
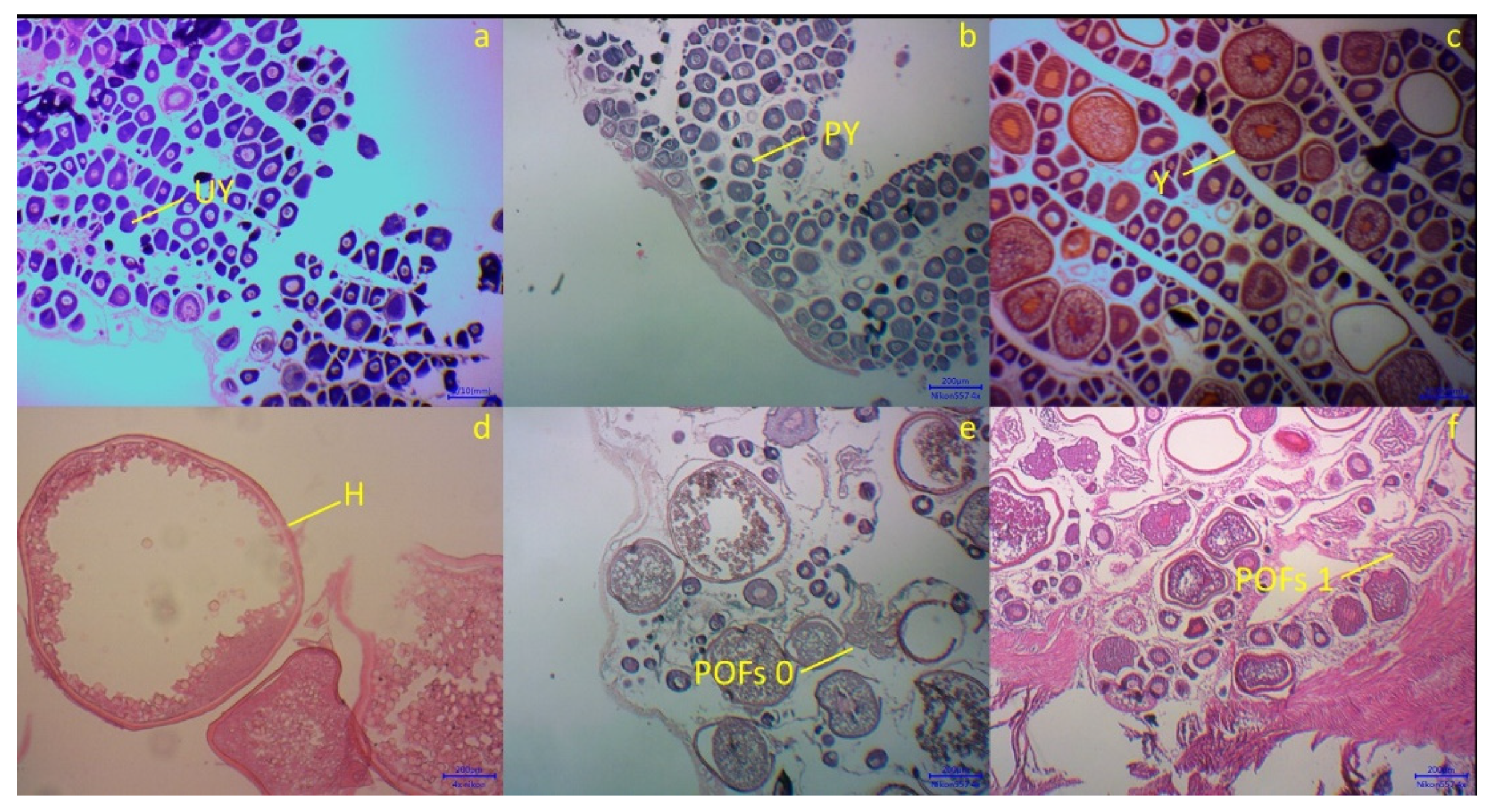
2.2. Histological Section and Ovary Classification
2.3. Fecundity and BF Estimation
2.4. Spawning Frequency Estimation
2.5. Size at 50% Maturity
3. Results
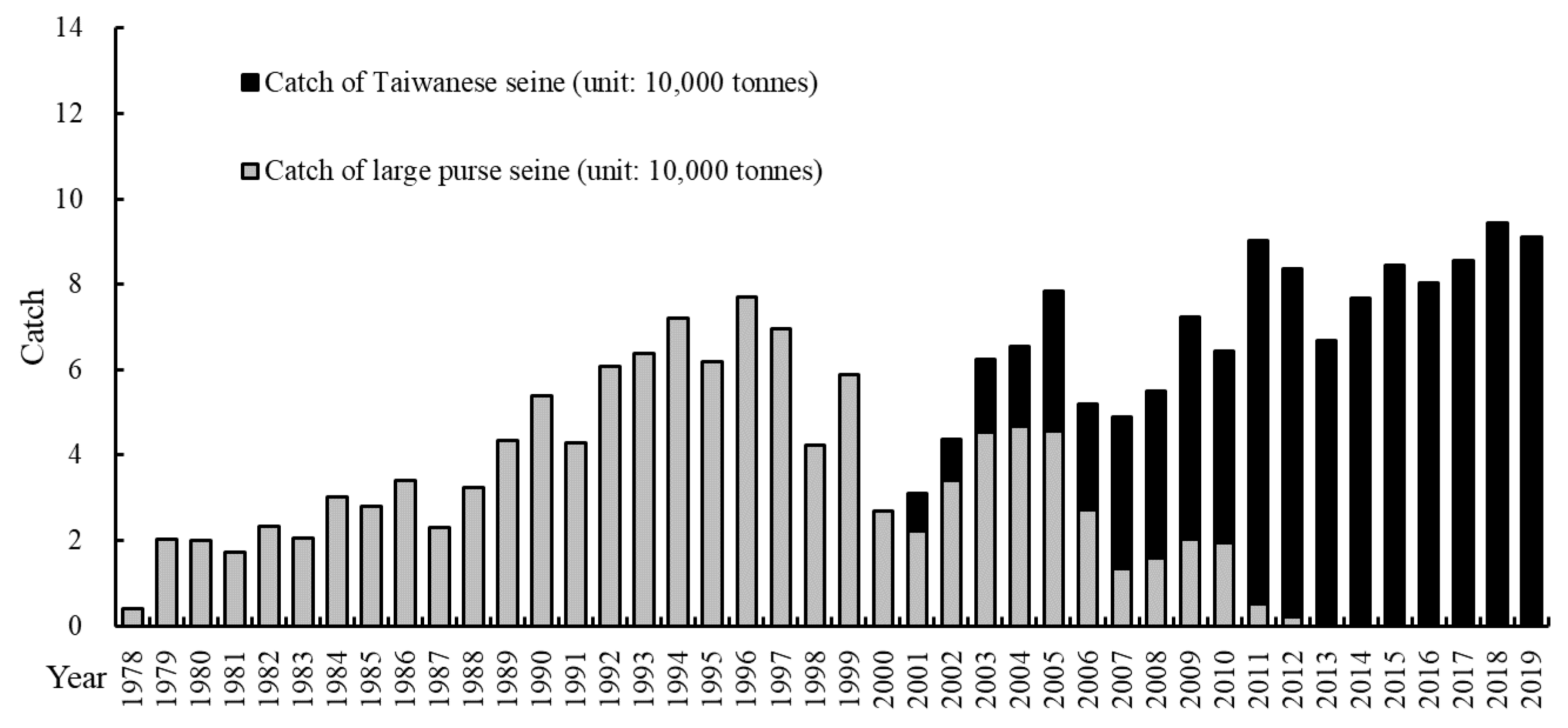
3.1. Sample Collection
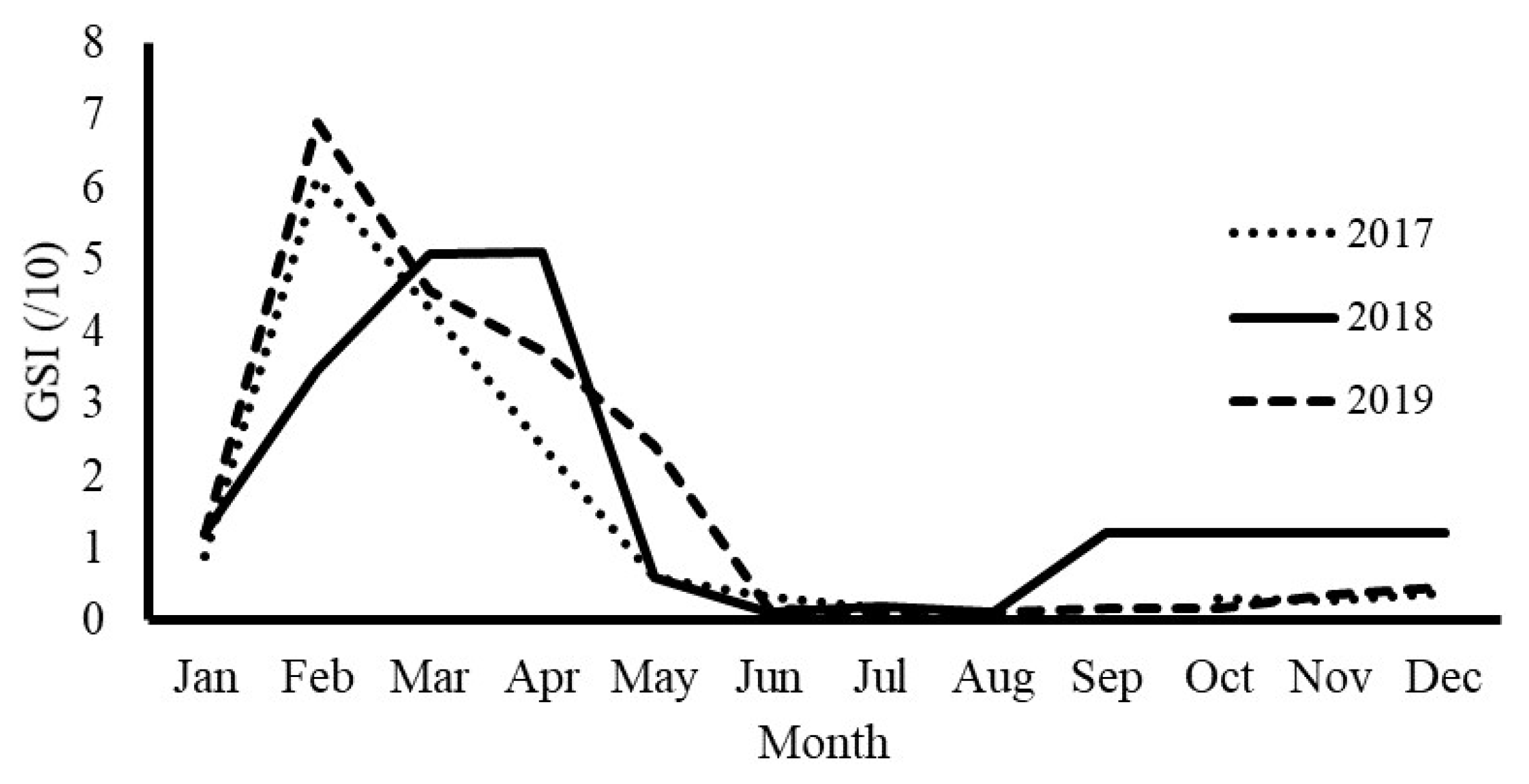
3.2. Spawning Season by GSI Observation
3.3. Observation of Oocyte Development
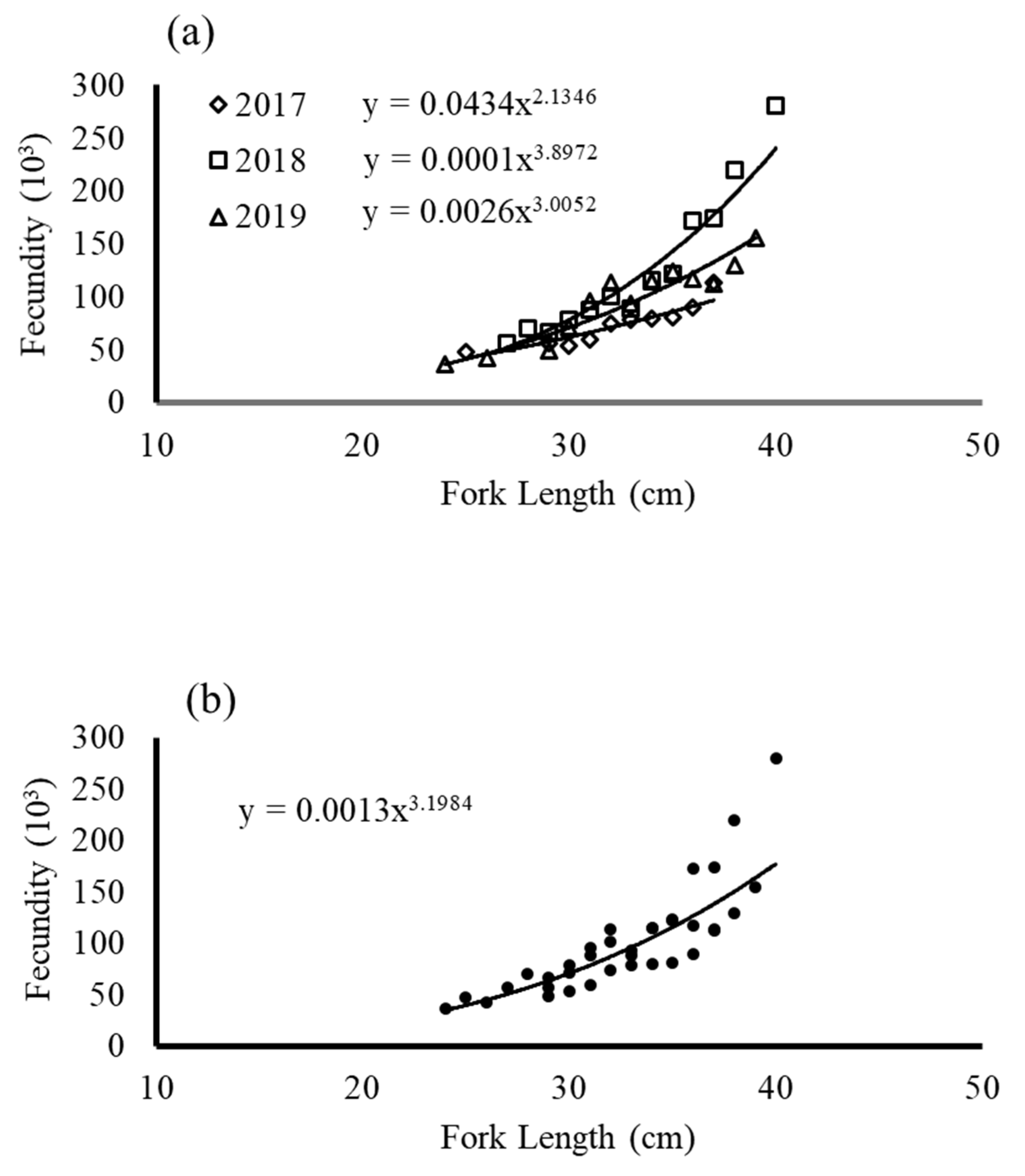
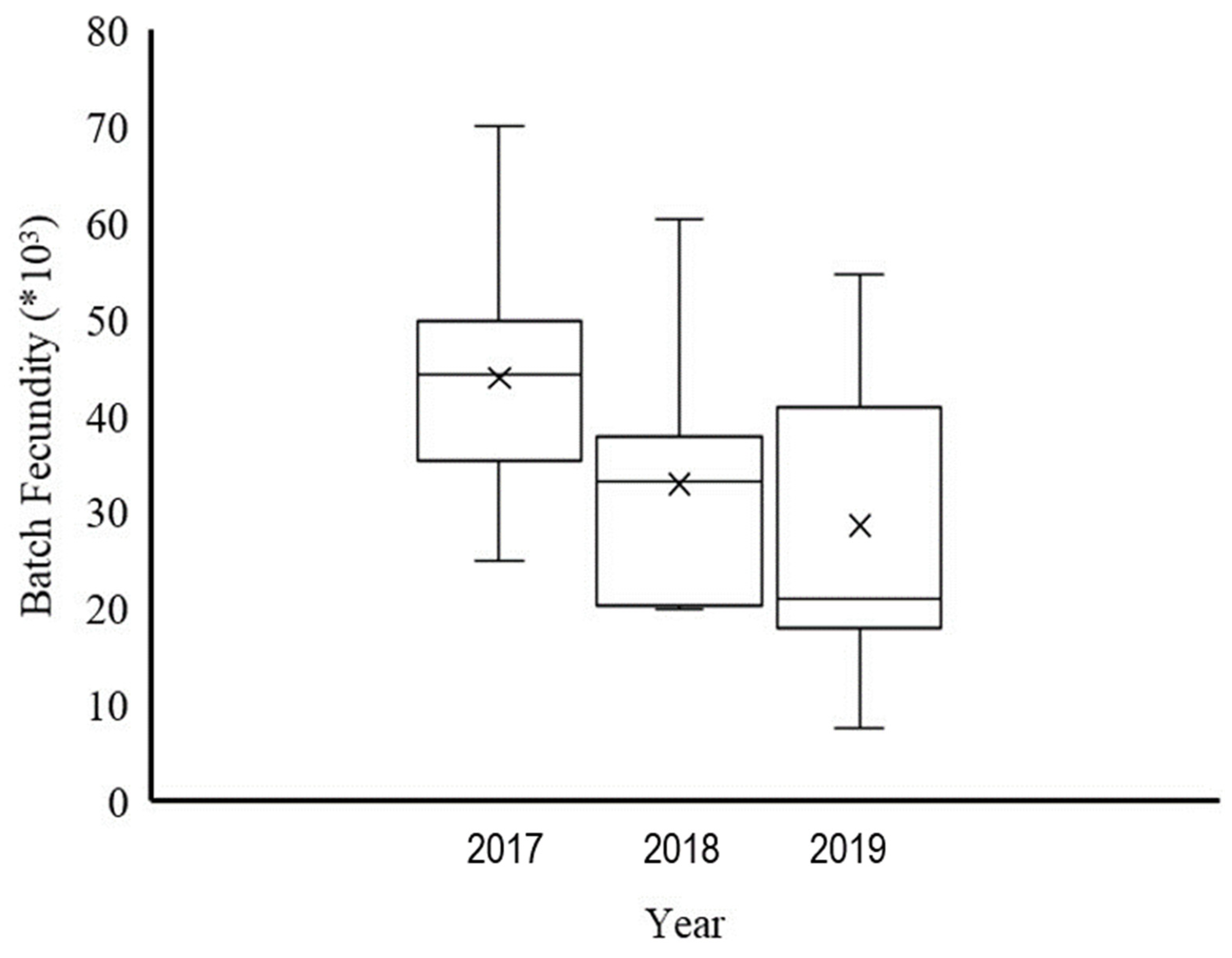
3.4. Total Fecundity and BF
3.5. Spawning Fraction and Spawning Frequency
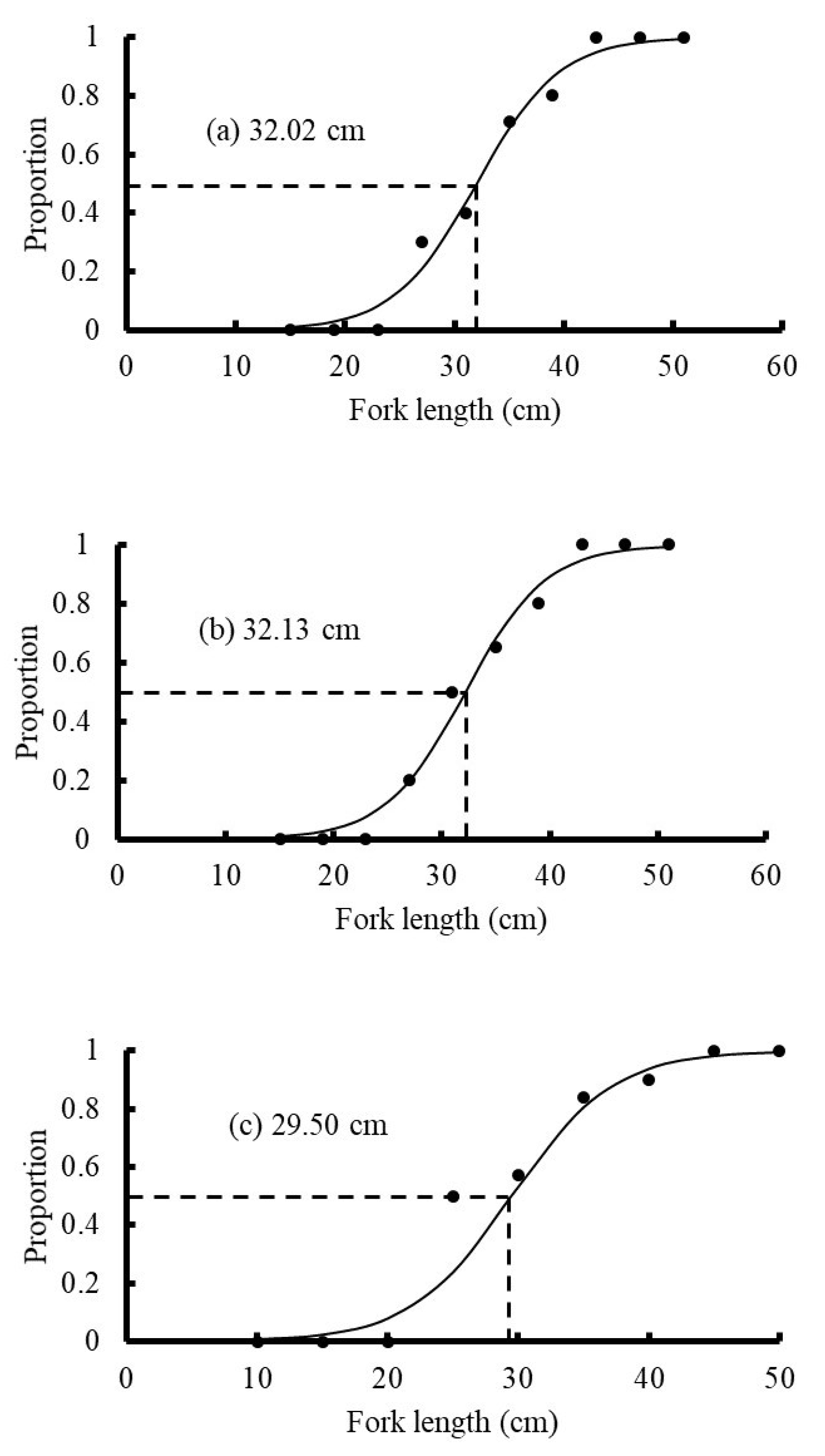
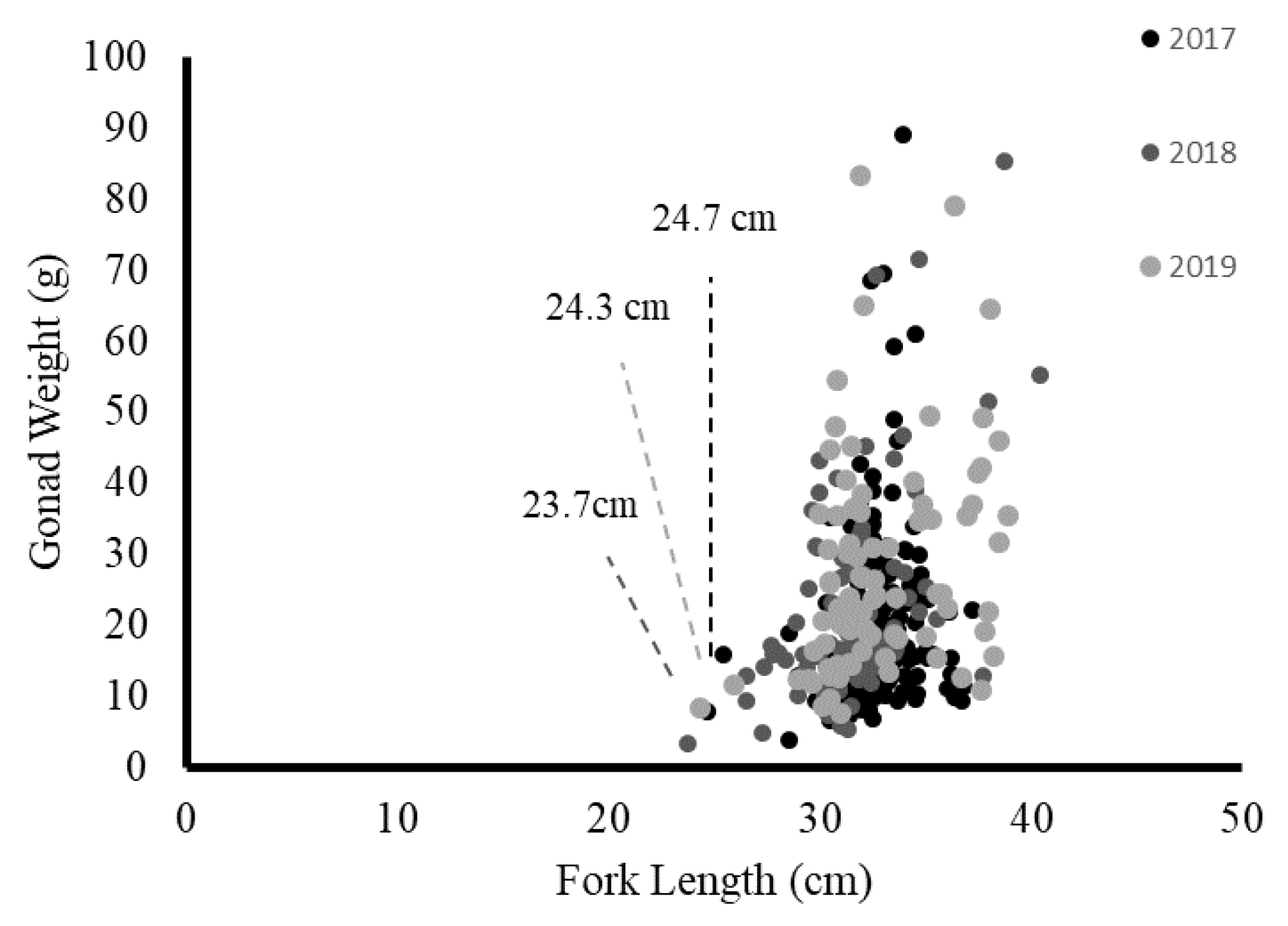
3.6. Size at 50% Maturity
4. Discussion
4.1. Spawning Mode
4.2. Fecundity and BF
4.3. Spawning Frequency
4.4. Sample Size
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Japan Statistic Yearbook. [Quantity of Catches of Marine Fisheries by Species]. 2018. Available online: https://www.stat.go.jp/english/data/nenkan/69nenkan/1431-08.html (accessed on 9 September 2021).
- Tzeng, T.D. Morphological variation between populations of spotted mackerel (Scomber australasicus) off Taiwan. Fish. Res. 2004, 68, 45–55. [Google Scholar] [CrossRef]
