
Preparation and Use of Decellularized Extracellular Matrix for Tissue Engineering



Abstract
:
1. Introduction
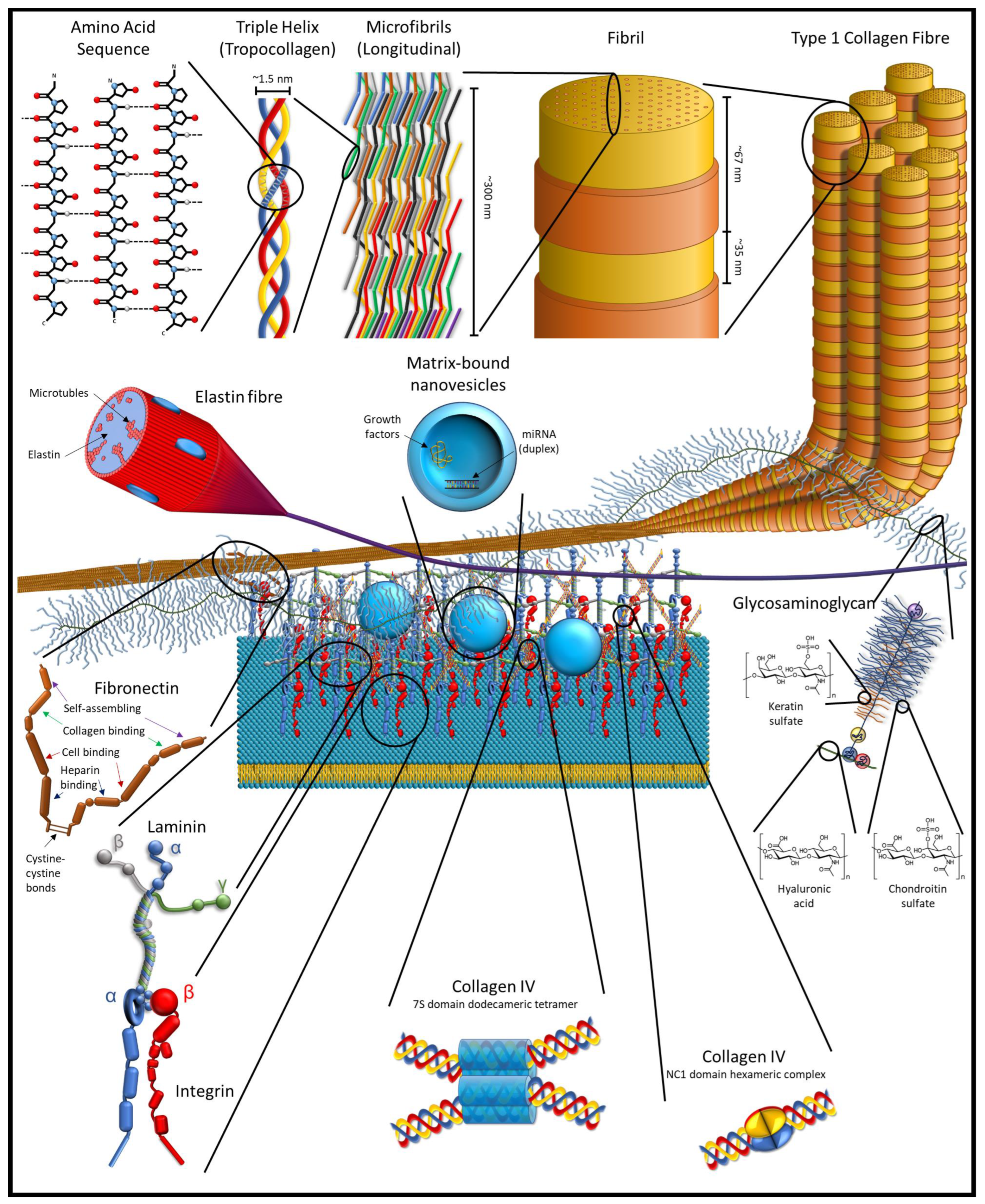
2. Structure and Properties of ECM
2.1. Xenogeneic Compatibility of ECM
2.2. Immune Response to dECM
2.3. Cryptome and Nanovesicles
- Class 1—peptides proteolytically cleaved in vivo that are novel and function very differently than their parent protein;
- Class 2—peptides proteolytically cleaved in vivo and have similar activity to their precursors; and
- Class 3—peptides produced in vitro through proteolytic digestion of proteins or recombinant technology, but may not be similar or identical to those found in vivo.

{kind=link}
{kind=link}
{kind=link}
| Parent ECM Protein | Protein Chain | Cryptein Name | MW (kDa) | Purpose in Parent Protein | Function | References |
|---|---|---|---|---|---|---|
| Collagen III | IIIα | AGVGGEKSGGF | ~1 | C terminus telopeptide | Chemotactic behaviour Increases the presence of Sox2+ and Sca1+, Lin− cells at wound site Influences osteogenesis and bone remodelling | [64,65] |
| Collagen IV | α1 | Arresten | 26 | NC1 domain | Inhibits angiogenesis (inhibits endothelial cell proliferation, migration, and tube formation) Inhibits tumour growth and metastasis | [66] |
| α2 | Canstatin | 24 | NC1 domain | Inhibited endothelial cell proliferation and migration Endothelial cell apoptosis | [67] | |
| α3 | Tumstatin | 28 | NC1 domain | Inhibits angiogenesis (amino acids 54–132) Promotes adhesion and inhibit proliferation of human melanoma cells (amino acids 185–203) Inhibit proliferation, promote apoptosis, and inhibit Akt activation (amino acids 185-191; CNYYSNS linear peptide) Reduces neovascularization (YSNSG cyclopeptide) | [68] | |
| Collagen XV | α1 | Restin | 22 | NC1 domain | Anti-angiogenic Tumour-growth inhibition | [28] |
| Collagen XVIII | Endostatin | 20 | NC1 domain | Inhibit angiogenesis Inhibits in vivo growth of primary and metastatic tumours | [28,69,70] | |
| Perlecan | Endorepellin | 81 | C terminus | Blocked adhesion of endothelial cell to fibronectin and type I collagen Binds and counter-acts endostatin | [71,72] | |
| Fibronectin | III1C | Anastellin | 10.18 | C terminus two-thirds of the first type III homology repeat | Suppress tumour growth and metastasis Inhibit angiogenesis Affects cell cycle progression | [73] |
| Laminin-332 | γ2 | EGF-like repeat | 30 | DIII | Stimulate cell migration without proliferation | [74,75] |
| Laminin-111 | β1 | β1–LN–LE1-4 fragment | 60 | N terminus | Regulates cell behaviour (e.g., epithelial-to-mesenchymal transition) Downregulates MMP2 expression | [76] |
| Elastin | xGxPGxGxG consensus sequence | ~0.75 | Stimulate cell migratory, proliferative, and morphogenic behaviours Stimulates angiogenesis Pro-tumour properties | [77,78,79,80,81] |
2.4. Effects on Cell Behaviour
3. Methods of Preparing dECM
- nuclear material not visible in tissue sections stained with either H&E or 4′,6-diamidino-2-phenylindole (DAPI);
- dsDNA content < 50 ng/mg of ECM (dry weight); and
3.1. Decellularization in Chemical Baths
3.2. Decellularization by Perfusion
3.3. Cell-Cultured ECM
3.4. Advances in ECM Decellularization
3.4.1. Vacuum
3.4.2. Hydrostatic Washing
3.4.3. Pulsatile Perfusion
3.4.4. Chemical–Penetration Enhancement
3.4.5. Sonication
3.4.6. Nonthermal Irreversible Electroporation (NTIRE)
3.4.7. Decellularizing Agents
3.4.8. Supercritical Fluids
3.4.9. Alternating Decellularizing Solutions
4. ECM Modification and Methods
4.1. Improving Structural Stability
4.1.1. Composite dECM Scaffolds
4.1.2. Cross-Linking
4.1.3. Structural Fabrication
4.2. Improving Fibrous Structure
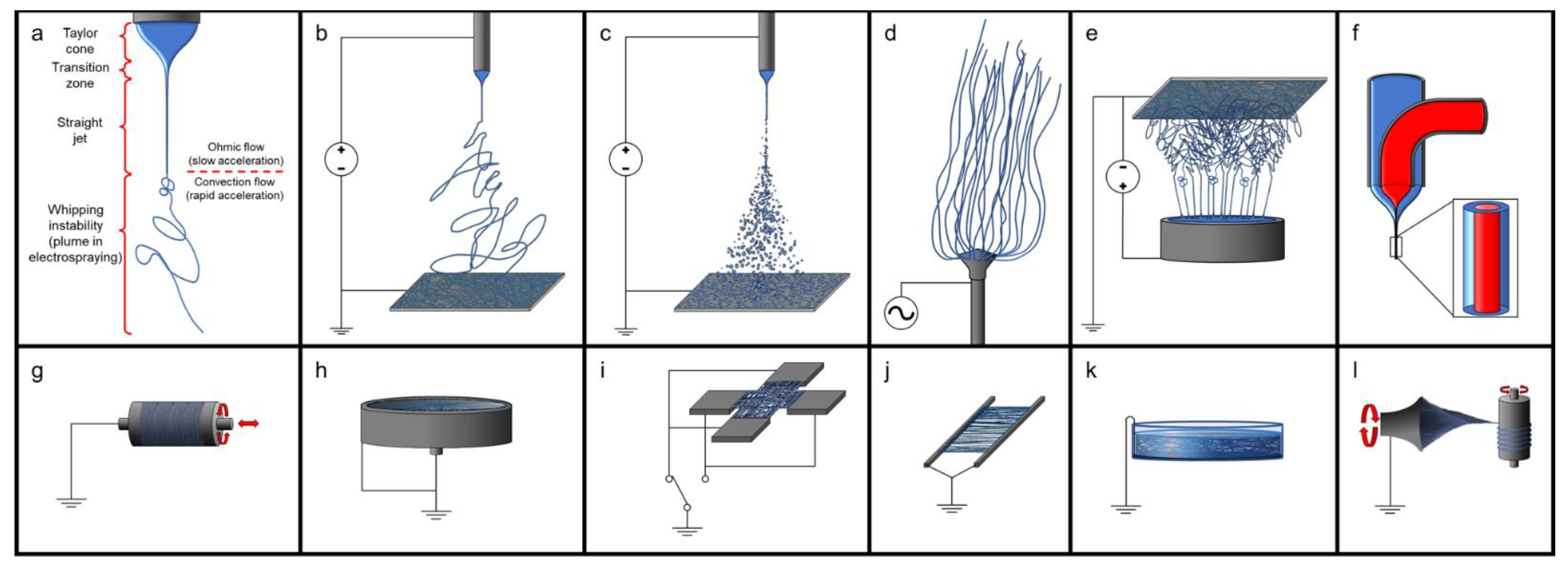
4.2.1. Electrospinning Solubilized dECM
4.2.2. Modifying dECM for Enhanced Engraftment
4.3. In Vivo Use of dECM
4.4. Solubilizing dECM for Bioinks
Inks and Bioinks
4.5. Sterilization
4.6. Cell Seeding of dECM
- Totipotent stem cells that can produce all of the cell types in a foetus (including the birth-associated tissues: placenta, amnion, etc., that are derived from the trophoectoderm);
- Pluripotent stem cells that can produce cells from all three germ layers (i.e., endoderm, mesoderm, and ectoderm);
- Multipotent stem cells that can produce a limited number of cell types of different lineages within one of the germ layers (though some research is showing that there is the potential for these stem cells to differentiate into the cell types of other germ layers);
- Oligopotent stem cells can produce two or more cell types within a specific tissue (these are sometimes divided from multipotent stem cells as an intermediate step); and
- Unipotent stem cells are terminally differentiated stem cells that can produce cells of only one type [351].
5. Applications of dECM
5.1. In Vitro Cell Culture with dECM
5.2. ECM Extract
5.3. Clinical Use of dECM
| Source | Targeted Condition | Phase | Recruitment Status | Outcome | Material | Year Posted | Reference |
|---|---|---|---|---|---|---|---|
| Porcine small intestine submucosa | Rotator cuff tear | 4 | Recruiting | N/A | ArthroFLEX ECM scaffold graft | 2018 | [368] |
| Fish skin | Chronic wounds | N/A | Completed | Not reported | MariGen Wound Dressing | 2011 | [369] |
| Adipose | Obesity | N/A | Completed | Not reported | Adipose allograft extracellular matrix | 2016 | [370] |
| N/A | Ischemic cardiomyopathy | 1/2 | Not yet recruiting | N/A | Wharton’s jelly-derived mesenchymal cells seeded onto an extracellular matrix patch | 2019 | [371] |
| Porcine urinary bladder | Neuropathic diabetic foot ulcer | N/A | Completed | Not reported | MatriStem | 2016 | [372] |
| Porcine myocardium | Myocardial infarction-induced heart failure | 1 | Completed | Not reported | VentriGel | 2014 | [373] |
| Human dECM | Articular cartilage repair in microfracture surgery | 1/2 | Recruiting | N/A | HST-003 | 2021 | [374] |
| Porcine small intestine submucosa | Pericardial reconstruction | N/A | Completed | Elevated pro-inflammatory proteins in blood for all patients (similar to control not treated with CorMatrix) No adverse events for the treatment or control groups | CorMatrix ECM | 2014 | [375] |
| Porcine small intestine submucosa | Implantable electronic device placement for cardiovascular diseases | N/A | Completed | Of the 1025 patients in the SECURE trial: 14 had an ECM-related adverse event, possibly related to CanGaroo 2 had an ECM-related adverse event, probably related to CanGaroo 12 had major pocket infections | Cormatrix CanGaroo ECM Envelope | 2015 | [376] |
| Porcine small intestine submucosa | Pericardial reconstruction following coronary artery bypass graft surgery | N/A | Terminated | Elevated pro-inflammatory proteins in blood for all patients (similar to control not treated with CorMatrix) No adverse events for the treatment or control groups | CorMatrix ECM | 2012 | [377] |
| Porcine small intestine submucosa | Chronic wounds | N/A | Completed | Not reported | Oasis Extracellular Matrix | 2018 | [378] |
| Human adipose tissue | Soft Tissue Injuries | 1 | Completed | Graft demonstrated satisfactory safety results No participants experienced serious nor unanticipated adverse events (all were expected and mild) | Acellular adipose tissue | 2016 | [379] |
| Human adipose tissue | Soft Tissue Injuries | 2 | Active, not recruiting | N/A | Acellular adipose tissue | 2018 | [380] |
| Porcine small intestine submucosa | Inguinal hernia repair | 4 | Completed | Not reported | Surgisis Inguinal Hernia Matrix | 2008 | [381] |
| Ovine forestomach | Reconstruction of soft tissues | 4 | Recruiting | N/A | Myriad Matrix, Myriad Morcells | 2022 | [382] |
| Porcine urinary bladder | Pressure ulcer | N/A | Completed | 2 of 20 patients had complete wound epithelization at 12 weeks | MicroMatrix ACell Cytal Wound Matrix 2-Layer | 2017 | [383] |
| Porcine dECM | Volumetric muscle loss | N/A | Completed | Average improvement of strength: 37.3% Average improvement in range-of-motion tasks: 27.1% No serious adverse events | ACell, Matristem Cook, BioDesign Bard, XenMatrix | 2011 | [361,384] |
| Porcine small intestine submucosa Fish skin | Punch biopsy wounds | N/A | Completed | Not reported | Oasis ECM (porcine) MariGen Wound ECM dressing (fish) | 2013 | [385] |
6. Future Directions
6.1. Optimizing Decellularization
6.2. Stabilizing dECM
6.3. Fabricating Custom Scaffolds
6.4. Recellularization
6.5. Mechanism of Action of ECM
6.6. Clinical Use of dECM for Regenerative Medicine
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Borrelli, M.R.; Hu, M.S.; Longaker, M.T.; Lorenz, H.P. Tissue Engineering and Regenerative Medicine in Craniofacial Reconstruction and Facial Aesthetics. J. Craniofacial Surg. 2020, 31, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Katari, R.; Peloso, A.; Orlando, G. Tissue Engineering and Regenerative Medicine: Semantic Considerations for an Evolving Paradigm. Front. Bioeng. Biotechnol. 2015, 2, 57. [Google Scholar] [CrossRef] [PubMed]
- Klemm, D.; Heublein, B.; Fink, H.-P.; Bohn, A. Cellulose: Fascinating Biopolymer and Sustainable Raw Material. Angew. Chem. Int. Ed. 2005, 44, 3358–3393. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 Paradigm of Macrophage Activation: Time for Reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Keane, T.J.; Londono, R.; Turner, N.J.; Badylak, S.F. Consequences of Ineffective Decellularization of Biologic Scaffolds on the Host Response. Biomaterials 2012, 33, 1771–1781. [Google Scholar] [CrossRef]
- Valentin, J.E. Macrophage Involvement in the Remodelling of an Extracellular Matrix Scaffold. Ph.D. Thesis, University of Pittsburgh, Pittsburgh, PA, USA, 2009. [Google Scholar]
- Sicari, B.M.; Dziki, J.L.; Siu, B.F.; Medberry, C.J.; Dearth, C.L.; Badylak, S.F. The Promotion of a Constructive Macrophage Phenotype by Solubilized Extracellular Matrix. Biomaterials 2014, 35, 8605–8612. [Google Scholar] [CrossRef]
- Dziki, J.L.; Wang, D.S.; Pineda, C.; Sicari, B.M.; Rausch, T.; Badylak, S.F. Solubilized Extracellular Matrix Bioscaffolds Derived from Diverse Source Tissues Differentially Influence Macrophage Phenotype. J. Biomed. Mater. Res. A 2017, 105, 138–147. [Google Scholar] [CrossRef]
- Loneker, A.E.; Faulk, D.M.; Hussey, G.S.; D’Amore, A.; Badylak, S.F. Solubilized Liver Extracellular Matrix Maintains Primary Rat Hepatocyte Phenotype In-Vitro. J. Biomed. Mater. Res. A 2016, 104, 957–965. [Google Scholar] [CrossRef]
- Sellaro, T.L.; Ravindra, A.K.; Stolz, D.B.; Badylak, S.F. Maintenance of Hepatic Sinusoidal Endothelial Cell Phenotype In Vitro Using Organ-Specific Extracellular Matrix Scaffolds. Tissue Eng 2007, 13, 2301–2310. [Google Scholar] [CrossRef]
- Sellaro, T.L.; Ranade, A.; Faulk, D.M.; McCabe, G.P.; Dorko, K.; Badylak, S.F.; Strom, S.C. Maintenance of Human Hepatocyte Function In Vitro by Liver-Derived Extracellular Matrix Gels. Tissue Eng. Part A 2010, 16, 1075–1082. [Google Scholar] [CrossRef]
- Ijima, H.; Nakamura, S.; Bual, R.P.; Yoshida, K. Liver-Specific Extracellular Matrix Hydrogel Promotes Liver-Specific Functions of Hepatocytes in Vitro and Survival of Transplanted Hepatocytes in Vivo. J. Biosci. Bioeng. 2019, 128, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Coronado, R.E.; Somaraki-Cormier, M.; Ong, J.L.; Halff, G.A. Hepatocyte-like Cells Derived from Human Amniotic Epithelial, Bone Marrow, and Adipose Stromal Cells Display Enhanced Functionality When Cultured on Decellularized Liver Substrate. Stem. Cell Re.s 2019, 38, 101471. [Google Scholar] [CrossRef]
- Chaimov, D.; Baruch, L.; Krishtul, S.; Meivar-levy, I.; Ferber, S.; Machluf, M. Innovative Encapsulation Platform Based on Pancreatic Extracellular Matrix Achieve Substantial Insulin Delivery. J. Control Release 2017, 257, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Werb, Z.; Kain, K.H. The Extracellular Matrix and Disease: An Interview with Zena Werb. Interviewed by Kristin H. Kain. Dis. Model. Mech. 2010, 3, 513–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornstein, P. Cell-Matrix Interactions: The View from the Outside. Methods Cell Biol. 2002, 69, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The Extracellular Matrix at a Glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hynes, R.O.; Naba, A. Overview of the Matrisome—An Inventory of Extracellular Matrix Constituents and Functions. Cold Spring Harb. Perspect. Biol. 2012, 4, a004903. [Google Scholar] [CrossRef] [Green Version]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The Extracellular Matrix: Tools and Insights for the “Omics” Era. Matrix Biol. 2016, 49, 10–24. [Google Scholar] [CrossRef]
- Kuljanin, M.; Brown, C.F.C.; Raleigh, M.J.; Lajoie, G.A.; Flynn, L.E. Collagenase Treatment Enhances Proteomic Coverage of Low-Abundance Proteins in Decellularized Matrix Bioscaffolds. Biomaterials 2017, 144, 130–143. [Google Scholar] [CrossRef]
- Bornstein, P. Matricellular Proteins: An Overview. J. Cell Commun. Signal. 2009, 3, 163–165. [Google Scholar] [CrossRef]
- Bornstein, P.; Sage, E.H. Matricellular Proteins: Extracellular Modulators of Cell Function. Curr. Opin. Cell Biol. 2002, 14, 608–616. [Google Scholar] [CrossRef]
- Huleihel, L.; Hussey, G.S.; Naranjo, J.D.; Zhang, L.; Dziki, J.L.; Turner, N.J.; Stolz, D.B.; Badylak, S.F. Matrix-Bound Nanovesicles within ECM Bioscaffolds. Sci. Adv. 2016, 2, e1600502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huleihel, L.; Bartolacci, J.G.; Dziki, J.L.; Vorobyov, T.; Arnold, B.; Scarritt, M.E.; Pineda Molina, C.; LoPresti, S.T.; Brown, B.N.; Naranjo, J.D.; et al. Matrix-Bound Nanovesicles Recapitulate Extracellular Matrix Effects on Macrophage Phenotype. Tissue Eng. Part A 2017, 23, 1283–1294. [Google Scholar] [CrossRef] [PubMed]
- Hussey, G.S.; Molina, C.P.; Cramer, M.C.; Tyurina, Y.Y.; Tyurin, V.A.; Lee, Y.C.; El-Mossier, S.O.; Murdock, M.H.; Timashev, P.S.; Kagan, V.E.; et al. Lipidomics and RNA Sequencing Reveal a Novel Subpopulation of Nanovesicle within Extracellular Matrix Biomaterials. Sci. Adv. 2020, 6, eaay4361. [Google Scholar] [CrossRef] [Green Version]
