
A Very Large Spawning Aggregation of a Deep-Sea Eel: Magnitude and Status

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sampling
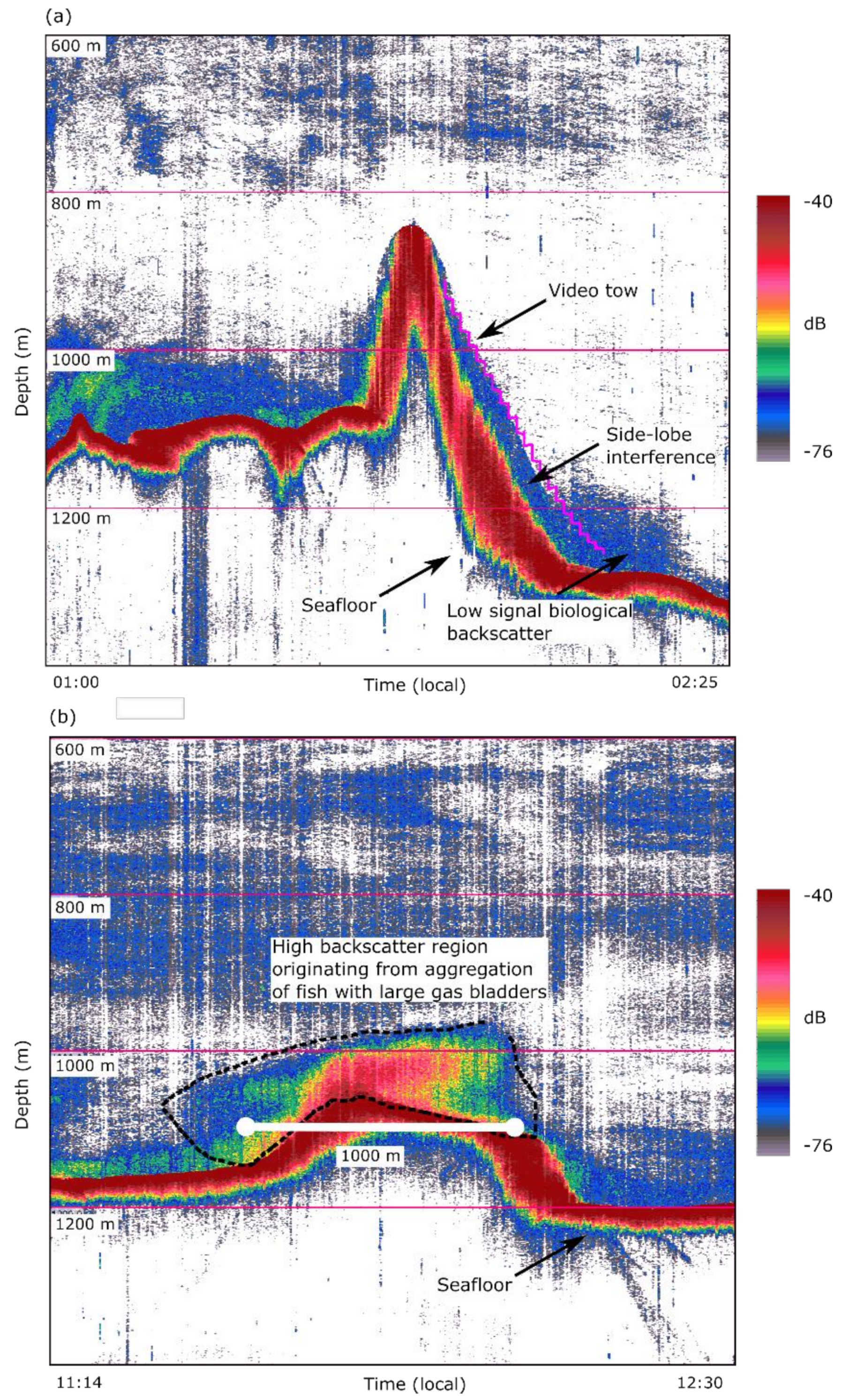
2.2. Acoustic Mapping of the Aggregation at Patience Seamount
2.3. Estimating Fish Abundance
- (1)
- Density: counts of individual fishes above (overlapping) polygons of known area were made during Survey #1 [35] for D. capensis and other fishes: predominantly orange roughy (Hoplostethus atlanticus), combined Oreosomatidae (Allocyttus verucosus and Neocyttus rhomboidalis); squaliform sharks, and macrourids. Polygon areas were estimated by using the stereo configuration of the video to determine absolute areas of seafloor in the high-resolution still images. Surface areas in the 3-dimensional perspective of the oblique field of view were calculated from calibrated stereo imagery using Photomeasure software (SeaGIS-TM V3.15 & SeaGIS-EM V5.51, www.seagis.com.au/index.html, accessed on 5 June 2021). Polygons of known area were drawn in the images by identifying a series of common points in corresponding frames from the stereo video and the still images. These polygons encompassed an average area of 5.0 m2. Differences in mean density between seamounts used a 2-tailed t-test.