- Hsiao, Y.Y.; Lu, H.J. Changes in the Sizes-at-Maturity of Spotted Mackerel (Scomber australicus) under Fishing and Environmental Variations. Master’s Thesis, Department of Environmental Biology and Fisheries Science College of Ocean and Resource National Taiwan Ocean University, Keelung, Taiwan, 2011. [Google Scholar]
- Bernal, M.; Borchers, D.L.; Valdes, L.; de Lanzós, A.L.; Buckland, S.T. A new ageing method for eggs of fish species with daily spawning synchronicity. Can. J. Fish. Aquat. Sci. 2001, 58, 2330–2340. [Google Scholar] [CrossRef]
- Parker, K. A direct method for estimating northern anchovy, Engraulis mordax, spawning biomass. Fish Bull. 1980, 78, 541–544. [Google Scholar]
- Rogers, P.J.; Ward, T.M.; McLeay, L.J.; Lowry, M.; Saunders, R.J.; Williams, D. Reproductive biology of blue mackerel, Scomber australasicus, off southern and eastern Australia: Suitability of the Daily Egg Production Method for stock assessment. Mar. Freshw. Res. 2009, 60, 187–202. [Google Scholar] [CrossRef]
- Murua, H.; Kraus, G.; Saborido-Rey, F.; Witthames, P.R.; Thorsen, A.; Junquera, S. Procedures to estimate fecundity of marine fish species from field samples in relation to reproductive strategy. J. Northwest Atl. Fish. Sci. 2003, 33, 33–54. [Google Scholar] [CrossRef]
- Hunter, J.R.; Macewicz, B.J. Sexual maturity, batch fecundity, spawning frequency, and temporal pattern of spawning for the northern anchovy, Engraulis mordax, during the 1979 spawning season. CalCOFI Rep. 1980, 21, 139–149. [Google Scholar]
