
Eco-Interactions of Engineered Nanomaterials in the Marine Environment: Towards an Eco-Design Framework

 ,
, 
 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
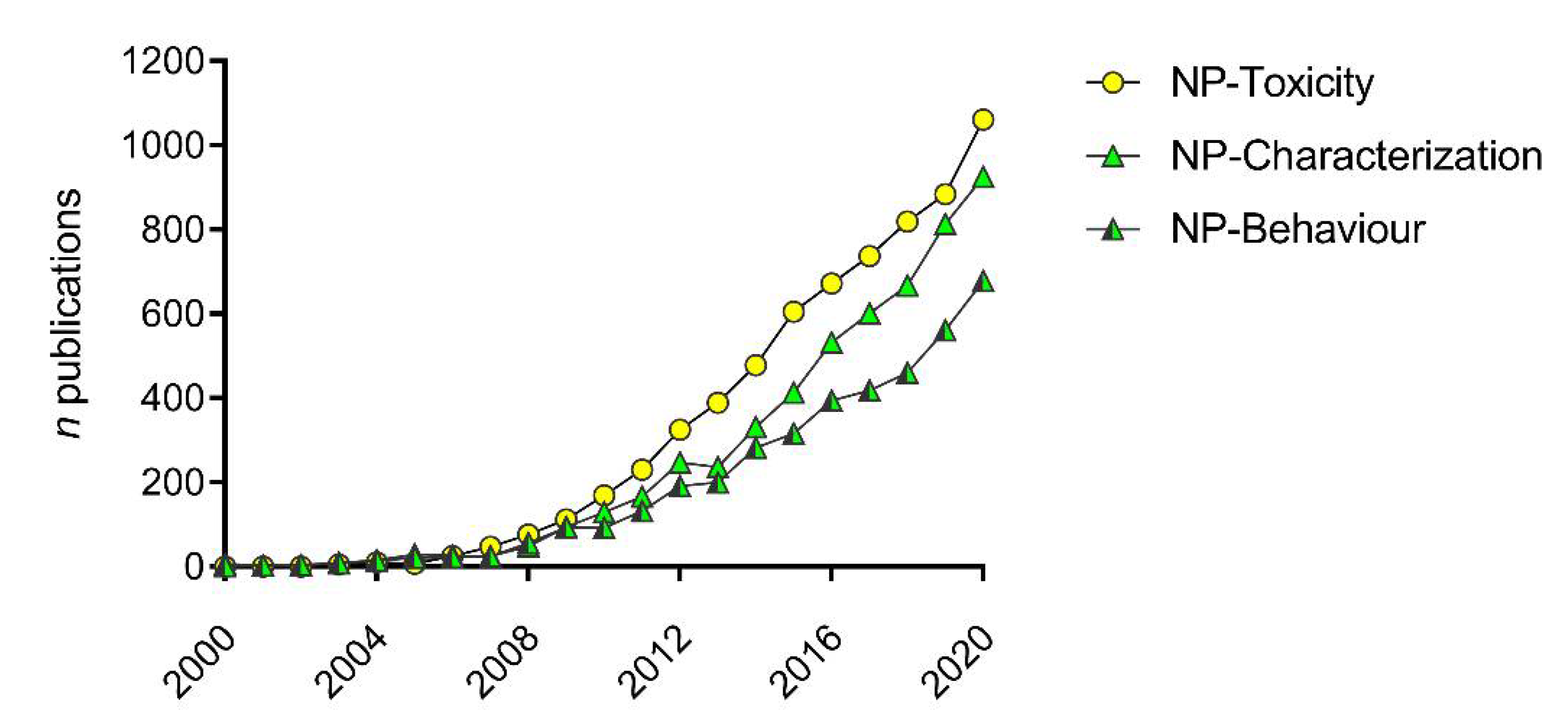
:1. Introduction
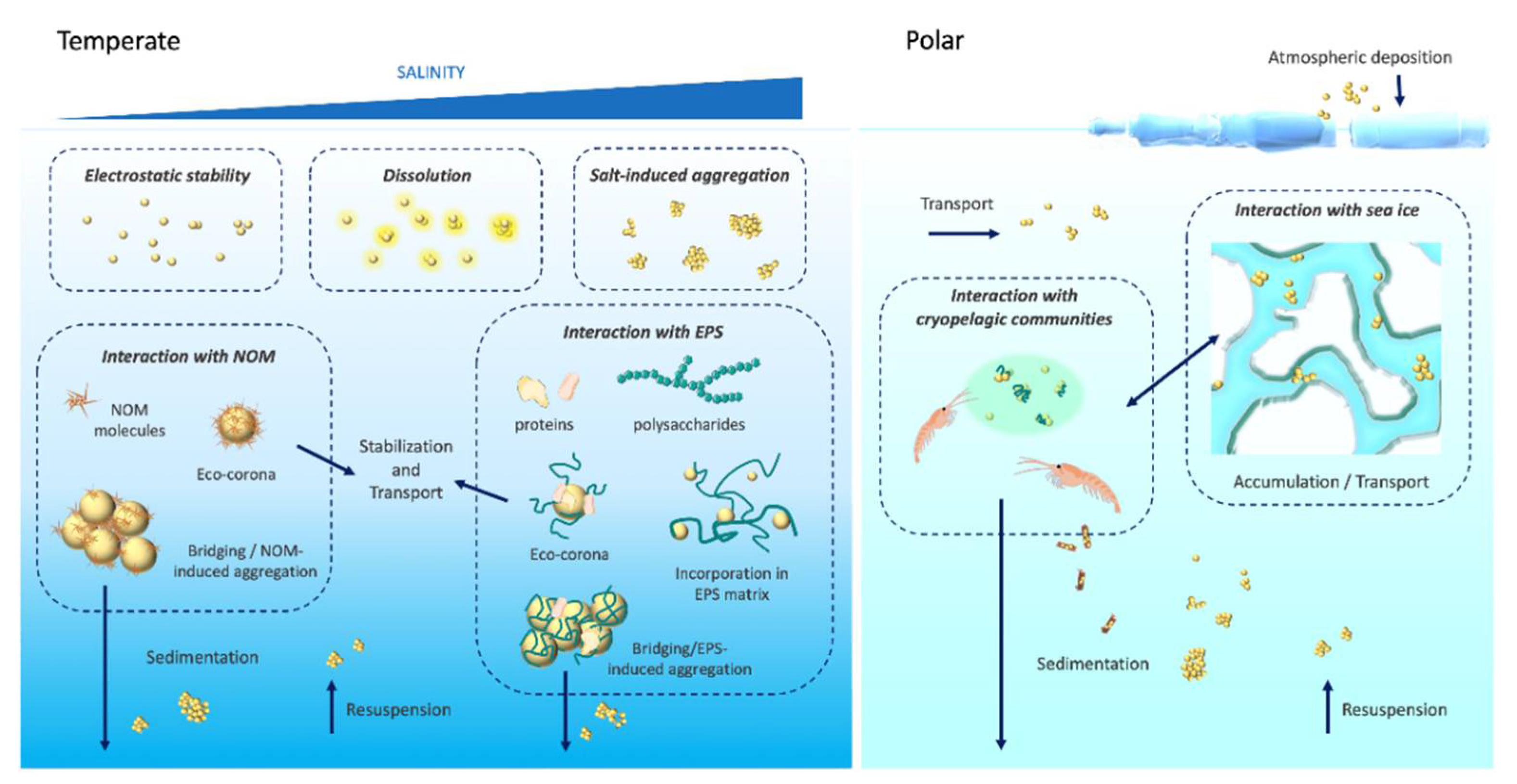
2. Behavior and Exposure Assessment of ENMs in Marine Waters
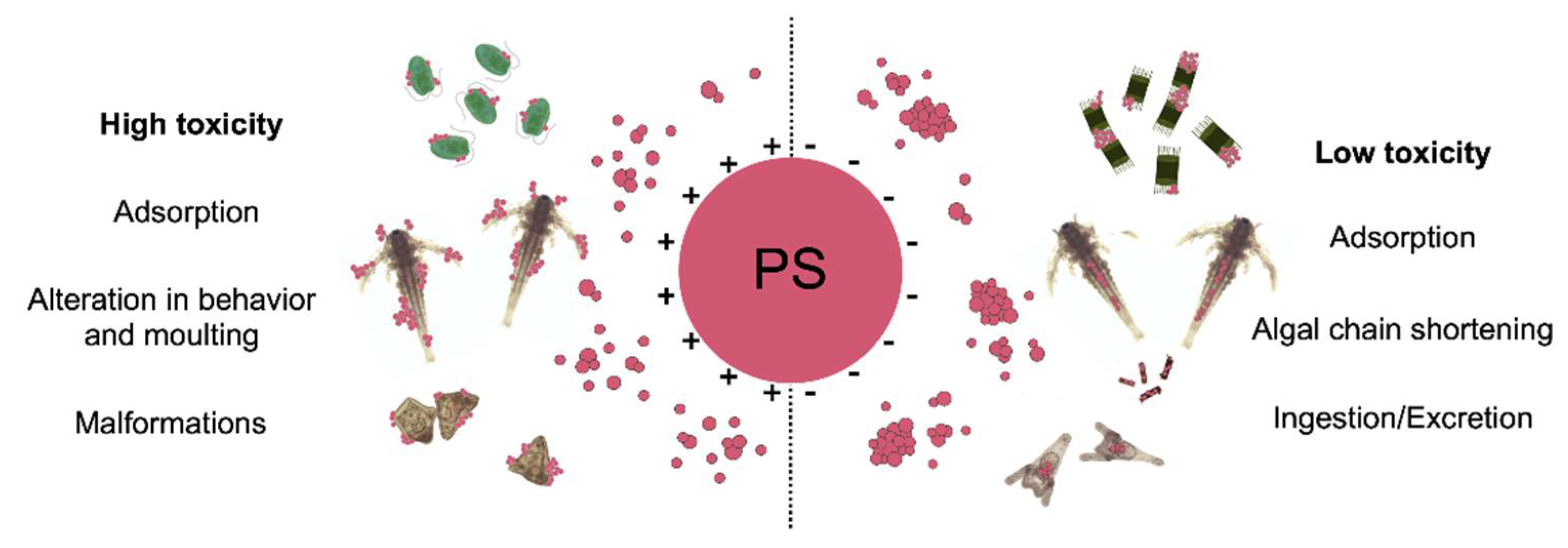
2.1. The Role of ENMs Surface Charges
2.2. The Interplay with Dissolved/Particulate NOM and Existing Chemicals in Seawater
2.3. Nano–Eco Interactions Scenarios Leading to Ecotoxicity
- Understand the interplay between ENMs, water media components and exposed organisms;
- Build ENM fate and transport models and identify major receiving compartments to which concentrate major efforts in terms of protection and eventual remediation actions [9].
