
Peptide Amphiphiles in Corneal Tissue Engineering



Abstract
:1. Introduction
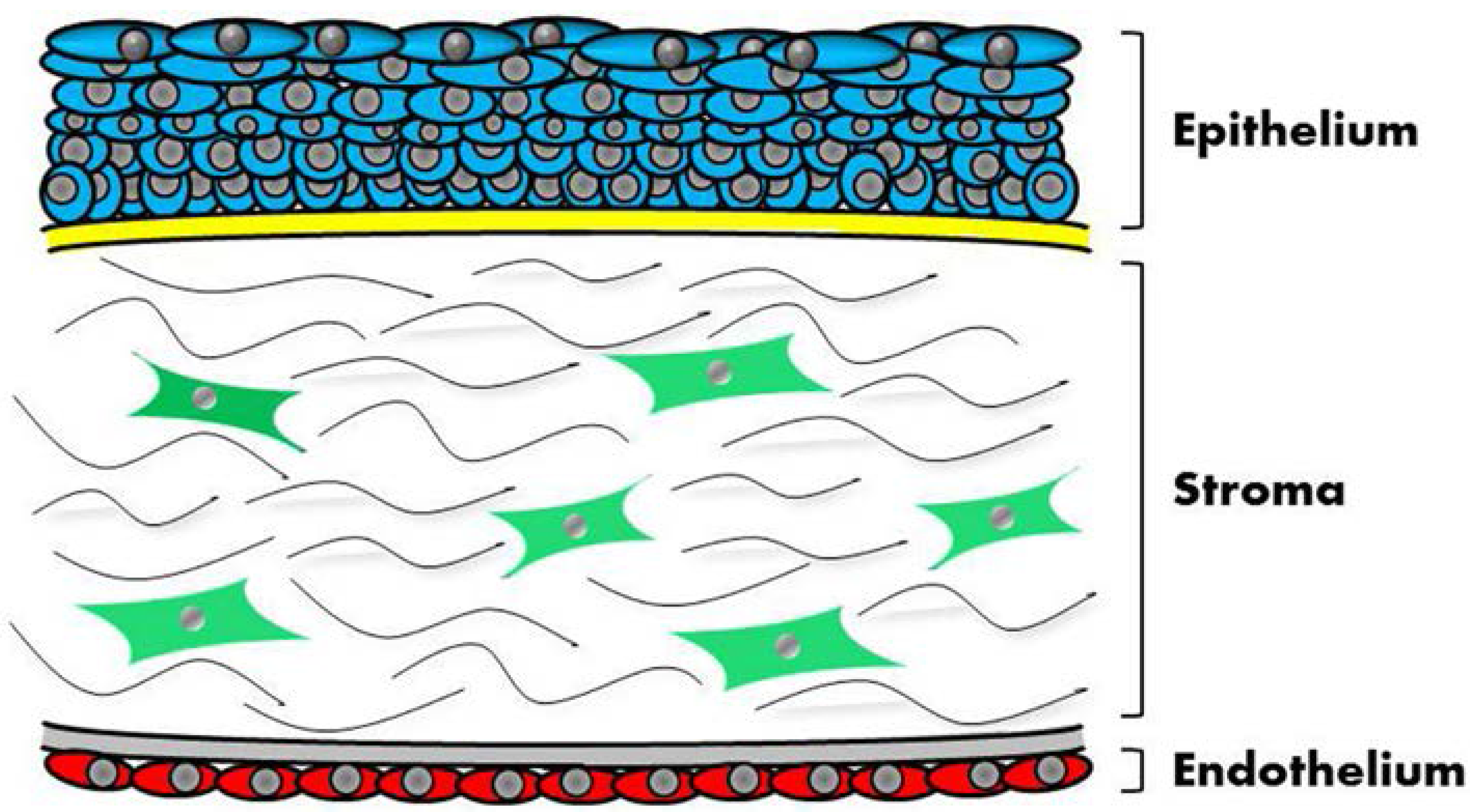
2. Challenges in Corneal Tissue Engineering