- Hunter, J.R.; Goldberg, S.R. Spawning incidence and batch fecundity in Northern Anchovy, Engraulis mordax. Collect. Repr. 1980, 77, 7964. [Google Scholar]
- Ward, T.M.; Rogers, P.J.; McLeay, L.J.; McGarvey, R. Evaluating the use of the daily egg production method for stock assessment of blue mackerel, Scomber australasicus. Mar. Freshw. Res. 2009, 60, 112–128. [Google Scholar] [CrossRef]
- Tzeng, T.D. Population structure and historical demography of the spotted mackerel (Scomber australasicus) off Taiwan inferred from mitochondrial control region sequencing. Zool. Stud. 2007, 46, 656. [Google Scholar]
- Bithy, K.; Miah, M.I.; Haque, M.S.; Hasan, K.R.; Islam, M.F. Estimation of the fecundity of Jat Puti, Puntius sophore (Hamilton). J. Environ. Sci. Nat. Resour. 2012, 5, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, P.; Costa, A.M.; Murta, A.G. Estimates of batch fecundity and spawning fraction for the southern stock of horse mackerel (Trachurus trachurus) in ICES Division IXa. ICES J. Mar. Sci. 2009, 66, 617–622. [Google Scholar] [CrossRef]
- Hunter, J.R.; Lo, N.C.; Leong, R.J. Batch fecundity in multiple spawning fishes. NOAA Tech. Rep. NMFS 1985, 36, 67–77. [Google Scholar]