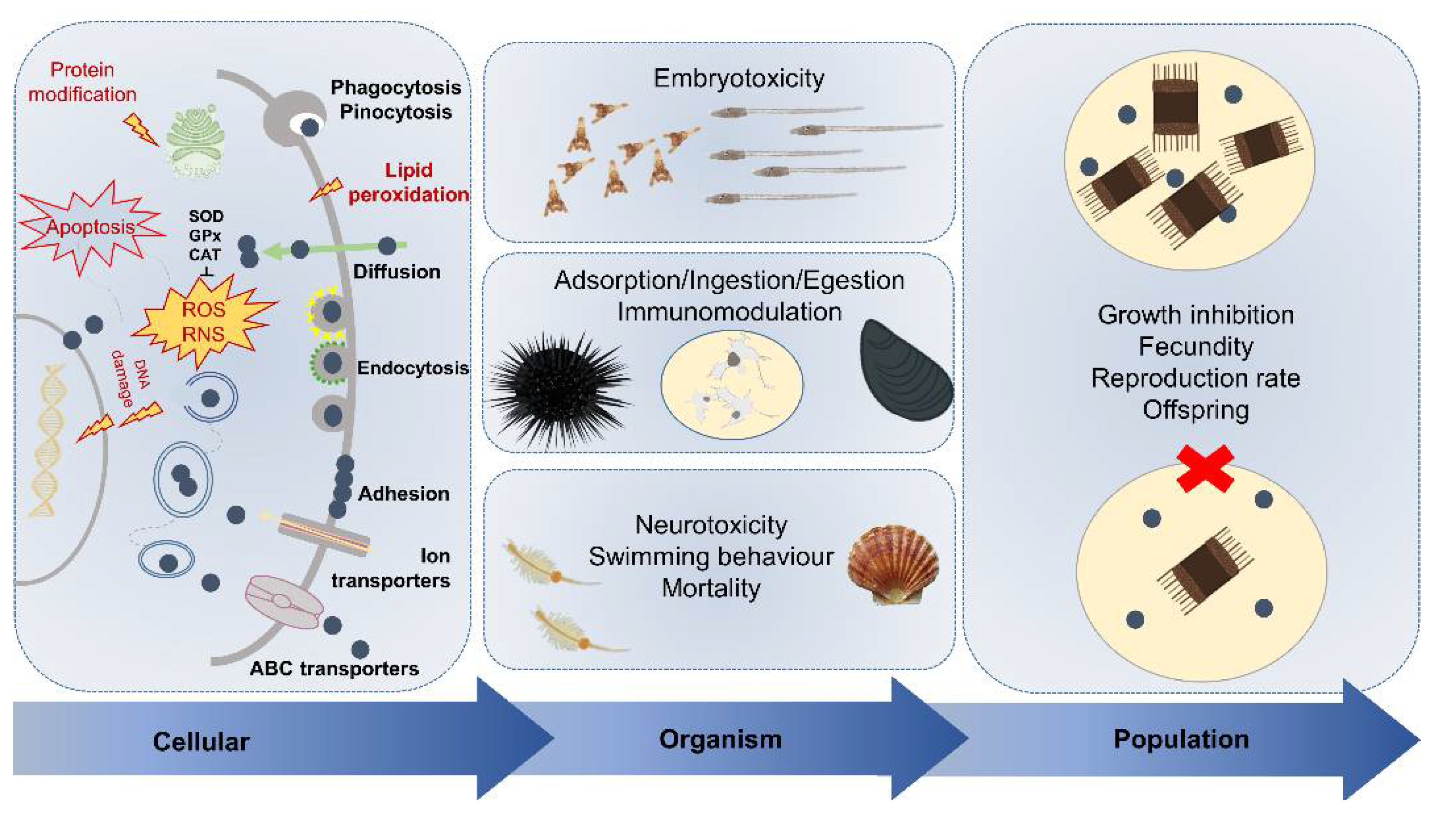
3. Current Knowledge on ENMs Marine Ecotoxicity
3.1. Cellular Uptake and Immunotoxicity
3.2. Neurotoxicity
3.3. Behavioral and Developmental Effects
3.4. From Impact on Single Species Up to Populations and Communities
4. Gaps to Be Filled
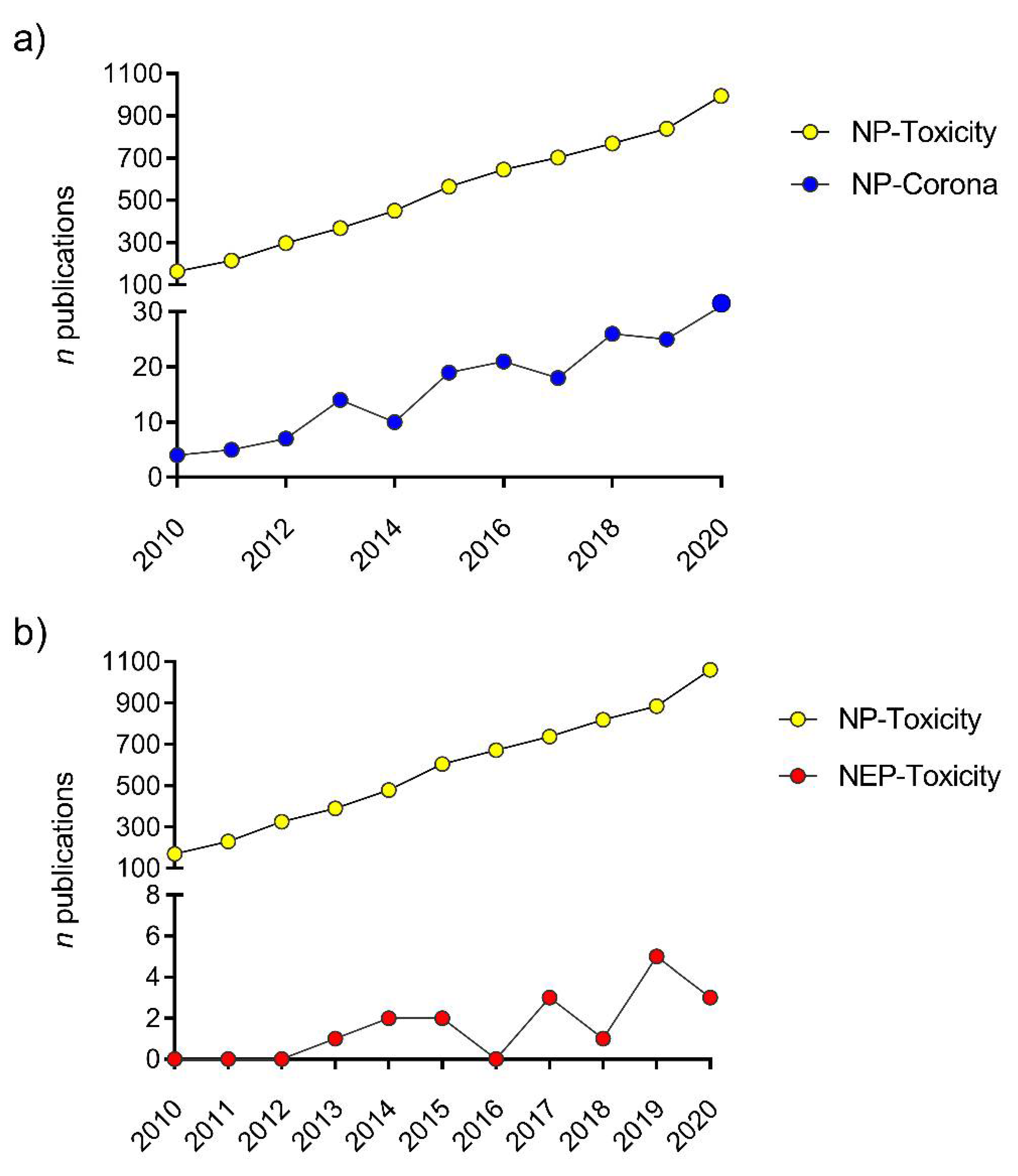
4.1. Real Exposure Scenarios: Nano-Enabled Products (NEPs)
4.2. Impact on Benthic Species Overlooked
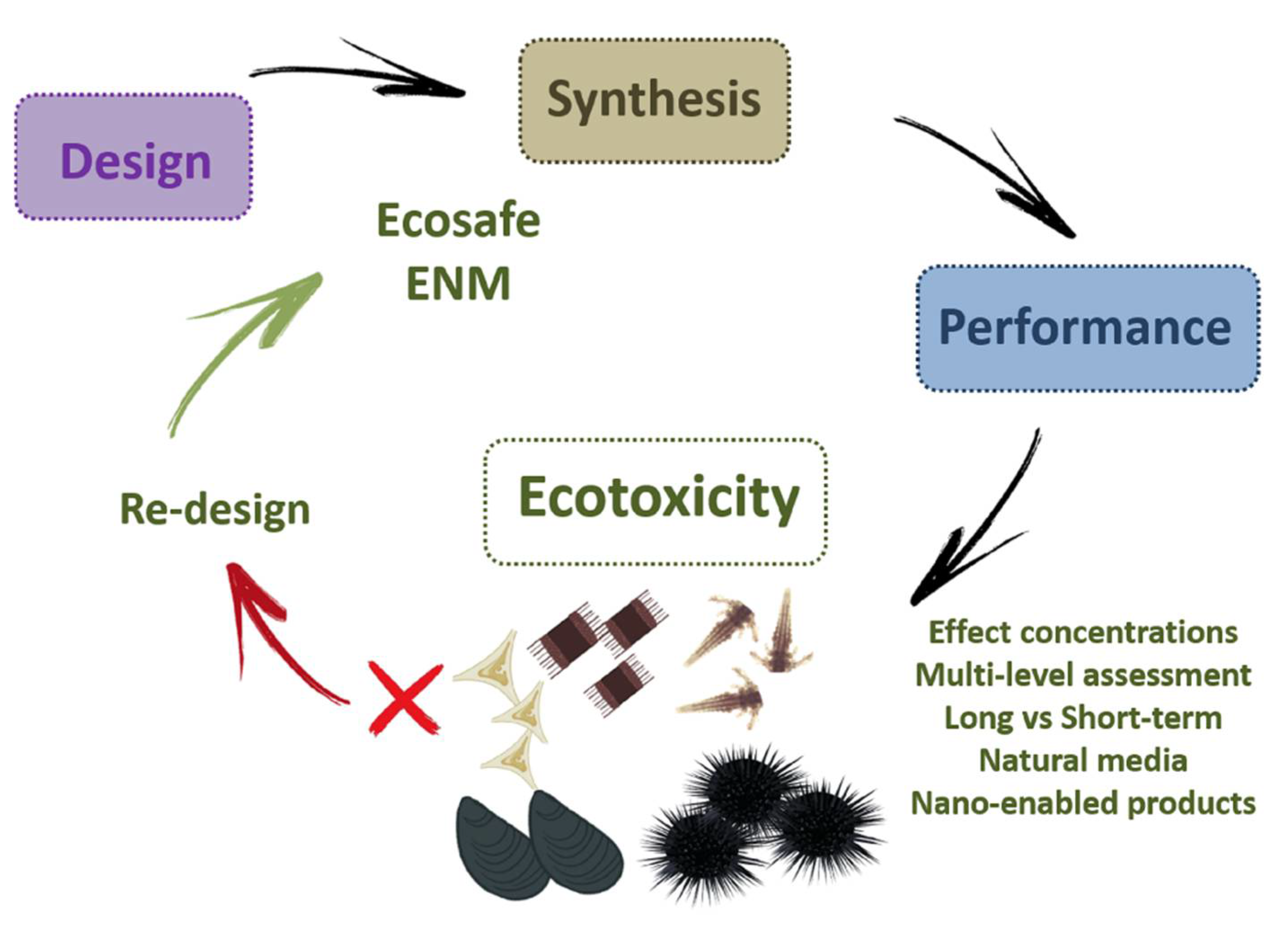
5. A Lesson to Learn: Using Ecotoxicity for a Safer ENMs Design (Eco-Design)
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barbier, E.B. Marine ecosystem services. Curr. Biol. 2017, 27, R507–R510. [Google Scholar] [CrossRef] [Green Version]
- UNESCO. The UN Decade of Ocean Science for Sustainable Development. Available online: https://en.unesco.org/ocean-decade (accessed on 17 June 2021).
- European Commission Marine Strategy Framework Directive. European Commission Directive 2008/56/EC. Available online: https://eur-lex.europa.eu/legal-content/en/ALL/?uri=CELEX%3A32008L0056 (accessed on 17 June 2021).
- European Commission Pathway to a Healthy Planet for All. EU Action Plan: Towards Zero Pollution for Air, Water and Soil 12.5.2021. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A52021DC0400&qid=1623311742827 (accessed on 17 June 2021).
- Landrigan, P.; Stegeman, J.; Fleming, L.; Allemand, D.; Anderson, D.; Backer, L.; Brucker-Davis, F.; Chevalier, N.; Corra, L.; Czerucka, D.; et al. Human health and ocean pollution. Ann. Glob. Health 2020, 151. [Google Scholar] [CrossRef]
- European Commission. 2011/696/EU commission recommendation of 18 October 2011 on the definition of nanomaterial. Off. J. Eur. Union 2011, L275, 38–54. Available online: https://eur-lex.europa.eu/eli/reco/2011/696/oj (accessed on 17 June 2021).