3. Previous Approaches to Engineer Corneal Tissue: Top-Down or Bottom-Up?
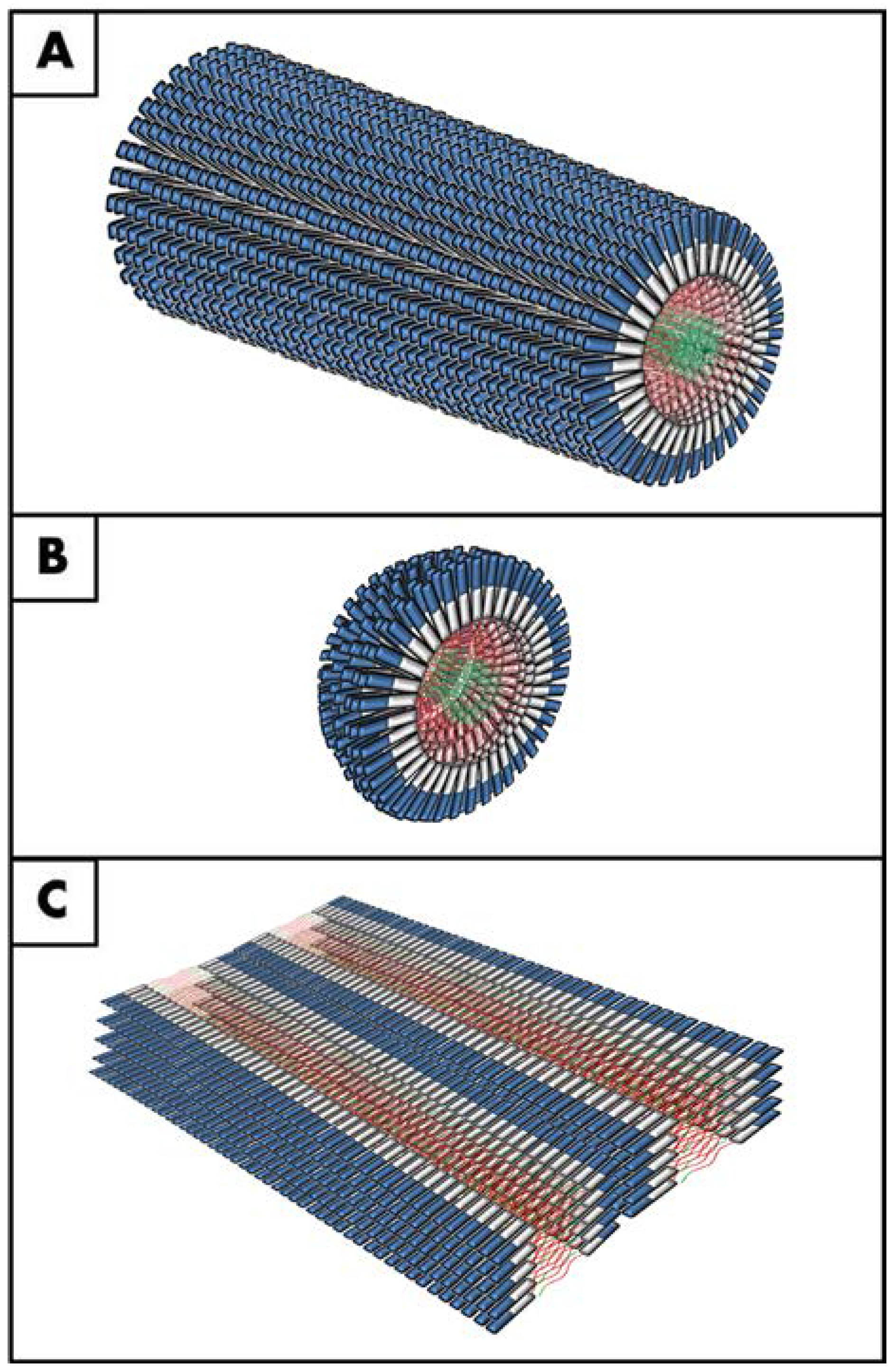
4. Peptide Amphiphiles in Tissue Engineering

5. Peptide Amphiphiles as Versatile Templates to Recreate Human Corneas in vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PA | Chemical Structure | Aim | Source | [PA] wt % | PA Form | Reference |
|---|---|---|---|---|---|---|
| C16-C4-G3-S-RGD |  | Bone regeneration | Fibronectin | 0.1 | coating | [83] |
| C12-HSNGLPLGGGS EEEAAAVVV(K) |  | Cartilage regeneration | De novo synthetized | 1 | hydrogel | [84] |
| C16-V2A2E2-NH2 |  | Cavernous nerve regeneration | De novo synthetized | 0.85 | hydrogel | [87] |
| C16-C4-G3- LRKKLGKA |  | Blood vessels regeneration | Heparin binding consensus sequence | 3 | hydrogel | [85] |
| C16-KTTKS |  | Skin regeneration | Procollagen I | 0.0003 | solution | [88] |
| PA | Source | Biological Effect | [PA] wt % | PA Form | Reference |
|---|---|---|---|---|---|
| C16-G3-RGD/RGDS + C16-ETTES | Fibronectin | Enhanced adhesion and proliferation of hCSFs | 1 to 0.005 | coating | [42,96] |
| A6-RGDS | Fibronectin | Enhanced adhesion and proliferation of hCSFs | 1 to 0.1 | coating | [97] |
| C16-TPGPQGIAGQ-RGDS | MMP cleavage sequence + Fibronectin | Promoted adhesion andgrowth of hCSFs. Stimulated collagen production. Governed tissues lift-up | 2 | coating | [44,74] |
| Fmoc-RGDS | Fibronectin | Enhanced cell attachment, proliferation and viability | 1 | solution | [43] |
| C16-KTTKS | Procollagen I | Stimulated collagen production from hCSFs | 0.002, 0.004, 0.008 | solution | [98,99] |
| C12-VVAGKYIGSR | Laminin | Enhanced keratocyte proliferation and migration, and stimulated collagen I synthesis | 0.2 | coating | [100] |
| C16-YEALRVANEVTLN | Lumican | Stimulated collagen I production | 0.01, 0.005, 0.0025 0.00125, 0.000625 | solution | [101] |


6. Future Perspectives
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ruberti, J.W.; Zieske, J.D. Prelude to corneal tissue engineering—Gaining control of collagen organization. Prog. Retin. Eye Res. 2008, 27, 549–577. [Google Scholar] [CrossRef] [PubMed]
- Whitcher, J.P.; Srinivasan, I.M.; Upadhyay, M.P. Corneal blindness: A global perspective. Bull. World Health Organ. 2001, 79, 214–221. [Google Scholar] [PubMed]
- Trinkaus-Randall, V. Principles of tissue engineering. Cornea 2000, 471–491. [Google Scholar]
- Thompson, R.W.J.; Price, M.O.; Bowers, P.J.; Price, F.W.J. Long-term graft survival after penetrating keratoplasty. Ophthalmology 2003, 110, 1396–1402. [Google Scholar] [CrossRef]
- McLaughlin, C.R.; Tsai, R.J.; Latorre, M.A.; Griffith, M. Bioengineered corneas for transplantation and in vitro toxicology. Front. Biosci. 2009, 14, 3326–3337. [Google Scholar] [CrossRef]
- Fagerholm, P.; Lagali, N.S.; Merrett, K.; Jackson, W.B.; Munger, R.; Liu, Y.; Polarek, J.W.; Söderqvist, M.; Griffith, M. A biosynthetic alternative to human donor tissue for inducing corneal regeneration: 24-month follow-up of a phase 1 clinical study. Sci. Transl. Med. 2010, 2, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.G.; Deng, C.; McLaughlin, C.R.; Fagerholm, P.; Lagali, N.S.; Heyne, B.; Scaiano, J.C.; Watsky, M.A.; Kato, Y.; Munger, R.; et al. Collagen-phosphorylcholine interpenetrating network hydrogels as corneal substitutes. Biomaterials 2009, 30, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Alaminos, M.; Del Carmen Sanchez-Quevedo, M.; Munoz-Avila, J.I.; Serrano, D.; Medialdea, S.; Carreras, I.; Campos, A. Construction of a complete rabbit cornea substitute using a fibrin-agarose scaffold. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3311–3317. [Google Scholar] [CrossRef] [PubMed]
- Levis, H.J.; Kureshi, A.K.; Massie, I.; Morgan, L.; Vernon, A.J.; Daniels, J.T. Tissue engineering the cornea: The evolution of raft. J. Funct. Biomater. 2015, 6, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, S.S.; Lim, R.R.; Lakshminarayanan, R.; Mohan, R.R. Nanomedicine approaches for corneal diseases. J. Funct. Biomater. 2015, 6, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Utheim, T.P. Limbal epithelial cell theraphy: Past, present, and future. In Corneal Regenerative Medicine: Methods and Protocols; Wright, B.C., Connon, C.J., Eds.; Humana Press: New York, NY, USA, 2013; Volume 1014, pp. 3–43. [Google Scholar]