- Shiraishi, T.; Ohta, K.; Yamaguchi, A.; Yoda, M.; Chuda, H.; Matsuyama, M. Reproductive parameters of the chub mackerel Scomber japonicus estimated from human chorionic gonadotropin-induced final oocyte maturation and ovulation in captivity. Fish. Sci. 2005, 71, 531–542. [Google Scholar] [CrossRef]
- Fitzhugh, G.A.; Fioramonti, C.A.; Walling, W.I.; Gamby, M.I.; Lyon, H.O.; Devries, D.O.; Ortiz, M.A. Batch fecundity and an attempt to estimate spawning frequency of king mackerel (Scomberomorus cavalla) in US waters. 61st Gulf and Caribbean Fisheries Institute. 2009, pp. 30–37. Available online: https://nsgl.gso.uri.edu/flsgp/flsgpw08001/data/papers/005.pdf (accessed on 9 September 2021).
- de Santana, H.S.; Cantarute Rodrigues, A.; Dei Tos, C. Patterns of reproduction and growth of the catfish Iheringichthys labrosus (Lütken, 1874) after a reservoir formation. J. Appl. Ichthyol. 2016, 32, 456–463. [Google Scholar] [CrossRef]
- Yukami, R.; Ohshimo, S.; Yoda, M.; Hiyama, Y. Estimation of the spawning grounds of chub mackerel Scomber japonicus and spotted mackerel Scomber australasicus in the East China Sea based on catch statistics and biometric data. Fish. Sci. 2009, 75, 167–174. [Google Scholar] [CrossRef]
- Hiyama, Y.; Yoda, M.; Ohshimo, S. Stock size fluctuations in chub mackerel (Scomber japonicus) in the East China Sea and the Japan/East Sea. Fish. Oceanogr. 2002, 11, 347–353. [Google Scholar] [CrossRef]