- Halden, R.U. Epistemology of contaminants of emerging concern and literature meta-analysis. J. Hazard. Mater. 2015, 282, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Corsi, I.; Cherr, G.N.; Lenihan, H.S.; Labille, J.; Hassellov, M.; Canesi, L.; Dondero, F.; Frenzilli, G.; Hristozov, D.; Puntes, V.; et al. Common strategies and technologies for the ecosafety assessment and design of nanomaterials entering the marine environment. ACS Nano 2014, 8, 9694–9709. [Google Scholar] [CrossRef] [Green Version]
- Garner, K.L.; Suh, S.; Keller, A.A. Assessing the risk of engineered nanomaterials in the environment: Development and application of the nanoFate model. Environ. Sci. Technol. 2017, 51, 5541–5551. [Google Scholar] [CrossRef] [Green Version]
- Mitrano, D.M.; Motellier, S.; Clavaguera, S.; Nowack, B. Review of nanomaterial aging and transformations through the life cycle of nano-enhanced products. Environ. Int. 2015, 77, 132–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.Y.; Bornhöft, N.A.; Hungerbühler, K.; Nowack, B. Dynamic probabilistic modeling of environmental emissions of engineered nanomaterials. Environ. Sci. Technol. 2016, 50, 4701–4711. [Google Scholar] [CrossRef] [PubMed]
- Brar, S.K.; Verma, M.; Tyagi, R.D.; Surampalli, R.Y. Engineered nanoparticles in wastewater and wastewater sludge—Evidence and impacts. Waste Manag. 2010, 30, 504–520. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Hernández, A.G.; Chiodoni, A.; Bocchini, S.; Vazquez-Duhalt, R. 3D printer waste, a new source of nanoplastic pollutants. Environ. Pollut. 2020, 267, 115609. [Google Scholar] [CrossRef]
- Hernandez, L.M.; Xu, E.G.; Larsson, H.C.E.; Tahara, R.; Maisuria, V.B.; Tufenkji, N. Plastic teabags release billions of microparticles and nanoparticles into tea. Environ. Sci. Technol. 2019, 53, 12300–12310. [Google Scholar] [CrossRef]
- Lambert, S.; Wagner, M. Formation of microscopic particles during the degradation of different polymers. Chemosphere 2016, 161, 510–517. [Google Scholar] [CrossRef]
- Plastic Europe. Plastic the Facts. 2019. Available online: https://www.plasticseurope.org/en/resources/publications/1804-plastics-facts-2019 (accessed on 17 June 2021).
- Lebreton, L.; Andrady, A. Future scenarios of global plastic waste generation and disposal. Palgrave Commun. 2019, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Frehland, S.; Kaegi, R.; Hufenus, R.; Mitrano, D.M. Long-term assessment of nanoplastic particle and microplastic fiber flux through a pilot wastewater treatment plant using metal-doped plastics. Water Res. 2020, 182, 115860. [Google Scholar] [CrossRef]
- Schirinzi, G.F.; Llorca, M.; Seró, R.; Moyano, E.; Barceló, D.; Abad, E.; Farré, M. Trace analysis of polystyrene microplastics in natural waters. Chemosphere 2019, 236, 124321. [Google Scholar] [CrossRef] [PubMed]
- Ter Halle, A.; Jeanneau, L.; Martignac, M.; Jardé, E.; Pedrono, B.; Brach, L.; Gigault, J. Nanoplastic in the North Atlantic subtropical gyre. Environ. Sci. Technol. 2017, 51, 13689–13697. [Google Scholar] [CrossRef] [PubMed]
- Bergami, E.; Pugnalini, S.; Vannuccini, M.L.; Manfra, L.; Faleri, C.; Savorelli, F.; Dawson, K.A.; Corsi, I. Long-term toxicity of surface-charged polystyrene nanoplastics to marine planktonic species Dunaliella tertiolecta and Artemia franciscana. Aquat. Toxicol. 2017, 189, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Della Torre, C.; Bergami, E.; Salvati, A.; Faleri, C.; Cirino, P.; Dawson, K.A.; Corsi, I. Accumulation and embryotoxicity of polystyrene nanoparticles at early stage of development of sea urchin embryos Paracentrotus lividus. Environ. Sci. Technol. 2014, 48, 12302–12311. [Google Scholar] [CrossRef]
- Manfra, L.; Rotini, A.; Bergami, E.; Grassi, G.; Faleri, C.; Corsi, I. Comparative ecotoxicity of polystyrene nanoparticles in natural seawater and reconstituted seawater using the rotifer Brachionus plicatilis. Ecotoxicol. Environ. Saf. 2017, 145, 557–563. [Google Scholar] [CrossRef]
- Kittler, S.; Greulich, C.; Diendorf, J.; Köller, M.; Epple, M. Toxicity of silver nanoparticles increases during storage because of slow Dissolution under release of silver ions. Chem. Mater. 2010, 22, 4548–4554. [Google Scholar] [CrossRef]
- Clément, L.; Hurel, C.; Marmier, N. Toxicity of TiO2 nanoparticles to cladocerans, algae, rotifers and plants—Effects of size and crystalline structure. Chemosphere 2013, 90, 1083–1090. [Google Scholar] [CrossRef]
- Johnson, M.S.; Ates, M.; Arslan, Z.; Farah, I.O.; Bogatu, C. Assessment of crystal morphology on uptake, particle dissolution, and toxicity of nanoscale titanium dioxide on Artemia salina. J. Nanotoxicol. Nanomed. 2017, 2, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Baalousha, M. Effect of nanomaterial and media physicochemical properties on nanomaterial aggregation kinetics. NanoImpact 2017, 6, 55–68. [Google Scholar] [CrossRef] [Green Version]
- Domingos, R.F.; Tufenkji, N.; Wilkinson, K.J. Aggregation of titanium dioxide nanoparticles: Role of a fulvic acid. Environ. Sci. Technol. 2009, 43, 1282–1286. [Google Scholar] [CrossRef] [Green Version]
- French, R.A.; Jacobson, A.R.; Kim, B.; Isley, S.L.; Penn, L.; Baveye, P.C. Influence of ionic strength, pH, and cation valence on aggregation kinetics of titanium dioxide nanoparticles. Environ. Sci. Technol. 2009, 43, 1354–1359. [Google Scholar] [CrossRef]
- Romanello, M.B.; Fidalgo de Cortalezzi, M.M. An experimental study on the aggregation of TiO2 nanoparticles under environmentally relevant conditions. Water Res. 2013, 47, 3887–3898. [Google Scholar] [CrossRef]
- Corsi, I.; Bergami, E.; Grassi, G. Behavior and bio-interactions of anthropogenic particles in marine environment for a more realistic ecological risk assessment. Front. Environ. Sci. 2020, 8, 60. [Google Scholar] [CrossRef]
- Tallec, K.; Huvet, A.; di Poi, C.; González-Fernández, C.; Lambert, C.; Petton, B.; le Goïc, N.; Berchel, M.; Soudant, P.; Paul-Pont, I. Nanoplastics impaired oyster free living stages, gametes and embryos. Environ. Pollut. 2018, 242, 1226–1235. [Google Scholar] [CrossRef] [Green Version]
- Della Torre, C.; Balbi, T.; Grassi, G.; Frenzilli, G.; Bernardeschi, M.; Smerilli, A.; Guidi, P.; Canesi, L.; Nigro, M.; Monaci, F.; et al. Titanium dioxide nanoparticles modulate the toxicological response to cadmium in the gills of Mytilus galloprovincialis. J. Hazard. Mater. 2015, 297, 92–100. [Google Scholar] [CrossRef]
- Bergami, E.; Bocci, E.; Vannuccini, M.L.; Monopoli, M.; Salvati, A.; Dawson, K.A.; Corsi, I. Nano-Sized Polystyrene affects feeding, behavior and physiology of brine shrimp Artemia franciscana larvae. Ecotoxicol. Environ. Saf. 2016, 123, 18–25. [Google Scholar] [CrossRef]
- Varó, I.; Perini, A.; Torreblanca, A.; Garcia, Y.; Bergami, E.; Vannuccini, M.L.; Corsi, I. Time-dependent effects of polystyrene nanoparticles in brine shrimp Artemia franciscana at physiological, biochemical and molecular levels. Sci. Total Environ. 2019, 675, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Hu, L.; Shi, H.; Ye, J.; Zhang, Y.; Kim, H. Effects of inorganic ions and natural organic matter on the aggregation of nanoplastics. Chemosphere 2018, 197, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Tiwari, E.; Khandelwal, N.; Darbha, G.K. Understanding the stability of nanoplastics in aqueous environments: Effect of ionic strength, temperature, dissolved organic matter, clay, and heavy metals. Environ. Sci. Nano 2019, 6, 2968–2976. [Google Scholar] [CrossRef]
- Wu, J.; Jiang, R.; Lin, W.; Ouyang, G. Effect of salinity and humic acid on the aggregation and toxicity of polystyrene nanoplastics with different functional groups and charges. Environ. Pollut. 2019, 245, 836–843. [Google Scholar] [CrossRef]
- Bergmann, M.; Mützel, S.; Primpke, S.; Tekman, M.B.; Trachsel, J.; Gerdts, G. White and wonderful? Microplastics prevail in snow from the alps to the arctic. Sci. Adv. 2019, 5, eaax1157. [Google Scholar] [CrossRef] [Green Version]
- Cózar, A.; Martí, E.; Duarte, C.M.; García-De-Lomas, J.; van Sebille, E.; Ballatore, T.J.; Eguíluz, V.M.; Ignacio González-Gordillo, J.; Pedrotti, M.L.; Echevarría, F.; et al. The Arctic Ocean as a dead end for floating plastics in the North Atlantic branch of the Thermohaline Circulation. Sci. Adv. 2017, 3, e1600582. [Google Scholar] [CrossRef] [Green Version]
- Obbard, R.W. Microplastics in polar regions: The role of long range transport. Curr. Opin. Environ. Sci. Health 2018, 1, 24–29. [Google Scholar] [CrossRef]
- Karl, M.; Leck, C.; Coz, E.; Heintzenberg, J. Marine nanogels as a source of atmospheric nanoparticles in the high Arctic. Geophys. Res. Lett. 2013, 40, 3738–3743. [Google Scholar] [CrossRef]
- Malakar, A.; Snow, D.D. Nanoparticles as sources of inorganic water pollutants. In Inorganic Pollutants in Water; Elsevier: Amsterdam, The Netherlands, 2020; pp. 337–370. [Google Scholar]
- Kumar, N.; Palmer, G.R.; Shah, V.; Walker, V.K. The effect of silver nanoparticles on seasonal change in arctic tundra bacterial and fungal assemblages. PLoS ONE 2014, 9, e99953. [Google Scholar] [CrossRef]
- Bergami, E.; Krupinski Emerenciano, A.; González-Aravena, M.; Cárdenas, C.A.; Hernández, P.; Silva, J.R.M.C.; Corsi, I. Polystyrene nanoparticles affect the innate immune system of the Antarctic sea urchin Sterechinus neumayeri. Polar Biol. 2019, 42, 743–757. [Google Scholar] [CrossRef]
- Petosa, A.R.; Jaisi, D.P.; Quevedo, I.R.; Elimelech, M.; Tufenkji, N. Aggregation and deposition of engineered nanomaterials in aquatic environments: Role of physicochemical interactions. Environ. Sci. Technol. 2010, 44, 6532–6549. [Google Scholar] [CrossRef] [Green Version]