- (2)
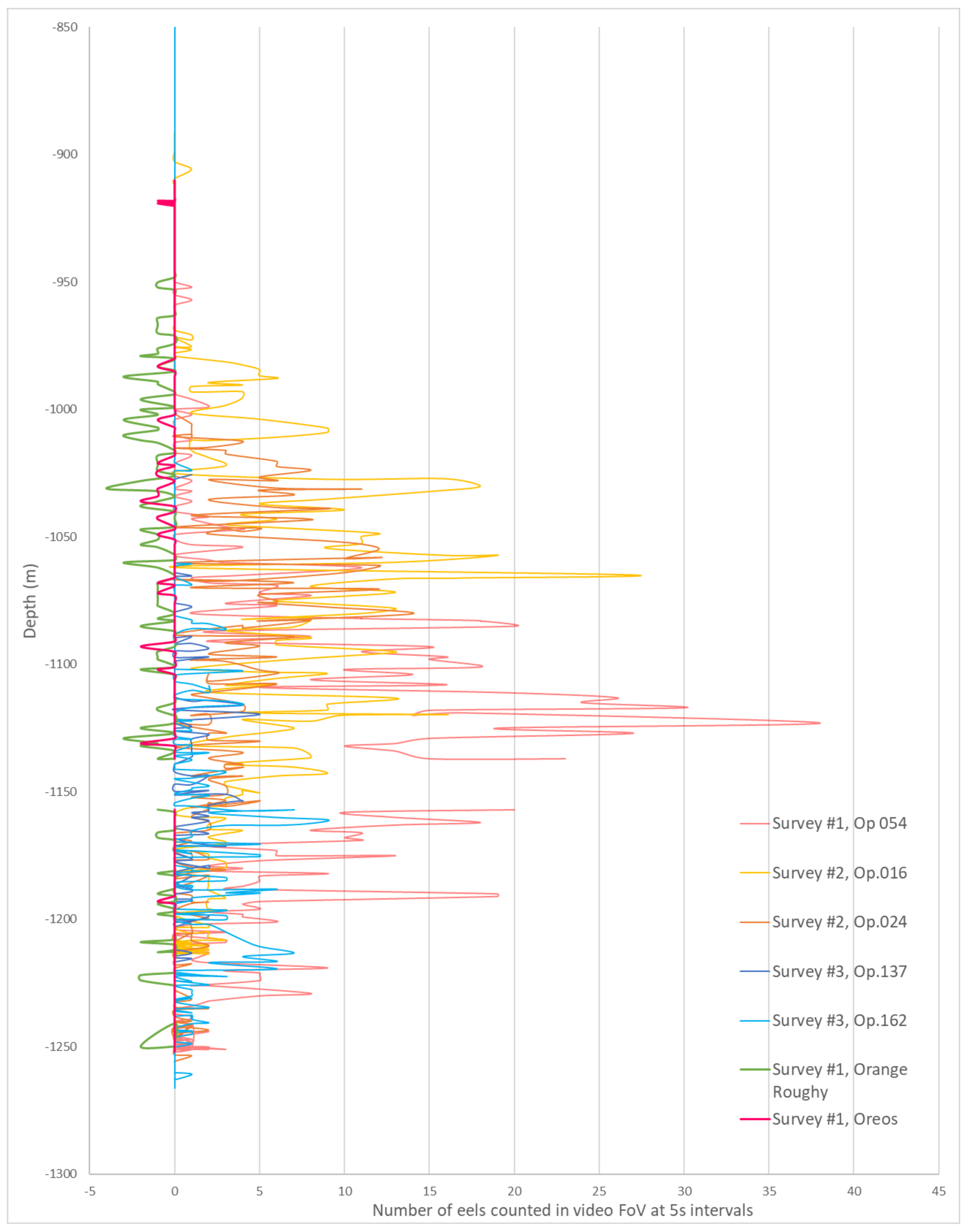
- Relative abundance: counts of individual D. capensis in the field of view in non-overlapping video-frames (5 s intervals) to provide an along-transect estimate of the total number of individuals. This provided a conservative sum of individuals because eels were not counted in the gaps between images. For survey #3, the video was viewed at half-speed and all eels in the field of view were counted. The observed number of eels was standardised to individuals per h of video tow.
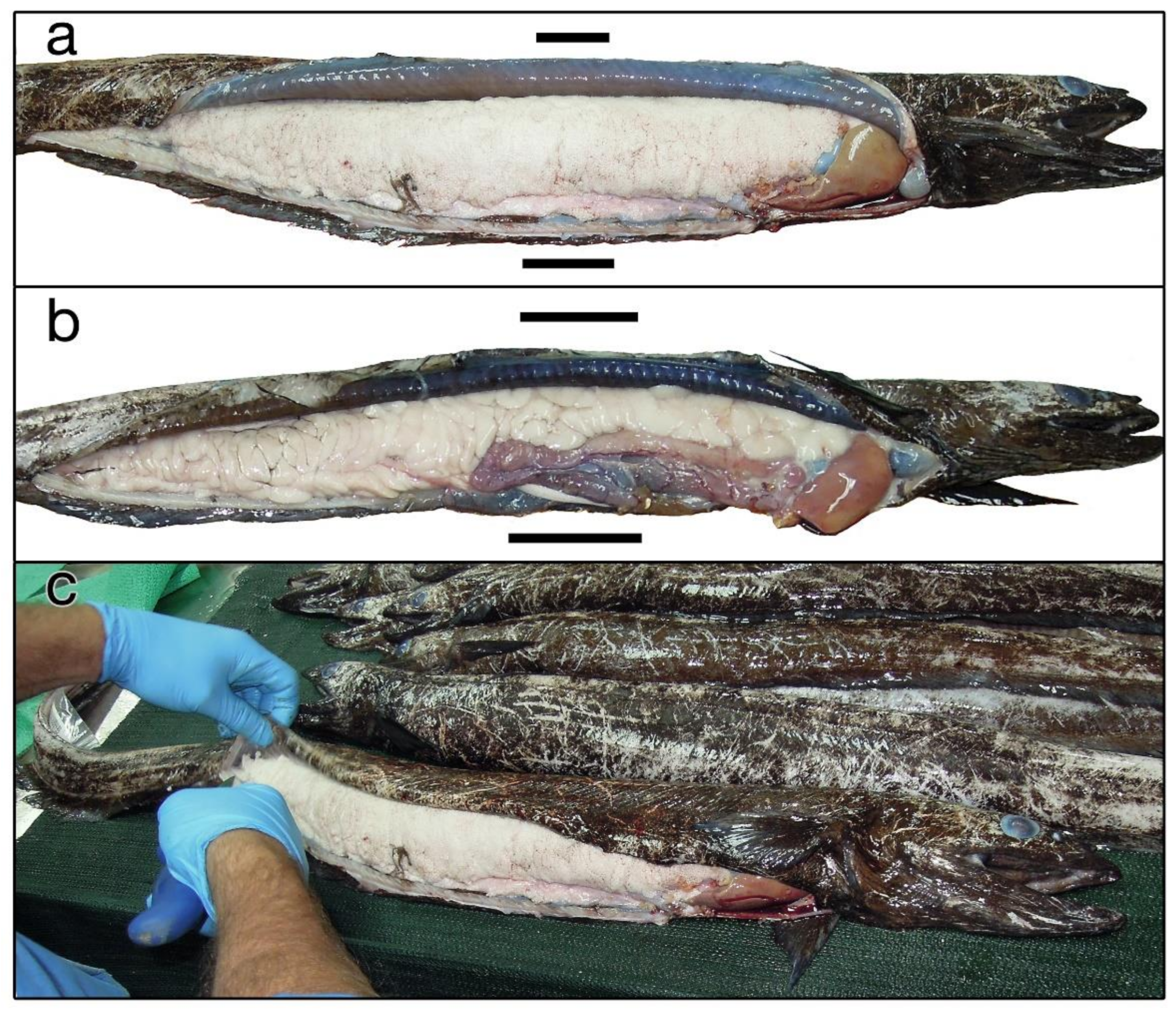
2.4. Reproductive Biology and Swimbladder of Diastobranchus capensis
3. Results
3.1. Abundance of Diastobranchus capensis
3.2. Reproductive and Swimbladder Characteristics of Diastobranchus capensis
3.3. Characteristics and Composition of Acoustic Marks at Patience Seamount
4. Discussion
4.1. Characteristics of the Aggregation of Diastobranchus capensis
4.2. Ecological Status of the Diastobranchus capensis Aggregation
4.3. Conservation Value and Recovery Monitoring
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer’s Catalog of Fishes: Genera/Species by Family/Subfamily; California Academy of Sciences: San Francisco, CA, USA, 2021; Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 12 May 2021).
- Linley, T.D.; Gerringer, M.E.; Yancey, P.H.; Drazen, J.C.; Weinstock, C.L.; Jamieson, A.J. Fishes of the hadal zone including new species, in situ observations and depth records of Liparidae. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 114, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Righton, D.; Piper, A.; Aarestrup, K.; Amilhat, E.; Belpaire, C.; Casselman, J.; Castonguay, M.; Díaz, E.; Dörner, H.; Faliex, E.; et al. Important questions to progress science and sustainable management of anguillid eels. Fish Fish. 2021, 1–27. [Google Scholar] [CrossRef]
