

Cell Instructive Behavior of Composite Scaffolds in a Co-Culture of Human Mesenchymal Stem Cells and Peripheral Blood Mononuclear Cells

 ,
,  , ,
, ,  , , ,
, , ,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Scaffolds
2.2. Cell Culture
2.3. Immunophenotypic Characterization of hBM-MSCs and hPBMCs
2.4. Co-Culture of Human Bone Marrow Mesenchymal Stem Cells (hBM-MSCs) and Peripheral Blood Mononuclear Cells (hPBMCs)
2.5. Cell Viability and Proliferation
2.5.1. Cell Viability Assessment
2.5.2. Adhesion and Morphology of hBM-MSCs and hPBMCs
2.6. Cell Differentiation via Biochemical Analysis
2.6.1. Alkaline Phosphatase (ALP) Activity
2.6.2. Tartrate-Resistant Acid Phosphatase (TRAP) Activity
2.7. Expression of ALP and Visualization by Means of Confocal Laser Microscopy (CLSM)
2.8. Expression of TRAP and Visualization Microscopically
2.9. Cell Differentiation via Real-Time Polymerase Chain Reaction
2.10. Statistical Analysis
3. Results
3.1. Scaffolds Characterization
3.2. Characterization of hBM-MSCs and hPBMCs
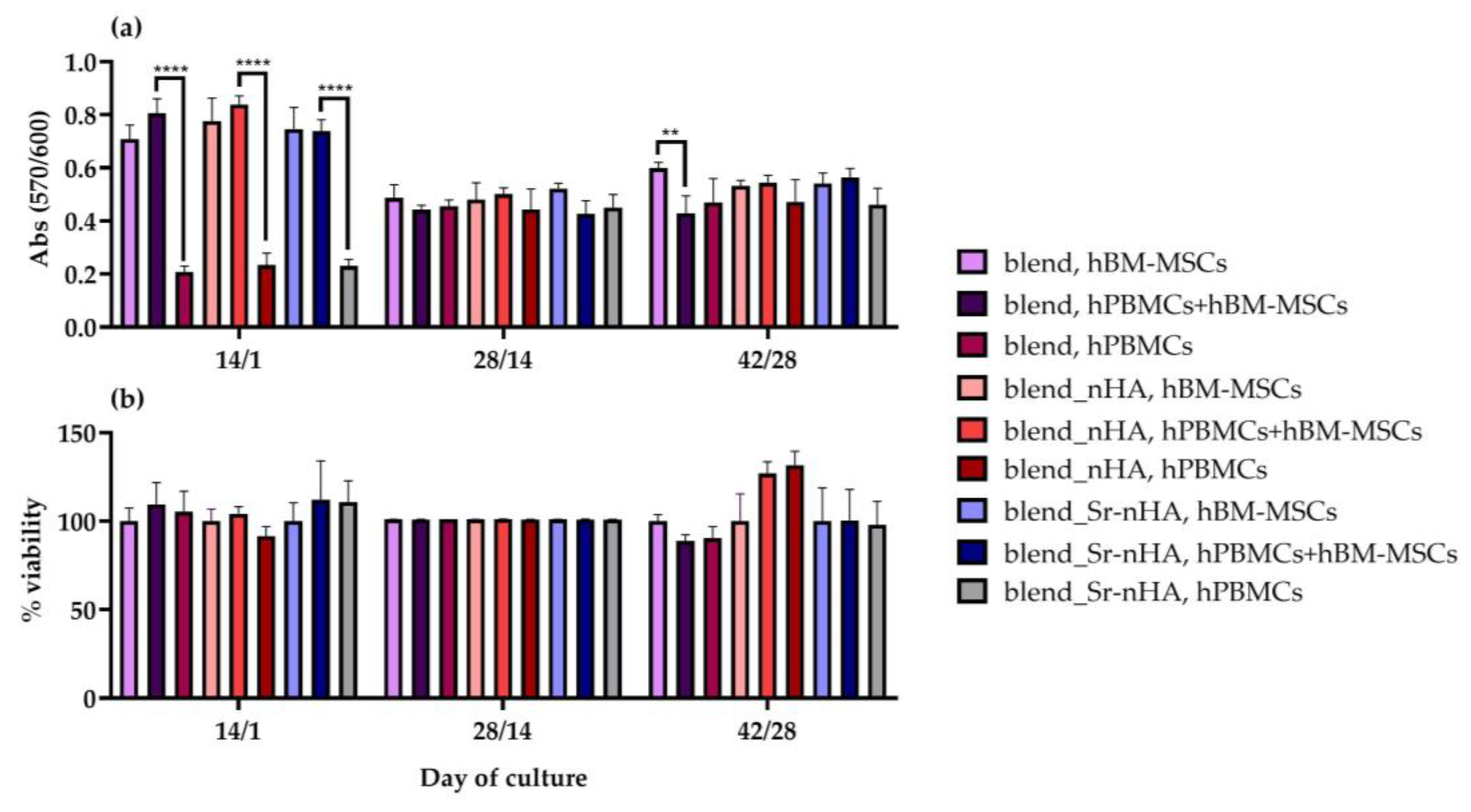
3.3. Cell Viability Assessment within the Scaffolds
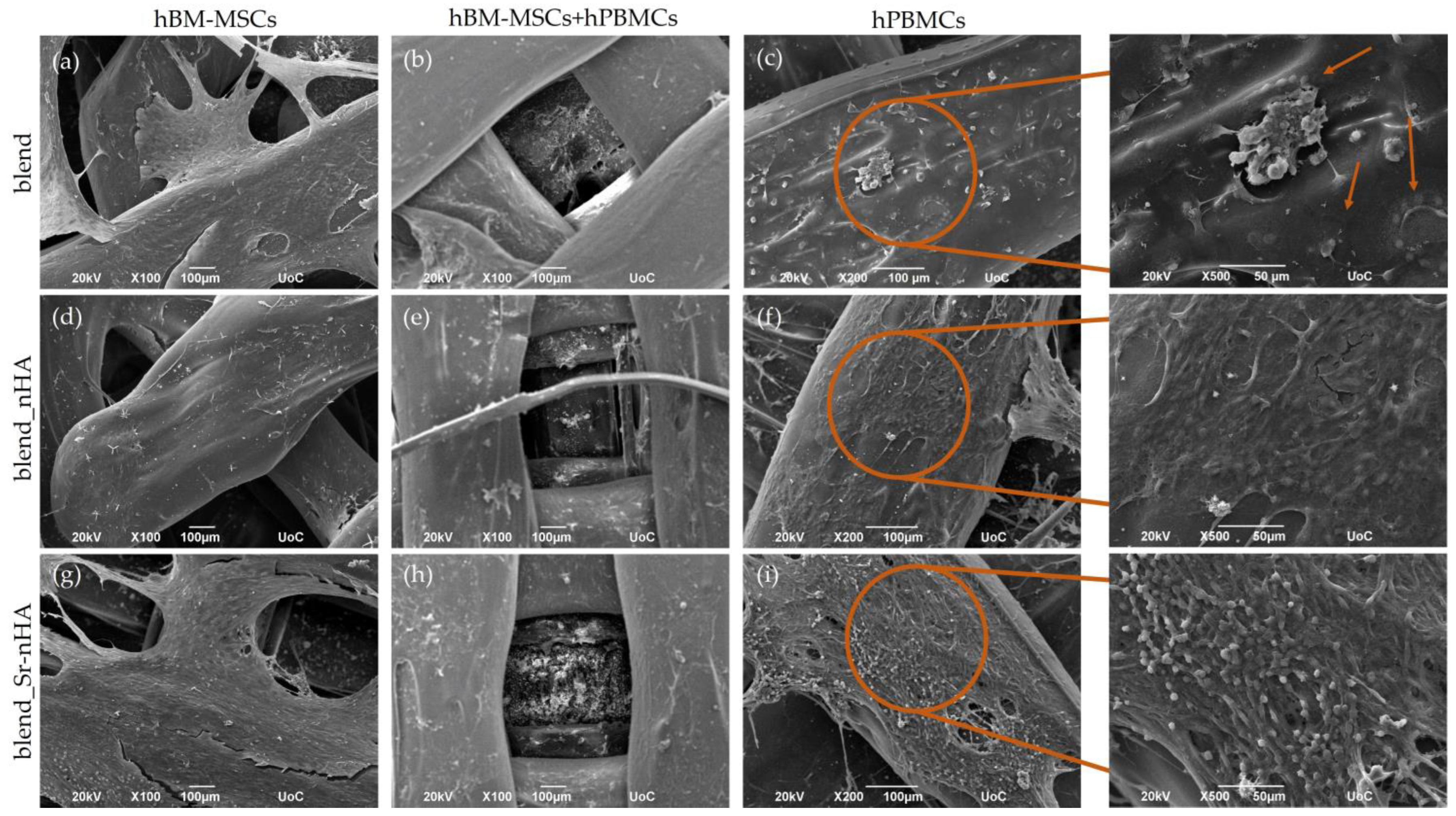
3.4. Cell Adhesion and Morphology
3.5. Biochemical Determination of ALP and TRAP Activity
3.6. Expression of ALP and Visualization by Means of CLSM
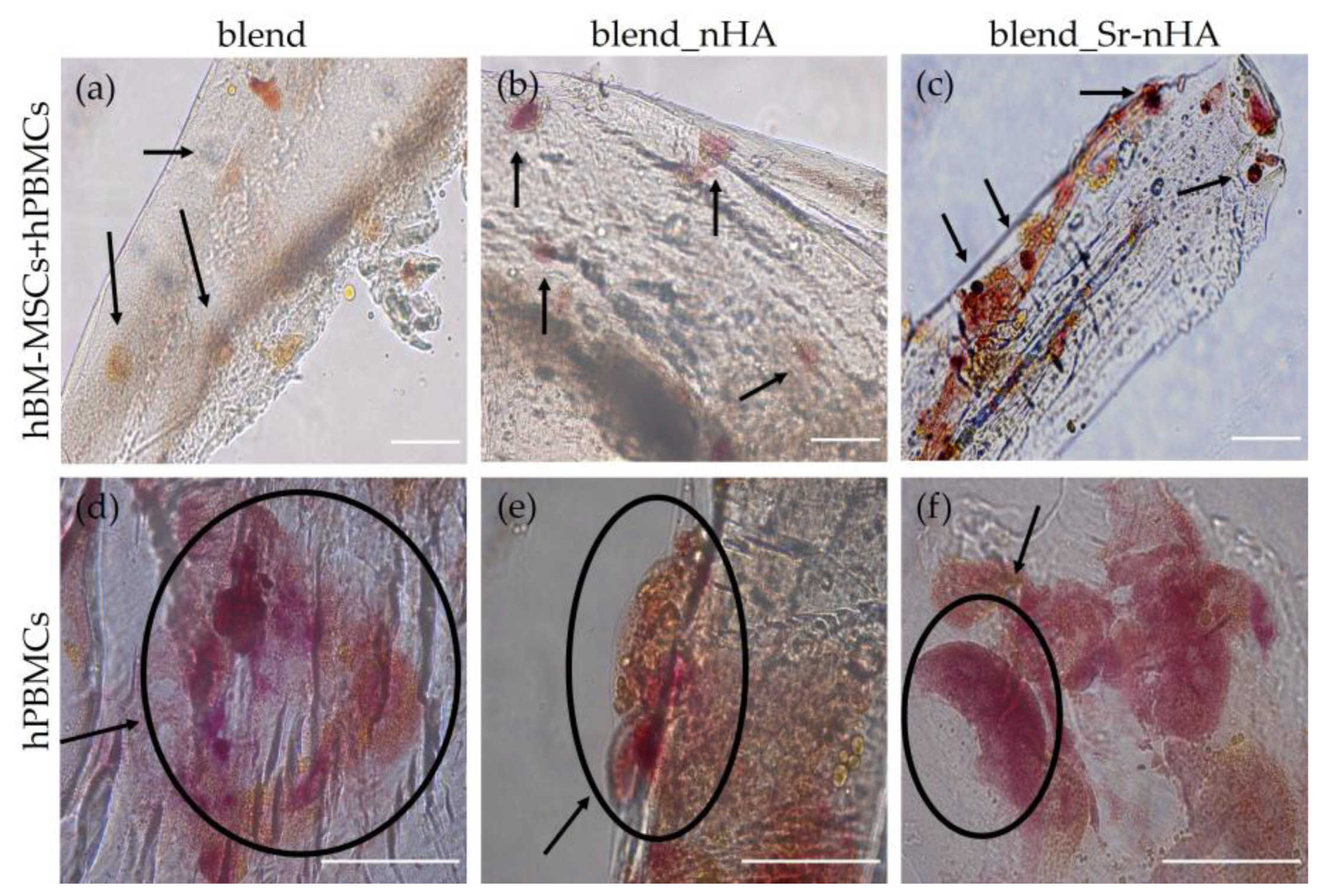
3.7. TRAP Expression and Visualization
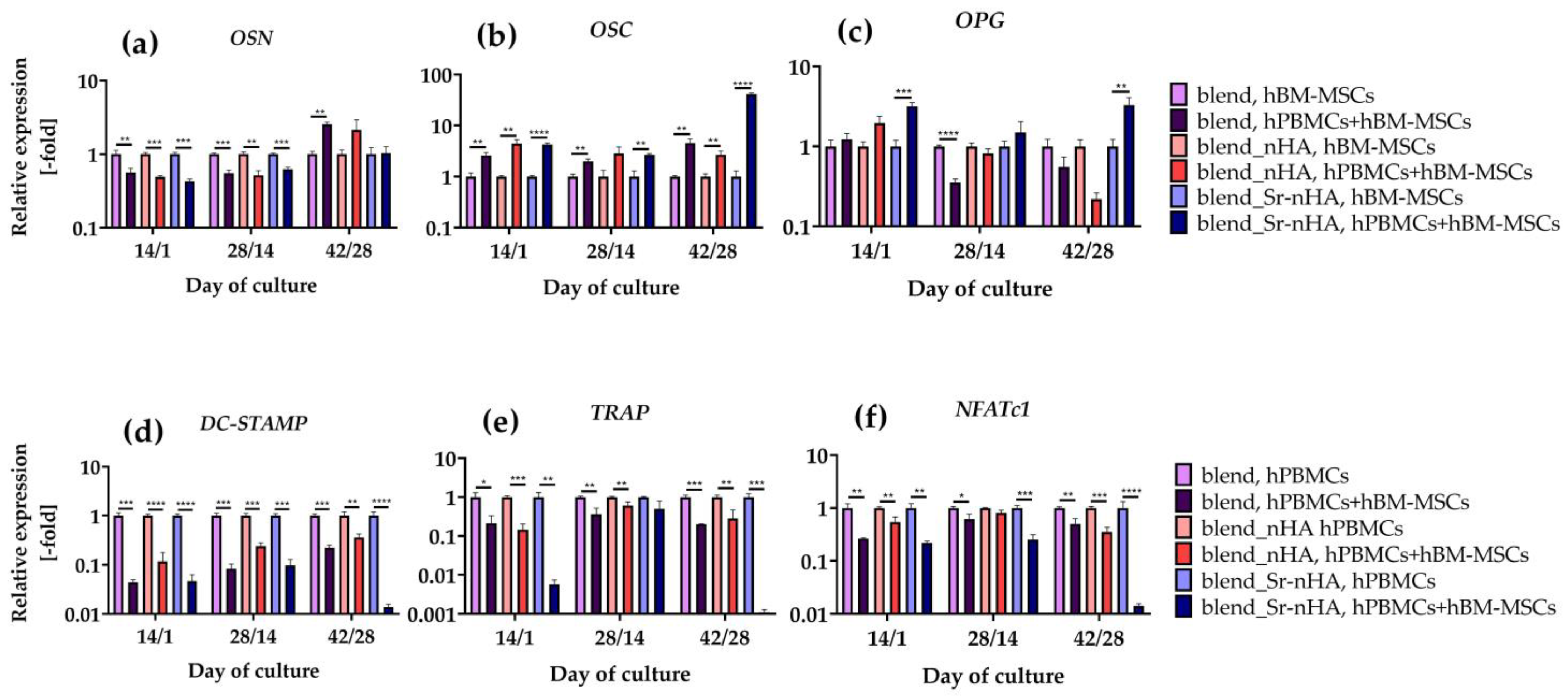
3.8. Analysis of Osteogenic and Osteoclastogenic Gene Expression in Culture with Pure and Substituted Polymeric Materials
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berner, A.; Reichert, J.C.; Müller, M.B.; Zellner, J.; Pfeifer, C.; Dienstknecht, T.; Nerlich, M.; Sommerville, S.; Dickinson, I.C.; Schütz, M.A.; et al. Treatment of long bone defects and non-unions: From research to clinical practice. Cell Tissue Res. 2012, 347, 501–519. [Google Scholar] [CrossRef] [PubMed]
- Moshiri, A.; Oryan, A. Role of tissue engineering in tendon reconstructive surgery and regenerative medicine: Current concepts, approaches and concerns. Hard Tissue 2012, 1, 11. [Google Scholar] [CrossRef]