- Peeken, I.; Primpke, S.; Beyer, B.; Gütermann, J.; Katlein, C.; Krumpen, T.; Bergmann, M.; Hehemann, L.; Gerdts, G. Arctic sea ice is an important temporal sink and means of transport for microplastic. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Dawson, A.L.; Kawaguchi, S.; King, C.K.; Townsend, K.A.; King, R.; Huston, W.M.; Bengtson Nash, S.M. Turning microplastics into nanoplastics through digestive fragmentation by Antarctic krill. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Bergami, E.; Manno, C.; Cappello, S.; Vannuccini, M.L.; Corsi, I. Nanoplastics affect moulting and faecal pellet sinking in Antarctic krill (Euphausia superba) juveniles. Environ. Int. 2020, 143, 105999. [Google Scholar] [CrossRef] [PubMed]
- Lespes, G.; Faucher, S.; Slaveykova, V.I. Natural nanoparticles, anthropogenic nanoparticles, where is the frontier? Front. Environ. Sci. 2020, 8, 71. [Google Scholar] [CrossRef]
- Tallec, K.; Blard, O.; González-Fernández, C.; Brotons, G.; Berchel, M.; Soudant, P.; Huvet, A.; Paul-Pont, I. Surface functionalization determines behavior of nanoplastic solutions in model aquatic environments. Chemosphere 2019, 225, 639–646. [Google Scholar] [CrossRef] [Green Version]
- Gigault, J.; Balaresque, M.; Tabuteau, H. Estuary-on-a-Chip: Unexpected results for the fate and transport of nanoparticles. Environ. Sci. Nano 2018, 5, 1231–1236. [Google Scholar] [CrossRef] [Green Version]
- Baalousha, M.; Afshinnia, K.; Guo, L. Natural organic matter composition determines the molecular nature of silver nanomaterial-NOM corona. Environ. Sci. Nano 2018, 5, 868–881. [Google Scholar] [CrossRef]
- Chetwynd, A.J.; Lynch, I. The rise of the nanomaterial metabolite corona, and emergence of the complete corona. Environ. Sci. Nano 2020, 7, 1041–1060. [Google Scholar] [CrossRef]
- Walczyk, D.; Bombelli, F.B.; Monopoli, M.P.; Lynch, I.; Dawson, K.A. What the cell “sees” in bionanoscience. J. Am. Chem. Soc. 2010, 132, 5761–5768. [Google Scholar] [CrossRef]
- Lynch, I.; Cedervall, T.; Lundqvist, M.; Cabaleiro-Lago, C.; Linse, S.; Dawson, K.A. The nanoparticle-protein complex as a biological entity; a complex fluids and surface science challenge for the 21st century. Adv. Colloid Interface Sci. 2007, 134, 167–174. [Google Scholar] [CrossRef]
- Lundqvist, M.; Stigler, J.; Elia, G.; Lynch, I.; Cedervall, T.; Dawson, K.A. Nanoparticle size and surface properties determine the protein corona with possible implications for biological impacts. Proc. Natl. Acad. Sci. USA 2008, 105, 14265–14270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahon, E.; Salvati, A.; Baldelli Bombelli, F.; Lynch, I.; Dawson, K.A. Designing the nanoparticle-biomolecule interface for “targeting and therapeutic delivery”. J. Control. Release 2012, 161, 164–174. [Google Scholar] [CrossRef]
- Oh, J.Y.; Kim, H.S.; Palanikumar, L.; Go, E.M.; Jana, B.; Park, S.A.; Kim, H.Y.; Kim, K.; Seo, J.K.; Kwak, S.K.; et al. Cloaking nanoparticles with protein corona shield for targeted drug delivery. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lynch, I.; Dawson, K.A.; Lead, J.R.; Valsami-Jones, E. Macromolecular Coronas and Their Importance in Nanotoxicology and Nanoecotoxicology. Front. Nanosci. 2014, 7, 127–156. [Google Scholar] [CrossRef]
- Westmeier, D.; Stauber, R.H.; Docter, D. The concept of bio-corona in modulating the toxicity of engineered nanomaterials (ENM). Toxicol. Appl. Pharmacol. 2016, 299, 53–57. [Google Scholar] [CrossRef]
- Walkey, C.D.; Olsen, J.B.; Song, F.; Liu, R.; Guo, H.; Olsen, D.W.H.; Cohen, Y.; Emili, A.; Chan, W.C.W. Protein corona fingerprinting predicts the cellular interaction of gold and silver nanoparticles. ACS Nano 2014, 8, 2439–2455. [Google Scholar] [CrossRef]
- Ritz, S.; Schöttler, S.; Kotman, N.; Baier, G.; Musyanovych, A.; Kuharev, J.; Landfester, K.; Schild, H.; Jahn, O.; Tenzer, S.; et al. Protein corona of nanoparticles: Distinct proteins regulate the cellular uptake. Biomacromolecules 2015, 16, 1311–1321. [Google Scholar] [CrossRef]
- Fleischer, C.C.; Payne, C.K. Secondary structure of corona proteins determines the cell surface receptors used by nanoparticles. J. Phys. Chem. B 2014, 118, 14017–14026. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, C.C.; Payne, C.K. Nanoparticle—Cell interactions: Molecular structure of the protein corona and cellular outcomes. Acc. Chem. Res. 2015, 47, 2651–2659. [Google Scholar] [CrossRef]
- Wan, S.; Kelly, P.M.; Mahon, E.; Stöckmann, H.; Rudd, P.M.; Caruso, F.; Dawson, K.A.; Yan, Y.; Monopoli, M.P. The “sweet” side of the protein corona: Effects of glycosylation on nanoparticle-cell interactions. ACS Nano 2015, 9, 2157–2166. [Google Scholar] [CrossRef] [Green Version]
- Adeleye, A.S.; Keller, A.A. Long-term colloidal stability and metal leaching of single wall carbon nanotubes: Effect of temperature and extracellular polymeric substances. Water Res. 2014, 49, 236–250. [Google Scholar] [CrossRef] [Green Version]
- Surette, M.C.; Nason, J.A. Effects of surface coating character and interactions with natural organic matter on the colloidal stability of gold nanoparticles. Environ. Sci. Nano 2016, 3, 1144–1152. [Google Scholar] [CrossRef]
- Surette, M.C.; Nason, J.A. Nanoparticle aggregation in a freshwater river: The role of engineered surface coatings. Environ. Sci. Nano 2019, 6, 540–553. [Google Scholar] [CrossRef]
- Barbero, F.; Mayall, C.; Drobne, D.; Saiz-Poseu, J.; Bastús, N.G.; Puntes, V. Formation and evolution of the nanoparticle environmental corona: The case of Au and humic acid. Sci. Total Environ. 2021, 768, 144792. [Google Scholar] [CrossRef]
- Grunér, M.S.; Kauscher, U.; Linder, M.B.; Monopoli, M.P. An environmental route of exposure affects the formation of nanoparticle coronas in blood plasma. J. Proteom. 2016, 137, 52–58. [Google Scholar] [CrossRef]
- Nasser, F.; Constantinou, J.; Lynch, I. Nanomaterials in the environment acquire an “eco-corona” impacting their toxicity to Daphnia magna—A call for updating toxicity testing policies. Proteomics 2020, 20, e1800412. [Google Scholar] [CrossRef] [Green Version]
- Canesi, L.; Corsi, I. Effects of nanomaterials on marine invertebrates. Sci. Total Environ. 2016, 565, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Canesi, L.; Balbi, T.; Fabbri, R.; Salis, A.; Damonte, G.; Volland, M.; Blasco, J. Biomolecular coronas in invertebrate species: Implications in the environmental impact of nanoparticles. NanoImpact 2017, 8, 89–98. [Google Scholar] [CrossRef]
- Wheeler, K.E.; Chetwynd, A.J.; Fahy, K.M.; Hong, B.S.; Tochihuitl, J.A.; Foster, L.A.; Lynch, I. Environmental dimensions of the protein corona. Nat. Nanotechnol. 2021, 16, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Ellis, L.J.A.; Lynch, I. Mechanistic insights into toxicity pathways induced by nanomaterials in: Daphnia magna from analysis of the composition of the acquired protein corona. Environ. Sci. Nano 2020, 7, 3343–3359. [Google Scholar] [CrossRef]
- Docter, D.; Westmeier, D.; Markiewicz, M.; Stolte, S.; Knauer, S.K.; Stauber, R.H. The Nanoparticle biomolecule corona: Lessons learned—Challenge accepted? Chem. Soc. Rev. 2015, 44, 6094–6121. [Google Scholar] [CrossRef] [Green Version]
- Nason, J.A.; McDowell, S.A.; Callahan, T.W. Effects of natural organic matter type and concentration on the aggregation of citrate-stabilized gold nanoparticles. J. Environ. Monit. 2012, 14, 1885–1892. [Google Scholar] [CrossRef]
- Vance, M.E.; Kuiken, T.; Vejerano, E.P.; McGinnis, S.P.; Hochella, M.F.; Hull, D.R. Nanotechnology in the real world: Redeveloping the nanomaterial consumer products inventory. Beilstein J. Nanotechnol. 2015, 6, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Mohd Zaidi, N.F.; Valsami-Jones, E.; Kreft, J.U. Time-resolved toxicity study reveals the dynamic interactions between uncoated silver nanoparticles and bacteria. Nanotoxicology 2017, 11, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Ratte, H.T. Bioaccumulation and toxicity of silver compounds: A review. Environ. Toxicol. Chem. 1999, 18, 89–108. [Google Scholar] [CrossRef]
- Yang, X.; Gondikas, A.P.; Marinakos, S.M.; Auffan, M.; Liu, J.; Hsu-Kim, H.; Meyer, J.N. Mechanism of silver nanoparticle toxicity is dependent on dissolved silver and surface coating in Caenorhabditis elegans. Environ. Sci. Technol. 2012, 46, 1119–1127. [Google Scholar] [CrossRef]
- Asadi Dokht Lish, R.; Johari, S.A.; Sarkheil, M.; Yu, I.J. On how environmental and experimental conditions affect the results of aquatic nanotoxicology on brine shrimp (Artemia salina): A case of silver nanoparticles toxicity. Environ. Pollut. 2019, 255, 113358. [Google Scholar] [CrossRef]
- McLaughlin, J.; Bonzongo, J.C.J. Effects of natural water chemistry on nanosilver behavior and toxicity to Ceriodaphnia dubia and Pseudokirchneriella subcapitata. Environ. Toxicol. Chem. 2012, 31, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Gondikas, A.P.; Morris, A.; Reinsch, B.C.; Marinakos, S.M.; Lowry, G.V.; Hsu-Kim, H. Cysteine-induced modifications of zero-valent silver nanomaterials: Implications for particle surface chemistry, aggregation, dissolution, and silver speciation. Environ. Sci. Technol. 2012, 46, 7037–7045. [Google Scholar] [CrossRef] [PubMed]
- Lekamge, S.; Miranda, A.F.; Trestrail, C.; Pham, B.; Ball, A.S.; Shukla, R.; Nugegoda, D. The toxicity of nonaged and aged coated silver nanoparticles to freshwater alga Raphidocelis subcapitata. Environ. Toxicol. Chem. 2019, 38, 2371–2382. [Google Scholar] [CrossRef]
- Schiavo, S.; Duroudier, N.; Bilbao, E.; Mikolaczyk, M.; Schäfer, J.; Cajaraville, M.P.; Manzo, S. Effects of PVP/PEI coated and uncoated silver NPs and PVP/PEI coating agent on three species of marine microalgae. Sci. Total Environ. 2017, 577, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Harper, B.J.; Harper, S.L. Differential dissolution and toxicity of surface functionalized silver nanoparticles in small-scale microcosms: Impacts of community complexity. Environ. Sci. Nano 2017, 4, 359–372. [Google Scholar] [CrossRef]