- Lee, D.; Son, S.; Kim, W.; Park, J.M.; Joo, H.; Lee, S.H. Spatio-temporal variability of the habitat suitability index for Chub Mackerel (Scomber japonicus) in the East/Japan sea and the South sea of South Korea. Remote Sens. 2018, 10, 938. [Google Scholar] [CrossRef] [Green Version]
- Yoda, M.; Shiraishi, T.; Yukami, R.; Ohshimo, S. Age and maturation of jack mackerel Trachurus japonicus in the East China Sea. Fish. Sci. 2014, 80, 61–68. [Google Scholar] [CrossRef]
- Morse, W.W. Spawning and fecundity of Atlantic mackerel, Scomber scombrus, in the Middle Atlantic Bight. Fish. Bull. 1980, 78, 103–108. [Google Scholar]
- Kjesbu, O.S.; Klungsøyr, J.; Kryvi, H.; Witthames, P.R.; Walker, M.G. Fecundity, atresia, and egg size of captive Atlantic cod (Gadus morhua) in relation to proximate body composition. Can. J. Fish. Aquat. Sci. 1991, 48, 2333–2343. [Google Scholar] [CrossRef]
- Yamada, T.; Aoki, I.; Mitani, I. Spawning time, spawning frequency and fecundity of Japanese chub mackerel, Scomber japonicus in the waters around the Izu Islands, Japan. Fish. Res. 1998, 38, 83–89. [Google Scholar] [CrossRef]
- Watanabe, C.; Hanai, T.; Meguro, K.; Ogino, R.; Kimura, R. Spawning biomass estimates of chub mackerel Scomber japonicus of Pacific subpopulation off central Japan by a daily egg production method. Nippon. Suisan Gakkaishi 1999, 65, 695–702. [Google Scholar] [CrossRef]