- Chow, S.; Kurogi, H.; Mochioka, N.; Yanagimoto, T. Massive spawning behavior of the triplespine deepwater cardinalfish Sphyraenops bairdianus Poey, 1861 observed on an offshore sea mount in the West Mariana Ridge. Environ. Biol. Fish. 2018, 101, 1701–1705. [Google Scholar] [CrossRef]
- Tsukamoto, K.; Chow, S.; Otake, T.; Kurogi, H.; Mochioka, N.; Miller, M.J.; Aoyama, J.; Kimura, S.; Watanabe, S.; Yoshinaga, T.; et al. Oceanic spawning ecology of freshwater eels in the western North Pacific. Nat. Commun. 2011, 2, 179. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.J. Contrasting migratory strategies of marine and freshwater eels. Fish. Sci. 2002, 68, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Ferraris, C.J., Jr. Redescription and Spawning Behavior of the Muraenid eel Gymnothorax herrei. Copeia. 1985, 2, 518–520. [Google Scholar] [CrossRef]
- Fricke, H.W. Ökologische und verhaltensbiologische Beobachtungen an den Rohrenaalen Gorgasia sillneri und Taenioconger hassi (Pisces, Apodes, Heterocongridae); Max-Planck-Institut für Verhaltensphysiologie, Seewiesen und Erling-Andechs: Munich, Germany, 1970; pp. 1076–1099. [Google Scholar]
- Moyer, J.T.; Zaiser, M.J. Reproductive Behavior of Moray Eels at Miyake-jima, Japan. Jpn. J. Ichthyol. 1982, 28, 466–468. [Google Scholar]
- Castle, P.H.J. Synaphobranch eels from the Southern Ocean. Deep Sea Res. 1968, 15, 393–396. [Google Scholar] [CrossRef]
- Cohen, D.M.; Dean, D. Sexual maturity and migratory behaviour of the tropical eel, Ahlia egmotis. Nature 1970, 227, 189–190. [Google Scholar] [CrossRef]
- Ross, S.W.; Rohde, F.C. Colections of Ophichthid eels on the surface at night off North Carolina. Bull. Mar. Sci. 2003, 72, 241–246. [Google Scholar]