- Schenk, S.; Quaranta, V. Tales from the Crypt[Ic] Sites of the Extracellular Matrix. Trends Cell Biol. 2003, 13, 366–375. [Google Scholar] [CrossRef]
- Ner, J.; Kotlinska, J.H.; Silberring, J. Crypteins—An Overlooked Piece of Peptide Systems. Curr Protein Pept Sci 2015, 16, 203–218. [Google Scholar] [CrossRef]
- Autelitano, D.J.; Rajic, A.; Smith, A.I.; Berndt, M.C.; Ilag, L.L.; Vadas, M. The Cryptome: A Subset of the Proteome, Comprising Cryptic Peptides with Distinct Bioactivities. Drug Discov. Today 2006, 11, 306–314. [Google Scholar] [CrossRef]
- Davis, G.E.; Bayless, K.J.; Davis, M.J.; Meininger, G.A. Regulation of Tissue Injury Responses by the Exposure of Matricryptic Sites within Extracellular Matrix Molecules. Am. J. Pathol. 2000, 156, 1489–1498. [Google Scholar] [CrossRef] [Green Version]
- Nishimichi, N.; Higashikawa, F.; Kinoh, H.H.; Tateishi, Y.; Matsuda, H.; Yokosaki, Y. Polymeric Osteopontin Employs Integrin A9β1 as a Receptor and Attracts Neutrophils by Presenting a de Novo Binding Site. J. Biol. Chem. 2009, 284, 14769–14776. [Google Scholar] [CrossRef] [Green Version]
- Schweitzer, M.H.; Suo, Z.; Avci, R.; Asara, J.M.; Allen, M.A.; Arce, F.T.; Horner, J.R. Analyses of Soft Tissue from Tyrannosaurus Rex Suggest the Presence of Protein. Science 2007, 316, 277–280. [Google Scholar] [CrossRef]
- Schweitzer, M.H.; Zheng, W.; Organ, C.L.; Avci, R.; Suo, Z.; Freimark, L.M.; Lebleu, V.S.; Duncan, M.B.; Vander Heiden, M.G.; Neveu, J.M.; et al. Biomolecular Characterization and Protein Sequences of the Campanian Hadrosaur B. Canadensis. Science 2009, 324, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Schroeter, E.R.; DeHart, C.J.; Cleland, T.P.; Zheng, W.; Thomas, P.M.; Kelleher, N.L.; Bern, M.; Schweitzer, M.H. Expansion for the Brachylophosaurus Canadensis Collagen I Sequence and Additional Evidence of the Preservation of Cretaceous Protein. J. Proteome. Res. 2017, 16, 920–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-C.; Chiang, C.-C.; Huang, P.-Y.; Chung, C.-Y.; Huang, T.D.; Wang, C.-C.; Chen, C.-I.; Chang, R.-S.; Liao, C.-H.; Reisz, R.R. Evidence of Preserved Collagen in an Early Jurassic Sauropodomorph Dinosaur Revealed by Synchrotron FTIR Microspectroscopy. Nat. Commun. 2017, 8, 14220. [Google Scholar] [CrossRef] [Green Version]
- Organ, C.L.; Schweitzer, M.H.; Zheng, W.; Freimark, L.M.; Cantley, L.C.; Asara, J.M. Molecular Phylogenetics of Mastodon and Tyrannosaurus Rex. Science 2008, 320, 499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stover, D.A.; Verrelli, B.C. Comparative Vertebrate Evolutionary Analyses of Type I Collagen: Potential of COL1a1 Gene Structure and Intron Variation for Common Bone-Related Diseases. Mol. Biol. Evol. 2011, 28, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Lopresti, S.T.; Brown, B.N. Host Response to Naturally Derived Biomaterials. In Host Response to Biomaterials: The Impact of Host Response on Biomaterial Selection; Badylak, S.F., Ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 53–79. ISBN 9780128001967. [Google Scholar]
- van der Rest, M.; Garrone, R. Collagen Family of Proteins. FASEB J. 1991, 5, 2814–2823. [Google Scholar] [CrossRef] [Green Version]
- Macrophage Polarization Mini Review; Bio-Rad Laboratories, Inc.: Kidlington, UK, 2016.
- Rőszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediators Inflamm 2015, 2015, 816460. [Google Scholar] [CrossRef] [Green Version]
- Huleihel, L.; Dziki, J.L.; Bartolacci, J.G.; Rausch, T.; Scarritt, M.E.; Cramer, M.C.; Vorobyov, T.; LoPresti, S.T.; Swineheart, I.T.; White, L.J.; et al. Macrophage Phenotype in Response to ECM Bioscaffolds. Semin Immunol 2017, 29, 2–13. [Google Scholar] [CrossRef]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS–) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef]
- Novak, M.L.; Koh, T.J. Macrophage Phenotypes during Tissue Repair. J. Leukoc. Biol. 2013, 93, 875–881. [Google Scholar] [CrossRef]
- Badylak, S.F.; Dziki, J.L.; Sicari, B.M.; Ambrosio, F.; Boninger, M.L. Mechanisms by Which Acellular Biologic Scaffolds Promote Functional Skeletal Muscle Restoration. Biomaterials 2016, 103, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Allman, A.J.; McPherson, T.B.; Badylak, S.F.; Merrill, L.C.; Kallakury, B.; Sheehan, C.; Raeder, R.H.; Metzger, D.W. Xenogeneic Extracellular Matrix Grafts Elicit a TH2-Restricted Immune Response. Transplantation 2001, 71, 1631–1640. [Google Scholar] [CrossRef] [PubMed]
- Allman, A.J.; McPherson, T.B.; Merrill, L.C.; Badylak, S.F.; Metzger, D.W. The Th2-Restricted Immune Response to Xenogeneic Small Intestinal Submucosa Does Not Influence Systemic Protective Immunity to Viral and Bacterial Pathogens. Tissue Eng. 2002, 8, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Witherel, C.E.; Sao, K.; Brisson, B.K.; Han, B.; Volk, S.W.; Petrie, R.J.; Han, L.; Spiller, K.L. Regulation of Extracellular Matrix Assembly and Structure by Hybrid M1/M2 Macrophages. Biomaterials 2021, 269, 120667. [Google Scholar] [CrossRef]
- Valentin, J.E.; Stewart-Akers, A.M.; Gilbert, T.W.; Badylak, S.F. Macrophage Participation in the Degradation and Remodelling of Extracellular Matrix Scaffolds. Tissue Eng. Part A 2009, 15, 1687–1694. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.N.; Londono, R.; Tottey, S.; Zhang, L.; Kukla, K.A.; Wolf, M.T.; Daly, K.A.; Reing, J.E.; Badylak, S.F. Macrophage Phenotype as a Predictor of Constructive Remodelling Following the Implantation of Biologically Derived Surgical Mesh Materials. Acta Biomater. 2012, 8, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.J.; Yates, A.J.; Weber, D.J.; Qureshi, I.R.; Stolz, D.B.; Gilbert, T.W.; Badylak, S.F. Xenogeneic Extracellular Matrix as an Inductive Scaffold for Regeneration of a Functioning Musculotendinous Junction. Tissue Eng. Part A 2010, 16, 3309–3317. [Google Scholar] [CrossRef]
- Turner, N.J.; Badylak, J.S.; Weber, D.J.; Badylak, S.F. Biologic Scaffold Remodelling in a Dog Model of Complex Musculoskeletal Injury. J. Surg. Res. 2012, 176, 490–502. [Google Scholar] [CrossRef]
- Qiu, X.; Liu, S.; Zhang, H.; Zhu, B.; Su, Y.; Zheng, C.; Tian, R.; Wang, M.; Kuang, H.; Zhao, X.; et al. Mesenchymal Stem Cells and Extracellular Matrix Scaffold Promote Muscle Regeneration by Synergistically Regulating Macrophage Polarization toward the M2 Phenotype. Stem. Cell Res. Ther. 2018, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Holder, E.J. The Story of Catgut. Postgrad. Med. J. 1949, 25, 427–433. [Google Scholar] [CrossRef]
- Ben Abdessalem, S.; Debbabi, F.; Jedda, H.; Elmarzougui, S.; Mokhtar, S. Tensile and Knot Performance of Polyester Braided Sutures. Text. Res. J. 2009, 79, 247–252. [Google Scholar] [CrossRef]
- Jackson, J.D. Immunology: Host Response to Biomaterials. In In Situ Tissue Regeneration: Host Cell Recruitment and Biomaterial Design; Lee, S.J., Atala, A., Yoo, J.J., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 35–47. ISBN 9780128022252. [Google Scholar]
- Kayabolen, A.; Keskin, D.; Aykan, A.; Karslıoglu, Y.; Zor, F.; Tezcaner, A. Native Extracellular Matrix/Fibroin Hydrogels for Adipose Tissue Engineering with Enhanced Vascularization. Biomed. Mater. 2017, 12, 035007. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.Y.; Seo, Y.; Davaa, G.; Kim, H.W.; Kim, S.H.; Hyun, J.K. Decellularized Brain Matrix Enhances Macrophage Polarization and Functional Improvements in Rat Spinal Cord Injury. Acta Biomater. 2020, 101, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.V.; Atala, A. 3D Bioprinting of Tissues and Organs. Nat. Biotechnol. 2014, 32, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Sage, E.H. Pieces of Eight: Bioactive Fragments of Extracellular Proteins as Regulators of Angiogenesis. Trends Cell Biol. 1997, 7, 182–186. [Google Scholar] [CrossRef]
- Asmussen, N.C.; Cohen, D.J.; Lin, Z.; McClure, M.J.; Boyan, B.D.; Schwartz, Z. Specific MicroRNAs Found in Extracellular Matrix Vesicles Regulate Proliferation and Differentiation in Growth Plate Chondrocytes. Calcif. Tissue Int. 2021, 109, 455–468. [Google Scholar] [CrossRef]
- de Castro Brás, L.E.; Frangogiannis, N.G. Extracellular Matrix-Derived Peptides in Tissue Remodelling and Fibrosis. Matrix Biol. 2020, 91–92, 176–187. [Google Scholar] [CrossRef]
- Vanderlugt, C.L.; Miller, S.D. Epitope Spreading in Immune-Mediated Diseases: Implications for Immunotherapy. Nat. Rev. Immunol. 2002, 2, 85–95. [Google Scholar] [CrossRef]
- Chen, L.; Qin, L.; Chen, C.; Hu, Q.; Wang, J.; Shen, J. Serum Exosomes Accelerate Diabetic Wound Healing by Promoting Angiogenesis and ECM Formation. Cell Biol. Int. 2021, 45, 1976–1985. [Google Scholar] [CrossRef]
- Agrawal, V.; Tottey, S.; Johnson, S.A.; Freund, J.M.; Siu, B.F.; Badylak, S.F. Recruitment of Progenitor Cells by an Extracellular Matrix Cryptic Peptide in a Mouse Model of Digit Amputation. Tissue Eng. Part A 2011, 17, 2435–2443. [Google Scholar] [CrossRef]
- Agrawal, V.; Kelly, J.; Tottey, S.; Daly, K.A.; Johnson, S.A.; Siu, B.F.; Reing, J.; Badylak, S.F. An Isolated Cryptic Peptide Influences Osteogenesis and Bone Remodelling in an Adult Mammalian Model of Digit Amputation. Tissue Eng. Part A 2011, 17, 3033–3044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colorado, P.C.; Torre, A.; Kamphaus, G.; Maeshima, Y.; Hopfer, H.; Takahashi, K.; Volk, R.; Zamborsky, E.D.; Herman, S.; Sarkar, P.K.; et al. Anti-Angiogenic Cues from Vascular Basement Membrane Collagen. Cancer Res. 2000, 60, 2520–2526. [Google Scholar] [PubMed]
- Kamphaus, G.D.; Colorado, P.C.; Panka, D.J.; Hopfer, H.; Ramchandran, R.; Torre, A.; Maeshima, Y.; Mier, J.W.; Sukhatme, V.P.; Kalluri, R. Canstatin, a Novel Matrix-Derived Inhibitor of Angiogenesis and Tumour Growth. J. Biol. Chem. 2000, 275, 1209–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeshima, Y.; Colorado, P.C.; Kalluri, R. Two RGD-Independent Avβ3 Integrin Binding Sites on Tumstatin Regulate Distinct Anti-Tumour Properties. J. Biol. Chem. 2000, 275, 23745–23750. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, M.S.; Boehm, T.; Shing, Y.; Fukai, N.; Vasios, G.; Lane, W.S.; Flynn, E.; Birkhead, J.R.; Olsen, B.R.; Folkman, J. Endostatin: An Endogenous Inhibitor of Angiogenesis and Tumour Growth. Cell 1997, 88, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Marneros, A.G.; Olsen, B.R. The Role of Collagen-Derived Proteolytic Fragments in Angiogenesis. Matrix Biol. 2001, 20, 337–345. [Google Scholar] [CrossRef]
- Goyal, A.; Pal, N.; Concannon, M.; Paul, M.; Doran, M.; Poluzzi, C.; Sekiguchi, K.; Whitelock, J.M.; Neill, T.; Iozzo, R.V. Endorepellin, the Angiostatic Module of Perlecan, Interacts with Both the A2β1 Integrin and Vascular Endothelial Growth Factor Receptor 2 (VEGFR2): A Dual Receptor Antagonism. J. Biol. Chem. 2011, 286, 25947–25962. [Google Scholar] [CrossRef] [Green Version]
- Mongiat, M.; Sweeney, S.M.; San Antonio, J.D.; Fu, J.; Iozzo, R.V. Endorepellin, a Novel Inhibitor of Angiogenesis Derived from the C Terminus of Perlecan. J. Biol. Chem. 2003, 278, 4238–4249. [Google Scholar] [CrossRef] [Green Version]
- Ambesi, A.; Klein, R.M.; Pumiglia, K.M.; McKeown-Longo, P.J. Anastellin, a Fragment of the First Type III Repeat of Fibronectin, Inhibits Extracellular Signal-Regulated Kinase and Causes G1 Arrest in Human Microvessel Endothelial Cells. Cancer Res. 2005, 65, 148–156. [Google Scholar] [CrossRef]
- Koshikawa, N.; Minegishi, T.; Sharabi, A.; Quaranta, V.; Seiki, M. Membrane-Type Matrix Metalloproteinase-1 (MT1-MMP) Is a Processing Enzyme for Human Laminin Γ2 Chain. J. Biol. Chem. 2005, 280, 88–93. [Google Scholar] [CrossRef]
- Schenk, S.; Hintermann, E.; Bilban, M.; Koshikawa, N.; Hojilla, C.; Khokha, R.; Quaranta, V. Binding to EGF Receptor of a Laminin-5 EGF-like Fragment Liberated during MMP-Dependent Mammary Gland Involution. J. Cell Biol. 2003, 161, 197–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horejs, C.M.; Serio, A.; Purvis, A.; Gormley, A.J.; Bertazzo, S.; Poliniewicz, A.; Wang, A.J.; DiMaggio, P.; Hohenester, E.; Stevens, M.M. Biologically-Active Laminin-111 Fragment That Modulates the Epithelial-to-Mesenchymal Transition in Embryonic Stem Cells. Proc. Natl. Acad. Sci. USA 2014, 111, 5908–5913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, J.; Lameiras, P.; Beljebbar, A.; Berquand, A.; Villemin, M.; Ramont, L.; Dukic, S.; Nuzillard, J.M.; Molinari, M.; Gautier, M.; et al. Structural Characterization and in Vivo Pro-Tumour Properties of a Highly Conserved Matrikine. Oncotarget 2018, 9, 17839–17857. [Google Scholar] [CrossRef] [PubMed]
- Pocza, P.; Süli-Vargha, H.; Darvas, Z.; Falus, A. Locally Generated VGVAPG and VAPG Elastin-Derived Peptides Amplify Melanoma Invasion via the Galectin-3 Receptor. Int. J. Cancer 2007, 122, 1972–1980. [Google Scholar] [CrossRef]
- Antonicelli, F.; Bellon, G.; Lorimier, S.; Hornebeck, W. Role of the Elastin Receptor Complex (S-Gal/Cath-A/Neu-1) in Skin Repair and Regeneration. Wound Repair Regen. 2009, 17, 631–638. [Google Scholar] [CrossRef]
- Robinet, A.; Fahem, A.; Cauchard, J.H.; Huet, E.; Vincent, L.; Lorimier, S.; Antonicelli, F.; Soria, C.; Crepin, M.; Hornebeck, W.; et al. Elastin-Derived Peptides Enhance Angiogenesis by Promoting Endothelial Cell Migration and Tubulogenesis through Upregulation of MT1-MMP. J. Cell Sci. 2005, 118, 343–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brassart, B.; Randoux, A.; Hornebeck, W.; Emonard, H. Regulation of Matrix Metalloproteinase-2 (Gelatinase A, MMP-2), Membrane-Type Matrix Metalloproteinase-1 (MT1-MMP) and Tissue Inhibitor of Metalloproteinases-2 (TIMP-2) Expression by Elastin-Derived Peptides in Human HT-1080 Fibrosarcoma Cell Line. Clin. Exp. Metastasis 1998, 16, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Pati, F.; Jang, J.; Ha, D.-H.; Won Kim, S.; Rhie, J.-W.; Shim, J.-H.; Kim, D.-H.; Cho, D.-W. Printing Three-Dimensional Tissue Analogues with Decellularized Extracellular Matrix Bioink. Nat. Commun. 2014, 5, 3935. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Singh, N.K.; Kim, J.J.; Kim, H.; Kim, B.S.; Park, J.Y.; Jang, J.; Cho, D.-W. Directed Differential Behaviours of Multipotent Adult Stem Cells from Decellularized Tissue/Organ Extracellular Matrix Bioinks. Biomaterials 2019, 224, 119496. [Google Scholar] [CrossRef]