- Rao, S.H.; Harini, B.S.; Shadamarshan, R.P.K.; Balagangadharan, K.; Selvamurugan, N. Natural and synthetic polymers/bioceramics/bioactive compounds-mediated cell signalling in bone tissue engineering. Int. J. Biol. Macromol. 2017, 110, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, D.; Zhao, X.; Pakvasa, M.; Tucker, A.B.; Luo, H.; Qin, K.H.; Hu, D.A.; Wang, E.J.; Li, A.J.; et al. Stem cell-friendly scaffold biomaterials: Applications for bone tissue engineering and regenerative medicine. Front. Bioeng. Biotechnol. 2020, 8, 598607. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaz, A.; Jayasuriya, A.C. Recent trends in the application of widely used natural and synthetic polymer nanocomposites in bone tissue regeneration. Mater. Sci. Eng. C 2020, 110, 110698. [Google Scholar] [CrossRef] [PubMed]
- Georgopoulou, A.; Kaliva, M.; Vamvakaki, M.; Chatzinikolaidou, M. Osteogenic potential of pre-osteoblastic cells on a chitosan-graft-polycaprolactone copolymer. Materials 2018, 11, 490. [Google Scholar] [CrossRef] [PubMed]
- Butscher, A.; Bohner, M.; Hofmann, S.; Gauckler, L.; Müller, R. Structural and material approaches to bone tissue engineering in powder-based three-dimensional printing. Acta Biomater. 2011, 7, 907–920. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, O.A.; Masood, S.H.; Bhowmik, J.L. Optimization of fused deposition modeling process parameters: A review of current research and future prospects. Adv. Manuf. 2015, 3, 42–53. [Google Scholar] [CrossRef]
- Calore, A.R.; Sinha, R.; Harings, J.; Bernaerts, K.V.; Mota, C.; Moroni, L. Additive manufacturing using melt extruded thermoplastics for tissue engineering. Methods Mol. Biol. 2021, 2147, 75–99. [Google Scholar] [PubMed]
- Seyednejad, H.; Ghassemi, A.H.; van Nostrum, C.F.; Vermonden, T.; Hennink, W.E. Functional aliphatic polyesters for biomedical and pharmaceutical applications. J. Control. Release Off. J. Control. Release Soc. 2011, 152, 168–176. [Google Scholar] [CrossRef] [PubMed]
- da Silva, D.; Kaduri, M.; Poley, M.; Adir, O.; Krinsky, N.; Shainsky-Roitman, J.; Schroeder, A. Biocompatibility, biodegradation and excretion of polylactic acid (pla) in medical implants and theranostic systems. Chem. Eng. J. 2018, 340, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Komal, U.K.; Lila, M.K.; Singh, I. Pla/banana fiber based sustainable biocomposites: A manufacturing perspective. Compos. Part B Eng. 2020, 180, 107535. [Google Scholar] [CrossRef]
- Mazzanti, V.; Malagutti, L.; Mollica, F. Fdm 3d printing of polymers containing natural fillers: A review of their mechanical properties. Polymers 2019, 11, 1094. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wang, W.; Bartolo, P. Investigation of polycaprolactone for bone tissue engineering scaffolds: In vitro degradation and biological studies. Mater. Des. 2022, 216, 110582. [Google Scholar] [CrossRef]