- Levard, C.; Hotze, E.M.; Colman, B.P.; Dale, A.L.; Truong, L.; Yang, X.Y.; Bone, A.J.; Brown, G.E.; Tanguay, R.L.; di Giulio, R.T.; et al. Sulfidation of silver nanoparticles: Natural antidote to their toxicity. Environ. Sci. Technol. 2013, 47, 13440–13448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prosposito, P.; Burratti, L.; Bellingeri, A.; Protano, G.; Faleri, C.; Corsi, I.; Battocchio, C.; Iucci, G.; Tortora, L.; Secchi, V.; et al. Bifunctionalized silver nanoparticles as Hg2+ plasmonic sensor in water: Synthesis, characterizations, and ecosafety. Nanomaterials 2019, 9, 1353. [Google Scholar] [CrossRef] [Green Version]
- Levard, C.; Hotze, E.M.; Lowry, G.V.; Brown, G.E. Environmental transformations of silver nanoparticles: Impact on stability and toxicity. Environ. Sci. Technol. 2012, 46, 6900–6914. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Su, M.; Wang, X.; Zou, X.; Sun, X.; Shi, J.; Zhang, H. Environmental fate and behavior of silver nanoparticles in natural estuarine systems. J. Environ. Sci. (China) 2020, 88, 248–259. [Google Scholar] [CrossRef]
- Sigg, L.; Lindauer, U. Silver nanoparticle dissolution in the presence of ligands and of hydrogen peroxide. Environ. Pollut. 2015, 206, 582–587. [Google Scholar] [CrossRef]
- Gunsolus, I.L.; Mousavi, M.P.S.; Hussein, K.; Bühlmann, P.; Haynes, C.L. Effects of humic and fulvic acids on silver nanoparticle stability, dissolution, and toxicity. Environ. Sci. Technol. 2015, 49, 8078–8086. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.J.; Chappell, M.A.; Bednar, A.J.; Ryan, A.C.; Laird, J.G.; Stanley, J.K.; Steevens, J.A. Impact of organic carbon on the stability and toxicity of fresh and stored silver nanoparticles. Environ. Sci. Technol. 2012, 46, 10772–10780. [Google Scholar] [CrossRef]
- Malysheva, A.; Voelcker, N.; Holm, P.E.; Lombi, E. Unraveling the complex behavior of AgNPs driving NP-cell interactions and toxicity to algal cells. Environ. Sci. Technol. 2016, 50, 12455–12463. [Google Scholar] [CrossRef]
- Kleiven, M.; Macken, A.; Oughton, D.H. Growth inhibition in Raphidocelis subcapita—Evidence of nanospecific toxicity of silver nanoparticles. Chemosphere 2019, 221, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Fadare, O.O.; Wan, B.; Liu, K.; Yang, Y.; Zhao, L.; Guo, L. Eco-corona vs. protein corona: Effects of humic substances on corona formation and nanoplastic particle toxicity in Daphnia magna. Environ. Sci. Technol. 2020, 54, 8001–8009. [Google Scholar] [CrossRef]
- Fadare, O.O.; Wan, B.; Guo, L.-H.; Xin, Y.; Qin, W.; Yang, Y. Humic acid alleviates the toxicity of polystyrene nanoplastic particles to Daphnia magna. Environ. Sci. Nano 2019, 6, 1466–1477. [Google Scholar] [CrossRef]
- Noventa, S.; Rowe, D.; Galloway, T. Mitigating effect of organic matter on the vivo toxicity of metal oxide nanoparticles in the marine environment. Environ. Sci. Nano 2018, 5, 1764–1777. [Google Scholar] [CrossRef] [Green Version]
- Morelli, E.; Gabellieri, E.; Bonomini, A.; Tognotti, D.; Grassi, G.; Corsi, I. TiO2 nanoparticles in seawater: Aggregation and interactions with the green alga Dunaliella tertiolecta. Ecotoxicol. Environ. Saf. 2018, 148, 184–193. [Google Scholar] [CrossRef]
- Grassi, G.; Gabellieri, E.; Cioni, P.; Paccagnini, E.; Faleri, C.; Lupetti, P.; Corsi, I.; Morelli, E. Interplay between extracellular polymeric substances (EPS) from a marine diatom and model nanoplastic through eco-corona formation. Sci. Total Environ. 2020, 725, 138457. [Google Scholar] [CrossRef]
- Canesi, L.; Ciacci, C.; Fabbri, R.; Balbi, T.; Salis, A.; Damonte, G.; Cortese, K.; Caratto, V.; Monopoli, M.P.; Dawson, K.; et al. Interactions of cationic polystyrene nanoparticles with marine bivalve hemocytes in a physiological environment: Role of soluble hemolymph proteins. Environ. Res. 2016, 150, 73–81. [Google Scholar] [CrossRef]
- Marques-Santos, L.F.; Grassi, G.; Bergami, E.; Faleri, C.; Balbi, T.; Salis, A.; Damonte, G.; Canesi, L.; Corsi, I. Cationic polystyrene nanoparticle and the sea urchin immune system: Biocorona formation, cell toxicity, and multixenobiotic resistance phenotype. Nanotoxicology 2018, 12, 847–867. [Google Scholar] [CrossRef]
- Grassi, G.; Landi, C.; Della Torre, C.; Bergami, E.; Bini, L.; Corsi, I. Proteomic profile of the hard corona of charged polystyrene nanoparticles exposed to sea urchin Paracentrotus lividus coelomic fluid highlights potential drivers of toxicity. Environ. Sci. Nano 2019. [Google Scholar] [CrossRef]
- Limbach, L.K.; Wick, P.; Manser, P.; Grass, R.N.; Bruinink, A.; Stark, W.J. Exposure of engineered nanoparticles to human lung epithelial cells: Influence of chemical composition and catalytic activity on oxidative stress. Environ. Sci. Technol. 2007, 41, 4158–4163. [Google Scholar] [CrossRef] [PubMed]
- Naasz, S.; Altenburger, R.; Kühnel, D. Environmental mixtures of nanomaterials and chemicals: The trojan-horse phenomenon and its relevance for ecotoxicity. Sci. Total Environ. 2018, 635, 1170–1181. [Google Scholar] [CrossRef]
- Bellingeri, A.; Bergami, E.; Grassi, G.; Faleri, C.; Redondo-Hasselerharm, P.; Koelmans, A.A.; Corsi, I. Combined effects of nanoplastics and copper on the freshwater alga Raphidocelis subcapitata. Aquat. Toxicol. 2019, 210, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Velzeboer, I.; Kwadijk, C.J.A.F.; Koelmans, A.A. Strong sorption of PCBs to nanoplastics, microplastics, carbon nanotubes, and fullerenes. Environ. Sci. Technol. 2014, 48, 4869–4876. [Google Scholar] [CrossRef]
- Lu, J.; Tian, S.; Lv, X.; Chen, Z.; Chen, B.; Zhu, X.; Cai, Z. TiO2 nanoparticles in the marine environment: Impact on the toxicity of phenanthrene and Cd2+ to marine zooplankton Artemia salina. Sci. Total Environ. 2018, 615, 375–380. [Google Scholar] [CrossRef]
- Rocco, L.; Santonastaso, M.; Nigro, M.; Mottola, F.; Costagliola, D.; Bernardeschi, M.; Guidi, P.; Lucchesi, P.; Scarcelli, V.; Corsi, I.; et al. Genomic and chromosomal damage in the marine mussel Mytilus galloprovincialis: Effects of the combined exposure to titanium dioxide nanoparticles and cadmium chloride. Mar. Environ. Res. 2015, 111, 144–148. [Google Scholar] [CrossRef]
- Banni, M.; Sforzini, S.; Balbi, T.; Corsi, I.; Viarengo, A.; Canesi, L. Combined effects of n-TiO2 and 2,3,7,8-TCDD in Mytilus galloprovincialis digestive gland: A transcriptomic and immunohistochemical study. Environ. Res. 2016, 145, 135–144. [Google Scholar] [CrossRef]
- Canesi, L.; Frenzilli, G.; Balbi, T.; Bernardeschi, M.; Ciacci, C.; Corsolini, S.; Della Torre, C.; Fabbri, R.; Faleri, C.; Focardi, S.; et al. Interactive effects of n-TiO2 and 2,3,7,8-TCDD on the marine bivalve Mytilus galloprovincialis. Aquat. Toxicol. 2014, 153, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Zhang, Y.; Song, C.; Zhu, X.; Xing, B. Bioaccumulation and biotransformation of polybrominated diphenyl ethers in the marine bivalve (Scapharca subcrenata): Influence of titanium dioxide nanoparticles. Mar. Pollut. Bull. 2015, 90, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Farkas, J.; Bergum, S.; Nilsen, E.W.; Olsen, A.J.; Salaberria, I.; Ciesielski, T.M.; Baczek, T.; Konieczna, L.; Salvenmoser, W.; Jenssen, B.M. The Impact of TiO2 Nanoparticles on Uptake and Toxicity of Benzo(a)Pyrene in the Blue Mussel (Mytilus edulis). Sci. Total Environ. 2015, 511, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Zhou, J.; Cai, Z. The Toxicity and Oxidative Stress of TiO2 Nanoparticles in Marine Abalone (Haliotis diversicolor supertexta). Mar. Pollut. Bull. 2011, 63, 334–338. [Google Scholar] [CrossRef]
- Shi, W.; Han, Y.; Guo, C.; Zhao, X.; Liu, S.; Su, W.; Zha, S.; Wang, Y.; Liu, G. Immunotoxicity of Nanoparticle nTiO2 to a Commercial Marine Bivalve Species, Tegillarca granosa. Fish Shellfish. Immunol. 2017, 66, 300–306. [Google Scholar] [CrossRef]
- Della Torre, C.; Buonocore, F.; Frenzilli, G.; Corsolini, S.; Brunelli, A.; Guidi, P.; Kocan, A.; Mariottini, M.; Mottola, F.; Nigro, M.; et al. Influence of Titanium Dioxide Nanoparticles on 2,3,7,8-Tetrachlorodibenzo-p-Dioxin Bioconcentration and Toxicity in the Marine Fish European Sea Bass (Dicentrarchus labrax). Environ. Pollut. 2015, 196, 185–193. [Google Scholar] [CrossRef]
- Vannuccini, M.L.; Grassi, G.; Leaver, M.J.; Corsi, I. Combination Effects of Nano-TiO2 and 2,3,7,8-Tetrachlorodibenzo-p-Dioxin (TCDD) on Biotransformation Gene Expression in the Liver of European Sea Bass Dicentrarchus labrax. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 176, 71–78. [Google Scholar] [CrossRef]
- Kulacki, K.J.; Cardinale, B.J.; Keller, A.A.; Bier, R.; Dickson, H. How Do Stream Organisms Respond to, and Influence, the Concentration of Titanium Dioxide Nanoparticles? A Mesocosm Study with Algae and Herbivores. Environ. Toxicol. Chem. 2012, 31, 2414–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jovanović, B.; Bezirci, G.; Çağan, A.S.; Coppens, J.; Levi, E.E.; Oluz, Z.; Tuncel, E.; Duran, H.; Beklioğlu, M. Food Web Effects of Titanium Dioxide Nanoparticles in an Outdoor Freshwater Mesocosm Experiment. Nanotoxicology 2016, 10, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Rist, S.; Hartmann, N.B. Aquatic ecotoxicity of microplastics and nanoplastics: Lessons learned from engineered nanomaterials. In Handbook of Environmental Chemistry; Springer: Berlin/Heidelberg, Germany, 2018; Volume 58, pp. 25–49. [Google Scholar] [CrossRef]