- Mao, Y.; Hoffman, T.; Wu, A.; Goyal, R.; Kohn, J. Cell Type–Specific Extracellular Matrix Guided the Differentiation of Human Mesenchymal Stem Cells in 3D Polymeric Scaffolds. J. Mater. Sci. Mater. Med. 2017, 28, 100. [Google Scholar] [CrossRef]
- Noh, Y.K.; Du, P.; Dos Santos Da Costa, A.; Park, K. Induction of Chondrogenesis of Human Placenta-Derived Mesenchymal Stem Cells via Heparin-Grafted Human Fibroblast Derived Matrix. Biomater. Res. 2018, 22, 12. [Google Scholar] [CrossRef] [PubMed]
- Rothrauff, B.B.; Yang, G.; Tuan, R.S. Tissue-Specific Bioactivity of Soluble Tendon-Derived and Cartilage-Derived Extracellular Matrices on Adult Mesenchymal Stem Cells. Stem. Cell Res. Ther. 2017, 8, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Q.-W.; Zhang, Y.; Luo, J.-C.; Zhang, D.; Xiong, B.-J.; Yang, J.-Q.; Xie, H.-Q.; Lv, Q. Hydrogel Derived from Decellularized Porcine Adipose Tissue as a Promising Biomaterial for Soft Tissue Augmentation. J. Biomed. Mater. Res. A 2017, 105, 1756–1764. [Google Scholar] [CrossRef]
- Lin, X.; Fang, X.; Wang, Q.; Hu, Z.; Chen, K.; Shan, Z.; Chen, S.; Wang, J.; Mo, J.; Ma, J.; et al. Decellularized Allogeneic Intervertebral Disc: Natural Biomaterials for Regenerating Disc Degeneration. Oncotarget 2016, 7, 12121–12136. [Google Scholar] [CrossRef] [Green Version]
- Shao, X.; Johnson, J.E.; Richardson, J.A.; Hiesberger, T.; Igarashi, P.; Adin, C.; Ellison, G.W.; Jorgensen, M.; Batich, C.D. A Minimal Ksp-Cadherin Promoter Linked to a Green Fluorescent Protein Reporter Gene Exhibits Tissue-Specific Expression in the Developing Kidney and Genitourinary Tract. J. Am. Soc. Nephrol. 2009, 13, 1824–1836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simões, I.N.; Vale, P.; Soker, S.; Atala, A.; Keller, D.; Noiva, R.; Carvalho, S.; Peleteiro, C.; Cabral, J.M.S.; Eberli, D.; et al. Acellular Urethra Bioscaffold: Decellularization of Whole Urethras for Tissue Engineering Applications. Sci. Rep. 2017, 7, 41934. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Pei, F.; Zhang, W.; Guo, W.; Li, R.; He, W.; Tian, W. Bone Marrow Mesenchymal Stem Cells Combine with Treated Dentin Matrix to Build Biological Root. Sci. Rep. 2017, 7, 44635. [Google Scholar] [CrossRef] [Green Version]
- Dziki, J.L.; Giglio, R.M.; Sicari, B.M.; Wang, D.S.; Gandhi, R.M.; Londono, R.; Dearth, C.L.; Badylak, S.F. The Effect of Mechanical Loading Upon Extracellular Matrix Bioscaffold-Mediated Skeletal Muscle Remodelling. Tissue Eng. Part A 2018, 24, 34–46. [Google Scholar] [CrossRef]
- Gao, L.-P.; Du, M.-J.; Lv, J.-J.; Schmull, S.; Huang, R.-T.; Li, J. Use of Human Aortic Extracellular Matrix as a Scaffold for Construction of a Patient-Specific Tissue Engineered Vascular Patch. Biomed. Mater. 2017, 12, 065006. [Google Scholar] [CrossRef]
- Smith, L.R.; Cho, S.; Discher, D.E. Stem Cell Differentiation Is Regulated by Extracellular Matrix Mechanics. Physiology 2018, 33, 16–25. [Google Scholar] [CrossRef]
- Watt, F.M.; Huck, W.T.S. Role of the Extracellular Matrix in Regulating Stem Cell Fate. Nat. Rev. Mol. Cell Biol. 2013, 14, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Willemse, J.; van Tienderen, G.; van Hengel, E.; Schurink, I.; van der Ven, D.; Kan, Y.; de Ruiter, P.; Rosmark, O.; Westergren-Thorsson, G.G.; Schneeberger, K.; et al. Hydrogels Derived from Decellularized Liver Tissue Support the Growth and Differentiation of Cholangiocyte Organoids. Biomaterials 2022, 284, 121473. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Zheng, J.; Li, Z.; Qi, X.; Tian, Y.; Zhang, M.; Zhang, J.; Huang, X. Printing 3D Vagina Tissue Analogues with Vagina Decellularized Extracellular Matrix Bioink. Int. J. Biol. Macromol. 2021, 180, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Nam, S.A.; Yi, J.; Kim, J.Y.; Lee, J.Y.; Park, S.; Sen, T.; Choi, Y.; Lee, J.Y.; Kim, H.L.; et al. Kidney Decellularized Extracellular Matrix Enhanced the Vascularization and Maturation of Human Kidney Organoids. Adv. Sci. 2022, 9, 2103526. [Google Scholar] [CrossRef]
- Goh, S.K.; Halfter, W.; Richardson, T.; Bertera, S.; Vaidya, V.; Candiello, J.; Bradford, M.; Banerjee, I. Organ-Specific ECM Arrays for Investigating Cell-ECM Interactions during Stem Cell Differentiation. Biofabrication 2020, 13, 015015. [Google Scholar] [CrossRef]
- Gao, L.; Li, X.; Tan, R.; Cui, J.; Schmull, S. Human-Derived Decellularized Extracellular Matrix Scaffold Incorporating Autologous Bone Marrow Stem Cells from Patients with Congenital Heart Disease for Cardiac Tissue Engineering. Biomed. Mater. Eng. 2022, Preprint, 1–15. [Google Scholar] [CrossRef]
- Wan, H.Y.; Shin, R.L.Y.; Chen, J.C.H.; Assunção, M.; Wang, D.; Nilsson, S.K.; Tuan, R.S.; Blocki, A. Dextran Sulfate-Amplified Extracellular Matrix Deposition Promotes Osteogenic Differentiation of Mesenchymal Stem Cells. Acta Biomater. 2022, 140, 163–177. [Google Scholar] [CrossRef]
- Mesquita, F.C.P.; Morrissey, J.; Monnerat, G.; Domont, G.B.; Nogueira, F.C.S.; Hochman-Mendez, C. Decellularized Extracellular Matrix Powder Accelerates Metabolic Maturation at Early Stages of Cardiac Differentiation in Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Cells Tissues Organs 2021, 1–13. [Google Scholar] [CrossRef]
- Weng, W.; Zanetti, F.; Bovard, D.; Braun, B.; Ehnert, S.; Uynuk-Ool, T.; Histing, T.; Hoeng, J.; Nussler, A.K.; Aspera-Werz, R.H. A Simple Method for Decellularizing a Cell-Derived Matrix for Bone Cell Cultivation and Differentiation. J. Mater. Sci. Mater. Med. 2021, 32, 124. [Google Scholar] [CrossRef]
- Carvalho, C.M.F.; Leonel, L.C.P.C.; Cañada, R.R.; Barreto, R.S.N.; Maria, D.A.; Del Sol, M.; Miglino, M.A.; Lobo, S.E. Comparison between Placental and Skeletal Muscle ECM: In Vivo Implantation. Connect. Tissue Res. 2021, 62, 629–642. [Google Scholar] [CrossRef]
- Ozudogru, E.; Isik, M.; Eylem, C.C.; Nemutlu, E.; Arslan, Y.E.; Derkus, B. Decellularized Spinal Cord Meninges Extracellular Matrix Hydrogel That Supports Neurogenic Differentiation and Vascular Structure Formation. J. Tissue Eng. Regen Med 2021, 15, 948–963. [Google Scholar] [CrossRef] [PubMed]
- Sasikumar, S.; Chameettachal, S.; Kingshott, P.; Cromer, B.; Pati, F. Influence of Liver Extracellular Matrix in Predicting Drug-Induced Liver Injury: An Alternate Paradigm. ACS Biomater. Sci. Eng. 2022, 8, 834–846. [Google Scholar] [CrossRef] [PubMed]
- Girardeau-Hubert, S.; Lynch, B.; Zuttion, F.; Label, R.; Rayee, C.; Brizion, S.; Ricois, S.; Martinez, A.; Park, E.; Kim, C.; et al. Impact of Microstructure on Cell Behaviour and Tissue Mechanics in Collagen and Dermal Decellularized Extra-Cellular Matrices. Acta Biomater. 2022, 143, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, W.; Dai, Y.; Huang, Y.; Chen, Z.; Xi, T.; Zhou, Z.; Liu, H. Decellularized Sturgeon Cartilage Extracellular Matrix Scaffold Inhibits Chondrocyte Hypertrophy in Vitro and in Vivo. J. Tissue Eng. Regen. Med. 2021, 15, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Ryzhuk, V.; Zeng, X.; Wang, X.; Melnychuk, V.; Lankford, L.; Farmer, D.; Wang, A. Human Amnion Extracellular Matrix Derived Bioactive Hydrogel for Cell Delivery and Tissue Engineering. Mater. Sci. Eng. C 2018, 85, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili Pourfarhangi, K.; Mashayekhan, S.; Asl, S.G.; Hajebrahimi, Z. Construction of Scaffolds Composed of Acellular Cardiac Extracellular Matrix for Myocardial Tissue Engineering. Biologicals 2018, 53, 10–18. [Google Scholar] [CrossRef]
- Becker, M.; Maring, J.; Schneider, M.; Herrera Martin, A.; Seifert, M.; Klein, O.; Braun, T.; Falk, V.; Stamm, C.; Becker, M.; et al. Towards a Novel Patch Material for Cardiac Applications: Tissue-Specific Extracellular Matrix Introduces Essential Key Features to Decellularized Amniotic Membrane. Int. J. Mol. Sci. 2018, 19, 1032. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Liu, Y.; Hui, L. Preparation and Characterization of Acellular Adipose Tissue Matrix Using a Combination of Physical and Chemical Treatments. Mol. Med. Rep. 2017, 17, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Fischer, I.; Westphal, M.; Rossbach, B.; Bethke, N.; Hariharan, K.; Ullah, I.; Reinke, P.; Kurtz, A.; Stachelscheid, H. Comparative Characterization of Decellularized Renal Scaffolds for Tissue Engineering. Biomed. Mater. 2017, 12, 045005. [Google Scholar] [CrossRef]
- Kim, J.; Shim, I.K.; Hwang, D.G.; Lee, Y.N.; Kim, M.; Kim, H.; Kim, S.W.; Lee, S.; Kim, S.C.; Cho, D.W.; et al. 3D Cell Printing of Islet-Laden Pancreatic Tissue-Derived Extracellular Matrix Bioink Constructs for Enhancing Pancreatic Functions. J. Mater. Chem. B 2019, 7, 1773–1781. [Google Scholar] [CrossRef]
- Xu, J.; Fang, H.; Zheng, S.; Li, L.; Jiao, Z.; Wang, H.; Nie, Y.; Liu, T.; Song, K. A Biological Functional Hybrid Scaffold Based on Decellularized Extracellular Matrix/Gelatin/Chitosan with High Biocompatibility and Antibacterial Activity for Skin Tissue Engineering. Int. J. Biol. Macromol. 2021, 187, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.C.; Rodrigues, S.C.; Caldeira, J.; Nunes, A.M.; Sampaio-Pinto, V.; Resende, T.P.; Oliveira, M.J.J.; Barbosa, M.A.M.A.; Thorsteinsdóttir, S.; Nascimento, D.S.A.; et al. Three-Dimensional Scaffolds of Fetal Decellularized Hearts Exhibit Enhanced Potential to Support Cardiac Cells in Comparison to the Adult. Biomaterials 2016, 104, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Gilpin, S.E.; Li, Q.; Evangelista-Leite, D.; Ren, X.; Reinhardt, D.P.; Frey, B.L.; Ott, H.C. Fibrillin-2 and Tenascin-C Bridge the Age Gap in Lung Epithelial Regeneration. Biomaterials 2017, 140, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Wu Young, M.Y.; Dolivo, D.M.; Hong, S.J.; Iyer, H.; Mustoe, T.A.; Galiano, R.D. Decellularized Fetal Matrix Suppresses Fibrotic Gene Expression and Promotes Myogenesis in a Rat Model of Volumetric Muscle Loss. Plast. Reconstr. Surg. 2020, 146, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Sicari, B.M.; Johnson, S.A.; Siu, B.F.; Crapo, P.M.; Daly, K.A.; Jiang, H.; Medberry, C.J.; Tottey, S.; Turner, N.J.; Badylak, S.F. The Effect of Source Animal Age upon the in Vivo Remodelling Characteristics of an Extracellular Matrix Scaffold. Biomaterials 2012, 33, 5524–5533. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Long, D.W.; Huang, Y.; Chen, W.C.W.; Kim, K.; Wang, Y. Decellularized Neonatal Cardiac Extracellular Matrix Prevents Widespread Ventricular Remodelling in Adult Mammals after Myocardial Infarction. Acta Biomater. 2019, 87, 140–151. [Google Scholar] [CrossRef]
- LoPresti, S.T.; Brown, B.N. Effect of Source Animal Age upon Macrophage Response to Extracellular Matrix Biomaterials. J. Immunol. Regen. Med. 2018, 1, 57–66. [Google Scholar] [CrossRef]
- Wang, X.; Pierre, V.; Liu, C.; Senapati, S.; Park, P.S.H.; Senyo, S.E. Exogenous Extracellular Matrix Proteins Decrease Cardiac Fibroblast Activation in Stiffening Microenvironment through CAPG. J. Mol. Cell Cardiol. 2021, 159, 105–119. [Google Scholar] [CrossRef]
- Balestrini, J.L.; Gard, A.L.; Gerhold, K.A.; Wilcox, E.C.; Liu, A.; Schwan, J.; Le, A.V.; Baevova, P.; Dimitrievska, S.; Zhao, L.; et al. Comparative Biology of Decellularized Lung Matrix: Implications of Species Mismatch in Regenerative Medicine. Biomaterials 2016, 102, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef]
- Mollica, P.A.; Booth-Creech, E.N.; Reid, J.A.; Zamponi, M.; Sullivan, S.M.; Palmer, X.-L.; Sachs, P.C.; Bruno, R.D. 3D Bioprinted Mammary Organoids and Tumouroids in Human Mammary Derived ECM Hydrogels. Acta Biomater. 2019, 95, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Helal-Neto, E.; Brandão-Costa, R.M.; Saldanha-Gama, R.; Ribeiro-Pereira, C.; Midlej, V.; Benchimol, M.; Morandi, V.; Barja-Fidalgo, C. Priming Endothelial Cells with a Melanoma-Derived Extracellular Matrix Triggers the Activation of Avβ3/VEGFR2 Axis. J. Cell Physiol. 2016, 231, 2464–2473. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wang, B.; Li, S.; Jin, Q.; Dai, Y. Human Breast Cancer Decellularized Scaffolds Promote Epithelial-to-mesenchymal Transitions and Stemness of Breast Cancer Cells in Vitro. J. Cell Physiol. 2019, 234, 9447–9456. [Google Scholar] [CrossRef]
- Pinto, M.L.; Rios, E.S.; Silva, A.C.; Neves, S.C.; Caires, H.R.; Pinto, A.T.; Durães, C.; Carvalho, F.A.; Cardoso, A.P.; Santos, N.C.; et al. Decellularized Human Colorectal Cancer Matrices Polarize Macrophages towards an Anti-Inflammatory Phenotype Promoting Cancer Cell Invasion via CCL18. Biomaterials 2017, 124, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Liu, G.; Li, S.; Yuan, H.; Yun, Z.; Zhang, W.; Zhang, S.; Dai, Y.; Ma, Y. Decellularized Breast Matrix as Bioactive Microenvironment for in Vitro Three-Dimensional Cancer Culture. J. Cell Physiol. 2019, 234, 3425–3435. [Google Scholar] [CrossRef]
- Mazza, G.; Telese, A.; Al-Akkad, W.; Frenguelli, L.; Levi, A.; Marrali, M.; Longato, L.; Thanapirom, K.; Vilia, M.G.; Lombardi, B.; et al. Cirrhotic Human Liver Extracellular Matrix 3D Scaffolds Promote Smad-Dependent TGF-Β1 Epithelial Mesenchymal Transition. Cells 2020, 9, 83. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Xu, J.; Huang, Y.; Tang, L.; Hong, Y. Regional-Specific Meniscal Extracellular Matrix Hydrogels and Their Effects on Cell-Matrix Interactions of Fibrochondrocytes. Biomed. Mater. 2022, 17, 014105. [Google Scholar] [CrossRef]
- Ning, L.-J.; Jiang, Y.-L.; Zhang, C.-H.; Zhang, Y.; Yang, J.-L.; Cui, J.; Zhang, Y.-J.; Yao, X.; Luo, J.-C.; Qin, T.-W. Fabrication and Characterization of a Decellularized Bovine Tendon Sheet for Tendon Reconstruction. J. Biomed. Mater. Res. A 2017, 105, 2299–2311. [Google Scholar] [CrossRef]