- Doyle, C.; Tanner, E.T.; Bonfield, W. In vitro and in vivo evaluation of polyhydroxybutyrate and of polyhydroxybutyrate reinforced with hydroxyapatite. Biomaterials 1991, 12, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Briso, A.L.; Serrano-Aroca, Á. Poly(3-hydroxybutyrate-co-3-hydroxyvalerate): Enhancement strategies for advanced applications. Polymers 2018, 10, 732. [Google Scholar] [CrossRef] [PubMed]
- Hassanajili, S.; Karami-Pour, A.; Oryan, A.; Talaei-Khozani, T. Preparation and characterization of pla/pcl/ha composite scaffolds using indirect 3d printing for bone tissue engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 104, 109960. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Fu, S.; Zhu, Y. 3d printing of ceramic-based scaffolds for bone tissue engineering: An overview. J. Mater. Chem. B 2018, 6, 4397–4412. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yuan, Z.; Huang, J.J.M.T. Substituted hydroxyapatite: A recent development. Mater. Technol. 2020, 35, 785–796. [Google Scholar] [CrossRef]
- Kothapalli, C.R.; Shaw, M.T.; Wei, M. Biodegradable ha-pla 3-d porous scaffolds: Effect of nano-sized filler content on scaffold properties. Acta Biomater. 2005, 1, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Jaidev, L.R.; Chatterjee, K. Surface functionalization of 3d printed polymer scaffolds to augment stem cell response. Mater. Des. 2019, 161, 44–54. [Google Scholar] [CrossRef]
- Brown, E.M. Is the calcium receptor a molecular target for the actions of strontium on bone? Osteoporos. Int. 2003, 14, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lu, X.; Meng, Y.; Weyant, C.M. Comparison study of biomimetic strontium-doped calcium phosphate coatings by electrochemical deposition and air plasma spray: Morphology, composition and bioactive performance. J. Mater. Sci. Mater. Med. 2012, 23, 2359–2368. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, J.; Christoffersen, M.R.; Kolthoff, N.; Bärenholdt, O. Effects of strontium ions on growth and dissolution of hydroxyapatite and on bone mineral detection. Bone 1997, 20, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Li, Z.; Zhong, X.; Cai, Z.; Ning, Z.; Hou, T.; Xiong, L.; Feng, Y.; Leung, F.; Lu, W.W.; et al. Strontium inhibits osteoclastogenesis by enhancing lrp6 and β-catenin-mediated opg targeted by mir-181d-5p. J. Cell Commun. Signal. 2019, 13, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, C.J. Developing cellular systems in vitro to simulate regeneration. Tissue Eng. Part A 2014, 20, 1355–1357. [Google Scholar] [CrossRef] [PubMed]