- Goldman, J.N.; Benedek, G.B.; Dohlman, C.H.; Kravitt, B. Structural alterations affecting transparency in swollen human corneas. Investig. Ophthalmol. 1968, 7, 501–519. [Google Scholar]
- Müller, L.J.; Pels, E.; Schurmans, L.R.H.M.; Vrensen, G.F.J.M. A new three-dimensional model of the organization of proteoglycans and collagen fibrils in the human corneal stroma. Exp. Eye Res. 2004, 78, 493–501. [Google Scholar] [CrossRef]
- Jester, J.V.; Petroll, W.M.; Cavanagh, H.D. Corneal stromal wound healing in refractive surgery: The role of the myofibroblast. Prog. Retin. Eye Res. 1999, 18, 311–356. [Google Scholar] [CrossRef]
- Jester, J.V.; Moller-Pedersen, T.; Huang, J.; Sax, C.M.; Kays, W.T.; Cavangh, H.D.; Petroll, W.M.; Piatigorsky, J. The cellular basis of corneal transparency: Evidence for “corneal crystallins”. J. Cell Sci. 1999, 112, 613–622. [Google Scholar] [PubMed]
- Azar, D.T. Corneal angiogenic privilege: Angiogenic and antiangiogenic factors in corneal avascularity, vasculogenesis, and wound healing. Trans. Am. Ophthalmol. Soc. 2006, 104, 264–302. [Google Scholar] [PubMed]
- Qazi, Y.; Wong, G.; Monson, B.; Stringham, J.; Ambati, B.K. Corneal transparency: Genesis, maintenance and dysfunction. Brain Res. Bull. 2010, 81, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K. Interaction between injured corneal epithelial cells and stromal cells. Cornea 2003, 22, S35–S47. [Google Scholar] [CrossRef] [PubMed]
- Joice, N.C. Proliferative capacity of the corneal endothelium. Prog. Retin. Eye Res. 2003, 22, 359–389. [Google Scholar] [CrossRef]
- De Castro, F.; Silos-Santiago, I.; Lopez de Armentia, M.; Barbacid, M.; Belmonte, C. Corneal innervation and sensitivity to noxious stimuli intrka knockout mice. Eur. J. Neurosci. 2010, 10, 146–152. [Google Scholar]
- Gonzalez-Andrades, M.; De la Cruz Cardona, J.; Ionescu, A.M.; Campos, A.; Del Mar Perez, M.; Alaminos, M. Generation of bioengineered corneas with decellularized xenografts and human keratocytes. Investig. Ophthalmol. Vis. Sci. 2011, 52, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Chen, J.; Funderburgh, J.L.; Zhu, X.; Li, L. Functional reconstruction of rabbit corneal epithelium by human limbal cells cultured on amniotic membrane. Mol. Vis. 2003, 9, 635–643. [Google Scholar] [PubMed]
- Mi, S.; Chen, B.; Wright, B.; Connon, C.J. Ex vivo construction of an artificial ocular surface by combination of corneal limbal epithelial cells and a compressed collagen scaffold containing keratocytes. Tissue Eng. Part A 2010, 1, 2091–2100. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, A.M.; de la Cruz Cardona, J.; Gonzalez-Andrades, M.; Alaminos, M.; Campos, A.; Hita, E.; del Mar Perez, M. UV absorbance of a bioengineered corneal stroma substitute in the 240–400 nm range. Cornea 2010, 29, 895–898. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, A.M.; Alaminos, M.; de la Cruz Cardona, J.; de Dios García-López Durán, J.; Gonzalez-Andrades, M.; Ghinea, R.; Campos, A.; Hita, E.; del Mar Pérez, M. Investigating a novel nanostructured fibrin-agarose biomaterial for human cornea tissue engineering: Rheological properties. J. Mech. Behav. Biomed. Mater. 2011, 4, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz Cardona, J.; Ionescu, A.M.; Gomez-Sotomayor, R.; Gonzalez-Andrades, M.; Campos, A.; Alaminos, M.; del Mar Pérez, M. Transparency in a fibrin and fibrin-agarose corneal stroma substitute generated by tissue engineering. Cornea 2001, 30, 1428–1435. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Ritch, R.; Lin, S.M.; Ni, M.H.; Chang, Y.C.; Lu, Y.L.; Lai, H.J.; Lin, F.H. A new fish scale derived scaffold for corneal regeneration. Eur. Cell. Mater. 2010, 19, 50–57. [Google Scholar] [PubMed]
- Liang, Y.; Liu, W.; Han, B.; Yang, C.; Ma, Q.; Zhao, W.; Rong, M.; Li, M. Fabrication and characters of a corneal endothelial cells scaffold based on chitosan. J. Mater. Sci. Mater. Med. 2011, 22, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Mohanty, S.; Gupta, D.; Jassal, M.; Agrawal, A.K.; Tandon, R. Cellular response of limbal epithelial cells on electrospun poly-epsilon-caprolactone nanofibrous scaffolds for ocular surface bioengineering: A preliminary in vitro study. Mol. Vis. 2011, 17, 2898–2910. [Google Scholar] [PubMed]