- Xu, S.; Lu, F.; Cheng, L.; Li, C.; Zhou, X.; Wu, Y.; Chen, H.; Zhang, K.; Wang, L.; Xia, J.; et al. Preparation and Characterization of Small-Diameter Decellularized Scaffolds for Vascular Tissue Engineering in an Animal Model. Biomed. Eng. Online 2017, 16, 55. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Wang, S.; Wang, G.; Su, M.; Song, L.; Chen, J.; Fan, S.; Lin, X. Preparation of Decellularized Biphasic Hierarchical Myotendinous Junction Extracellular Matrix for Muscle Regeneration. Acta Biomater. 2018, 68, 15–28. [Google Scholar] [CrossRef]
- Rajabi, S.; Pahlavan, S.; Ashtiani, M.K.; Ansari, H.; Abbasalizadeh, S.; Sayahpour, F.A.; Varzideh, F.; Kostin, S.; Aghdami, N.; Braun, T.; et al. Human Embryonic Stem Cell-Derived Cardiovascular Progenitor Cells Efficiently Colonize in BFGF-Tethered Natural Matrix to Construct Contracting Humanized Rat Hearts. Biomaterials 2018, 154, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Park, H.-J.; Kim, S.-W.; Kim, H.; Park, J.Y.; Na, S.J.; Kim, H.J.; Park, M.N.; Choi, S.H.; Park, S.H.; et al. 3D Printed Complex Tissue Construct Using Stem Cell-Laden Decellularized Extracellular Matrix Bioinks for Cardiac Repair. Biomaterials 2017, 112, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, H.; Bai, A.; Jiang, W.; Li, X.; Wang, X.; Mao, Y.; Lu, C.; Qian, R.; Guo, F.; et al. Functional Engineered Human Cardiac Patches Prepared from Nature’s Platform Improve Heart Function after Acute Myocardial Infarction. Biomaterials 2016, 105, 52–65. [Google Scholar] [CrossRef] [PubMed]
- D’Amore, A.; Yoshizumi, T.; Luketich, S.K.; Wolf, M.T.; Gu, X.; Cammarata, M.; Hoff, R.; Badylak, S.F.; Wagner, W.R. Bi-Layered Polyurethane—Extracellular Matrix Cardiac Patch Improves Ischemic Ventricular Wall Remodelling in a Rat Model. Biomaterials 2016, 107, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; PR, A.K.; Yoo, J.J.; Zahran, F.; Atala, A.; Lee, S.J. A Photo-Crosslinkable Kidney ECM-Derived Bioink Accelerates Renal Tissue Formation. Adv. Healthc. Mater. 2019, 8, 1800992. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Butler, J.P.; Ingber, D.E. Mechanotransduction across the Cell Surface and through the Cytoskeleton. Science 1993, 260, 1124–1127. [Google Scholar] [CrossRef]
- Fraldi, M.; Palumbo, S.; Carotenuto, A.R.; Cutolo, A.; Deseri, L.; Pugno, N. Buckling Soft Tensegrities: Fickle Elasticity and Configurational Switching in Living Cells. J. Mech. Phys. Solids 2019, 124, 299–324. [Google Scholar] [CrossRef] [Green Version]
- Ingber, D.E. Control of Capillary Growth and Differentiation by Extracellular Matrix: Use of a Tensegrity (Tensional Integrity) Mechanism for Signal Processing. Chest 1991, 99, 34S–40S. [Google Scholar] [CrossRef]
- Allbritton-King, J.D.; Kimicata, M.; Fisher, J.P. Incorporating a Structural Extracellular Matrix Gradient into a Porcine Urinary Bladder Matrix-based Hydrogel Dermal Scaffold. J. Biomed. Mater. Res. A 2021, 109, 1893–1904. [Google Scholar] [CrossRef]
- Schwartz, D.M.; Pehlivaner Kara, M.O.; Goldstein, A.M.; Ott, H.C.; Ekenseair, A.K. Spray Delivery of Intestinal Organoids to Reconstitute Epithelium on Decellularized Native Extracellular Matrix. Tissue Eng. Part C Methods 2017, 23, 565–573. [Google Scholar] [CrossRef]
- Jank, B.J.; Xiong, L.; Moser, P.T.; Guyette, J.P.; Ren, X.; Cetrulo, C.L.; Leonard, D.A.; Fernandez, L.; Fagan, S.P.; Ott, H.C. Engineered Composite Tissue as a Bioartificial Limb Graft. Biomaterials 2015, 61, 246–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, E.E.; Lanier, S.T.; Niknam-Bienia, S.; Arenas, G.A.; Rajendran, D.; Wertheim, J.A.; Galiano, R.D. Residual Sodium Dodecyl Sulfate in Decellularized Muscle Matrices Leads to Fibroblast Activation in Vitro and Foreign Body Response in Vivo. J. Tissue Eng. Regen. Med. 2018, 12, e1704–e1715. [Google Scholar] [CrossRef]
- Ghorbani, F.; Ekhtiari, M.; Moeini Chaghervand, B.; Moradi, L.; Mohammadi, B.; Kajbafzadeh, A.M. Detection of the Residual Concentration of Sodium Dodecyl Sulfate in the Decellularized Whole Rabbit Kidney Extracellular Matrix. Cell Tissue Bank. 2022, 23, 119–128. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Callanan, A.; Lagaras, K.; Steele, J.A.M.; Stevens, M.M. Optimization of SDS Exposure on Preservation of ECM Characteristics in Whole Organ Decellularization of Rat Kidneys. J. Biomed. Mater. Res. B Appl. Biomater. 2017, 105, 1352–1360. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Kirita, Y.; Kami, D.; Kitani, T.; Ozaki, C.; Itakura, Y.; Toyoda, M.; Gojo, S. Novel Detergent for Whole Organ Tissue Engineering. J. Biomed. Mater. Res. A 2015, 103, 3364–3373. [Google Scholar] [CrossRef] [PubMed]
- Uhl, F.E.; Zhang, F.; Pouliot, R.A.; Uriarte, J.J.; Rolandsson Enes, S.; Han, X.; Ouyang, Y.; Xia, K.; Westergren-Thorsson, G.; Malmström, A.; et al. Functional Role of Glycosaminoglycans in Decellularized Lung Extracellular Matrix. Acta Biomater. 2020, 102, 231–246. [Google Scholar] [CrossRef]
- Liu, X.; Li, N.; Gong, D.; Xia, C.; Xu, Z. Comparison of Detergent-Based Decellularization Protocols for the Removal of Antigenic Cellular Components in Porcine Aortic Valve. Xenotransplantation 2018, 25, e12380. [Google Scholar] [CrossRef]
- Robertson, M.J.; Dries-Devlin, J.L.; Kren, S.M.; Burchfield, J.S.; Taylor, D.A. Optimizing Recellularization of Whole Decellularized Heart Extracellular Matrix. PLoS ONE 2014, 9, e90406. [Google Scholar] [CrossRef] [Green Version]
- Campo, H.; Baptista, P.M.; López-Pérez, N.; Faus, A.; Cervelló, I.; Simón, C. De- and Recellularization of the Pig Uterus: A Bioengineering Pilot Study. Biol. Reprod. 2016, 96, 34–45. [Google Scholar] [CrossRef]
- Chirco, K.R.; Worthington, K.S.; Flamme-Wiese, M.J.; Riker, M.J.; Andrade, J.D.; Ueberheide, B.M.; Stone, E.M.; Tucker, B.A.; Mullins, R.F. Preparation and Evaluation of Human Choroid Extracellular Matrix Scaffolds for the Study of Cell Replacement Strategies. Acta Biomater. 2017, 57, 293–303. [Google Scholar] [CrossRef]
- Lohan, A.; Kohl, B.; Meier, C.; Schulze-Tanzil, G. Tenogenesis of Decellularized Porcine Achilles Tendon Matrix Reseeded with Human Tenocytes in the Nude Mice Xenograft Model. Int. J. Mol. Sci. 2018, 19, 2059. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Gao, Z.; Xu, J.; Zhu, Z.; Fan, Z.; Zhang, C.; Wang, J.; Wang, S. Decellularized Swine Dental Pulp as a Bioscaffold for Pulp Regeneration. Biomed. Res. Int. 2017, 2017, 9342714. [Google Scholar] [CrossRef] [PubMed]
- Kajbafzadeh, A.-M.; Abbasioun, R.; Sabetkish, N.; Sabetkish, S.; Habibi, A.A.; Tavakkolitabassi, K. In Vivo Human Corpus Cavernosum Regeneration: Fabrication of Tissue-Engineered Corpus Cavernosum in Rat Using the Body as a Natural Bioreactor. Int. Urol. Nephrol. 2017, 49, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.K.; Du, R.X.; Zhang, L.; Li, Y.N.; Zhang, M.L.; Zhao, S.; Huang, X.H.; Xu, Y.F. A New Material for Tissue Engineered Vagina Reconstruction: Acellular Porcine Vagina Matrix. J. Biomed. Mater. Res. A 2017, 105, 1949–1959. [Google Scholar] [CrossRef]
- Das, S.; Kim, S.-W.; Choi, Y.-J.; Lee, S.; Lee, S.-H.; Kong, J.-S.; Park, H.-J.; Cho, D.-W.; Jang, J. Decellularized Extracellular Matrix Bioinks and the External Stimuli to Enhance Cardiac Tissue Development in Vitro. Acta Biomater. 2019, 95, 188–200. [Google Scholar] [CrossRef]
- Gao, G.; Lee, J.H.; Jang, J.; Lee, D.H.; Kong, J.-S.; Kim, B.S.; Choi, Y.-J.; Jang, W.B.; Hong, Y.J.; Kwon, S.-M.; et al. Tissue Engineered Bio-Blood-Vessels Constructed Using a Tissue-Specific Bioink and 3D Coaxial Cell Printing Technique: A Novel Therapy for Ischemic Disease. Adv. Funct. Mater. 2017, 27, 1700798. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.; Kim, T.G.; Kim, B.S.; Kim, S.-W.; Kwon, S.-M.; Cho, D.-W. Tailoring Mechanical Properties of Decellularized Extracellular Matrix Bioink by Vitamin B2-Induced Photo-Crosslinking. Acta Biomater. 2016, 33, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Han, W.; Kim, H.; Ha, D.-H.; Jang, J.; Kim, B.S.; Cho, D.-W. Development of Liver Decellularized Extracellular Matrix Bioink for Three-Dimensional Cell Printing-Based Liver Tissue Engineering. Biomacromolecules 2017, 18, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Ijima, H.; Nakamura, S.; Bual, R.; Shirakigawa, N.; Tanoue, S. Physical Properties of the Extracellular Matrix of Decellularized Porcine Liver. Gels 2018, 4, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remuzzi, A.; Figliuzzi, M.; Bonandrini, B.; Silvani, S.; Azzollini, N.; Nossa, R.; Benigni, A.; Remuzzi, G. Experimental Evaluation of Kidney Regeneration by Organ Scaffold Recellularization. Sci. Rep. 2017, 7, 43502. [Google Scholar] [CrossRef]
- Park, S.M.; Yang, S.; Rye, S.-M.; Choi, S.W. Effect of Pulsatile Flow Perfusion on Decellularization. Biomed. Eng. Online 2018, 17, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guler, S.; Aydin, H.M.; Lü, L.-X.; Yang, Y. Improvement of Decellularization Efficiency of Porcine Aorta Using Dimethyl Sulfoxide as a Penetration Enhancer. Artif. Organs 2018, 42, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Z.; Wong, M.L.; Griffiths, L.G. Effect of Bovine Pericardial Extracellular Matrix Scaffold Niche on Seeded Human Mesenchymal Stem Cell Function. Sci. Rep. 2016, 6, 37089. [Google Scholar] [CrossRef] [Green Version]
- Grover, G.N.; Rao, N.; Christman, K.L. Myocardial Matrix-Polyethylene Glycol Hybrid Hydrogels for Tissue Engineering. Nanotechnology 2014, 25, 12. [Google Scholar] [CrossRef]
- Xiang, J.; Liu, P.; Zheng, X.; Dong, D.; Fan, S.; Dong, J.; Zhang, X.; Liu, X.; Wang, B.; Lv, Y. The Effect of Riboflavin/UVA Cross-Linking on Anti-Degeneration and Promoting Angiogenic Capability of Decellularized Liver Matrix. J. Biomed. Mater. Res. A 2017, 105, 2662–2669. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, M.; Jundziłł, A.; Nazarewski, Ł.; Kotela, A.; Kloskowski, T.; Skopińska-Wisniewska, J.; Bodnar, M.; Łukasiewicz, A.; Nazarewski, S.; Kotela, I.; et al. Blood Vessel Matrix Seeded with Cells: A Better Alternative for Abdominal Wall Reconstruction—A Long-Term Study. Biomed. Res. Int. 2015, 2015, 890613. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.L.; Yan, M.; Su, J.; Shah, R.N. Directing the Growth and Alignment of Biliary Epithelium within Extracellular Matrix Hydrogels. Acta Biomater. 2019, 85, 84–93. [Google Scholar] [CrossRef]
- Kim, S.; Min, S.; Choi, Y.S.; Jo, S.-H.; Jung, J.H.; Han, K.; Kim, J.; An, S.; Ji, Y.W.; Kim, Y.-G.; et al. Tissue Extracellular Matrix Hydrogels as Alternatives to Matrigel for Culturing Gastrointestinal Organoids. Nat. Commun. 2022, 13, 1692. [Google Scholar] [CrossRef]
- Ergun, C.; Parmaksiz, M.; Vurat, M.T.; Elçin, A.E.; Elçin, Y.M. Decellularized Liver ECM-Based 3D Scaffolds: Compositional, Physical, Chemical, Rheological, Thermal, Mechanical, and in Vitro Biological Evaluations. Int. J. Biol. Macromol. 2022, 200, 110–123. [Google Scholar] [CrossRef]
- Fu, Z.; Naghieh, S.; Xu, C.; Wang, C.; Sun, W.; Chen, X. Printability in Extrusion Bioprinting. Biofabrication 2021, 13, 033001. [Google Scholar] [CrossRef]
- Barthold, J.E.; McCreery, K.P.; Martinez, J.; Bellerjeau, C.; Ding, Y.; Bryant, S.J.; Whiting, G.L.; Neu, C.P. Particulate ECM Biomaterial Ink Is 3D Printed and Naturally Crosslinked to Form Structurally-Layered and Lubricated Cartilage Tissue Mimics. Biofabrication 2022, 14, 025021. [Google Scholar] [CrossRef] [PubMed]
- Fares, A.E.; Gabr, H.; ShamsEldeen, A.M.; Farghali, H.A.M.; Rizk, M.M.S.M.; Mahmoud, B.E.; Tammam, A.B.A.; Mahmoud, A.M.A.; Suliman, A.A.M.; Ayyad, M.A.A.; et al. Implanted Subcutaneous versus Intraperitoneal Bioscaffold Seeded with Hepatocyte-like Cells: Functional Evaluation. Stem. Cell Res. Ther. 2021, 12, 441. [Google Scholar] [CrossRef] [PubMed]
- Demko, P.; Hillebrandt, K.H.; Napierala, H.; Haep, N.; Tang, P.; Gassner, J.M.G.V.; Kluge, M.; Everwien, H.; Polenz, D.; Reutzel-Selke, A.; et al. Perfusion-Based Recellularization of Rat Livers with Islets of Langerhans. J. Med. Biol. Eng. 2022, 42, 271–280. [Google Scholar] [CrossRef]
- Hou, N.; Lv, D.; Xu, X.; Lu, Y.; Li, J.; Ma, R.; Tang, Y.; Zheng, Y. Development of a Decellularized Hypopharynx with Vascular Pedicle Scaffold for Use in Reconstructing Hypopharynx. Artif. Organs 2022, 46, 1268–1280. [Google Scholar] [CrossRef] [PubMed]
- Greco, K.V.; Jones, L.G.; Obiri-Yeboa, I.; Ansari, T. Creation of an Acellular Vaginal Matrix for Potential Vaginal Augmentation and Cloacal Repair. J. Pediatr. Adolesc. Gynecol. 2018, 31, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Butler, C.R.; Hynds, R.E.; Crowley, C.; Gowers, K.H.C.; Partington, L.; Hamilton, N.J.; Carvalho, C.; Platé, M.; Samuel, E.R.; Burns, A.J.; et al. Vacuum-Assisted Decellularization: An Accelerated Protocol to Generate Tissue-Engineered Human Tracheal Scaffolds. Biomaterials 2017, 124, 95–105. [Google Scholar] [CrossRef]
- Yang, J.-L.; Yao, X.; Qing, Q.; Zhang, Y.; Jiang, Y.-L.; Ning, L.-J.; Luo, J.-C.; Qin, T.-W. An Engineered Tendon/Ligament Bioscaffold Derived from Decellularized and Demineralized Cortical Bone Matrix. J. Biomed. Mater. Res. A 2018, 106, 468–478. [Google Scholar] [CrossRef]
- Sun, D.; Liu, Y.; Wang, H.; Deng, F.; Zhang, Y.; Zhao, S.; Ma, X.; Wu, H.; Sun, G. Novel Decellularized Liver Matrix-Alginate Hybrid Gel Beads for the 3D Culture of Hepatocellular Carcinoma Cells. Int. J. Biol. Macromol. 2018, 109, 1154–1163. [Google Scholar] [CrossRef]
- Lange, P.; Shah, H.; Birchall, M.A.; Sibbons, P.D.; Ansari, T. Characterization of a Biologically Derived Rabbit Tracheal Scaffold. J. Biomed. Mater. Res. B Appl. Biomater. 2017, 105, 2126–2135. [Google Scholar] [CrossRef]
- Den Hondt, M.; Vanaudenaerde, B.M.; Maughan, E.F.; Butler, C.R.; Crowley, C.; Verbeken, E.K.; Verleden, S.E.; Vranckx, J.J. An Optimized Non-Destructive Protocol for Testing Mechanical Properties in Decellularized Rabbit Trachea. Acta Biomater. 2017, 60, 291–301. [Google Scholar] [CrossRef]