- Borciani, G.; Montalbano, G.; Baldini, N.; Cerqueni, G.; Vitale-Brovarone, C.; Ciapetti, G. Co-culture systems of osteoblasts and osteoclasts: Simulating in vitro bone remodeling in regenerative approaches. Acta Biomater. 2020, 108, 22–45. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, A.; Thieme, S.; Domaschke, H.; Springer, A.; Rösen-Wolff, A.; Gelinsky, M. Crosstalk of osteoblast and osteoclast precursors on mineralized collagen—Towards an in vitro model for bone remodeling. J. Biomed. Mater. Res. Part A 2010, 95, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.L.; Motta, A.; Marshall, M.J.; El Haj, A.J.; Cartmell, S.H. Osteoblast: Osteoclast co-cultures on silk fibroin, chitosan and plla films. Biomaterials 2009, 30, 5376–5384. [Google Scholar] [CrossRef] [PubMed]
- Tsuneto, M.; Yamazaki, H.; Yoshino, M.; Yamada, T.; Hayashi, S.-I. Ascorbic acid promotes osteoclastogenesis from embryonic stem cells. Biochem. Biophys. Res. Commun. 2005, 335, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Hozumi, A.; Osaki, M.; Goto, H.; Sakamoto, K.; Inokuchi, S.; Shindo, H. Bone marrow adipocytes support dexamethasone-induced osteoclast differentiation. Biochem. Biophys. Res. Commun. 2009, 382, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Noh, A.L.; Yim, M. Beta-glycerophosphate accelerates rankl-induced osteoclast formation in the presence of ascorbic acid. Die Pharm. 2011, 66, 195–200. [Google Scholar]
- Naseem, R.; Montalbano, G.; German, M.J.; Ferreira, A.M.; Gentile, P.; Dalgarno, K. Influence of pcl and phbv on plla thermal and mechanical properties in binary and ternary polymer blends. Molecules 2022, 27, 7633. [Google Scholar] [CrossRef] [PubMed]
- Kontogianni, G.-I.; Bonatti, A.F.; De Maria, C.; Naseem, R.; Melo, P.; Coelho, C.; Vozzi, G.; Dalgarno, K.; Quadros, P.; Vitale-Brovarone, C.; et al. Promotion of in vitro osteogenic activity by melt extrusion-based plla/pcl/phbv scaffolds enriched with nano-hydroxyapatite and strontium substituted nano-hydroxyapatite. Polymers 2023, 15, 1052. [Google Scholar] [CrossRef] [PubMed]