- Wu, J.; Du, Y.; Watkins, S.C.; Funderburgh, J.L.; Wagner, W.R. The engineering of organized human corneal tissue through the spatial guidance of corneal stromal stem cells. Biomaterials 2012, 33, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Gan, L.; Carlsson, D.J.; Fagerholm, P.; Lagali, N.; Watsky, M.A.; Munger, R.; Hodge, W.G.; Priest, D.; Griffith, M. A simple, cross-linked collagen tissue substitute for corneal implantation. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1869–1875. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Merrett, K.; Griffith, M.; Fagerholm, P.; Dravida, S.; Heyne, B.; Scaiano, J.C.; Watsky, M.A.; Shinozaki, N.; Lagali, N.; et al. Recombinant human collagen for tissue engineered corneal substitutes. Biomaterials 2008, 29, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; McLaughlin, C.; Griffith, M.; Sheardown, H. Biofunctionalization of collagen for improved biological response: Scaffolds for corneal tissue engineering. Biomaterials 2007, 28, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Ott, H.C.; Matthiesen, T.S.; Goh, S.K.; Black, L.D.; Kren, S.M.; Netoff, T.I.; Taylor, D.A. Perfusion-decellularized matrix: Using nature’s platform to engineer a bioartificial heart. Nat. Med. 2008, 14, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Dean, D.M.; Napolitano, A.P.; Youssef, J.; Morgan, J.R. Rods, tori, and honeycombs: The directed self-assembly of microtissues with prescribed microscale geometries. FASEB J. 2007, 21, 4005–4012. [Google Scholar] [CrossRef] [PubMed]
- Yeh, J.; Ling, Y.; Karp, J.M.; Gantze, J.; Chandawarkard, A.; Eng, G.; Blumling, J.; Langer, R.; Khademhosseini, A. Micromolding of shape-controlled, harvestable cell-laden hydrogels. Biomaterials 2006, 27, 5391–5398. [Google Scholar] [CrossRef] [PubMed]
- Karamichos, D.; Funderburgh, M.L.; Hutcheon, A.E.; Zieske, J.D.; Du, Y.; Wu, J.; Funderburgh, J.L. A role for topographic cues in the organization of collagenous matrix by corneal fibroblasts and stem cells. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- L’Heureux, N.; Paquet, S.; Labbe, R.; Germain, L.; Auger, F.A. A completely biological tissue-engineered human blood vessel. FASEB J. 1998, 12, 47–56. [Google Scholar] [PubMed]
- Mironov, V.; Boland, T.; Trusk, T.; Forgacs, G.; Markwald, R.R. Organ printing: Computer-aided jet-based 3d tissue engineering. Trends Biotechnol. 2003, 21, 157–161. [Google Scholar] [CrossRef]
- Tsang, V.L.; Chen, A.A.; Cho, L.M.; Jadin, K.D.; Sah, R.L.; DeLong, S.; West, J.L.; Bhatia, S.N. Fabrication of 3D hepatic tissues by additive photopatterning of cellular hydrogels. FASEB J. 2007, 21, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Connon, C.J. Approaches to corneal tissue engineering: Top-down or bottom-up? Procedia Eng. 2015, 110, 15–20. [Google Scholar] [CrossRef]
- Gouveia, R.M.; Castelletto, V.; Alcock, S.G.; Hamley, I.W.; Connon, C.J. Bioactive films produced from self-assembling peptide amphiphiles as versatile substrates for tuning cell adhesion and tissue architecture in serum-free conditions. J. Mater. Chem. B 2013, 1, 6157–6169. [Google Scholar] [CrossRef]
- Gouveia, R.M.; Jones, R.R.; Hamley, I.W.; Connon, C.J. The bioactivity of composite fmoc-rgds-collagen gels. Biomater. Sci. 2014, 2, 1222–1229. [Google Scholar] [CrossRef]
- Dehsorkhi, A.; Gouveia, R.M.; Smith, A.M.; Hamley, I.W.; Castelletto, V.; Connon, C.J.; Reza, M.; Ruokolainen, J. Self-assembly of a dual functional bioactive peptide amphiphile incorporating both matrix metalloprotease substrate and cell adhesion motifs. Soft Matter 2015, 11, 3115–3124. [Google Scholar] [CrossRef] [PubMed]
- Storrie, H.; Guler, M.O.; Abu-Amara, S.N.; Volberg, T.; Rao, M.; Geiger, B.; Stupp, S.I. Supramolecular crafting of cell adhesion. Biomaterials 2007, 28, 4608–4618. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Smith, A.M.; Das, A.K.; Hodson, N.W.; Collins, R.F.; Ulijin, R.V.; Gough, J.E. Self-assembled peptide-based hydrogels as scaffolds for anchorage-dependent cells. Biomaterials 2009, 30, 2523–2530. [Google Scholar] [CrossRef] [PubMed]