- Petersen, E.J.; Diamond, S.A.; Kennedy, A.J.; Goss, G.G.; Ho, K.; Lead, J.; Hanna, S.K.; Hartmann, N.B.; Hund-Rinke, K.; Mader, B.; et al. Adapting OECD Aquatic Toxicity Tests for Use with Manufactured Nanomaterials: Key Issues and Consensus Recommendations. Environ. Sci. Technol. 2015, 49, 9532–9547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johari, S.A.; Rasmussen, K.; Gulumian, M.; Ghazi-Khansari, M.; Tetarazako, N.; Kashiwada, S.; Asghari, S.; Park, J.W.; Yu, I.J. Introducing a New Standardized Nanomaterial Environmental Toxicity Screening Testing Procedure, ISO/TS 20787: Aquatic Toxicity Assessment of Manufactured Nanomaterials in Saltwater Lakes Using Artemia Sp. Nauplii. Toxicol. Mech. Methods 2019, 29, 95–109. [Google Scholar] [CrossRef]
- Kach, D.J.; Ward, J.E. The Role of Marine Aggregates in the Ingestion of Picoplankton-Size Particles by Suspension-Feeding Molluscs. Mar. Biol. 2008, 153, 797–805. [Google Scholar] [CrossRef]
- Abtahi, S.M.H.; Trevisan, R.; di Giulio, R.; Murphy, C.J.; Saleh, N.B.; Vikesland, P.J. Implications of Aspect Ratio on the Uptake and Nanotoxicity of Gold Nanomaterials. NanoImpact 2019, 14, 100153. [Google Scholar] [CrossRef]
- Bouallegui, Y.; ben Younes, R.; Bellamine, H.; Oueslati, R. Histopathology and Analyses of Inflammation Intensity in the Gills of Mussels Exposed to Silver Nanoparticles: Role of Nanoparticle Size, Exposure Time, and Uptake Pathways. Toxicol. Mech. Methods 2017, 27, 582–591. [Google Scholar] [CrossRef]
- Eliso, M.C.; Bergami, E.; Manfra, L.; Spagnuolo, A.; Corsi, I. Toxicity of Nanoplastics during the Embryogenesis of the Ascidian Ciona robusta (Phylum Chordata). Nanotoxicology 2020, 14, 1415–1431. [Google Scholar] [CrossRef]
- Bellingeri, A.; Casabianca, S.; Capellacci, S.; Faleri, C.; Paccagnini, E.; Lupetti, P.; Koelmans, A.A.; Penna, A.; Corsi, I. Impact of Polystyrene Nanoparticles on Marine Diatom Skeletonema marinoi Chain Assemblages and Consequences on Their Ecological Role in Marine Ecosystems. Environ. Pollut. 2020, 262, 114268. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Chi, T.; Li, Z.; Wang, J.; Yu, M.; Wu, M.; Zhou, H. Combined Effect of Polystyrene Plastics and Triphenyltin Chloride on the Green Algae Chlorella pyrenoidosa. Environ. Sci. Pollut. Res. 2019, 26, 15011–15018. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.J.; Bennett, S.; Keller, A.A.; Pease, S.; Lenihan, H.S. TiO2 Nanoparticles Are Phototoxic to Marine Phytoplankton. PLoS ONE 2012, 7, e30321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sendra, M.; Staffieri, E.; Yeste, M.P.; Moreno-Garrido, I.; Gatica, J.M.; Corsi, I.; Blasco, J. Are the Primary Characteristics of Polystyrene Nanoplastics Responsible for Toxicity and Ad/Absorption in the Marine Diatom Phaeodactylum tricornutum? Environ. Pollut. 2019, 249, 610–619. [Google Scholar] [CrossRef]
- Handy, R.D.; Owen, R.; Valsami-Jones, E. The Ecotoxicology of Nanoparticles and Nanomaterials: Current Status, Knowledge Gaps, Challenges, and Future Needs. Ecotoxicology 2008, 17, 315–325. [Google Scholar] [CrossRef]
- Lesniak, A.; Fenaroli, F.; Monopoli, M.P.; Åberg, C.; Dawson, K.A.; Salvati, A. Effects of the Presence or Absence of a Protein Corona on Silica Nanoparticle Uptake and Impact on Cells. ACS Nano 2012, 6, 5845–5857. [Google Scholar] [CrossRef]
- Monopoli, M.P.; Åberg, C.; Salvati, A.; Dawson, K.A. Biomolecular Coronas Provide the Biological Identity of Nanosized Materials. Nat. Nanotechnol. 2012, 7, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.N. Do Nanoparticles Present Ecotoxicological Risks for the Health of the Aquatic Environment? Environ. Int. 2006, 32, 967–976. [Google Scholar] [CrossRef]
- Firdessa, R.; Oelschlaeger, T.A.; Moll, H. Identification of Multiple Cellular Uptake Pathways of Polystyrene Nanoparticles and Factors Affecting the Uptake: Relevance for Drug Delivery Systems. Eur. J. Cell Biol. 2014, 93, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Luoma, S.N.; Khan, F.R.; Croteau, M.N. Bioavailability and Bioaccumulation of Metal-Based Engineered Nanomaterials in Aquatic Environments. Concepts and Processes. In Frontiers of Nanoscience; Elsevier: Amsterdam, The Netherlands, 2014; Volume 7, pp. 157–193. [Google Scholar] [CrossRef]
- Roiter, Y.; Ornatska, M.; Rammohan, A.R.; Balakrishnan, J.; Heine, D.R.; Minko, S. Interaction of Nanoparticles with Lipid Membrane. Nano Lett. 2008, 8, 941–944. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Barnoud, J.; Monticelli, L. Polystyrene Nanoparticles Perturb Lipid Membranes. J. Phys. Chem. Lett. 2014, 5, 241–246. [Google Scholar] [CrossRef]
- Rocha, T.L.; Gomes, T.; Sousa, V.S.; Mestre, N.C.; Bebianno, M.J. Ecotoxicological Impact of Engineered Nanomaterials in Bivalve Molluscs: An Overview. Mar. Environ. Res. 2015, 111, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, T.R.; Chi, R.J.; Parrow, M.W.; Ringwood, A.H. Cellular Bioreactivity of Micro- and Nano-Plastic Particles in Oysters. Front. Mar. Sci. 2018, 5, 345. [Google Scholar] [CrossRef]
- Sendra, M.; Saco, A.; Yeste, M.P.; Romero, A.; Novoa, B.; Figueras, A. Nanoplastics: From Tissue Accumulation to Cell Translocation into Mytilus galloprovincialis Hemocytes. Resilience of Immune Cells Exposed to Nanoplastics and Nanoplastics plus Vibrio splendidus Combination. J. Hazard. Mater. 2020, 388. [Google Scholar] [CrossRef]
- Noventa, S.; Hacker, C.; Correia, A.; Drago, C.; Galloway, T. Gold Nanoparticles Ingested by Oyster Larvae Are Internalized by Cells through an Alimentary Endocytic Pathway. Nanotoxicology 2018, 12, 901–913. [Google Scholar] [CrossRef] [Green Version]
- van der Zande, M.; Jemec Kokalj, A.; Spurgeon, D.J.; Loureiro, S.; Silva, P.v.; Khodaparast, Z.; Drobne, D.; Clark, N.J.; van den Brink, N.W.; Baccaro, M.; et al. The Gut Barrier and the Fate of Engineered Nanomaterials: A View from Comparative Physiology. Environ. Sci. Nano 2020, 7, 1874–1898. [Google Scholar] [CrossRef]
- Zhang, Y.; Pu, S.; Lv, X.; Gao, Y.; Ge, L. Global Trends and Prospects in Microplastics Research: A Bibliometric Analysis. J. Hazard. Mater. 2020, 400, 123110. [Google Scholar] [CrossRef]
- Fu, P.P.; Xia, Q.; Hwang, H.M.; Ray, P.C.; Yu, H. Mechanisms of Nanotoxicity: Generation of Reactive Oxygen Species. J. Food Drug Anal. 2014, 22, 64–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, T.; Araújo, O.; Pereira, R.; Almeida, A.C.; Cravo, A.; Bebianno, M.J. Genotoxicity of Copper Oxide and Silver Nanoparticles in the Mussel Mytilus galloprovincialis. Mar. Environ. Res. 2013, 84, 51–59. [Google Scholar] [CrossRef]
- Jovanović, B.; Palić, D.Š. Immunotoxicology of Non-Functionalized Engineered Nanoparticles in Aquatic Organisms with Special Emphasis on Fish-Review of Current Knowledge, Gap Identification, and Call for Further Research. Aquat. Toxicol. 2012, 118, 141–151. [Google Scholar] [CrossRef]
- Matranga, V.; Corsi, I. Toxic Effects of Engineered Nanoparticles in the Marine Environment: Model Organisms and Molecular Approaches. Mar. Environ. Res. 2012, 76, 32–40. [Google Scholar] [CrossRef]
- Canesi, L.; Ciacci, C.; Fabbri, R.; Marcomini, A.; Pojana, G.; Gallo, G. Bivalve Molluscs as a Unique Target Group for Nanoparticle Toxicity. Mar. Environ. Res. 2012, 76, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Matranga, V.; Pinsino, A.; Celi, M.; di Bella, G.; Natoli, A. Impacts of UV-B Radiation on Short-Term Cultures of Sea Urchin Coelomocytes. Mar. Biol. 2006, 149, 25–34. [Google Scholar] [CrossRef]
- Migliaccio, O.; Pinsino, A.; Maffioli, E.; Smith, A.M.; Agnisola, C.; Matranga, V.; Nonnis, S.; Tedeschi, G.; Byrne, M.; Gambi, M.C.; et al. Living in Future Ocean Acidification, Physiological Adaptive Responses of the Immune System of Sea Urchins Resident at a CO2 Vent System. Sci. Total Environ. 2019, 672, 938–950. [Google Scholar] [CrossRef]
- Murano, C.; Agnisola, C.; Caramiello, D.; Castellano, I.; Casotti, R.; Corsi, I.; Palumbo, A. How Sea Urchins Face Microplastics: Uptake, Tissue Distribution and Immune System Response. Environ. Pollut. 2020, 264, 114685. [Google Scholar] [CrossRef] [PubMed]
- Matranga, V.; Toia, G.; Bonaventura, R.; Müller, W.E.G. Cellular and Biochemical Responses to Environmental and Experimentally Induced Stress in Sea Urchin Coelomocytes. Cell Stress Chaperones 2000, 5, 113–120. [Google Scholar] [CrossRef]
- Canesi, L.; Ciacci, C.; Bergami, E.; Monopoli, M.P.; Dawson, K.A.; Papa, S.; Canonico, B.; Corsi, I. Evidence for Immunomodulation and Apoptotic Processes Induced by Cationic Polystyrene Nanoparticles in the Hemocytes of the Marine Bivalve Mytilus. Mar. Environ. Res. 2015, 111, 34–40. [Google Scholar] [CrossRef]
- Auguste, M.; Lasa, A.; Pallavicini, A.; Gualdi, S.; Vezzulli, L.; Canesi, L. Exposure to TiO2 Nanoparticles Induces Shifts in the Microbiota Composition of Mytilus galloprovincialis Hemolymph. Sci. Total Environ. 2019, 670, 129–137. [Google Scholar] [CrossRef]
- Auguste, M.; Balbi, T.; Ciacci, C.; Canonico, B.; Papa, S.; Borello, A.; Vezzulli, L.; Canesi, L. Shift in Immune Parameters After Repeated Exposure to Nanoplastics in the Marine Bivalve Mytilus. Front. Immunol. 2020, 11, 436. [Google Scholar] [CrossRef] [PubMed]
- Murano, C.; Bergami, E.; Liberatori, G.; Palumbo, A.; Corsi, I. Interplay Between Nanoplastics and the Immune System of the Mediterranean Sea Urchin Paracentrotus lividus. Front. Mar. Sci. 2021, 8, 647394. [Google Scholar] [CrossRef]
- Xia, B.; Zhu, L.; Han, Q.; Sun, X.; Chen, B.; Qu, K. Effects of TiO2 Nanoparticles at Predicted Environmental Relevant Concentration on the Marine Scallop Chlamys farreri: An Integrated Biomarker Approach. Environ. Toxicol. Pharmacol. 2017, 50, 128–135. [Google Scholar] [CrossRef]
- Guan, X.; Shi, W.; Zha, S.; Rong, J.; Su, W.; Liu, G. Neurotoxic Impact of Acute TiO2 Nanoparticle Exposure on a Benthic Marine Bivalve Mollusk, Tegillarca granosa. Aquat. Toxicol. 2018, 200, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Saidani, W.; Sellami, B.; Khazri, A.; Mezni, A.; Dellali, M.; Joubert, O.; Sheehan, D.; Beyrem, H. Metal Accumulation, Biochemical and Behavioral Responses on the Mediterranean Clams Ruditapes decussatus Exposed to Two Photocatalyst Nanocomposites (TiO2 NPs and AuTiO2 NPs). Aquat. Toxicol. 2019, 208, 71–79. [Google Scholar] [CrossRef]