- Melo, P.; Naseem, R.; Corvaglia, I.; Montalbano, G.; Pontremoli, C.; Azevedo, A.; Quadros, P.; Gentile, P.; Ferreira, A.M.; Dalgarno, K.; et al. Processing of sr2+ containing poly l-lactic acid-based hybrid composites for additive manufacturing of bone scaffolds. Front. Mater. 2020, 7, 601645. [Google Scholar] [CrossRef]
- Yeo, C.; Saunders, N.; Locca, D.; Flett, A.; Preston, M.; Brookman, P.; Davy, B.; Mathur, A.; Agrawal, S. Ficoll-paque versus lymphoprep: A comparative study of two density gradient media for therapeutic bone marrow mononuclear cell preparations. Regen. Med. 2009, 4, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Batsali, A.K.; Pontikoglou, C.; Koutroulakis, D.; Pavlaki, K.I.; Damianaki, A.; Mavroudi, I.; Alpantaki, K.; Kouvidi, E.; Kontakis, G.; Papadaki, H.A. Differential expression of cell cycle and wnt pathway-related genes accounts for differences in the growth and differentiation potential of wharton’s jelly and bone marrow-derived mesenchymal stem cells. Stem Cell Res. Ther. 2017, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, C.; Heinemann, S.; Rößler, S.; Kruppke, B.; Wiesmann, H.P.; Hanke, T. Organically modified hydroxyapatite (ormohap) nanospheres stimulate the differentiation of osteoblast and osteoclast precursors: A co-culture study. Biomed. Mater. 2019, 14, 035015. [Google Scholar] [CrossRef] [PubMed]
- Loukelis, K.; Papadogianni, D.; Chatzinikolaidou, M. Kappa-carrageenan/chitosan/gelatin scaffolds enriched with potassium chloride for bone tissue engineering. Int. J. Biol. Macromol. 2022, 209, 1720–1730. [Google Scholar] [CrossRef] [PubMed]
- Gentleman, E.; Fredholm, Y.C.; Jell, G.; Lotfibakhshaiesh, N.; O’Donnell, M.D.; Hill, R.G.; Stevens, M.M. The effects of strontium-substituted bioactive glasses on osteoblasts and osteoclasts in vitro. Biomaterials 2010, 31, 3949–3956. [Google Scholar] [CrossRef] [PubMed]
- Martí, M.; Mulero, L.; Pardo, C.; Morera, C.; Carrió, M.; Laricchia-Robbio, L.; Esteban, C.R.; Izpisua Belmonte, J.C. Characterization of pluripotent stem cells. Nat. Protoc. 2013, 8, 223–253. [Google Scholar] [CrossRef]
- Wei, L.; Ke, J.; Prasadam, I.; Miron, R.J.; Lin, S.; Xiao, Y.; Chang, J.; Wu, C.; Zhang, Y. A comparative study of sr-incorporated mesoporous bioactive glass scaffolds for regeneration of osteopenic bone defects. Osteoporos. Int. J. Establ. Result Coop. Eur. Found. Osteoporos. Natl. Osteoporos. Found. USA 2014, 25, 2089–2096. [Google Scholar] [CrossRef] [PubMed]