- Garriboli, M.; Deguchi, K.; Totonelli, G.; Georgiades, F.; Urbani, L.; Ghionzoli, M.; Burns, A.J.; Sebire, N.J.; Turmaine, M.; Eaton, S.; et al. Development of a Porcine Acellular Bladder Matrix for Tissue-Engineered Bladder Reconstruction. Pediatr. Surg. Int. 2022, 38, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Chiti, M.; Vanacker, J.; Ouni, E.; Tatic, N.; Viswanath, A.; des Rieux, A.; Dolmans, M.; White, L.J.; Amorim, C.A. Ovarian Extracellular Matrix-based Hydrogel for Human Ovarian Follicle Survival in Vivo: A Pilot Work. J. Biomed. Mater. Res. B Appl. Biomater. 2022, 110, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Meder, T.J.; Prest, T.A.; Skillen, C.; Marchal, L.; Yupanqui, V.T.; Soletti, L.; Gardner, P.; Cheetham, J.; Brown, B.N. Nerve-Specific Extracellular Matrix Hydrogel Promotes Functional Regeneration Following Nerve Gap Injury. NPJ Regen. Med. 2021, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- Naeem, E.M.; Sajad, D.; Talaei-Khozani, T.; Khajeh, S.; Azarpira, N.; Alaei, S.; Tanideh, N.; Reza, T.M.; Razban, V. Decellularized Liver Transplant Could Be Recellularized in Rat Partial Hepatectomy Model. J. Biomed. Mater. Res. A 2019, 107, 2576–2588. [Google Scholar] [CrossRef]
- Yaghoubi, A.; Azarpira, N.; Karbalay-Doust, S.; Daneshi, S.; Vojdani, Z.; Talaei-Khozani, T. Prednisolone and Mesenchymal Stem Cell Preloading Protect Liver Cell Migration and Mitigate Extracellular Matrix Modification in Transplanted Decellularized Rat Liver. Stem. Cell Res. Ther. 2022, 13, 36. [Google Scholar] [CrossRef]
- Obata, T.; Tsuchiya, T.; Akita, S.; Kawahara, T.; Matsumoto, K.; Miyazaki, T.; Masumoto, H.; Kobayashi, E.; Niklason, L.E.; Nagayasu, T. Utilization of Natural Detergent Potassium Laurate for Decellularization in Lung Bioengineering. Tissue Eng. Part C Methods 2019, 25, 459–471. [Google Scholar] [CrossRef]
- Hudson, T.W.; Zawko, S.; Deister, C.; Lundy, S.; Hu, C.Y.; Lee, K.; Schmidt, C.E. Optimized Acellular Nerve Graft Is Immunologically Tolerated and Supports Regeneration. Tissue Eng. 2004, 10, 1641–1651. [Google Scholar] [CrossRef]
- Nagao, R.J.; Ouyang, Y.; Keller, R.; Lee, C.; Suggs, L.J.; Schmidt, C.E. Preservation of Capillary-Beds in Rat Lung Tissue Using Optimized Chemical Decellularization. J. Mater. Chem. B 2013, 1, 4801–4808. [Google Scholar] [CrossRef] [Green Version]
- Uygun, B.E.; Price, G.; Saedi, N.; Izamis, M.-L.; Berendsen, T.; Yarmush, M.; Uygun, K. Decellularization and Recellularization of Whole Livers. J. Vis. Exp. 2011, 48, e2394. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Xu, R.; Huang, J.; Bao, X.; Zhao, B. Treatment of Rabbit Growth Plate Injuries with Oriented ECM Scaffold and Autologous BMSCs. Sci. Rep. 2017, 7, 44140. [Google Scholar] [CrossRef]
- White, L.J.; Keane, T.J.; Smoulder, A.; Zhang, L.; Castleton, A.A.; Reing, J.E.; Turner, N.J.; Dearth, C.L.; Badylak, S.F. The Impact of Sterilization upon Extracellular Matrix Hydrogel Structure and Function. J. Immunol. Regen. Med. 2018, 2, 11–20. [Google Scholar] [CrossRef]
- Ghassemi, T.; Saghatolslami, N.; Matin, M.M.; Gheshlaghi, R.; Moradi, A. CNT-Decellularized Cartilage Hybrids for Tissue Engineering Applications. Biomed. Mater. 2017, 12, 065008. [Google Scholar] [CrossRef] [PubMed]
- Nasiri, B.; Mashayekhan, S. Fabrication of Porous Scaffolds with Decellularized Cartilage Matrix for Tissue Engineering Application. Biologicals 2017, 48, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Kristofik, N.J.; Qin, L.; Calabro, N.E.; Dimitrievska, S.; Li, G.; Tellides, G.; Niklason, L.E.; Kyriakides, T.R. Improving in Vivo Outcomes of Decellularized Vascular Grafts via Incorporation of a Novel Extracellular Matrix. Biomaterials 2017, 141, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Soffer-Tsur, N.; Peer, D.; Dvir, T. ECM-Based Macroporous Sponges Release Essential Factors to Support the Growth of Hematopoietic Cells. J. Control Release 2017, 257, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.T.; Daly, K.A.; Brennan-Pierce, E.P.; Johnson, S.A.; Carruthers, C.A.; D’Amore, A.; Nagarkar, S.P.; Velankar, S.S.; Badylak, S.F. A Hydrogel Derived from Decellularized Dermal Extracellular Matrix. Biomaterials 2012, 33, 7028–7038. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.; Jung, Y.; Kim, S.H. Decellularized Heart ECM Hydrogel Using Supercritical Carbon Dioxide for Improved Angiogenesis. Acta Biomater. 2018, 67, 270–281. [Google Scholar] [CrossRef]
- Sarmin, A.M.; Connelly, J.T. Fabrication of Human Skin Equivalents Using Decellularized Extracellular Matrix. Curr. Protoc. 2022, 2, e393. [Google Scholar] [CrossRef]
- Mousavi, A.; Mashayekhan, S.; Baheiraei, N.; Pourjavadi, A. Biohybrid Oxidized Alginate/Myocardial Extracellular Matrix Injectable Hydrogels with Improved Electromechanical Properties for Cardiac Tissue Engineering. Int. J. Biol. Macromol. 2021, 180, 692–708. [Google Scholar] [CrossRef]
- Alksne, M.; Kalvaityte, M.; Simoliunas, E.; Gendviliene, I.; Barasa, P.; Rinkunaite, I.; Kaupinis, A.; Seinin, D.; Rutkunas, V.; Bukelskiene, V. Dental Pulp Stem Cell-Derived Extracellular Matrix: Autologous Tool Boosting Bone Regeneration. Cytotherapy 2022, 24, 597–607. [Google Scholar] [CrossRef]
- Song, E.S.; Park, J.H.; Ha, S.S.; Cha, P.H.; Kang, J.T.; Park, C.Y.; Park, K. Novel Corneal Endothelial Cell Carrier Couples a Biodegradable Polymer and a Mesenchymal Stem Cell-Derived Extracellular Matrix. ACS Appl. Mater. Interfaces 2022, 14, 12116–12129. [Google Scholar] [CrossRef] [PubMed]
- Lopera Higuita, M.; Lopera Giraldo, J.F.; Sarrafian, T.L.; Griffiths, L.G. Tissue Engineered Bovine Saphenous Vein Extracellular Matrix Scaffolds Produced via Antigen Removal Achieve High in Vivo Patency Rates. Acta Biomater. 2021, 134, 144–159. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, H.; Morimoto, N.; Yamaoka, T. Decellularization of Submillimeter-Diameter Vascular Scaffolds Using Peracetic Acid. J. Artif. Organs 2020, 23, 156–162. [Google Scholar] [CrossRef]
- Sengyoku, H.; Tsuchiya, T.; Obata, T.; Doi, R.; Hashimoto, Y.; Ishii, M.; Sakai, H.; Matsuo, N.; Taniguchi, D.; Suematsu, T.; et al. Sodium Hydroxide Based Non-Detergent Decellularizing Solution for Rat Lung. Organogenesis 2018, 14, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Nellinger, S.; Mrsic, I.; Keller, S.; Heine, S.; Southan, A.; Bach, M.; Volz, A.; Chassé, T.; Kluger, P.J. Cell-derived and Enzyme-based Decellularized Extracellular Matrix Exhibit Compositional and Structural Differences That Are Relevant for Its Use as a Biomaterial. Biotechnol. Bioeng. 2022, 119, 1142–1156. [Google Scholar] [CrossRef]
- Xu, Y.; Guo, X.; Yang, S.; Li, L.; Zhang, P.; Sun, W.; Liu, C.; Mi, S. Construction of Bionic Tissue Engineering Cartilage Scaffold Based on Three-Dimensional Printing and Oriented Frozen Technology. J. Biomed. Mater. Res. A 2018, 106, 1664–1676. [Google Scholar] [CrossRef]
- Arslan, Y.E.; Galata, Y.F.; Sezgin Arslan, T.; Derkus, B. Trans-Differentiation of Human Adipose-Derived Mesenchymal Stem Cells into Cardiomyocyte-like Cells on Decellularized Bovine Myocardial Extracellular Matrix-Based Films. J. Mater. Sci. Mater. Med. 2018, 29, 127. [Google Scholar] [CrossRef]
- Terpstra, M.L.; Li, J.; Mensinga, A.; de Ruijter, M.; van Rijen, M.H.P.; Androulidakis, C.; Galiotis, C.; Papantoniou, I.; Matsusaki, M.; Malda, J.; et al. Bioink with Cartilage-Derived Extracellular Matrix Microfibers Enables Spatial Control of Vascular Capillary Formation in Bioprinted Constructs. Biofabrication 2022, 14, 034104. [Google Scholar] [CrossRef]
- Chen, M.; Feng, Z.; Guo, W.; Yang, D.; Gao, S.; Li, Y.; Shen, S.; Yuan, Z.; Huang, B.; Zhang, Y.; et al. PCL-MECM-Based Hydrogel Hybrid Scaffolds and Meniscal Fibrochondrocytes Promote Whole Meniscus Regeneration in a Rabbit Meniscectomy Model. ACS Appl. Mater. Interfaces 2019, 11, 41626–41639. [Google Scholar] [CrossRef]
- Nguyen, D.T.; O’Hara, M.; Graneli, C.; Hicks, R.; Miliotis, T.; Nyström, A.-C.; Hansson, S.; Davidsson, P.; Gan, L.-M.; Magnone, M.C.; et al. Humanizing Miniature Hearts through 4-Flow Cannulation Perfusion Decellularization and Recellularization. Sci. Rep. 2018, 8, 7458. [Google Scholar] [CrossRef]
- Liu, H.; Yang, L.; Zhang, E.; Zhang, R.; Cai, D.; Zhu, S.; Ran, J.; Bunpetch, V.; Cai, Y.; Heng, B.C.; et al. Biomimetic Tendon Extracellular Matrix Composite Gradient Scaffold Enhances Ligament-to-Bone Junction Reconstruction. Acta Biomater. 2017, 56, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Azhim, A.; Syazwani, N.; Morimoto, Y.; Furukawa, K.; Ushida, T. The Use of Sonication Treatment to Decellularize Aortic Tissues for Preparation of Bioscaffolds. J Biomater Appl 2014, 29, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Suss, P.H.; Ribeiro, V.S.T.; Motooka, C.E.; de Melo, L.C.; Tuon, F.F. Comparative Study of Decellularization Techniques to Obtain Natural Extracellular Matrix Scaffolds of Human Peripheral-Nerve Allografts. Cell Tissue Bank. 2021, 23, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.T.; Zhou, V.X.; Rubinsky, B. Using Non-Thermal Irreversible Electroporation to Create an in Vivo Niche for Exogenous Cell Engraftment. Biotechniques 2017, 62, 229–231. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.A.; Narayan, R.; Padath, T.; Rubinsky, B. Irreversible Electroporation on the Small Intestine. Br. J. Cancer 2012, 106, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.; Maor, E.; Rubinsky, B. Nonthermal Irreversible Electroporation for Tissue Decellularization. J. Biomech. Eng. 2010, 132, 091003. [Google Scholar] [CrossRef]
- Phillips, M.; Maor, E.; Rubinsky, B. Principles of Tissue Engineering with Nonthermal Irreversible Electroporation. J. Heat. Transfer. 2010, 133, 011004. [Google Scholar] [CrossRef]
- Sano, M.B.; Neal, R.E.; Garcia, P.A.; Gerber, D.; Robertson, J.; Davalos, R. V Towards the Creation of Decellularized Organ Constructs Using Irreversible Electroporation and Active Mechanical Perfusion. Biomed. Eng. Online 2010, 9, 83. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Tuffin, J.; Lei, I.M.; Ruggeri, F.S.; Lewis, N.S.; Gill, E.L.; Savin, T.; Huleihel, L.; Badylak, S.F.; Knowles, T.; et al. Solution Fibre Spinning Technique for the Fabrication of Tuneable Decellularised Matrix-Laden Fibres and Fibrous Micromembranes. Acta Biomater. 2018, 78, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Karthika, S.; Anoop, S.; Devanand, C.B.; Narayanan, M.K.; Unni, M.; Eassow, S.; Anilkumar, T. A Porcine-Cholecyst-Derived Scaffold for Treating Full Thickness Lacerated Skin Wounds in Dogs. Vet. Res. Commun. 2018, 42, 233–242. [Google Scholar] [CrossRef]
- Costa, A.; Naranjo, J.D.; Turner, N.J.; Swinehart, I.T.; Kolich, B.D.; Shaffiey, S.A.; Londono, R.; Keane, T.J.; Reing, J.E.; Johnson, S.A.; et al. Mechanical Strength vs. Degradation of a Biologically-Derived Surgical Mesh over Time in a Rodent Full Thickness Abdominal Wall Defect. Biomaterials 2016, 108, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Dziki, J.L.; Keane, T.J.; Shaffiey, S.; Cognetti, D.; Turner, N.; Nagle, D.; Hackam, D.; Badylak, S.F. Bioscaffold-Mediated Mucosal Remodelling Following Short-Segment Colonic Mucosal Resection. J. Surg. Res. 2017, 218, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Ghuman, H.; Gerwig, M.; Nicholls, F.J.; Liu, J.R.; Donnelly, J.; Badylak, S.F.; Modo, M. Long-Term Retention of ECM Hydrogel after Implantation into a Sub-Acute Stroke Cavity Reduces Lesion Volume. Acta Biomater. 2017, 63, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Poornejad, N.; Momtahan, N.; Salehi, A.S.M.; Scott, D.R.; Fronk, C.A.; Roeder, B.L.; Reynolds, P.R.; Bundy, B.C.; Cook, A.D. Efficient Decellularization of Whole Porcine Kidneys Improves Reseeded Cell Behaviour. Biomed. Mater. 2016, 11, 025003. [Google Scholar] [CrossRef] [PubMed]
- Willemse, J.; Verstegen, M.M.A.; Vermeulen, A.; Schurink, I.J.; Roest, H.P.; van der Laan, L.J.W.; de Jonge, J. Fast, Robust and Effective Decellularization of Whole Human Livers Using Mild Detergents and Pressure Controlled Perfusion. Mater. Sci. Eng. C 2020, 108, 110200. [Google Scholar] [CrossRef]
- Reisbig, N.A.; Hussein, H.A.; Pinnell, E.; Bertone, A.L. Evaluation of Equine Synovial-Derived Extracellular Matrix Scaffolds Seeded with Equine Synovial-Derived Mesenchymal Stem Cells. Am. J. Vet. Res. 2018, 79, 124–133. [Google Scholar] [CrossRef]
- Gil-Ramírez, A.; Rosmark, O.; Spégel, P.; Swärd, K.; Westergren-Thorsson, G.; Larsson-Callerfelt, A.-K.; Rodríguez-Meizoso, I. Pressurized Carbon Dioxide as a Potential Tool for Decellularization of Pulmonary Arteries for Transplant Purposes. Sci. Rep. 2020, 10, 4031. [Google Scholar] [CrossRef] [Green Version]
- Sawada, K.; Terada, D.; Yamaoka, T.; Kitamura, S.; Fujisato, T. Cell Removal with Supercritical Carbon Dioxide for Acellular Artificial Tissue. J. Chem. Technol. Biotechnol. 2008, 83, 943–949. [Google Scholar] [CrossRef]
- Crapo, P.M.; Gilbert, T.W.; Badylak, S.F. An Overview of Tissue and Whole Organ Decellularization Processes. Biomaterials 2011, 32, 3233–3243. [Google Scholar] [CrossRef] [Green Version]
- Keane, T.J.; Swinehart, I.T.; Badylak, S.F. Methods of Tissue Decellularization Used for Preparation of Biologic Scaffolds and in Vivo Relevance. Methods 2015, 84, 25–34. [Google Scholar] [CrossRef]
- Mora-Navarro, C.; Garcia, M.E.; Sarker, P.; Ozpinar, E.W.; Enders, J.R.; Khan, S.; Branski, R.C.; Freytes, D.O. Monitoring Decellularization via Absorbance Spectroscopy during the Derivation of Extracellular Matrix Scaffolds. Biomed. Mater. 2022, 17, 015008. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Liu, S.; Hao, C.; Guo, W.; Gao, S.; Wang, M.; Chen, M.; Sun, Z.; Xu, Y.; Wang, Y.; et al. AMECM/DCB Scaffold Prompts Successful Total Meniscus Reconstruction in a Rabbit Total Meniscectomy Model. Biomaterials 2016, 111, 13–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bejleri, D.; Streeter, B.W.; Nachlas, A.L.Y.; Brown, M.E.; Gaetani, R.; Christman, K.L.; Davis, M.E. A Bioprinted Cardiac Patch Composed of Cardiac-Specific Extracellular Matrix and Progenitor Cells for Heart Repair. Adv. Healthc. Mater. 2018, 7, 1800672. [Google Scholar] [CrossRef] [PubMed]
- Chmura, D.; Leigh, J. Replacing Body Parts. NOVA ScienceNOW, 2011, Season 5, Episode 2.