- Staudigel, H.; Hart, S.T.; Pile, A.; Bailey, B.E.; Baker, E.T.; Brooke, S.; Connelly, D.P.; Haucke, L.; German, C.R.; Hudson, I.; et al. Vailulu’u Seamount, Samoa: Life and death on an active submarine volcano. Proc. Natl. Acad. Sci. USA 2006, 103, 6448–6453. [Google Scholar] [CrossRef] [Green Version]
- Fujikura, K.; Tsuchida, S.; Ueno, H.; Ishibashi, J.; Gaze, W.; Maki, Y. Investigation of the deep-sea chemosynthetic ecosystem and submarine volcano at the Kasuga 2 and 3 seamounts in the Northern Mariana Trough, Western Pacific. JAMSTEC J. Deep Sea Res. 1998, 14, 127–138. [Google Scholar]
- Leitner, A.B.; Durden, J.M.; Smith, C.R.; Klingberg, E.D.; Drazen, J.C. Synaphobranchid eel swarms on abyssal seamounts: Largest aggregation of fishes ever observed at abyssal depths. Deep Sea Res. Part I 2021, 167, 103423. [Google Scholar] [CrossRef]
- Castle, P.H.J. Leptocephali of the Nemichthyidae, Serrivomeridae, Synaphobranchidae and Nettastmidae in Australasian waters. Trans. R. Soc. N. Z. Zool. 1965, 5, 131–146. [Google Scholar]
- Sulak, K.J.; Shcherbachev, Y.N. Zoogeography and systematics of six deep-living genera of Synaphobranchid eels, with a key to taxa and description of two new species of Ilyophis. Bull. Mar. Sci. 1997, 60, 1158–1194. [Google Scholar]
- Moore, J.A.; Vecchione, M.; Collette, B.B.; Gibbons, R.; Hartel, K.E. Selected fauna of Bear Seamount (New England Seamount chain), and the presence of “natural invader” species. Arch. Fish. Mar. Res. 2004, 51, 241–250. [Google Scholar]
- Svendsen, F.M.; Langhelle, G.; Byrkjedal, I. First record of a synaphobranchid eel in the Arctic: Diastobranchus capensis caught near Spitsbergen. Cybium 2011, 35, 255–256. [Google Scholar]
- Tracey, D.M.; Bull, B.; Clark, M.R.; MaCkay, K.A. Fish species composition on seamounts and adjacent slope in New Zealand waters. N. Z. J. Mar. Freshw. Res. 2004, 38, 163–182. [Google Scholar] [CrossRef]