- Papadogiannis, F.; Batsali, A.; Klontzas, M.E.; Karabela, M.; Georgopoulou, A.; Mantalaris, A.; Zafeiropoulos, N.E.; Chatzinikolaidou, M.; Pontikoglou, C. Osteogenic differentiation of bone marrow mesenchymal stem cells on chitosan/gelatin scaffolds: Gene expression profile and mechanical analysis. Biomed. Mater. 2020, 15, 064101. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Tang, L.; Jiang, N.; Zheng, R.; Li, W.; Gu, Y.; Wang, M. Characterization of peripheral blood mononuclear cells isolated using two kinds of leukocyte filters. Transfus. Clin. Biol. J. Soc. Fr. Transfus. Sang. 2020, 27, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Tiedemann, K.; Le Nihouannen, D.; Fong, J.E.; Hussein, O.; Barralet, J.E.; Komarova, S.V. Regulation of osteoclast growth and fusion by mtor/raptor and mtor/rictor/akt. Front. Cell Dev. Biol. 2017, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Bonatti, A.F.C.I.; Micalizzi, S.; Vozzi, G.; De Maria, C. Bioprinting for bone tissue engineering 3D Print. Orthop. Traumatol. 2021, 72, 376–394. [Google Scholar]
- Milazzo, M.; Negrini, N.C.; Scialla, S.; Marelli, B.; Farè, S.; Danti, S.; Buehler, M. Additive manufacturing approaches for hydroxyapatite-reinforced composites. Adv. Funct. Mater. 2019, 29, 1903055. [Google Scholar] [CrossRef]
- Klinder, A.; Markhoff, J.; Jonitz-Heincke, A.; Sterna, P.; Salamon, A.; Bader, R. Comparison of different cell culture plates for the enrichment of non-adherent human mononuclear cells. Exp. Ther. Med. 2019, 17, 2004–2012. [Google Scholar] [CrossRef] [PubMed]
- Salo, J.; Metsikkö, K.; Palokangas, H.; Lehenkari, P.; Väänänen, H.K. Bone-resorbing osteoclasts reveal a dynamic division of basal plasma membrane into two different domains. J. Cell Sci. 1996, 109, 301–307. [Google Scholar] [CrossRef]
- Caudrillier, A.; Hurtel-Lemaire, A.-S.; Wattel, A.; Cournarie, F.; Godin, C.; Petit, L.; Petit, J.-P.; Terwilliger, E.; Kamel, S.; Brown, E.M.; et al. Strontium ranelate decreases receptor activator of nuclear factor-κb ligand-induced osteoclastic differentiation in vitro: Involvement of the calcium-sensing receptor. Mol. Pharmacol. 2010, 78, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Morelli, S.; Salerno, S.; Holopainen, J.; Ritala, M.; De Bartolo, L. Osteogenic and osteoclastogenic differentiation of co-cultured cells in polylactic acid-nanohydroxyapatite fiber scaffolds. J. Biotechnol. 2015, 204, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Young, M.F.; Kerr, J.M.; Ibaraki, K.; Heegaard, A.M.; Robey, P.G. Structure, expression, and regulation of the major noncollagenous matrix proteins of bone. Clin. Orthop. Relat. Res. 1992, 281, 275–294. [Google Scholar] [CrossRef]