- Layland, J.; Carrick, D.; Lee, M.; Oldroyd, K.; Berry, C. Adenosine: Physiology, Pharmacology, and Clinical Applications. JACC Cardiovasc. Interv. 2014, 7, 581–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, S. The Interaction of Triton X-100 with Soluble Proteins: Possible Implications for the Transport of Proteins across Membranes. Biochem. Biophys. Res. Commun. 1977, 79, 46–52. [Google Scholar] [CrossRef]
- Ott, H.C.; Matthiesen, T.S.; Goh, S.-K.; Black, L.D.; Kren, S.M.; Netoff, T.I.; Taylor, D.A. Perfusion-Decellularized Matrix: Using Nature’s Platform to Engineer a Bioartificial Heart. Nat. Med. 2008, 14, 213–221. [Google Scholar] [CrossRef]
- Ott, H.C.; Clippinger, B.; Conrad, C.; Schuetz, C.; Pomerantseva, I.; Ikonomou, L.; Kotton, D.; Vacanti, J.P. Regeneration and Orthotopic Transplantation of a Bioartificial Lung. Nat. Med. 2010, 16, 927–933. [Google Scholar] [CrossRef]
- Song, J.J.; Guyette, J.; Gilpin, S.; Gonzalez, G.; Vacanti, J.P.; Ott, H.C. Regeneration and Experimental Orthotopic Transplantation of a Bioengineered Kidney. Nat. Med. 2013, 19, 646–651. [Google Scholar] [CrossRef]
- Gerli, M.F.M.; Guyette, J.P.; Evangelista-Leite, D.; Ghoshhajra, B.B.; Ott, H.C. Perfusion Decellularization of a Human Limb: A Novel Platform for Composite Tissue Engineering and Reconstructive Surgery. PLoS ONE 2018, 13, e0191497. [Google Scholar] [CrossRef] [Green Version]
- Struecker, B.; Butter, A.; Hillebrandt, K.; Polenz, D.; Reutzel-Selke, A.; Tang, P.; Lippert, S.; Leder, A.; Rohn, S.; Geisel, D.; et al. Improved Rat Liver Decellularization by Arterial Perfusion under Oscillating Pressure Conditions. J. Tissue Eng. Regen. Med. 2017, 11, 531–541. [Google Scholar] [CrossRef]
- Hou, N.; Xu, X.; Lv, D.; Lu, Y.; Li, J.; Cui, P.; Ma, R.; Luo, X.; Tang, Y.; Zheng, Y. Tissue-Engineered Esophagus: Recellular Esophageal Extracellular Matrix Based on Perfusion-Decellularized Technique and Mesenchymal Stem Cells. Biomed. Mater. 2021, 16, 055017. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Lee, M.S.; Yang, H.S. Differentiated Osteoblasts Derived Decellularized Extracellular Matrix to Promote Osteogenic Differentiation. Biomater. Res. 2018, 22, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.G.; Gil, C.H.; Seo, J.; Park, S.J.; Subbiah, R.; Jung, T.H.; Kim, J.S.; Jeong, Y.H.; Chung, H.M.; Lee, J.H.; et al. Mechanotransduction of Human Pluripotent Stem Cells Cultivated on Tunable Cell-Derived Extracellular Matrix. Biomaterials 2018, 150, 100–111. [Google Scholar] [CrossRef]
- Carvalho, M.S.; Silva, J.C.; Cabral, J.M.S.; Silva, C.L.; Vashishth, D. Cultured Cell-derived Extracellular Matrices to Enhance the Osteogenic Differentiation and Angiogenic Properties of Human Mesenchymal Stem/Stromal Cells. J. Tissue Eng. Regen. Med. 2019, 13, 1544–1558. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Sharma, D.; Jia, W.; Radke, D.; Kamp, T.; Zhao, F. Engineering Stem Cell Cardiac Patch with Microvascular Features Representative of Native Myocardium. Theranostics 2019, 9, 2143–2157. [Google Scholar] [CrossRef]
- Yan, J.; Chen, X.; Pu, C.; Zhao, Y.; Liu, X.; Liu, T.; Pan, G.; Lin, J.; Pei, M.; Yang, H.; et al. Synovium Stem Cell-Derived Matrix Enhances Anti-Inflammatory Properties of Rabbit Articular Chondrocytes via the SIRT1 Pathway. Mater. Sci. Eng. C 2020, 106, 110286. [Google Scholar] [CrossRef]
- Han, C.; Luo, X.; Zou, D.; Li, J.; Zhang, K.; Yang, P.; Huang, N. Nature-Inspired Extracellular Matrix Coating Produced by Micro-Patterned Smooth Muscle and Endothelial Cells Endows Cardiovascular Materials with Better Biocompatibility. Biomater. Sci. 2019, 7, 2686–2701. [Google Scholar] [CrossRef]
- Zhai, Y.; Wang, Q.; Zhu, Z.; Zheng, W.; Ma, S.; Hao, Y.; Yang, L.; Cheng, G. Cell-Derived Extracellular Matrix Enhanced by Collagen-Binding Domain-Decorated Exosomes to Promote Neural Stem Cells Neurogenesis. Biomed. Mater. 2022, 17, 14104. [Google Scholar] [CrossRef]
- Wang, Z.; Han, L.; Sun, T.; Ma, J.; Sun, S.; Ma, L.; Wu, B. Extracellular Matrix Derived from Allogenic Decellularized Bone Marrow Mesenchymal Stem Cell Sheets for the Reconstruction of Osteochondral Defects in Rabbits. Acta Biomater. 2020, 118, 54–68. [Google Scholar] [CrossRef]
- Wang, Y.; Duan, M.; Rahman, M.; Yang, M.; Zhao, W.; Zhou, S.; Gao, G.; Fu, Q. Use of Bioactive Extracellular Matrix Fragments as a Urethral Bulking Agent to Treat Stress Urinary Incontinence. Acta Biomater. 2020, 117, 156–166. [Google Scholar] [CrossRef]
- Mertsch, S.; Hasenzahl, M.; Reichl, S.; Geerling, G.; Schrader, S. Decellularized Human Corneal Stromal Cell Sheet as a Novel Matrix for Ocular Surface Reconstruction. J. Tissue Eng. Regen. Med. 2020, 14, 1318–1332. [Google Scholar] [CrossRef] [PubMed]
- Magnan, L.; Labrunie, G.; Fénelon, M.; Dusserre, N.; Foulc, M.P.; Lafourcade, M.; Svahn, I.; Gontier, E.; Vélez, V.J.H.; McAllister, T.N.; et al. Human Textiles: A Cell-Synthesized Yarn as a Truly “Bio” Material for Tissue Engineering Applications. Acta Biomater. 2020, 105, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, C.; Zhong, Y.; Zhao, J. A Novel Approach to Utilize Icariin as Icariin-Derived ECM on Small Intestinal Submucosa Scaffold for Bone Repair. Ann. Biomed. Eng. 2017, 45, 2673–2682. [Google Scholar] [CrossRef]
- Mielan, B.; Sousa, D.; Krok-Borkowicz, M.; Eloy, P.; Dupont, C.; Lamghari, M.; Pamuła, E. Polymeric Microspheres/Cells/Extracellular Matrix Constructs Produced by Auto-Assembly for Bone Modular Tissue Engineering. Int. J. Mol. Sci. 2021, 22, 7897. [Google Scholar] [CrossRef] [PubMed]
- Burgio, F.; Rimmer, N.; Pieles, U.; Buschmann, J.; Beaufils-Hugot, M. Characterization and in Ovo Vascularization of a 3D-Printed Hydroxyapatite Scaffold with Different Extracellular Matrix Coatings under Perfusion Culture. Biol. Open. 2018, 7, bio034488. [Google Scholar] [CrossRef] [Green Version]
- Blum, J.C.; Schenck, T.L.; Birt, A.; Giunta, R.E.; Wiggenhauser, P.S. Artificial Decellularized Extracellular Matrix Improves the Regenerative Capacity of Adipose Tissue Derived Stem Cells on 3D Printed Polycaprolactone Scaffolds. J. Tissue Eng. 2021, 12, 20417314211022242. [Google Scholar] [CrossRef]
- Li, W.; Midgley, A.C.; Bai, Y.; Zhu, M.; Chang, H.; Zhu, W.; Wang, L.; Wang, Y.; Wang, H.; Kong, D. Subcutaneously Engineered Autologous Extracellular Matrix Scaffolds with Aligned Microchannels for Enhanced Tendon Regeneration: Aligned Microchannel Scaffolds for Tendon Repair. Biomaterials 2019, 224, 119488. [Google Scholar] [CrossRef]
- Lange, P.; Greco, K.; Partington, L.; Carvalho, C.; Oliani, S.; Birchall, M.A.; Sibbons, P.D.; Lowdell, M.W.; Ansari, T. Pilot Study of a Novel Vacuum-Assisted Method for Decellularization of Tracheae for Clinical Tissue Engineering Applications. J. Tissue Eng. Regen. Med. 2017, 11, 800–811. [Google Scholar] [CrossRef]
- Xu, K.; Kuntz, L.A.; Foehr, P.; Kuempel, K.; Wagner, A.; Tuebel, J.; Deimling, C.V.; Burgkart, R.H. Efficient Decellularization for Tissue Engineering of the Tendon-Bone Interface with Preservation of Biomechanics. PLoS ONE 2017, 12, e0171577. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, N.; Ito, A.; Kimura, T.; Kishida, A. Extracellular Matrix Induces Periodontal Ligament Reconstruction In Vivo. Int. J. Mol. Sci. 2019, 20, 3277. [Google Scholar] [CrossRef]
- Kobayashi, M.; Ohara, M.; Hashimoto, Y.; Nakamura, N.; Fujisato, T.; Kimura, T.; Kishida, A. In Vitro Evaluation of Surface Biological Properties of Decellularized Aorta for Cardiovascular Use. J. Mater. Chem. B 2020, 8, 10977–10989. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Kadota, J.; Hashimoto, Y.; Fujisato, T.; Nakamura, N.; Kimura, T.; Kishida, A. Elastic Modulus of ECM Hydrogels Derived from Decellularized Tissue Affects Capillary Network Formation in Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 6304. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ishida, N.; Hashimoto, Y.; Negishi, J.; Saga, H.; Sasaki, Y.; Akiyoshi, K.; Kimura, T.; Kishida, A. Extraction and Biological Evaluation of Matrix-Bound Nanovesicles (MBVs) from High-Hydrostatic Pressure-Decellularized Tissues. Int. J. Mol. Sci. 2022, 23, 8868. [Google Scholar] [CrossRef] [PubMed]
- Charoensombut, N.; Kawabata, K.; Kim, J.; Chang, M.; Kimura, T.; Kishida, A.; Ushida, T.; Furukawa, K.S. Internal Radial Perfusion Bioreactor Promotes Decellularization and Recellularization of Rat Uterine Tissue. J. Biosci. Bioeng. 2022, 133, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, S.; Hashimoto, Y.; Funamoto, S.; Murata, K.; Yamashita, A.; Yamazaki, K.; Ikeda, T.; Minatoya, K.; Kishida, A.; Masumoto, H. In Vivo Recellularization of Xenogeneic Vascular Grafts Decellularized with High Hydrostatic Pressure Method in a Porcine Carotid Arterial Interpose Model. PLoS ONE 2021, 16, e0254160. [Google Scholar] [CrossRef]
- Tchoukalova, Y.D.; Hintze, J.M.; Hayden, R.E.; Lott, D.G. Tracheal Decellularization Using a Combination of Chemical, Physical and Bioreactor Methods. Int. J. Artif. Organs 2018, 41, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Mao, H.; Moser, M.A.J.; Zhang, W.; Qian, Z.; Zhang, B. Irreversible Electroporation Enhanced by Radiofrequency Ablation: An In Vitro and Computational Study in a 3D Liver Tumour Model. Ann. Biomed. Eng. 2021, 49, 2126–2138. [Google Scholar] [CrossRef]
- Ding, L.; Moser, M.; Luo, Y.; Zhang, W.; Zhang, B. Treatment Planning Optimization in Irreversible Electroporation for Complete Ablation of Variously Sized Cervical Tumours: A Numerical Study. J. Biomech. Eng. 2020, 143, 014503. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, Y.; Ding, L.; Moser, M.A.J.; Zhang, E.M.; Zhang, W. Tumour Ablation Enhancement by Combining Radiofrequency Ablation and Irreversible Electroporation: An In Vitro 3D Tumour Study. Ann Biomed. Eng. 2019, 47, 694–705. [Google Scholar] [CrossRef]
- Yang, Y.; Moser, M.A.J.; Zhang, E.; Zhang, W.; Zhang, B. Development of a Statistical Model for Cervical Cancer Cell Death with Irreversible Electroporation in Vitro. PLoS ONE 2018, 13, e0195561. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, B.; Moser, M.; Zhang, E.; Zhang, W. Analysis and Optimization of Determining Factors in Irreversible Electroporation for Large Ablation Zones without Thermal Damage. In Proceedings of the ASME 2017 International Mechanical Engineering Congress and Exposition, Tampa, FL, USA, 3–9 November 2017. [Google Scholar] [CrossRef]
- Del Amo, C.; Fernández-San Argimiro, X.; Cascajo-Castresana, M.; Perez-Valle, A.; Madarieta, I.; Olalde, B.; Andia, I. Wound-Microenvironment Engineering through Advanced-Dressing Bioprinting. Int. J. Mol. Sci. 2022, 23, 2836. [Google Scholar] [CrossRef] [PubMed]
- Shojarazavi, N.; Mashayekhan, S.; Pazooki, H.; Mohsenifard, S.; Baniasadi, H. Alginate/Cartilage Extracellular Matrix-Based Injectable Interpenetrating Polymer Network Hydrogel for Cartilage Tissue Engineering. J. Biomater. Appl. 2021, 36, 803–817. [Google Scholar] [CrossRef] [PubMed]
- Saeedi Garakani, S.; Khanmohammadi, M.; Atoufi, Z.; Kamrava, S.K.; Setayeshmehr, M.; Alizadeh, R.; Faghihi, F.; Bagher, Z.; Davachi, S.M.; Abbaspourrad, A. Fabrication of Chitosan/Agarose Scaffolds Containing Extracellular Matrix for Tissue Engineering Applications. Int. J. Biol. Macromol. 2020, 143, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Chu, T.L.; Tripathi, G.; Ho, B.S.; Lee, B.T. In-Vitro and in-Vivo Hemostat Evaluation of Decellularized Liver Extra Cellular Matrix Loaded Chitosan/Gelatin Spongy Scaffolds for Liver Injury. Int. J. Biol. Macromol. 2021, 193, 638–646. [Google Scholar] [CrossRef]
- Frost, S.J.; Mawad, D.; Wuhrer, R.; Myers, S.; Lauto, A. Semitransparent Bandages Based on Chitosan and Extracellular Matrix for Photochemical Tissue Bonding. Biomed. Eng. Online 2018, 17, 7. [Google Scholar] [CrossRef] [Green Version]
- Raj, R.; Sobhan, P.K.; Pratheesh, K.V.; Anilkumar, T.V. A Cholecystic Extracellular Matrix-based Hybrid Hydrogel for Skeletal Muscle Tissue Engineering. J. Biomed. Mater. Res. A 2020, 108, 1922–1933. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Ahn, G.; Kim, C.; Lee, J.-S.; Lee, I.-G.; An, S.-H.; Yun, W.-S.; Kim, S.-Y.; Shim, J.-H. Synergistic Effects of Beta Tri-Calcium Phosphate and Porcine-Derived Decellularized Bone Extracellular Matrix in 3D-Printed Polycaprolactone Scaffold on Bone Regeneration. Macromol. Biosci. 2018, 18, 1800025. [Google Scholar] [CrossRef]
- Guo, W.; Chen, M.; Wang, Z.; Tian, Y.; Zheng, J.; Gao, S.; Li, Y.; Zheng, Y.; Li, X.; Huang, J.; et al. 3D-Printed Cell-Free PCL–MECM Scaffold with Biomimetic Micro-Structure and Micro-Environment to Enhance in Situ Meniscus Regeneration. Bioact. Mater. 2021, 6, 3620–3633. [Google Scholar] [CrossRef]
- Xie, X.; Wang, W.; Cheng, J.; Liang, H.; Lin, Z.; Zhang, T.; Lu, Y.; Li, Q. Bilayer Pifithrin-α Loaded Extracellular Matrix/PLGA Scaffolds for Enhanced Vascularized Bone Formation. Colloids Surf. B Biointerfaces 2020, 190, 110903. [Google Scholar] [CrossRef]
- Singh, H.; Purohit, S.D.; Bhaskar, R.; Yadav, I.; Bhushan, S.; Gupta, M.K.; Mishra, N.C. Curcumin in Decellularized Goat Small Intestine Submucosa for Wound Healing and Skin Tissue Engineering. J. Biomed. Mater. Res. B Appl. Biomater. 2022, 110, 210–219. [Google Scholar] [CrossRef]
- Chen, P.; Zheng, L.; Wang, Y.; Tao, M.; Xie, Z.; Xia, C.; Gu, C.; Chen, J.; Qiu, P.; Mei, S.; et al. Desktop-Stereolithography 3D Printing of a Radially Oriented Extracellular Matrix/Mesenchymal Stem Cell Exosome Bioink for Osteochondral Defect Regeneration. Theranostics 2019, 9, 2439–2459. [Google Scholar] [CrossRef] [PubMed]
- Homan, K.A.; Kolesky, D.B.; Skylar-Scott, M.A.; Herrmann, J.; Obuobi, H.; Moisan, A.; Lewis, J.A. Bioprinting of 3D Convoluted Renal Proximal Tubules on Perfusable Chips. Sci. Rep. 2016, 6, 34845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolesky, D.B.; Truby, R.L.; Gladman, A.S.; Busbee, T.A.; Homan, K.A.; Lewis, J.A. 3D Bioprinting of Vascularized, Heterogeneous Cell-Laden Tissue Constructs. Adv. Mater. 2014, 26, 3124–3130. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; DeConinck, A.; Lewis, J.A. Omnidirectional Printing of 3D Microvascular Networks. Adv. Mater. 2011, 23, H178–H183. [Google Scholar] [CrossRef]
- Shin, Y.J.; Shafranek, R.T.; Tsui, J.H.; Walcott, J.; Nelson, A.; Kim, D.H. 3D Bioprinting of Mechanically Tuned Bioinks Derived from Cardiac Decellularized Extracellular Matrix. Acta Biomater. 2021, 119, 75–88. [Google Scholar] [CrossRef]
- Jeon, E.Y.; Joo, K.I.; Cha, H.J. Body Temperature-Activated Protein-Based Injectable Adhesive Hydrogel Incorporated with Decellularized Adipose Extracellular Matrix for Tissue-Specific Regenerative Stem Cell Therapy. Acta Biomater. 2020, 114, 244–255. [Google Scholar] [CrossRef]
- Nyambat, B.; Manga, Y.B.; Chen, C.-H.; Gankhuyag, U.; Pratomo WP, A.; Kumar Satapathy, M.; Chuang, E.-Y. New Insight into Natural Extracellular Matrix: Genipin Cross-Linked Adipose-Derived Stem Cell Extracellular Matrix Gel for Tissue Engineering. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Nishiguchi, A.; Taguchi, T. A PH-Driven Genipin Gelator to Engineer Decellularized Extracellular Matrix-Based Tissue Adhesives. Acta Biomater. 2021, 131, 211–221. [Google Scholar] [CrossRef]
- Smith, M.J.; Dempsey, S.G.; Veale, R.W.F.; Duston-Fursman, C.G.; Rayner, C.A.F.; Javanapong, C.; Gerneke, D.; Dowling, S.G.; Bosque, B.A.; Karnik, T.; et al. Further Structural Characterization of Ovine Forestomach Matrix and Multi-Layered Extracellular Matrix Composites for Soft Tissue Repair. J. Biomater. Appl. 2022, 36, 996–1010. [Google Scholar] [CrossRef]
- Valtera, J.; Kalous, T.; Pokorny, P.; Batka, O.; Bilek, M.; Chvojka, J.; Mikes, P.; Kostakova, E.K.; Zabka, P.; Ornstova, J.; et al. Fabrication of Dual-Functional Composite Yarns with a Nanofibrous Envelope Using High Throughput AC Needleless and Collectorless Electrospinning. Sci. Rep. 2019, 9, 1801. [Google Scholar] [CrossRef] [Green Version]
- Sivan, M.; Madheswaran, D.; Valtera, J.; Kostakova, E.K.; Lukas, D. Alternating Current Electrospinning: The Impacts of Various High-Voltage Signal Shapes and Frequencies on the Spinnability and Productivity of Polycaprolactone Nanofibers. Mater. Des. 2022, 213, 110308. [Google Scholar] [CrossRef]
- Balogh, A.; Cselkó, R.; Démuth, B.; Verreck, G.; Mensch, J.; Marosi, G.; Nagy, Z.K. Alternating Current Electrospinning for Preparation of Fibrous Drug Delivery Systems. Int. J. Pharm. 2015, 495, 75–80. [Google Scholar] [CrossRef]
- Partheniadis, I.; Nikolakakis, I.; Laidmäe, I.; Heinämäki, J. A Mini-Review: Needleless Electrospinning of Nanofibers for Pharmaceutical and Biomedical Applications. Processes 2020, 8, 673. [Google Scholar] [CrossRef]