- Nagai, Y.; Unsworth, L.D.; Koutsopoulos, S.; Zhang, S.G. Slow release of molecules in self-assembling peptide nanofiber scaffold. J. Control. Release 2006, 115, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Branco, M.C.; Pochan, D.J.; Wagner, N.J.; Schneider, J.P. Macromolecular diffusion and release from self-assembled beta-hairpin peptide hydrogels. Biomaterials 2009, 30, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Salick, D.A.; Kretsinger, J.K.; Pochan, D.J.; Schneider, J.P. Inherent antibacterial activity of a peptide-based beta-hairpin hydrogel. J. Am. Chem. Soc. 2007, 129, 14793–14799. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.M.; Liang, G.L.; Guo, Z.F.; Guo, Z.H.; Xu, B. Intracellular hydrogelation of small molecules inhibits bacterial growth. Angew. Chem. Intl. Ed. 2007, 46, 8216–8219. [Google Scholar] [CrossRef] [PubMed]
- Sundar, S.; Chen, Y.; Tong, Y.W. Delivery of therapeutics and molecules using self-assembled peptides. Curr. Med. Chem. 2014, 21, 2469–2479. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.G.; Webber, M.J.; Stupp, S.I. Self-assembly of peptide amphiphiles: From molecules to nanostructures to biomaterials. Biopolymers 2010, 94, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Webber, M.J.; Tongers, J.; Renault, M.A.; Roncalli, J.G.; Losordo, D.W.; Stupp, S.I. Development of bioactive peptide amphiphiles for therapeutic cell delivery. Acta Biomater. 2010, 6, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Beniash, E.; Hartgerink, J.D.; Storrie, H.; Stendahl, J.C.; Stupp, S.I. Self-assembling peptide amphiphile nanofiber matrices for cell entrapment. Acta Biomater. 2005, 1, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Galler, K.M.; Cavender, A.; Yuwono, V.; Dong, H.; Shi, S.T.; Schmalz, G.; Hartgerink, J.D.; D’Souza, R.N. Self-assembling peptide amphiphile nanofibers as a scaffold for dental stem cells. Tissue Eng. Part A 2008, 14, 2051–2058. [Google Scholar] [CrossRef] [PubMed]
- Weib, N.; Klee, D.; Hocker, H. Konzept zur bioaktiven ausrüstung. Biomaterialien 2001, 2, 81–86. [Google Scholar]
- Shroff, K.; Rexeisen, E.L.; Arunagirinathan, M.A.; Kokkoli, E. Fibronectin-mimetic peptide-amphiphile nanofiber gels support increased cell adhesion and promote ecm production. Soft Matter 2010, 6, 5064–5072. [Google Scholar] [CrossRef]
- Rexeisen, E.L.; Fan, W.; Pangburn, T.O.; Taribagil, R.R.; Bates, F.S.; Lodge, T.P.; Tsapatsis, M.; Kokkoli, E. Self-assembly of fibronectin mimetic peptide-amphiphile nanofibers. Langmuir 2010, 26, 1953–1959. [Google Scholar] [CrossRef] [PubMed]
- Stupp, S.I.; Pralle, M.U.; Tew, G.N.; Li, L.M.; Sayar, M.; Zubarev, E.R. Self-assembly of organic nano-objects into functional materials. MRS Bull. 2000, 25, 42–48. [Google Scholar] [CrossRef]
- Ghadiri, M.R.; Granja, J.R.; Milligan, R.A.; McRee, D.E.; Khazanovich, N. Self-assembling organic nanotubes based on a cyclic peptide architecture. Nature 1993, 366, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Muraoka, T.; Cheetham, A.G.; Stupp, S.I. Self-assembly of giant peptide nanobelts. Nano Lett. 2009, 9, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Stupp, S.I.; Zha, R.H.; Palmer, L.C.; Cui, H.G.; Bitton, R. Self-assembly of biomolecular soft matter. Faraday Discuss. 2013, 166, 9–30. [Google Scholar] [CrossRef] [PubMed]
- Hamley, I.W. Self-assembly of amphiphilic peptides. Soft Matter 2011, 7, 4122–4138. [Google Scholar] [CrossRef]
- Maude, S.; Tai, L.R.; Davies, R.P.W.; Liu, B.; Harris, S.A.; Kocienski, P.J.; Aggeli, A. Peptide synthesis and self-assembly. Top. Curr. Chem. 2012, 310, 27–69. [Google Scholar] [PubMed]
- Nagarkar, R.P.; Schneider, J.P. Synthesis and primary characterization of self-assembled peptide-based hydrogels. Methods Mol. Biol. 2008, 474, 61–77. [Google Scholar] [PubMed]
- Cavalli, S.; Albericio, F.; Kros, A. Amphiphilic peptides and their cross-disciplinary role as building blocks for nanoscience. Chem. Soc. Rev. 2010, 39, 241–263. [Google Scholar] [CrossRef] [PubMed]