- Shiraishi, T.; Ketkar, S.D.; Katoh, Y.; Nyuji, M.; Yamaguchi, A.; Matsuyama, M. Spawning frequency of the Tsushima Current subpopulation of chub mackerel Scomber japonicus off Kyushu, Japan. Fish. Sci. 2009, 75, 649–655. [Google Scholar] [CrossRef]
- Dickerson, T.L.; Macewicz, B.J.; Hunter, J.R. Spawning frequency and batch fecundity of chub mackerel, Scomber japonicus, during 1985. CalCOFI Rep. 1992, 33, 130–140. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Classification | Characteristics |
|---|---|---|
| I | Immature/Unyolked | Thin ovaries with very small oocytes (diameter 0.1–0.2 mm); invisible nucleus. |
| II | Maturing/Partially Yolked | Oocytes more rounded than in stage I (diameter 0.3–0.4 mm); ovaries translucent pink color; microscopically oocytes have big nucleus. |
| III | Mature/Yolked | Oocytes larger than in stage II (diameter 0.5–0.7 mm); ovaries and oocytes yellow in color; microscopically big oocytes with small nucleus. |
| IV | Hydrated | This stage showed bigger oocytes (diameter 0.8 mm–1 cm); translucent/hydrated oocytes distributed with yolked oocytes. |
| V | Spent/Atretic | Small oocytes with a small proportion of each stage dominated by immature oocytes. In this stage, POFs (POFs 0 and POFs 1) will appear, indicating recent spawning. |