- Clark, M.R.; Althaus, F.; Williams, A.; Niklitschek, E.; Menezes, G.M.; Hareide, N.-R.; Sutton, P.; O’Donnell, C. Are deep-sea demersal fish assemblages globally homogenous? Insights from seamounts. Mar. Ecology. 2010, 31, 39–51. [Google Scholar] [CrossRef]
- Zintzen, V.; Anderson, M.J.; Roberts, C.D.; Harvey, E.S.; Stewart, A.L.; Struthers, C.D. Diversity and composition of demersal fishes along a depth gradient assessed by baited remote underwater stereo-video. PLoS ONE 2012, 7, e48522. [Google Scholar] [CrossRef] [Green Version]
- Koslow, J.A.; Bulman, C.M.; Lyle, J.M. The mid-slope demersal fish community off southeastern Australia. Deep Sea Res. I 1994, 41, 113–141. [Google Scholar] [CrossRef]
- Bulman, C.M.; He, X.; Koslow, J.A. Trophic ecology of the mid-slope demersal fish community off southern Tasmania, Australia. Mar. Freshw. Res. 2002, 53, 59–72. [Google Scholar] [CrossRef]
- Anderson, M.E. Food habits of some deep-sea fish off South Africa’s west coast. 2. Eels and spiny eels (Anguilliformes and Notacanthiformes). Afr. J. Mar. Sci. 2005, 27, 557–566. [Google Scholar] [CrossRef]
- Clark, M.R.; Anderson, O.F.; Francis, R.I.C.C.; Tracey, D.M. The effects of commercial exploitation on orange roughy (Hoplestethus atlanticus) from the continental slope of the Chatham Rise, New Zealand, from 1979 to 1997. Fish. Res. 2000, 45, 217–238. [Google Scholar] [CrossRef]
- Jones, M.R.L.; Breen, B.B. Role of scavenging in a synaphobranchid eel (Diastobranchus capensis, Barnard, 1923), from northeastern Chatham Rise, New Zealand. Deep Sea Res. I 2014, 85, 118–123. [Google Scholar] [CrossRef]
- Zintzen, V.; Roberts, C.D.; Clark, M.R.; Williams, A.; Althaus, F.; Last, P.R. Composition, distribution and regional affinities of the deepwater ichthyofauna of the Lord Howe Rise and Norfolk Ridge, south-west Pacific Ocean. Deep Sea Res Part II Top. Stud. Oceanogr. 2011, 58, 933–947. [Google Scholar] [CrossRef]