- Pashuck, E.T.; Cui, H.; Stupp, S.I. Tuning supramolecular rigidity of peptide fibers through molecular structure. J. Am. Chem. Soc. 2010, 132, 6041–6046. [Google Scholar] [CrossRef] [PubMed]
- Matson, J.B.; Stupp, S.I. Self-assembling peptide scaffolds for regenerative medicine. Chem. Commun. 2012, 48, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Behanna, H.A.; Donners, J.J.J.M.; Gordon, A.C.; Stupp, S.I. Coassembly of amphiphiles with opposite peptide polarities into nanofibers. J. Am. Chem. Soc. 2005, 127, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Kokkoli, E.; Mardilovich, A.; Wedekind, A.; Rexeisen, E.L.; Garg, A.; Craig, J.A. Self-assembly and applications of biomimetic and bioactive peptide amphiphiles. Soft Matter 2006, 2, 1015–1024. [Google Scholar] [CrossRef]
- Kunitake, T. Synthetic bilayer membranes: Molecular design, self-organization, and application. Angew. Chem. Intl. Ed. 1992, 31, 709–726. [Google Scholar] [CrossRef]
- Gore, T.; Dori, Y.; Talmon, Y.; Tirrell, M.; Bianco-Peled, H. Self-assembly of model collagen peptide amphiphiles. Langmuir 2001, 17, 5352–5360. [Google Scholar] [CrossRef]
- Yu, Y.C.; Tirrell, M.; Fields, G.B. Minimal lipidation stabilizes protein-like molecular architecture. J. Am. Chem. Soc. 1998, 120, 9979–9987. [Google Scholar] [CrossRef]
- Gouveia, R.M.; Castelletto, V.; Hamley, I.W.; Connon, C.J. New self-assembling multi-functional templates for the biofabrication and controlled self-release of cultured tissues. Tissue Eng. Part A 2015, 21, 1772–1784. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.D.; Jin, Y.; Liu, Y.; Zhang, X.Z.; Zhuo, R.X. Self-assembly behaviour of peptide amphiphiles (PAs) with different length of hydrophobic alkyl tails. Colloid Surf. B 2010, 81, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Morandat, S.; Bortolato, M.; Anker, G.; Doutheau, A.; Lagarde, M.; Chauvet, J.P.; Roux, B. Plasmalogens protect unsaturated lipids against uv-induced oxidation in monolayer. BBA Biomembr. 2003, 1616, 137–146. [Google Scholar] [CrossRef]
- Reinitz, S.D.; Currier, B.H.; Levine, R.A.; Van Citters, D.W. Crosslink density, oxidation and chain scission in retrieved, highly cross-linked uhmwpe tibial bearings. Biomaterials 2014, 35, 4436–4440. [Google Scholar] [CrossRef] [PubMed]
- Aulisa, L.; Forraz, N.; McGuckin, C.; Hartgerink, J.D. Inhibition of cancer cell proliferation by designed peptide amphiphiles. Acta Biomater. 2009, 5, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Hashidzume, A.; Harada, A. Micelles and vesicles. In Encyclopedia of polymeric Nanomaterials; Kobayashi, S., Müllen, K., Eds.; Springer Berlin Heidelberg: Berlin, Jermany, 2015; pp. 1–5. [Google Scholar]
- El-Sherbiny, I.M.; Yacoub, M.H. Hydrogel scaffolds for tissue engineering: Progress and challenges. Glob. cardiol. Sci. Pract. 2013, 2013, 316–342. [Google Scholar] [CrossRef] [PubMed]
- Aszódi, A.; Legate, K.R.; Nakchbandi, I.; Fässler, R. What mouse mutants teach us about extracellular matrix function. Annu. Rev. Cell. Dev. Biol. 2006, 22, 591–621. [Google Scholar] [CrossRef] [PubMed]
- Sargeant, T.D.; Gulerb, M.O.; Oppenheimera, S.M.; Mata, A.; Satcher, R.L.; Dunanda, D.C.; Stupp, S.I. Hybrid bone implants: Self-assembly of peptide amphiphile nanofibers within porous titanium. Biomaterials 2008, 29, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Hartgerink, J.D.; Beniash, E.; Stupp, S.I. Self-assembly and mineralization of peptide-amphiphile nanofibers. Science 2001, 294, 1684–1688. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.M.; Shah, N.A.; Del Rosario Lim, M.M.; Hsieh, C.; Nuber, G.; Stupp, S.I. Supramolecular design of self-assembling nanofibers for cartilage regeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 3293–3298. [Google Scholar] [CrossRef] [PubMed]
- Rajangam, K.; Behanna, H.A.; Hui, M.J.; Han, X.; Hulvat, J.F.; Lomasney, J.W.; Stupp, S.I. Heparin binding nanostructures to promote growth of blood vessels. Nano Lett. 2006, 6, 2086–2090. [Google Scholar] [CrossRef] [PubMed]