| 2017 | 2018 | 2019 | ||||
|---|---|---|---|---|---|---|
| Month | Fecundity | Batch Fecundity | Fecundity | Batch Fecundity | Fecundity | Batch Fecundity |
| Jan | 1 | 0 | 7 | 0 | 12 | 0 |
| Feb | 17 | 4 | 29 | 0 | 30 | 5 |
| Mar | 68 | 15 | 18 | 3 | 12 | 1 |
| Apr | 28 | 0 | 56 | 10 | 21 | 5 |
| May | 3 | 0 | 2 | 0 | 4 | 0 |
| Total | 112 | 19 | 113 | 13 | 79 | 11 |
| Year | Month | Sample Number | No. POF Day-0 | No. POF Day 1+ | No. Hydrated | Spawning Fraction |
|---|---|---|---|---|---|---|
| 2017 | Jan | 8 | 0 | 0 | 1 | 0 |
| Feb | 41 | 8 | 5 | 8 | 0.15 | |
| Mar | 107 | 18 | 11 | 15 | 0.13 | |
| Apr | 40 | 2 | 11 | 0 | 0.16 | |
| May | 8 | 0 | 0 | 0 | 0 | |
| Subtotal | 204 | 28 | 27 | 24 | 0.13 * | |
| 2018 | Jan | 22 | 2 | 0 | 2 | 0.04 |
| Feb | 67 | 3 | 4 | 6 | 0.05 | |
| Mar | 36 | 0 | 2 | 9 | 0.03 | |
| Apr | 73 | 6 | 5 | 13 | 0.08 | |
| May | 5 | 0 | 0 | 0 | 0 | |
| Subtotal | 203 | 11 | 11 | 30 | 0.05 * | |
| 2019 | Jan | 20 | 1 | 2 | 1 | 0.08 |
| Feb | 41 | 3 | 2 | 10 | 0.06 | |
| Mar | 18 | 2 | 3 | 2 | 0.13 | |
| Apr | 26 | 3 | 1 | 7 | 0.08 | |
| May | 7 | 0 | 3 | 2 | 0.2 | |
| Subtotal | 112 | 9 | 11 | 22 | 0.09 * | |
| Total | 519 | 0.10 ** | ||||
| Year | Months | Total Spawn Female | Total Number of Females | Monthly Spawning (%) |