- Whitley, G.P. Some noteworthy fishes from eastern Australia. Proc. R. Zool. Soc. N. S. W. 1950, 27–32. [Google Scholar]
- Olsen, A. New fish records and notes on some uncommon Tasmanian species. Pap. Proc. R. Soc. Tasman. 1958, 92, 155–159. Available online: https://eprints.utas.edu.au/14369/ (accessed on 5 June 2021).
- Australian Marine Parks Huon Marine Park. 2021. Available online: https://parksaustralia.gov.au/marine/parks (accessed on 5 June 2021).
- Williams, A.; Althaus, F.; Maguire, K.; Green, M.; Untiedt, C.; Alderslade, P.; Clark, M.R.; Bax, N.; Schlacher, T.A. The Fate of Deep-Sea Coral Reefs on Seamounts in a Fishery-Seascape: What Are the Impacts, What Remains, and What Is Protected? Front. Mar. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Marouchos, A.; Sherlock, M.; Filisetti, A.; Williams, A. Underwater imaging on self-contained tethered systems. In Proceedings of the IEEE Oceans, Anchorage, AK, USA, 18–21 September 2017; Volume 5. [Google Scholar]
- Lewis, M. The CSIRO 4m Beam Trawl; CSIRO Marine and Atmospheric Research Paper 033; CSIRO Marine and Atmospheric Research: Hobart, Australia, 2010; 17p. [Google Scholar]
- Shortis, M.R.; Seager, J.W.; Williams, A.; Barker, B.A.; Sherlock, M. Using Stereo-Video for Deep Water Benthic Habitat Surveys. Mar. Technol. Soc. J. 2008, 42, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Bulman, C.; Wayte, S.E.; Elliott, N. Orange Roughy Surveys, 1988 and 1989 Part A Abundance Indices Part B Biological Data; CSIRO Marine Laboratories: Hobart, Australia, 1994; 40p, Available online: http://www.cmar.csiro.au/e-print/open/CMReport_215.pdf (accessed on 5 June 2021).
- AFMA Observer Data. Data Collected through the Australian Fisheries Management Authority Observer Program. 2021. Available online: https://www.afma.gov.au/monitoring-enforcement/observer-program (accessed on 5 June 2021).