- Stendahl, J.C.; Wang, L.J.; Chow, L.W.; Kaufman, D.B.; Stupp, S.I. Growth factor delivery from self-assembling nanofibers to facilitate islet transplantation. Transplantation 2008, 86, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, N.L.; Bond, C.W.; Tang, Y.; Harrington, D.A.; Zhang, S.; Stupp, S.I.; McKenna, K.E.; Podlasek, C.A. Regeneration of the cavernous nerve by sonic hedgehog using aligned peptide amphiphile nanofibers. Biomaterials 2011, 32, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.R.; Fitzgerald, N.C.; Doughty, D.G.; Dawes, N.C.; Berge, C.A.; Bissett, D.L. Topical palmitoyl pentapeptide provides improvement in photoaged human facial skin. Int. J. Cosmet. Sci. 2005, 27, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.S.; Marques, A.P.; Reisa, R.L.; Azevedo, H.S. Hyaluronan and self-assembling peptides as building blocks to reconstruct the extracellular environment in skin tissue. Biomater. Sci. 2013, 1, 952–964. [Google Scholar] [CrossRef] [Green Version]
- Germain, L.; Auger, F.A.; Grandbois, E.; Guignard, R.; Giasson, M.; Boisjoly, H.; Guérin, S.L. Reconstructed human cornea produced in vitro by tissue engineering. Pathobiology 1999, 67, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Griffith, M.; Hakim, M.; Shimmura, S.; Watsky, M.A.; Li, F.; Carlsson, D.; Doillon, C.J.; Nakamura, M.; Suuronen, E.; Shinozaki, N.; et al. Artificial human corneas: Scaffolds for transplantation and host regeneration. Cornea 2002, 21, 54–61. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Funamoto, S.; Sasaki, S.; Honda, T.; Hattori, S.; Nam, K.; Kimura, T.; Mochizuki, M.; Fujisato, T.; Kobayashi, H.; et al. Preparation and characterization of decellularized cornea using high-hydrostatic pressurization for corneal tissue engineering. Biomaterials 2010, 31, 3941–3948. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Jones, R.R.; Mi, S.; Foster, J.; Alcock, S.; Hamley, I.W.; Connon, C.J. The mechanical properties of amniotic membrane influence its effect as a biomaterial for ocular surface repair. Soft Matter 2012, 8, 8379–8387. [Google Scholar] [CrossRef]
- Gomes, J.A.; Dos Santos, M.S.; Cunha, M.C.; Mascaro, V.L.; Barros, J.d.; De Sousa, L.B. Amniotic membrane transplantation for partial and total limbal stem cell deficiency secondary to chemical burn. Opthalmology 2003, 110, 466–473. [Google Scholar] [CrossRef]
- Carsson, D.J.; Li, F.; Shimmura, S.; Griffith, M. Bioengineered corneas: How close are we? Curr. Opin. Opthalmol. 2003, 14, 192–197. [Google Scholar] [CrossRef]
- Castelletto, V.; Gouveia, R.M.; Connon, C.J.; Hamley, I.W. New rgd-peptide amphiphile mixtures containing a negatively charged diluent. Faraday Discuss. 2013, 166, 381–397. [Google Scholar] [CrossRef] [PubMed]
- Castelletto, V.; Gouveia, R.M.; Connon, C.J.; Hamley, I.W.; Seitsonen, J.; Nykänen, A.; Ruokolainen, J. Alanine-rich amphiphilic peptide containing the rgd cell adhesion motif: A coating material for human fibroblast attachment and culture. Biomater. Sci. 2014, 2, 362–369. [Google Scholar] [CrossRef]
- Castelletto, V.; Hamley, I.W.; Perez, J.; Abezgauz, L.; Danino, D. Fibrillar superstructure from extended nanotapes formed by a collagen-stimulating peptide. Chem. Commun. 2010, 46, 9185–9187. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.R.; Castelletto, V.; Connon, C.J.; Hamley, I.W. Collagen stimulating effect of peptide amphiphile c16-kttks on human fibroblasts. Mol. Pharm. 2013, 10, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Uzunalli, G.; Soran, Z.; Erkal, T.S.; Dagdas, Y.S.; Dinc, E.; Hondur, A.M.; Bilgihan, K.; Aydin, B.; Guler, M.O.; Tekinay, A.B. Bioactive self-assembled peptide nanofibers for corneal stroma regeneration. Acta Biomater. 2014, 10, 1156–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamley, I.W.; Dehsorkhi, A.; Walter, M.N.M.; Connon, C.J.; Reza, M.; Ruokolainen, J. Self-assembly and collagen stimulating activity of a peptide amphiphile incorporating a peptide sequence from lumican. Langmuir 2015, 31, 4490–4495. [Google Scholar] [CrossRef] [PubMed]
- Gribova, V.; Crouzier, T.; Picart, C. A material's point of view on recent developments of polymeric biomaterials: Control of mechanical and biochemical properties. J. Mater. Chem. 2011, 21, 14354–14366. [Google Scholar] [CrossRef] [PubMed]
- Hersel, U.; Dahmen, C.; Kessler, H. Rgd modified polymers: Biomaterials for stimulated cell adhesion and beyond. Biomaterials 2003, 24, 4385–4415. [Google Scholar] [CrossRef] [PubMed]