|---|---|---|---|---|
| 2017 | Jan | 0 | 8 | - |
| Feb | 13 | 41 | 31.7 | |
| Mar | 29 | 107 | 27.1 | |
| Apr | 13 | 40 | 32.5 | |
| May | 0 | 8 | - | |
| 2018 | Jan | 2 | 22 | 10 |
| Feb | 5 | 67 | 7.5 | |
| Mar | 2 | 36 | 5.6 | |
| Apr | 11 | 73 | 15 | |
| May | 0 | 5 | - | |
| 2019 | Jan | 3 | 20 | 15 |
| Feb | 5 | 41 | 12.2 | |
| Mar | 5 | 18 | 27.8 | |
| Apr | 4 | 26 | 15.4 | |
| May | 2 | 7 | 28.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinaga, S.; Lu, H.-J.; Lin, J.-R. Mackerel (Scomber australasicus) Reproduction in Northeastern Taiwan. J. Mar. Sci. Eng. 2021, 9, 1341. https://doi.org/10.3390/jmse9121341
Sinaga S, Lu H-J, Lin J-R. Mackerel (Scomber australasicus) Reproduction in Northeastern Taiwan. Journal of Marine Science and Engineering. 2021; 9(12):1341. https://doi.org/10.3390/jmse9121341
Chicago/Turabian StyleSinaga, Sunarti, Hsueh-Jung Lu, and Jia-Rong Lin. 2021. "Mackerel (Scomber australasicus) Reproduction in Northeastern Taiwan" Journal of Marine Science and Engineering 9, no. 12: 1341. https://doi.org/10.3390/jmse9121341
APA StyleSinaga, S., Lu, H. -J., & Lin, J. -R. (2021). Mackerel (Scomber australasicus) Reproduction in Northeastern Taiwan. Journal of Marine Science and Engineering, 9(12), 1341. https://doi.org/10.3390/jmse9121341