- Drews, U. Cholinesterase in Embryonic Development. Prog. Histochem. Cytochem. 1975, 7, 5–7. [Google Scholar] [CrossRef]
- Gambardella, C.; Aluigi, M.G.; Ferrando, S.; Gallus, L.; Ramoino, P.; Gatti, A.M.; Rottigni, M.; Falugi, C. Developmental Abnormalities and Changes in Cholinesterase Activity in Sea Urchin Embryos and Larvae from Sperm Exposed to Engineered Nanoparticles. Aquat. Toxicol. 2013, 130, 77–85. [Google Scholar] [CrossRef]
- Jeong, C.B.; Kang, H.M.; Lee, M.C.; Kim, D.H.; Han, J.; Hwang, D.S.; Souissi, S.; Lee, S.J.; Shin, K.H.; Park, H.G.; et al. Adverse Effects of Microplastics and Oxidative Stress-Induced MAPK/Nrf2 Pathway-Mediated Defense Mechanisms in the Marine Copepod Paracyclopina nana. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, C.B.; Won, E.J.; Kang, H.M.; Lee, M.C.; Hwang, D.S.; Hwang, U.K.; Zhou, B.; Souissi, S.; Lee, S.J.; Lee, J.S. Microplastic Size-Dependent Toxicity, Oxidative Stress Induction, and p-JNK and p-P38 Activation in the Monogonont Rotifer (Brachionus koreanus). Environ. Sci. Technol. 2016, 50, 8849–8857. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.; Figueira, E.; Libralato, G.; Soares, A.M.V.M.; Guida, M.; Freitas, R. Comparative Sensitivity of Crassostrea angulata and Crassostrea gigas Embryo-Larval Development to As under Varying Salinity and Temperature. Mar. Environ. Res. 2018, 140, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Balbi, T.; Camisassi, G.; Montagna, M.; Fabbri, R.; Franzellitti, S.; Carbone, C.; Dawson, K.; Canesi, L. Impact of Cationic Polystyrene Nanoparticles (PS-NH2) on Early Embryo Development of Mytilus galloprovincialis: Effects on Shell Formation. Chemosphere 2017, 186, 1–9. [Google Scholar] [CrossRef]
- Auguste, M.; Ciacci, C.; Balbi, T.; Brunelli, A.; Caratto, V.; Marcomini, A.; Cuppini, R.; Canesi, L. Effects of Nanosilver on Mytilus galloprovincialis Hemocytes and Early Embryo Development. Aquat. Toxicol. 2018, 203, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Manzo, S.; Miglietta, M.L.; Rametta, G.; Buono, S.; di Francia, G. Embryotoxicity and Spermiotoxicity of Nanosized ZnO for Mediterranean Sea Urchin Paracentrotus lividus. J. Hazard. Mater. 2013, 254, 1–9. [Google Scholar] [CrossRef]
- Maisano, M.; Cappello, T.; Catanese, E.; Vitale, V.; Natalotto, A.; Giannetto, A.; Barreca, D.; Brunelli, E.; Mauceri, A.; Fasulo, S. Developmental Abnormalities and Neurotoxicological Effects of CuO NPs on the Black Sea Urchin Arbacia lixula by Embryotoxicity Assay. Mar. Environ. Res. 2015, 111, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Torres-Duarte, C.; Adeleye, A.S.; Pokhrel, S.; Mädler, L.; Keller, A.A.; Cherr, G.N. Developmental Effects of Two Different Copper Oxide Nanomaterials in Sea Urchin (Lytechinus pictus) Embryos. Nanotoxicology 2016, 10, 671–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siller, L.; Lemloh, M.L.; Piticharoenphun, S.; Mendis, B.G.; Horrocks, B.R.; Brümmer, F.; Medaković, D. Silver Nanoparticle Toxicity in Sea Urchin Paracentrotus lividus. Environ. Pollut. (Barking Essex 1987) 2013, 178, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Pinsino, A.; Bergami, E.; Della Torre, C.; Vannuccini, M.L.; Addis, P.; Secci, M.; Dawson, K.A.; Matranga, V.; Corsi, I. Amino-Modified Polystyrene Nanoparticles Affect Signalling Pathways of the Sea Urchin (Paracentrotus lividus) Embryos. Nanotoxicology 2017, 11, 201–209. [Google Scholar] [CrossRef]
- Long, M.; Moriceau, B.; Gallinari, M.; Lambert, C.; Huvet, A.; Raffray, J.; Soudant, P. Interactions between Microplastics and Phytoplankton Aggregates: Impact on Their Respective Fates. Mar. Chem. 2015, 175, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Bellingeri, A.; Scattoni, M.; Venditti, I.; Battocchio, C.; Faleri, C.; Protano, G.; Corsi, I. Innocent Until Proven Guilty: Chronic Toxicity of AgNPcitLcys for Water Remediation Reveals the Need to Reconsider Ecosafety Testing. Environ. Sci. Nano 2021. submitted. [Google Scholar]
- Cervantes-Avilés, P.; Huang, Y.; Keller, A.A. Multi-Technique Approach to Study the Stability of Silver Nanoparticles at Predicted Environmental Concentrations in Wastewater. Water Res. 2019, 166, 115072. [Google Scholar] [CrossRef]
- Nabi, M.M.; Wang, J.; Meyer, M.; Croteau, M.N.; Ismail, N.; Baalousha, M. Concentrations and Size Distribution of TiO2 and Ag Engineered Particles in Five Wastewater Treatment Plants in the United States. Sci. Total Environ. 2021, 753, 142017. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Stoiber, M.; Wimmer, A.; Xu, Z.; Lindenblatt, C.; Helmreich, B.; Schuster, M. To What Extent Can Full-Scale Wastewater Treatment Plant Effluent Influence the Occurrence of Silver-Based Nanoparticles in Surface Waters? Environ. Sci. Technol. 2016, 50, 6327–6333. [Google Scholar] [CrossRef]
- Lankoff, A.; Sandberg, W.J.; Wegierek-Ciuk, A.; Lisowska, H.; Refsnes, M.; Sartowska, B.; Schwarze, P.E.; Meczynska-Wielgosz, S.; Wojewodzka, M.; Kruszewski, M. The Effect of Agglomeration State of Silver and Titanium Dioxide Nanoparticles on Cellular Response of HepG2, A549 and THP-1 Cells. Toxicol. Lett. 2012, 208, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Tsiola, A.; Pitta, P.; Callol, A.J.; Kagiorgi, M.; Kalantzi, I.; Mylona, K.; Santi, I.; Toncelli, C.; Pergantis, S.; Tsapakis, M. The Impact of Silver Nanoparticles on Marine Plankton Dynamics: Dependence on Coating, Size and Concentration. Sci. Total Environ. 2017, 601, 1838–1848. [Google Scholar] [CrossRef]
- Labille, J.; Slomberg, D.; Catalano, R.; Robert, S.; Apers-Tremelo, M.L.; Boudenne, J.L.; Manasfi, T.; Radakovitch, O. Assessing UV Filter Inputs into Beach Waters during Recreational Activity: A Field Study of Three French Mediterranean Beaches from Consumer Survey to Water Analysis. Sci. Total Environ. 2020, 706, 136010. [Google Scholar] [CrossRef] [PubMed]
- Lazareva, A.; Keller, A.A. Estimating Potential Life Cycle Releases of Engineered Nanomaterials from Wastewater Treatment Plants. ACS Sustain. Chem. Eng. 2014, 2, 1656–1665. [Google Scholar] [CrossRef]
- Maurer-Jones, M.A.; Gunsolus, I.L.; Murphy, C.J.; Haynes, C.L. Toxicity of Engineered Nanoparticles in the Environment. Anal. Chem. 2013, 85, 3036–3049. [Google Scholar] [CrossRef] [Green Version]
- Dedman, C.J.; Newson, G.C.; Davies, G.L.; Christie-Oleza, J.A. Mechanisms of Silver Nanoparticle Toxicity on the Marine Cyanobacterium Prochlorococcus under Environmentally-Relevant Conditions. Sci. Total Environ. 2020, 747, 141229. [Google Scholar] [CrossRef]
- Angel, B.M.; Batley, G.E.; Jarolimek, C.v.; Rogers, N.J. The Impact of Size on the Fate and Toxicity of Nanoparticulate Silver in Aquatic Systems. Chemosphere 2013, 93, 359–365. [Google Scholar] [CrossRef]
- Kos, M.; Kahru, A.; Drobne, D.; Singh, S.; Kalčíková, G.; Kühnel, D.; Rohit, R.; Gotvajn, A.Ž.; Jemec, A. A Case Study to Optimise and Validate the Brine Shrimp Artemia franciscana Immobilisation Assay with Silver Nanoparticles: The Role of Harmonisation. Environ. Pollut. 2016, 213, 173–183. [Google Scholar] [CrossRef] [PubMed]
- An, H.J.; Sarkheil, M.; Park, H.S.; Yu, I.J.; Johari, S.A. Comparative Toxicity of Silver Nanoparticles (AgNPs) and Silver Nanowires (AgNWs) on Saltwater Microcrustacean, Artemia salina. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 218, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Sendra, M.; Yeste, M.P.; Gatica, J.M.; Moreno-Garrido, I.; Blasco, J. Direct and Indirect Effects of Silver Nanoparticles on Freshwater and Marine Microalgae (Chlamydomonas reinhardtii and Phaeodactylum tricornutum). Chemosphere 2017, 179, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Becaro, A.A.; Jonsson, C.M.; Puti, F.C.; Siqueira, M.C.; Mattoso, L.H.C.; Correa, D.S.; Ferreira, M.D. Toxicity of PVA-Stabilized Silver Nanoparticles to Algae and Microcrustaceans. Environ. Nanotechnol. Monit. Manag. 2015, 3, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Hamouda, T.; Ibrahim, H.M.; Kafafy, H.H.; Mashaly, H.M.; Mohamed, N.H.; Aly, N.M. Preparation of Cellulose-Based Wipes Treated with Antimicrobial and Antiviral Silver Nanoparticles as Novel Effective High-Performance Coronavirus Fighter. Int. J. Biol. Macromol. 2021, 181, 990–1002. [Google Scholar] [CrossRef]
- Valdez-Salas, B.; Beltran-Partida, E.; Cheng, N.; Salvador-Carlos, J.; Valdez-Salas, E.A.; Curiel-Alvarez, M.; Ibarra-Wiley, R. Promotion of Surgical Masks Antimicrobial Activity by Disinfection and Impregnation with Disinfectant Silver Nanoparticles. Int. J. Nanomed. 2021, 16, 2689–2702. [Google Scholar] [CrossRef] [PubMed]
- Syafiuddin, A.; Salmiati; Salim, M.; Kueh, A.; Hadibarata, T.; Nur, H. A Review of Silver Nanoparticles: Research Trends, Global Consumption, Synthesis, Properties, and Future Challenges. J. Chin. Chem. Soc. 2017, 64, 732–756. [Google Scholar] [CrossRef]
- Dalzon, B.; Aude-Garcia, C.; Diemer, H.; Bons, J.; Marie-Desvergne, C.; Pérard, J.; Dubosson, M.; Collin-Faure, V.; Carapito, C.; Cianférani, S.; et al. The Longer the Worse: A Combined Proteomic and Targeted Study of the Long-Term: Versus Short-Term Effects of Silver Nanoparticles on Macrophages. Environ. Sci. Nano 2020, 7, 2032–2046. [Google Scholar] [CrossRef]
- Lead, J.R.; Batley, G.E.; Alvarez, P.J.J.; Croteau, M.N.; Handy, R.D.; McLaughlin, M.J.; Judy, J.D.; Schirmer, K. Nanomaterials in the Environment: Behavior, Fate, Bioavailability, and Effects—An Updated Review. Environ. Toxicol. Chem. 2018, 37, 2029–2063. [Google Scholar] [CrossRef] [PubMed]