- Pierschbacher, M.D.; Ruoslahti, E. Cell attachment activity of fibronectin can be duplicated by small synthetic fragments of the molecule. Nature 1984, 309, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, R.M.; Connon, C.J. The effects of retinoic acid on human corneal stromal keratocytes cultured in vitro under serum-free conditions. Invest. Opthalmol. Vis. Sci. 2013, 54, 7483–7491. [Google Scholar] [CrossRef] [PubMed]
- Bard, J.B.L.; Hay, E.D. The behavior of fibroblasts from the developing avian cornea: Morphology and movement in situ and in vitro. J. Cell Biol. 1975, 67, 400–418. [Google Scholar] [CrossRef] [PubMed]
- Cintron, C.; Covington, H.; Kublin, C.L. Morphogenesis of rabbit corneal stroma. Investig. Ophthalmol. Vis. Sci. 1983, 24, 543–556. [Google Scholar]
- Hay, E.D. The mesenchymal cell, its role in the embryo, and the remarkable signaling mechanisms that create it. Dev. Dyn. 2005, 233, 706–720. [Google Scholar] [CrossRef] [PubMed]
- Maurice, D.M. The structure and transparency of the cornea. J. Physiol. 1957, 136, 263–286. [Google Scholar] [CrossRef] [PubMed]
- Trelstad, R.L.; Coulombre, A.J. Morphogenesis of the collagenous stroma in the chick cornea. J. Cell Biol. 1971, 50, 840–858. [Google Scholar] [CrossRef] [PubMed]
- Petroll, W.M.; New, K.; Sachdev, M.; Cavanagh, H.D.; Jester, J.V. Radial keratotomy. III: Relationship between wound gape and corneal curvature in primate eyes. Investig. Ophthalmol. Vis. Sci. 1992, 33, 3283–3291. [Google Scholar]
- Moller-Pedersen, T.; Vogel, M.D.; Li, H.; Petroll, W.M.; Cavanagh, H.D.; Jester, J.V. Quantification of stromal thinning, epithelial thickness, and corneal haze following photorefractive keratectomy using in vivo confocal microscopy. Ophthalmology 1997, 104, 360–368. [Google Scholar] [CrossRef]
- Karamichos, D.; Lakshman, N.; Petroll, W.M. An experimental model for assessing fibroblast migration in 3-d collagen matrices. Cell Motil. Cytoskelet. 2009, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Lakshman, N.; Karamichos, D.; Petroll, W.M. Growth factor regulation of corneal keratocyte differentiation and migration in compressed collagen matrices. Investig. Ophthalmol. Vis. Sci. 2010, 51, 864–875. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Zhou, C.; Lakshman, N.; Petroll, W.M. Corneal stromal cells use both high- and low-contractility migration mechanisms in 3-D collagen matrices. Exp. Cell Res. 2012, 318, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, O.; Yuan, Y.; Coulson-Thomas, V.J.; Gesteira, T.F.; Call, M.K.; Zhang, Y.J.; Zhang, J.H.; Chang, S.H.; Xie, C.C.; Liu, C.Y.; et al. Lumican binds alk5 to promote epithelium wound healing. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Carlson, E.C.; Lin, M.; Liu, C.Y.; Kao, W.W.; Perez, V.L.; Pearlman, E. Keratocan and lumican regulate neutrophil infiltration and corneal clarity in lipopolysaccharide-induced keratitis by direct interaction with CXCL1. J. Biol. Chem. 2007, 282, 35502–35509. [Google Scholar] [CrossRef] [PubMed]
- Saika, S.; Shiraishi, A.; Liu, C.Y.; Funderburgh, J.L.; Kao, C.W.; Converse, R.L.; Kao, W.W. Role of lumican in the corneal epithelium during wound healing. J. Biol. Chem. 2000, 275, 2607–2612. [Google Scholar] [CrossRef] [PubMed]
- Aumailley, M.; Smyth, N. The role of laminins in basement membrane function. J. Anat. 1998, 193, 1–21. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miotto, M.; Gouveia, R.M.; Connon, C.J. Peptide Amphiphiles in Corneal Tissue Engineering. J. Funct. Biomater. 2015, 6, 687-707. https://doi.org/10.3390/jfb6030687
Miotto M, Gouveia RM, Connon CJ. Peptide Amphiphiles in Corneal Tissue Engineering. Journal of Functional Biomaterials. 2015; 6(3):687-707. https://doi.org/10.3390/jfb6030687
Chicago/Turabian StyleMiotto, Martina, Ricardo M. Gouveia, and Che J. Connon. 2015. "Peptide Amphiphiles in Corneal Tissue Engineering" Journal of Functional Biomaterials 6, no. 3: 687-707. https://doi.org/10.3390/jfb6030687
APA StyleMiotto, M., Gouveia, R. M., & Connon, C. J. (2015). Peptide Amphiphiles in Corneal Tissue Engineering. Journal of Functional Biomaterials, 6(3), 687-707. https://doi.org/10.3390/jfb6030687