- Grant, R.; Hallett, J.; Forbes, S.; Hay, D.; Callanan, A. Blended Electrospinning with Human Liver Extracellular Matrix for Engineering New Hepatic Microenvironments. Sci. Rep. 2019, 9, 6293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smoak, M.M.; Hogan, K.J.; Grande-Allen, K.J.; Mikos, A.G. Bioinspired Electrospun DECM Scaffolds Guide Cell Growth and Control the Formation of Myotubes. Sci. Adv. 2021, 7, 4123–4137. [Google Scholar] [CrossRef] [PubMed]
- Smoak, M.M.; Han, A.; Watson, E.; Kishan, A.; Grande-Allen, K.J.; Cosgriff-Hernandez, E.; Mikos, A.G. Fabrication and Characterization of Electrospun Decellularized Muscle-Derived Scaffolds. Tissue Eng. Part C Methods 2019, 25, 276–287. [Google Scholar] [CrossRef]
- Patel, K.H.; Dunn, A.J.; Talovic, M.; Haas, G.J.; Marcinczyk, M.; Elmashhady, H.; Kalaf, E.G.; Sell, S.A.; Garg, K. Aligned Nanofibers of Decellularized Muscle ECM Support Myogenic Activity in Primary Satellite Cells in Vitro. Biomed. Mater. 2019, 14, 035010. [Google Scholar] [CrossRef]
- Montero, R.B.; Vial, X.; Nguyen, D.T.; Farhand, S.; Reardon, M.; Pham, S.M.; Tsechpenakis, G.; Andreopoulos, F.M. BFGF-Containing Electrospun Gelatin Scaffolds with Controlled Nano-Architectural Features for Directed Angiogenesis. Acta Biomater. 2012, 8, 1778–1791. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Pérez, J.; Kador, K.E.; Lynch, A.P.; Ahearne, M. Characterization of Extracellular Matrix Modified Poly(ε-Caprolactone) Electrospun Scaffolds with Differing Fiber Orientations for Corneal Stroma Regeneration. Mater. Sci. Eng. C 2020, 108, 110415. [Google Scholar] [CrossRef]
- Li, X.; Li, Z.; Wang, L.; Ma, G.; Meng, F.; Pritchard, R.H.; Gill, E.L.; Liu, Y.; Huang, Y.Y.S. Low-Voltage Continuous Electrospinning Patterning. ACS Appl. Mater. Interfaces 2016, 8, 32120–32131. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, M.S.; Silva, J.C.; Udangawa, R.N.; Cabral, J.M.S.; Ferreira, F.C.; da Silva, C.L.; Linhardt, R.J.; Vashishth, D. Co-Culture Cell-Derived Extracellular Matrix Loaded Electrospun Microfibrous Scaffolds for Bone Tissue Engineering. Mater. Sci. Eng. C 2019, 99, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Sobreiro-Almeida, R.; Fonseca, D.R.; Neves, N.M. Extracellular Matrix Electrospun Membranes for Mimicking Natural Renal Filtration Barriers. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 103, 109866. [Google Scholar] [CrossRef]
- Liu, C.; Li, Y.; Zhang, Y.; Xu, H. The Experimental Study of Regeneration of Annulus Fibrosus Using Decellularized Annulus Fibrosus Matrix/Poly(Ether Carbonate Urethane)Urea-blended Fibrous Scaffolds with Varying Elastic Moduli. J. Biomed. Mater. Res. A 2022, 110, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Mejía Suaza, M.L.; Hurtado Henao, Y.; Moncada Acevedo, M.E. Wet Electrospinning and Its Applications: A Review. TecnoLógicas 2022, 25, e2223. [Google Scholar] [CrossRef]
- Zainuddin, S.; Scheibel, T. Continuous Yarn Electrospinning. Textiles 2022, 2, 124–141. [Google Scholar] [CrossRef]
- Song, J.-H.; Kim, H.-E.; Kim, H.-W. Production of Electrospun Gelatin Nanofiber by Water-Based Co-Solvent Approach. J. Mater. Sci. Mater. Med. 2008, 19, 95–102. [Google Scholar] [CrossRef]
- Kornmuller, A.; Flynn, L.E. Development and Characterization of Matrix-derived Microcarriers from Decellularized Tissues Using Electrospraying Techniques. J. Biomed. Mater. Res. A 2022, 110, 559–575. [Google Scholar] [CrossRef]
- Xia, B.; Kim, D.H.; Bansal, S.; Bae, Y.; Mauck, R.L.; Heo, S.J. Development of a Decellularized Meniscus Matrix-Based Nanofibrous Scaffold for Meniscus Tissue Engineering. Acta Biomater. 2021, 128, 175–185. [Google Scholar] [CrossRef]
- Reid, J.A.; Callanan, A. Hybrid Cardiovascular Sourced Extracellular Matrix Scaffolds as Possible Platforms for Vascular Tissue Engineering. J. Biomed. Mater. Res. B Appl Biomater 2020, 108, 910–924. [Google Scholar] [CrossRef]
- Piore, A.; Lewis, S. How Pig Guts Became the Next Bright Hope for Regenerating Human Limbs. Discov. Mag. 2011, 68–73. [Google Scholar]
- Lantz, G.C.; Badylak, S.F.; Coffey, A.C.; Geddes, L.A.; Blevins, W.E. Small Intestinal Submucosa as a Small-Diameter Arterial Graft in the Dog. J. Investig. Surg. 1990, 3, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Badylak, S.F.; Lantz, G.C.; Coffey, A.; Geddes, L.A. Small Intestinal Submucosa as a Large Diameter Vascular Graft in the Dog. J. Surg. Res. 1989, 47, 74–80. [Google Scholar] [CrossRef]
- Badylak, S.F.; Tullius, R.; Kokini, K.; Shelbourne, K.D.; Klootwyk, T.; Voytik, S.L.; Kraine, M.R.; Simmons, C. The Use of Xenogeneic Small Intestinal Submucosa as a Biomaterial for Achille’s Tendon Repair in a Dog Model. J. Biomed. Mater. Res. 1995, 29, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Browe, D.C.; Díaz-Payno, P.J.; Freeman, F.E.; Schipani, R.; Burdis, R.; Ahern, D.P.; Nulty, J.M.; Guler, S.; Randall, L.D.; Buckley, C.T.; et al. Bilayered Extracellular Matrix Derived Scaffolds with Anisotropic Pore Architecture Guide Tissue Organization during Osteochondral Defect Repair. Acta Biomater. 2022, 143, 266–281. [Google Scholar] [CrossRef] [PubMed]
- Raj, R. Preparation and Characterization of Cholecystic Extracellular-Matrix Powder; SCTIMST: Kerala, India, 2015. [Google Scholar]
- Gilbert, T.W.; Stolz, D.B.; Biancaniello, F.; Simmons-Byrd, A.; Badylak, S.F. Production and Characterization of ECM Powder: Implications for Tissue Engineering Applications. Biomaterials 2005, 26, 1431–1435. [Google Scholar] [CrossRef] [PubMed]
- Song, B.R.; Yang, S.S.; Jin, H.; Lee, S.H.; Park, D.Y.; Lee, J.H.; Park, S.R.; Park, S.H.; Min, B.H. Three Dimensional Plotted Extracellular Matrix Scaffolds Using a Rapid Prototyping for Tissue Engineering Application. Tissue Eng. Regen Med 2015, 12, 172–180. [Google Scholar] [CrossRef]
- Freytes, D.O.; Martin, J.; Velankar, S.S.; Lee, A.S.; Badylak, S.F. Preparation and Rheological Characterization of a Gel Form of the Porcine Urinary Bladder Matrix. Biomaterials 2008, 29, 1630–1637. [Google Scholar] [CrossRef]
- Zhao, F.; Cheng, J.; Zhang, J.; Yu, H.; Dai, W.; Yan, W.; Sun, M.; Ding, G.; Li, Q.; Meng, Q.; et al. Comparison of Three Different Acidic Solutions in Tendon Decellularized Extracellular Matrix Bio-Ink Fabrication for 3D Cell Printing. Acta Biomater. 2021, 131, 262–275. [Google Scholar] [CrossRef]
- Zhao, F.; Cheng, J.; Sun, M.; Yu, H.; Wu, N.; Li, Z.; Zhang, J.; Li, Q.; Yang, P.; Liu, Q.; et al. Digestion Degree Is a Key Factor to Regulate the Printability of Pure Tendon Decellularized Extracellular Matrix Bio-Ink in Extrusion-Based 3D Cell Printing. Biofabrication 2020, 12, 45011. [Google Scholar] [CrossRef]
- Li, M.; Tian, X.; Schreyer, D.J.; Chen, X. Effect of Needle Geometry on Flow Rate and Cell Damage in the Dispensing-Based Biofabrication Process. Biotechnol. Prog. 2011, 27, 1777–1784. [Google Scholar] [CrossRef]
- Li, M.; Tian, X.; Zhu, N.; Schreyer, D.J.; Chen, X. Modelling Process-Induced Cell Damage in the Biodispensing Process. Tissue Eng. Part C Methods 2010, 16, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.X.B. Extrusion Bioprinting of Scaffolds for Tissue Engineering Applications; Springer International Publishing, 2019; ISBN 9783030034603. In Extrusion Bioprinting of Scaffolds for Tissue Engineering Applications; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; ISBN 9783030034603. [Google Scholar]
- Ning, L.; Betancourt, N.; Schreyer, D.J.; Chen, X. Characterization of Cell Damage and Proliferative Ability during and after Bioprinting. ACS Biomater. Sci. Eng. 2018, 4, 3906–3918. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Tian, X.; Kozinski, J.A.; Chen, X.; Hwang, D. Modelling Mechanical Cell Damage in the Bioprinting Process Employing a Conical Needle. J. Mech. Med. Biol. 2015, 15, 1550073. [Google Scholar] [CrossRef]
- Li, M.G.; Tian, X.Y.Y.; Chen, X. Temperature Effect on the Shear-Induced Cell Damage in Biofabrication. Artif Organs 2011, 35, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Ferris, C.J.; Gilmore, K.J.; Beirne, S.; McCallum, D.; Wallace, G.G.; in het Panhuis, M. Bio-Ink for on-Demand Printing of Living Cells. Biomater. Sci. 2013, 1, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Naghieh, S.; Chen, D. Printability—A Key Issue in Extrusion-Based Bioprinting. J. Pharm. Anal. 2021, 11, 564–579. [Google Scholar] [CrossRef]
- Naghieh, S.; Sarker, M.D.; Karamooz-Ravari, M.R.; McInnes, A.D.; Chen, X. Modelling of the Mechanical Behaviour of 3D Bioplotted Scaffolds Considering the Penetration in Interlocked Strands. Appl. Sci. 2018, 8, 1422. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, L.; Yao, R.; Zhao, Y.; Sun, W. Effect of Bioink Properties on Printability and Cell Viability for 3D Bioplotting of Embryonic Stem Cells. Biofabrication 2016, 8, 035020. [Google Scholar] [CrossRef]
- Rajaram, A.; Schreyer, D.; Chen, D. Bioplotting Alginate/Hyaluronic Acid Hydrogel Scaffolds with Structural Integrity and Preserved Schwann Cell Viability. 3D Print Addit. Manuf. 2014, 1, 194–203. [Google Scholar] [CrossRef]
- Landers, R.; Mülhaupt, R. Desktop Manufacturing of Complex Objects, Prototypes and Biomedical Scaffolds by Means of Computer-Assisted Design Combined with Computer-Guided 3D Plotting of Polymers and Reactive Oligomers. Macromol. Mater. Eng. 2000, 282, 17–21. [Google Scholar] [CrossRef]
- Kim, M.K.; Jeong, W.; Lee, S.M.; Kim, J.B.; Jin, S.; Kang, H.W. Decellularized Extracellular Matrix-Based Bio-Ink with Enhanced 3D Printability and Mechanical Properties. Biofabrication 2020, 12, 025003. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hong, J.; Kim, W.J.; Kim, G.H. Bone-Derived DECM/Alginate Bioink for Fabricating a 3D Cell-Laden Mesh Structure for Bone Tissue Engineering. Carbohydr. Polym. 2020, 250, 116914. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Wang, Y.; Li, Y.; Juengpanich, S.; Li, W.; Chen, M.; Yin, J.; Fu, J.; Cai, X. Fabrication of Liver Microtissue with Liver Decellularized Extracellular Matrix (DECM) Bioink by Digital Light Processing (DLP) Bioprinting. Mater. Sci. Eng. C 2020, 109, 110625. [Google Scholar] [CrossRef] [PubMed]
- Elomaa, L.; Keshi, E.; Sauer, I.M.; Weinhart, M. Development of GelMA/PCL and DECM/PCL Resins for 3D Printing of Acellular in Vitro Tissue Scaffolds by Stereolithography. Mater. Sci. Eng. C 2020, 112, 110958. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Guo, Y.; Zhu, J.; Liu, F.; Xue, Y.; Huang, Y.; Zhu, B.; Wu, D.; Pan, H.; Gong, T.; et al. Hyaluronic Acid Methacrylate/Pancreatic Extracellular Matrix as a Potential 3D Printing Bioink for Constructing Islet Organoids. Acta Biomater. 2022. [Google Scholar] [CrossRef]
- Behre, A.; Tashman, J.W.; Dikyol, C.; Shiwarski, D.J.; Crum, R.J.; Johnson, S.A.; Kommeri, R.; Hussey, G.S.; Badylak, S.F.; Feinberg, A.W. 3D Bioprinted Patient-Specific Extracellular Matrix Scaffolds for Soft Tissue Defects. Adv. Healthc Mater. 2022, 2200866. [Google Scholar] [CrossRef]
- Chen, L.; Li, Z.; Zheng, Y.; Zhou, F.; Zhao, J.; Zhai, Q.; Zhang, Z.; Liu, T.; Chen, Y.; Qi, S. 3D-Printed Dermis-Specific Extracellular Matrix Mitigates Scar Contraction via Inducing Early Angiogenesis and Macrophage M2 Polarization. Bioact. Mater. 2022, 10, 236–246. [Google Scholar] [CrossRef]
- Kim, B.S.; Kwon, Y.W.; Kong, J.S.; Park, G.T.; Gao, G.; Han, W.; Kim, M.B.; Lee, H.; Kim, J.H.; Cho, D.W. 3D Cell Printing of in Vitro Stabilized Skin Model and in Vivo Pre-Vascularized Skin Patch Using Tissue-Specific Extracellular Matrix Bioink: A Step towards Advanced Skin Tissue Engineering. Biomaterials 2018, 168, 38–53. [Google Scholar] [CrossRef]
- de Sousa Iwamoto, L.A.; Duailibi, M.T.; Iwamoto, G.Y.; de Oliveira, D.C.; Duailibi, S.E. Evaluation of Ethylene Oxide, Gamma Radiation, Dry Heat and Autoclave Sterilization Processes on Extracellular Matrix of Biomaterial Dental Scaffolds. Sci. Rep. 2022, 12, 4299. [Google Scholar] [CrossRef]
- Łabuś, W.; Kitala, D.; Klama-Baryła, A.; Szapski, M.; Kraut, M.; Smętek, W.; Glik, J.; Kucharzewski, M.; Rojczyk, E.; Utrata-Wesołek, A.; et al. Influence of Electron Beam Irradiation on Extracellular Matrix of the Human Allogeneic Skin Grafts. J. Biomed. Mater. Res. B Appl. Biomater. 2022, 110, 547–563. [Google Scholar] [CrossRef]
- Balestrini, J.L.; Liu, A.; Gard, A.L.; Huie, J.; Blatt, K.M.S.; Schwan, J.; Zhao, L.; Broekelmann, T.J.; Mecham, R.P.; Wilcox, E.C.; et al. Sterilization of Lung Matrices by Supercritical Carbon Dioxide. Tissue Eng. Part C Methods 2015, 22, 260–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smoulder, A.; Castleton, A.; Keane, T.; White, L.; Zhang, L.; Badylak, S.F. The Effects of Terminal Sterilization on the Mechanical Properties of Extracellular Matrix Hydrogels; 2015. [Google Scholar]
- Kolios, G.; Moodley, Y. Introduction to Stem Cells and Regenerative Medicine. Respiration 2013, 85, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Huang, L.; Yu, S.; Zheng, J.; Wang, H.; Zhang, Y. Decellularized Tongue Tissue as an in Vitro Model for Studying Tongue Cancer and Tongue Regeneration. Acta Biomater. 2017, 58, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Alabi, B.R.; LaRanger, R.; Shay, J.W. Decellularized Mice Colons as Models to Study the Contribution of the Extracellular Matrix to Cell Behaviour and Colon Cancer Progression. Acta Biomater. 2019, 100, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Kort-Mascort, J.; Bao, G.; Elkashty, O.; Flores-Torres, S.; Munguia-Lopez, J.G.; Jiang, T.; Ehrlicher, A.J.; Mongeau, L.; Tran, S.D.; Kinsella, J.M. Decellularized Extracellular Matrix Composite Hydrogel Bioinks for the Development of 3D Bioprinted Head and Neck in Vitro Tumour Models. ACS Biomater. Sci. Eng. 2021, 7, 5288–5300. [Google Scholar] [CrossRef]
- Bortot, B.; Apollonio, M.; Rampazzo, E.; Valle, F.; Brucale, M.; Ridolfi, A.; Ura, B.; Addobbati, R.; Di Lorenzo, G.; Romano, F.; et al. Small Extracellular Vesicles from Malignant Ascites of Patients with Advanced Ovarian Cancer Provide Insights into the Dynamics of the Extracellular Matrix. Mol. Oncol. 2021, 15, 3596–3614. [Google Scholar] [CrossRef]
- Ferreira, L.P.; Gaspar, V.M.; Mendes, L.; Duarte, I.F.; Mano, J.F. Organotypic 3D Decellularized Matrix Tumour Spheroids for High-Throughput Drug Screening. Biomaterials 2021, 275, 120983. [Google Scholar] [CrossRef]
- Muffly, T.M.; Tizzano, A.P.; Walters, M.D. The History and Evolution of Sutures in Pelvic Surgery. J. R. Soc. Med. 2011, 104, 107–112. [Google Scholar] [CrossRef]
- Snyder, C.C. On the History of the Suture. Plast. Reconstr. Surg. 1976, 58, 401–406. [Google Scholar] [CrossRef]
- Badylak, S.F.; Hoppo, T.; Nieponice, A.; Gilbert, T.W.; Davison, J.M.; Jobe, B.A. Esophageal Preservation in Five Male Patients after Endoscopic Inner-Layer Circumferential Resection in the Setting of Superficial Cancer: A Regenerative Medicine Approach with a Biologic Scaffold. Tissue Eng. Part A 2011, 17, 1643–1650. [Google Scholar] [CrossRef]
- Mase, V.J.J.; Hsu, J.R.; Wolf, S.E.; Wenke, J.C.; Baer, D.G.; Owens, J.; Badylak, S.F.; Walters, T.J. Clinical Application of an Acellular Biologic Scaffold for Surgical Repair of a Large, Traumatic Quadriceps Femoris Muscle Defect. Orthopedics 2010, 33, 511. [Google Scholar] [CrossRef] [PubMed]
- Dziki, J.; Badylak, S.F.; Yabroudi, M.; Sicari, B.; Ambrosio, F.; Stearns, K.; Turner, N.; Wyse, A.; Boninger, M.L.; Brown, E.H.P.; et al. An Acellular Biologic Scaffold Treatment for Volumetric Muscle Loss: Results of a 13-Patient Cohort Study. NPJ Regen. Med. 2016, 1, 16008. [Google Scholar] [CrossRef] [Green Version]
- Raya-Rivera, A.M.; Esquiliano, D.; Fierro-Pastrana, R.; López-Bayghen, E.; Valencia, P.; Ordorica-Flores, R.; Soker, S.; Yoo, J.J.; Atala, A. Tissue-Engineered Autologous Vaginal Organs in Patients: A Pilot Cohort Study. Lancet 2014, 384, 329–336. [Google Scholar] [CrossRef]
- ACOG Committee Opinion. No. 728: Müllerian Agenesis Diagnosis, Management, And Treatment. Obstet. Gynecol. 2018, 131, e35–e42. [CrossRef]
- Cairo, S.B.; Tabak, B.; Harmon, C.M.; Bass, K.D. Novel Use of Porcine Extracellular Matrix in Recurrent Stricture Following Repair of Tracheoesophageal Fistula. Pediatr. Surg. Int. 2017, 33, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Ansari, S.; Karram, M. Two Cases of Female Urethral Reconstruction with Acellular Porcine Urinary Bladder Matrix. Int. Urogynecol. J. 2017, 28, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Kesavan, R.; Sheela Sasikumar, C.; Narayanamurthy, V.B.; Rajagopalan, A.; Kim, J. Management of Diabetic Foot Ulcer with MA–ECM (Minimally Manipulated Autologous Extracellular Matrix) Using 3D Bioprinting Technology—An Innovative Approach. Int. J. Low. Extrem. Wounds 2021. [Google Scholar] [CrossRef]
- Cua, C.L.; Johnston, P.; Harmon, J.; Shelton, K.; McConnell, P.I. Custom Extracellular Matrix Cylinder Mitral Valve in a Pediatric Patient. Echocardiography 2017, 34, 1956–1958. [Google Scholar] [CrossRef]
- Identifier: NCT03551509, LifeNet: Extracellular Matrix Graft in Rotator Cuff Repair. Available online: https://clinicaltrials.gov/ct2/show/NCT03551509 (accessed on 16 October 2022).