- Rosset, E.M.; Bradshaw, A.D. Sparc/osteonectin in mineralized tissue. Matrix Biol. J. Int. Soc. Matrix Biol. 2016, 52–54, 78–87. [Google Scholar] [CrossRef]
- Zhu, Y.-S.; Gu, Y.; Jiang, C.; Chen, L. Osteonectin regulates the extracellular matrix mineralization of osteoblasts through p38 signaling pathway. J. Cell. Physiol. 2020, 235, 2220–2231. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.; Halbout, P. opg/rankl: Role and therapeutic target in osteoporosis. Med. Sci. M/S 2008, 24, 105–110. [Google Scholar]
- Atkins, G.; Welldon, K.; Halbout, P.; Findlay, D. Strontium ranelate treatment of human primary osteoblasts promotes an osteocyte-like phenotype while eliciting an osteoprotegerin response. Osteoporos. Int. 2009, 20, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Liu, X.S.; Huang, S.; Li, Z.; Pan, H.; Zhen, W.; Luk, K.; Guo, X.E.; Lu, W.W. The cross-talk between osteoclasts and osteoblasts in response to strontium treatment: Involvement of osteoprotegerin. Bone 2011, 49, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Yagi, M.; Miyamoto, T.; Sawatani, Y.; Iwamoto, K.; Hosogane, N.; Fujita, N.; Morita, K.; Ninomiya, K.; Suzuki, T.; Miyamoto, K.; et al. Dc-stamp is essential for cell-cell fusion in osteoclasts and foreign body giant cells. J. Exp. Med. 2005, 202, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, N. Regulation of nfatc1 in osteoclast differentiation. J. Bone Metab. 2014, 21, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Kruppke, B.; Ray, S.; Alt, V.; Rohnke, M.; Kern, C.; Kampschulte, M.; Heinemann, C.; Budak, M.; Adam, J.; Döhner, N.; et al. Gelatin-modified calcium/strontium hydrogen phosphates stimulate bone regeneration in osteoblast/osteoclast co-culture and in osteoporotic rat femur defects—In vitro to in vivo translation. Molecules 2020, 25, 5103. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scaffold Designation | Composition |
|---|---|
| blend | Polymeric blend comprising 90/5/5%wt poly(lactic acid)/poly(ε-caprolactone)/poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PLLA/PCL/PHBV) |