- McClatchie, S.; Coombs, R. Low target strength fish in mixed species assemblages: The case of orange roughy. Fish. Res. 2005, 72, 185–192. [Google Scholar] [CrossRef]
- Ona, E.; Mitson, R.B. Acoustic sampling and signal processing near the seabed: The deadzone revisited. ICES J. Mar Sci. 1996, 53, 677–690. [Google Scholar] [CrossRef]
- Kloser, R.J. Improved precision of acoustic surveys of benthopelagic fish by means of a deep-towed transducer. ICES J. Mar. Sci. 1996, 53, 407–413. [Google Scholar] [CrossRef]
- Foote, K.G.; Knudsen, H.P.; Vestnes, G.; McLennan, D.N.; Simmonds, E.J. Calibration of Acoustic Instruments for Fish Density Estimation: A Practical Guide; Cooperative Research Report No.144; Marine Laboratory: Aberdeen, UK, 1987; 72p, Available online: http://courses.washington.edu/fish538/resources/CRR%20144%20acoustic%20calibration.pdf (accessed on 5 June 2021).
- Hazen, E.L.; Horne, J.K.A. method for evaluating the effects of biological factors on fish target strength. ICES J. Mar. Sci. 2003, 60, 555–562. [Google Scholar] [CrossRef]
- Kloser, R.J.; Macaulay, G.; Ryan, T.E.; Lewis, M. Identification and target strength of orange roughy (Hopletethus atlanticus) measured in situ. J. Acoust. Soc. Am. 2013, 134, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.R. Deep sea seamount fisheries: A review of global status and future prospects. Lat. Am. J. Aquat. Res. 2009, 37, 501–512. [Google Scholar] [CrossRef]
- Upston, J.; Punt, A.E.; Wayte, S.; Ryan, T.; Day, J.; Sporcic, M. Orange Roughy (Hoplostethus atlanticus) Eastern Zone Stock Assessment Incorporating Data Up to 2014; CSIRO Oceans and Atmosphere—Australian Fisheries Management Authority: Hobart, Australia, 2014. [Google Scholar]
- Morato, T.; Cheung, W.W.L.; Pitcher, T.J. Vulnerability of seamount fish to fishing: Fuzzy analysis of life-history attributes. J. Fish Biol. 2006, 68, 209–221. [Google Scholar] [CrossRef]
- Cheung, W.W.L.; Watson, R.; Morato, T.; Pitcher, T.J.; Pauly, D. Intrinsic vulnerability in the global fish catch. Mar. Ecol. Prog. Ser. 2007, 333, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Drazen, J.C.; Haedrich, R.L. A continuum of life histories in deep-sea demersal fishes. Deep Sea Res. Part I. 2012, 61, 34–42. [Google Scholar] [CrossRef]
- McMillan, P.J.; Hart, A.C.; Horn, P.L.; Tracey, D.M.; Maolagáin, C.Ó.; Finucci, B.; Dunn, M.R. Review of life history parameters and preliminary age estimates of some New Zealand deep-sea fishes. N. Z. J. Mar. Freshw. Res. 2020, 1–27. [Google Scholar] [CrossRef]
- Livingston, M.E.; Clark, M.R.; Baird, S.J. Trends in Incidental Catch of Major Fisheries on the Chatham Rise for Fishing Years 1989–90 to 1998–99; NZ Fisheries Assessment Report 2003/52; Ministry of Fisheries: Wellington, New Zealand, 2003.