- Identifier: NCT01348581, Use of Fish Skin Extracellular Matrix (ECM) to Facilitate Chronic Wound Healing. Available online: https://clinicaltrials.gov/ct2/show/NCT01348581 (accessed on 16 October 2022).
- Identifier: NCT02845180, In-Vivo Assessment of Adipose Allograft Extracellular Matrix in Abdominoplasty Patients. Available online: https://clinicaltrials.gov/ct2/show/NCT02845180 (accessed on 16 October 2022).
- Identifier: NCT04011059, Randomized Study of Coronary Revascularization Surgery with Injection of WJ-MSCs and Placement of an Epicardial Extracellular Matrix (Scorem-Cells). Available online: https://clinicaltrials.gov/ct2/show/NCT04011059 (accessed on 16 October 2022).
- Identifier: NCT02750280, A Longitudinal Study to Evaluate an Extracellular Matrix (MatriStem®) for the Treatment of Diabetic Foot Ulcers (M-S-DFU-RCT). Available online: https://clinicaltrials.gov/ct2/show/NCT02750280 (accessed on 16 October 2022).
- Identifier: NCT02305602, A Study of VentriGel in Post-MI Patients. Available online: https://clinicaltrials.gov/ct2/show/NCT02305602 (accessed on 16 October 2022).
- Identifier: NCT05082831, Human ECM Implanted within Microfracture Interstices & the Cartilage Defect in the Knee to Regenerate Hyaline Cartilage. Available online: https://clinicaltrials.gov/ct2/show/NCT05082831 (accessed on 16 October 2022).
- Identifier: NCT02073331, Obtain Additional Information on Use of CorMatrix ECM (Extracellular Matrix) (RECON). Available online: https://clinicaltrials.gov/ct2/show/NCT02073331 (accessed on 16 October 2022).
- Identifier: NCT02530970, A Post Market Study on the Use of Cormatrix® Cangaroo ECM® (Extracellular Matrix) Envelope (SECURE). Available online: https://clinicaltrials.gov/ct2/show/NCT02530970 (accessed on 16 October 2022).
- Identifier: NCT01535807, CorMatrix ECM Study: To Identify Inflammatory Markers Following CABG with/without ECM (CorMatrix). Available online: https://clinicaltrials.gov/ct2/show/NCT01535807 (accessed on 16 October 2022).
- Identifier: NCT03632031, Treatment of Wounds Using Oasis® ECM. Available online: https://clinicaltrials.gov/ct2/show/NCT03632031 (accessed on 16 October 2022).
- Identifier: NCT02817984, Safety Study of Acellular Adipose Tissue for Soft Tissue Reconstruction. Available online: https://clinicaltrials.gov/ct2/show/NCT02817984 (accessed on 16 October 2022).
- Identifier: NCT03544632, Acellular Adipose Tissue (AAT) for Soft Tissue Reconstruction. Available online: https://clinicaltrials.gov/ct2/show/NCT03544632 (accessed on 16 October 2022).
- Identifier: NCT00614419, Prospective Randomized Trial Comparing Lichtenstein’s Repair of Inguinal Hernia with Polypropylene Mesh Versus Surgisis. Available online: https://clinicaltrials.gov/ct2/show/NCT00614419 (accessed on 16 October 2022).
- Identifier: NCT05243966, MyriadTM Augmented Soft Tissue Reconstruction Registry (MASTRR). Available online: https://clinicaltrials.gov/ct2/show/NCT05243966 (accessed on 16 October 2022).
- Identifier: NCT03283787, Comparing Concomitant Use of ACell MicroMatrix® and ACell CytalTM to Standard of Care in Stage 3 or 4 Pressure Injuries. Available online: https://clinicaltrials.gov/ct2/show/NCT03283787 (accessed on 16 October 2022).
- Identifier: NCT01292876, Musculotendinous Tissue Repair Unit and Reinforcement (MTURR). Available online: https://clinicaltrials.gov/ct2/show/NCT01292876 (accessed on 16 October 2022).
- Identifier: NCT01917591, Healing of Punch Graft Wounds. Comparative Study Between MariGen and Oasis: A Non-Inferiority Study. Available online: https://clinicaltrials.gov/ct2/show/NCT01917591 (accessed on 16 October 2022).


| Method | Types | Mechanism | References |
|---|---|---|---|
| Chemical | |||
| Detergents | |||
| Ionic | Solubilize cell and nucleic membranes, denature proteins, and remove cell debris; can disrupt ECM structure and remove desirable biological molecules (e.g., GAG, growth factors, etc.) | ||
| Sodium dodecyl sulfate | [82,87,88,90,93,109,110,111,112,114,115,116,117,118,128,134,135,136,137,143,144,146,148,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177,178] | ||
| Sodium deoxycholate | [56,113,123,138,151,179,180,181,182,183,184,185,186,187] | ||
| Sodium lauryl ether sulfate | [127,149,188,189] | ||
| Sodium lauroyl sarcosinate | [125] | ||
| Potassium laurate | [190] | ||
| Triton X-200 | [172,191,192] | ||
| Nonionic | Disrupt DNA, lipid, and protein interactions; renatures proteins; less effective than SDS | ||
| Triton X-100 | [14,23,82,83,86,88,90,96,97,107,109,110,113,114,115,117,123,133,134,135,136,137,138,139,144,151,152,153,154,155,156,157,158,159,160,161,162,173,174,177,178,179,180,181,182,183,186,187,193,194,195,196,197,198,199,200,201,202,203,204,205] | ||
| Zwitterionic | Exhibit properties of non-ionic and ionic detergents; minimal disruption of ECM structure | ||
| CHAPS | [198] | ||
| Sulfobetaine-10 and -16 | [191,192] | ||
| Amidosulfobetaine-14 | [167,206] | ||
| Acid/Base | |||
| Acids | Denature proteins, solubilize the cytoplastic contents of the cell, and degrade genetic material | ||
| Peracetic Acid | [131,173,207] | ||
| Base | |||
| Sodium hydroxide | [208] | ||
| Ammonium hydroxide | [14,90,139,155,172,176,204,205,209] | ||
| Buffer | |||
| Tris | Extract soluble cytoplasmic proteins, increase permeability of cell membranes, and degrade genetic material | [20,23,83,97,116,128,151,158,166,167,194,197,199,200,202,210] | |
| Other | |||
| Hypotonic/hypertonic | |||
| Sodium chloride | Osmotic shock that can cause lysis or shrinking of cells | [143,172,180,185,199] | |
| Potassium iodide | [134,206] | ||
| Potassium chloride | [134,206] | ||
| Chelating | |||
| EDTA, EGTA | Bind to metal ions that have the potential to interfere with the activity of enzymes used in decellularization, disrupt cell adhesion to the ECM, and inhibit metalloproteases | [14,20,23,83,84,86,91,107,111,112,116,128,131,133,134,135,137,138,143,154,155,166,172,186,187,194,195,196,197,198,199,200,202,211] | |
| Penetration enhancement | |||
| DMSO | Protect ECM structure, increase the penetration of detergents, and shorten decellularization time | [151,166] | |
| Biological | |||
| Enzymes | |||
| DNase | Degrade genetic material through hydrolysis, leading to a reduction in the fragment size | [56,82,83,86,88,93,96,112,113,115,116,128,132,134,139,154,156,158,166,170,172,175,179,180,181,183,184,194,198,201,204,205,210,212] | |
| RNase | [56,82,112,132,156,158,179,180,181,183,194,204,205,210,212] | ||
| Benzoase | [123] | ||
| Trypsin | Digest proteins and help to disrupt cell attachment to the ECM | [23,107,112,115,135,138,155,186,187,196,197,202,210,212,213] | |
| Non-enzymatic agents | |||
| PMSF | Inhibit proteases that are released during cell lysis that have the potential to damage the ECM | [20,86,158,199] | |
| Physical | |||
| Freezing | Disrupt cell membranes due to ice crystal formation and expansion | [20,56,82,84,86,87,88,93,109,111,112,113,116,125,132,133,134,135,137,153,154,155,163,169,179,180,182,183,197,199,200,210,211,214,215] | |
| Heating | Denature cell proteins, inactivate enzymes, disrupt cell membranes | [103] | |
| Sonication | Aid in cell lysis and removal of cell debris | [216,217] | |
| Electroporation | Disrupt cell membranes | [218,219,220,221,222] | |
| Vacuum | Cause cell lysis, improves penetration of decellularization solutions | [118,146,180,183] | |
| Mechanical | Physically remove unwanted tissue layers to allow more effective decellularization | [23,92,158,195,200,223,224,225,226,227] | |
| Techniques of applying agents | |||
| Perfusion | Thoroughly deliver decellularization fluid to all parts of a tissue | [90,117,123,135,137,138,139,144,148,152,153,157,164,165,169,193,214,228,229] | |
| Agitation | Ensure adequate mixing of solutions used in decellularization and aid in cell lysis and removal of cell debris | [20,23,86,88,116,125,127,128,130,133,134,146,153,158,167,180,181,182,194,196,200,210,223,227,230] | |
| Supercritical fluid | Disrupt the cell membrane and cause cell lysis with a minimal effect on the ECM and ECM components | [201,231,232] | |
| Method | Purpose | Results | References |
|---|---|---|---|
| Vacuum | Enhanced penetration of decellularizing solution and clearance to cell debris | Effective at removing DNA, MHC-1, and other cellular content Faster and more efficient decellularization Minimal effect on collagen, GAG content, and biomechanical properties Potential to damage ECM microstructures in weaker tissues | [118,146,180,183,263] |
| Hydrostatic washing | Enhanced penetration of dense, fibrous tissues | Faster and more efficient decellularization | [264] |
| Pulsatile perfusion | More closely mimics in natura state of tissue perfusion | More profound decellularization More homogeneous decellularization Lower residual DNA content Little to no difference in collagen and GAG content | [165,245] |
| Chemical penetration enhancement | Improve solubility and penetration of detergents and disrupt cell membranes | Faster and more efficient decellularization Reduced DNA content Better preserved and protected GAG, elastin, and collagen | [166] |
| Sonication | Disrupt cell membranes due to cavitation | Minimal effect on the fibrous structure of ECM More effective at removing cells Greatly reduced the decellularization time | [216,217] |
| Nonthermal irreversible electroporation (NTIRE) | Cause irreversible damage to the cell membrane | No effect on the ECM structure Causes tissue disruption, cell delamination, and cell death | [165] |
| Decellularizing agents | Different levels of reactivity with biological molecules in addition to disruption of cell membranes and clearance of cellular debris | Potassium laurate Better retention of ECM compounds Better-preserved architecture Increased cell viability and proliferation in vitro Lower inflammatory response and better cell distribution in vivo | [127,149,188,190,208] |
| Sodium lauryl ether sulfate Better GAG and collagen retention Better preserved microarchitecture Lower inflammatory response and platelet adhesion in vivo Better host cell migration into scaffold Slower at decellularizing | |||
| NaOH As effective as detergents Similar collagen, GAG, and adhesion protein retention Better DNA clearance Equal to detergents for in vivo cell migration | |||
| Supercritical fluids | Enhance penetration of cell membrane and clearance of cell debris | Comparable to detergents in cell and DNA removal Superior retention of GAG, soluble collagen, adhesion proteins, and angiogenic factors Need to add collagen to gel for gelation to occur Superior neovascularization No difference in immune response | [201,232] |
| Alternating decellularizing solutions | Minimize exposure time to detergents in combination with hypertonic and hypotonic shock to cells, clearance of detergents, and clearance of cell debris | Slower decellularization Equal DNA removal Superior retention of GAG and soluble collagen Similar retention of growth factors Higher cell viability and gene expression profiles in vitro | [228] |
| Method | Pros | Cons | References |
|---|---|---|---|
| Peracetic acid and ethanol (together or separately) | Minimal effect on structural and biological properties of ECM Peracetic acid is a strong oxidizer with effective bactericidal, viricidal, fungicidal, and sporicidal properties Can be used to decellularize tissues in combination | Can be difficult to remove Residual chemicals can negatively affect cell viability | [23,83,92,93,109,114,135,136,138,144,155,158,159,161,162,169,179,182,187,189,200,211,223,225,226,227,230] |
| Ethylene oxide | Effective at sterilizing biomaterials | Residual gas can negatively affect cell viability Minimally effective in hydrogels Can affect the mechanical properties of the final hydrogel | [195,224,236,347] |
| UV | Can improve mechanical properties due to cross-linking | Harmful to cells Can cause premature gelation of dECM hydrogels | [118,133,153,169,211] |
| Antimicrobials | Can be added to culture media Can be used during decellularization to limit contamination during processing | Can be difficult to remove Can affect cell behaviour | [112,113,116,117,127,133,135,137,144,148,152,154,156,164,166,167,168,179,184,193,214] |
| γ radiation | Highly effective in reducing bioburden No effect on cell response | High levels of exposure can prevent gelation of dECM hydrogels | [158,194,195,347] |
| Electron beam (β radiation) | Highly effective in reducing bioburden | High levels of exposure can prevent gelation of dECM hydrogels | [195,348] |
| Sterile filtration | Can be used to filter ECM extracts | Loss of proteins in high concentration dECM solutions and colloidal dispersions | [86,91] |
| Supercritical CO2 | Highly effective at reducing bioburden Can be combined with other sterilizing agents (e.g., ethanol, peracetic acid) | Can affect the mechanical properties of the final hydrogel | [195,201,231,349,350] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McInnes, A.D.; Moser, M.A.J.; Chen, X. Preparation and Use of Decellularized Extracellular Matrix for Tissue Engineering. J. Funct. Biomater. 2022, 13, 240. https://doi.org/10.3390/jfb13040240
McInnes AD, Moser MAJ, Chen X. Preparation and Use of Decellularized Extracellular Matrix for Tissue Engineering. Journal of Functional Biomaterials. 2022; 13(4):240. https://doi.org/10.3390/jfb13040240
Chicago/Turabian StyleMcInnes, Adam D., Michael A. J. Moser, and Xiongbiao Chen. 2022. "Preparation and Use of Decellularized Extracellular Matrix for Tissue Engineering" Journal of Functional Biomaterials 13, no. 4: 240. https://doi.org/10.3390/jfb13040240
APA StyleMcInnes, A. D., Moser, M. A. J., & Chen, X. (2022). Preparation and Use of Decellularized Extracellular Matrix for Tissue Engineering. Journal of Functional Biomaterials, 13(4), 240. https://doi.org/10.3390/jfb13040240