| blend_nHA | 90/5/5%wt PLLA/PCL/PHBV, enriched with 2.5% w/v nHA (100% of Ca2+) |
| blend_Sr-nHA | 90/5/5%wt PLLA/PCL/PHBV, enriched with 2.5% w/v Sr-nHA (50% of Sr2+ substitution into the nHA) |
| Gene Symbol | Forward (5′-3′) | Reverse (5′-3′) | Amplicon Size (bp) |
|---|---|---|---|
| B2M | TGTCTTTCAGCAAGGACTGGT | ACATGTCTCGATCCCACTTAAC | 138 |
| SDHA | GCATGCCAGGGAAGACTACA | GCCAACGTCCACATAGGACA | 127 |
| OPG | GTGTGCGAATGCAAGGAAGG | AGCAGGAGACCAAAGACACTG | 209 |
| OSC | GTGCAGCCTTTGTGTCCAAG | TCAGCCAACTCGTCACAGTC | 157 |
| OSN | GAAACCGAAGAGGAGGTGGTG | AGAAGTGGCAGGAAGAGTCGAA | 196 |
| DC-STAMP | CCACAGAGGTGTTGTCCTCC | CCACAAGGGCCCAAAAATCG | 109 |
| NFATC1 | GTCTGGGAGATGGAAGCGAAAACT | CTGGTACTGGCTTCGCTTTCTCTT | 111 |
| TRAP | GGCAGGCAGGGAGGGAATAAA | AGTCACCCACGGCTACAAAGC | 200 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kontogianni, G.-I.; Bonatti, A.F.; De Maria, C.; Naseem, R.; Coelho, C.; Alpantaki, K.; Batsali, A.; Pontikoglou, C.; Quadros, P.; Dalgarno, K.; et al. Cell Instructive Behavior of Composite Scaffolds in a Co-Culture of Human Mesenchymal Stem Cells and Peripheral Blood Mononuclear Cells. J. Funct. Biomater. 2024, 15, 116. https://doi.org/10.3390/jfb15050116
Kontogianni G-I, Bonatti AF, De Maria C, Naseem R, Coelho C, Alpantaki K, Batsali A, Pontikoglou C, Quadros P, Dalgarno K, et al. Cell Instructive Behavior of Composite Scaffolds in a Co-Culture of Human Mesenchymal Stem Cells and Peripheral Blood Mononuclear Cells. Journal of Functional Biomaterials. 2024; 15(5):116. https://doi.org/10.3390/jfb15050116
Chicago/Turabian StyleKontogianni, Georgia-Ioanna, Amedeo Franco Bonatti, Carmelo De Maria, Raasti Naseem, Catarina Coelho, Kalliopi Alpantaki, Aristea Batsali, Charalampos Pontikoglou, Paulo Quadros, Kenneth Dalgarno, and et al. 2024. "Cell Instructive Behavior of Composite Scaffolds in a Co-Culture of Human Mesenchymal Stem Cells and Peripheral Blood Mononuclear Cells" Journal of Functional Biomaterials 15, no. 5: 116. https://doi.org/10.3390/jfb15050116
APA StyleKontogianni, G. -I., Bonatti, A. F., De Maria, C., Naseem, R., Coelho, C., Alpantaki, K., Batsali, A., Pontikoglou, C., Quadros, P., Dalgarno, K., Vozzi, G., Vitale-Brovarone, C., & Chatzinikolaidou, M. (2024). Cell Instructive Behavior of Composite Scaffolds in a Co-Culture of Human Mesenchymal Stem Cells and Peripheral Blood Mononuclear Cells. Journal of Functional Biomaterials, 15(5), 116. https://doi.org/10.3390/jfb15050116