- Bailey, D.M.; Collins, M.A.; Gordon, J.D.M.; Zuur, A.F.; Priede, I.G. Long-term changes in deep-water fish populations in the North East Atlantic: Deeper-reaching effect of fisheries? Proc. R. Soc. Lond. Ser. B. 2009, 276, 1965–1969. [Google Scholar]
- Priede, I.G.; Godbold, J.A.; Niedzielski, T.; Collins, M.A.; Bailey, D.M.; Gordon, J.D.M.; Zuur, A.F. A review of the spatial extent of fishery effects and species vulnerability of the deep-sea demersal fish assemblage of the Porcupine Seabight, Northeast Atlantic Ocean (ICES Subarea VII). ICES J. Mar. Sci. 2011, 68, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Ciannelli, L.; Bailey, K.; Olsen, E.M. Evolutionary and ecological constraints of fish spawning habitats. ICES J. Mar. Sci. 2015, 72, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Koslow, J. Continental slope and deep-sea fisheries: Implications for a fragile ecosystem. ICES J. Mar. Sci. 2000, 57, 548–557. [Google Scholar] [CrossRef] [Green Version]
- Hayes, K.R.; Dunstan, P.; Woolley, S.; Barrett, N.; Foster, S.; Monk, J.; Peel, D.; Hosack, G.R.; Howe, S.A.; Samson, C.R.; et al. Designing a Targeted Monitoring Program to Support Evidence-Based Management of Australian Marine Parks: A Pilot in the South-East Marine Park Network; NESP Marine Biodiversity Hub: Hobart, Australia, 2021. [Google Scholar]
- Chollett, I.; Priest, M.; Fulton, S.; Heyman, W.D. Should we protect extirpated fish spawning aggregation sites. Biol. Conserv. 2020, 241, 108395. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SURVEY #1 | SURVEY #2 | SURVEY #3 | ||||||
|---|---|---|---|---|---|---|---|---|
| Seamount | Seamount Peak Depth (m) | Depth Range Sampled (m) | No. Images | Average No. D. capensis per m2 | No. Video Transects | Average No. D. capensis per h | No. Video Transects | Average no. D. capensis per h |
| Patience | 817 | 904–1240 | 123 | 0.259 | 2 | 220.0 | 2 | 69.59 |
| Corvina Group | 750 | 750–1250 | 0 | -- | 0 | -- | 6 | 2.07 |
| Z5 | 1178 | 1176–1250 | 22 | 0.000 | 0 | -- | 1 | 0 |
| Pedra | 714 | 725–1343 | 448 | 0.002 | 0 | -- | 7 | 4.19 |
| Mongrel | 723 | 778–1244 | 286 | 0.001 | 0 | -- | 0 | -- |
| Z16 | 1004 | 1057–1404 | 54 | 0.000 | 0 | -- | 14 | 1.15 |
| Sisters | 807 | 864–1350 | 458 | 0.001 | 0 | -- | 10 | 6.7 |
| Hill B1 | 1064 | 1139–1405 | 60 | 0.003 | 0 | -- | 0 | -- |
| Hill K1 | 1238 | 1248–1817 | 251 | 0.001 | 0 | -- | 8 | 7.82 |
| Hill U | 1083 | 1099–1413 | 247 | 0.002 | 0 | -- | 11 | 6.29 |
| Dory Hill | 1054 | 1091–1469 | 113 | 0.000 | 0 | -- | 0 | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, A.; Osterhage, D.; Althaus, F.; Ryan, T.; Green, M.; Pogonoski, J. A Very Large Spawning Aggregation of a Deep-Sea Eel: Magnitude and Status. J. Mar. Sci. Eng. 2021, 9, 723. https://doi.org/10.3390/jmse9070723
Williams A, Osterhage D, Althaus F, Ryan T, Green M, Pogonoski J. A Very Large Spawning Aggregation of a Deep-Sea Eel: Magnitude and Status. Journal of Marine Science and Engineering. 2021; 9(7):723. https://doi.org/10.3390/jmse9070723
Chicago/Turabian StyleWilliams, Alan, Deborah Osterhage, Franziska Althaus, Timothy Ryan, Mark Green, and John Pogonoski. 2021. "A Very Large Spawning Aggregation of a Deep-Sea Eel: Magnitude and Status" Journal of Marine Science and Engineering 9, no. 7: 723. https://doi.org/10.3390/jmse9070723
APA StyleWilliams, A., Osterhage, D., Althaus, F., Ryan, T., Green, M., & Pogonoski, J. (2021). A Very Large Spawning Aggregation of a Deep-Sea Eel: Magnitude and Status. Journal of Marine Science and Engineering, 9(7), 723. https://doi.org/10.3390/jmse9070723