- Chae, Y.; An, Y.J. Effects of Micro- and Nanoplastics on Aquatic Ecosystems: Current Research Trends and Perspectives. Mar. Pollut. Bull. 2017, 124, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Venâncio, C.; Ferreira, I.; Martins, M.A.; Soares, A.M.V.M.; Lopes, I.; Oliveira, M. The Effects of Nanoplastics on Marine Plankton: A Case Study with Polymethylmethacrylate. Ecotoxicol. Environ. Saf. 2019, 184, 109632. [Google Scholar] [CrossRef]
- Lenz, R.; Enders, K.; Nielsen, T.G. Microplastic Exposure Studies Should Be Environmentally Realistic. Proc. Natl. Acad. Sci. USA 2016, 113, E4121–E41222. [Google Scholar] [CrossRef] [Green Version]
- Llorca, M.; Álvarez-Muñoz, D.; Ábalos, M.; Rodríguez-Mozaz, S.; Santos, L.H.M.L.M.; León, V.M.; Campillo, J.A.; Martínez-Gómez, C.; Abad, E.; Farré, M. Microplastics in Mediterranean Coastal Area: Toxicity and Impact for the Environment and Human Health. Trends Environ. Anal. Chem. 2020, 27, e00090. [Google Scholar] [CrossRef]
- Pourzahedi, L.; Vance, M.; Eckelman, M.J. Life Cycle Assessment and Release Studies for 15 Nanosilver-Enabled Consumer Products: Investigating Hotspots and Patterns of Contribution. Environ. Sci. Technol. 2017, 51, 7148–7158. [Google Scholar] [CrossRef]
- Salieri, B.; Turner, D.A.; Nowack, B.; Hischier, R. Life Cycle Assessment of Manufactured Nanomaterials: Where Are We? NanoImpact 2018, 10, 108–120. [Google Scholar] [CrossRef]
- Moeta, P.J.; Wesley-Smith, J.; Maity, A.; Thwala, M. Nano-Enabled Products in South Africa and the Assessment of Environmental Exposure Potential for Engineered Nanomaterials. SN Appl. Sci. 2019, 1, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Nowack, B.; Mitrano, D.M. Procedures for the Production and Use of Synthetically Aged and Product Released Nanomaterials for Further Environmental and Ecotoxicity Testing. NanoImpact 2018, 10, 70–80. [Google Scholar] [CrossRef]
- Vílchez, A.; Fernández-Rosas, E.; González-Gálvez, D.; Vázquez-Campos, S. Nanomaterials Release from Nano-Enabled Products. In Handbook of Environmental Chemistry; Springer: Berlin/Heidelberg, Germany, 2016; Volume 48, pp. 127–158. [Google Scholar] [CrossRef]
- Rai, M.; Birla, S.; Ingle, A.P.; Gupta, I.; Gade, A.; Abd-Elsalam, K.; Marcato, P.D.; Duran, N. Nanosilver: An Inorganic Nanoparticle with Myriad Potential Applications. Nanotechnol. Rev. 2014, 3, 281–309. [Google Scholar] [CrossRef]
- Abu-Qdais, H.A.; Abu-Dalo, M.A.; Hajeer, Y.Y. Impacts of Nanosilver-Based Textile Products Using a Life Cycle Assessment. Sustainability 2021, 13, 3436. [Google Scholar] [CrossRef]
- Lehutso, R.F.; Tancu, Y.; Maity, A.; Thwala, M. Aquatic Toxicity of Transformed and Product-Released Engineered Nanomaterials: An Overview of the Current State of Knowledge. Process. Saf. Environ. Prot. 2020, 138, 39–56. [Google Scholar] [CrossRef]
- Ale, A.; Liberatori, G.; Vannuccini, M.L.; Bergami, E.; Ancora, S.; Mariotti, G.; Bianchi, N.; Galdopórpora, J.M.; Desimone, M.F.; Cazenave, J.; et al. Exposure to a Nanosilver-Enabled Consumer Product Results in Similar Accumulation and Toxicity of Silver Nanoparticles in the Marine Mussel Mytilus galloprovincialis. Aquat. Toxicol. 2019, 211, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Gomes, T.; Pereira, C.G.; Cardoso, C.; Bebianno, M.J. Differential Protein Expression in Mussels Mytilus galloprovincialis Exposed to Nano and Ionic Ag. Aquat. Toxicol. 2013, 136, 79–90. [Google Scholar] [CrossRef]
- Gomes, T.; Pereira, C.G.; Cardoso, Ć.; Sousa, V.S.; Teixeira, M.R.; Pinheiro, J.P.; Bebianno, M.J. Effects of Silver Nanoparticles Exposure in the Mussel Mytilus galloprovincialis. Mar. Environ. Res. 2014, 101, 208–214. [Google Scholar] [CrossRef]
- Katsumiti, A.; Gilliland, D.; Arostegui, I.; Cajaraville, M.P. Mechanisms of Toxicity of Ag Nanoparticles in Comparison to Bulk and Ionic Ag on Mussel Hemocytes and Gill Cells. PLoS ONE 2015, 10, e0129039. [Google Scholar] [CrossRef]
- Duroudier, N.; Katsumiti, A.; Mikolaczyk, M.; Schäfer, J.; Bilbao, E.; Cajaraville, M.P. Dietary Exposure of Mussels to PVP/PEI Coated Ag Nanoparticles Causes Ag Accumulation in Adults and Abnormal Embryo Development in Their Offspring. Sci. Total Environ. 2019, 655, 48–60. [Google Scholar] [CrossRef]
- Reynolds, C.S. Benthic Ecology. Encycl. Environ. 2006. [Google Scholar] [CrossRef]
- Simpson, S.L.; Batley, G.E. Predicting Metal Toxicity in Sediments: A Critique of Current Approaches. Integr. Environ. Assess. Manag. 2007, 3, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Antizar-Ladislao, B.; Bhattacharya, B.D.; Ray Chaudhuri, S.; Sarkar, S.K. Impact of Silver Nanoparticles on Benthic Prokaryotes in Heavy Metal-Contaminated Estuarine Sediments in a Tropical Environment. Mar. Pollut. Bull. 2015, 99, 104–111. [Google Scholar] [CrossRef]
- Chan, C.Y.S.; Chiu, J.M.Y. Chronic Effects of Coated Silver Nanoparticles on Marine Invertebrate Larvae: A Proof of Concept Study. PLoS ONE 2015, 10, e0132457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Ho, K.T.; Scheckel, K.G.; Wu, F.; Cantwell, M.G.; Katz, D.R.; Horowitz, D.B.; Boothman, W.S.; Burgess, R.M. Toxicity, Bioaccumulation, and Biotransformation of Silver Nanoparticles in Marine Organisms. Environ. Sci. Technol. 2014, 48, 13711–13717. [Google Scholar] [CrossRef] [PubMed]
- Buffet, P.E.; Zalouk-Vergnoux, A.; Châtel, A.; Berthet, B.; Métais, I.; Perrein-Ettajani, H.; Poirier, L.; Luna-Acosta, A.; Thomas-Guyon, H.; Risso-de Faverney, C.; et al. A Marine Mesocosm Study on the Environmental Fate of Silver Nanoparticles and Toxicity Effects on Two Endobenthic Species: The Ragworm Hediste diversicolor and the Bivalve Mollusc Scrobicularia plana. Sci. Total Environ. 2014, 470–471, 1151–1159. [Google Scholar] [CrossRef]
- Cong, Y.; Banta, G.T.; Selck, H.; Berhanu, D.; Valsami-Jones, E.; Forbes, V.E. Toxicity and Bioaccumulation of Sediment-Associated Silver Nanoparticles in the Estuarine Polychaete, Nereis (Hediste) diversicolor. Aquat. Toxicol. 2014, 156, 106–115. [Google Scholar] [CrossRef]
- García-Alonso, J.; Khan, F.R.; Misra, S.K.; Turmaine, M.; Smith, B.D.; Rainbow, P.S.; Luoma, S.N.; Valsami-Jones, E. Cellular Internalization of Silver Nanoparticles in Gut Epithelia of the Estuarine Polychaete Nereis diversicolor. Environ. Sci. Technol. 2011, 45, 4630–4636. [Google Scholar] [CrossRef]
- Cozzari, M.; Elia, A.C.; Pacini, N.; Smith, B.D.; Boyle, D.; Rainbow, P.S.; Khan, F.R. Bioaccumulation and Oxidative Stress Responses Measured in the Estuarine Ragworm (Nereis diversicolor) Exposed to Dissolved, Nano- and Bulk-Sized Silver. Environ. Pollut. 2015, 198, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Ciacci, C.; Grimmelpont, M.V.; Corsi, I.; Bergami, E.; Curzi, D.; Burini, D.; Bouchet, V.M.P.; Ambrogini, P.; Gobbi, P.; Ujiié, Y.; et al. Nanoparticle-Biological Interactions in a Marine Benthic Foraminifer. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Wang, Z.; Yin, L.; Zhao, J.; Xing, B. Trophic Transfer and Accumulation of TiO2 Nanoparticles from Clamworm (Perinereis aibuhitensis) to Juvenile Turbot (Scophthalmus maximus) along a Marine Benthic Food Chain. Water Res. 2016, 95, 250–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.S.S.; Oliveira, M.; Valente, P.; Figueira, E.; Martins, M.; Pires, A. Behavior and Biochemical Responses of the Polychaeta Hediste diversicolor to Polystyrene Nanoplastics. Sci. Total Environ. 2020, 707, 134434. [Google Scholar] [CrossRef] [PubMed]
- Banta, G.T.; Andersen, O. Bioturbation and the Fate of Sediment Pollutants-Experimental Case Studies of Selected Infauna Species. Vie Et Milieu/Life Environ. Obs. Océanologique Lab. Arago 2003, 53, 233–248. [Google Scholar]
- Chariton, A.A.; Sun, M.; Gibson, J.; Webb, J.A.; Leung, K.M.Y.; Hickey, C.W.; Hose, G.C. Emergent Technologies and Analytical Approaches for Understanding the Effects of Multiple Stressors in Aquatic Environments. Mar. Freshw. Res. 2016, 67, 414–428. [Google Scholar] [CrossRef]
- Hjorth, R.; van Hove, L.; Wickson, F. What Can Nanosafety Learn from Drug Development? The Feasibility of “Safety by Design”. Nanotoxicology 2017, 11, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Bartolozzi, I.; Daddi, T.; Punta, C.; Fiorati, A.; Iraldo, F. Life Cycle Assessment of Emerging Environmental Technologies in the Early Stage of Development: A Case Study on Nanostructured Materials. J. Ind. Ecol. 2020, 24, 101–115. [Google Scholar] [CrossRef]
- Fiorati, A.; Grassi, G.; Graziano, A.; Liberatori, G.; Pastori, N.; Melone, L.; Bonciani, L.; Pontorno, L.; Punta, C.; Corsi, I. Eco-Design of Nanostructured Cellulose Sponges for Sea-Water Decontamination from Heavy Metal Ions. J. Clean. Prod. 2020, 246, 119009. [Google Scholar] [CrossRef]
- Esposito, M.C.; Corsi, I.; Russo, G.L.; Punta, C.; Tosti, E.; Gallo, A. The Era of Nanomaterials: A Safe Solution or a Risk for Marine Environmental Pollution? Biomolecules 2021, 11, 441. [Google Scholar] [CrossRef]
- Corsi, I.; Fiorati, A.; Grassi, G.; Bartolozzi, I.; Daddi, T.; Melone, L.; Punta, C. Environmentally Sustainable and Ecosafe Polysaccharide-Based Materials for Water Nano-Treatment: An Eco-Design Study. Materials 2018, 11, 1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melone, L.; Rossi, B.; Pastori, N.; Panzeri, W.; Mele, A.; Punta, C. TEMPO-Oxidized Cellulose Cross-Linked with Branched Polyethyleneimine: Nanostructured Adsorbent Sponges for Water Remediation. ChemPlusChem 2015, 80, 1408–1415. [Google Scholar] [CrossRef] [PubMed]
- Riva, L.; Pastori, N.; Panozzo, A.; Antonelli, M.; Punta, C. Nanostructured Cellulose-Based Sorbent Materials for Water Decontamination from Organic Dyes. Nanomaterials 2020, 10, 1570. [Google Scholar] [CrossRef]
- Omidi, Y.; Kafil, V. Cytotoxic Impacts of Linear and Branched Polyethylenimine Nanostructures in A431 Cells. Bioimpact 2011, 1, 23–30. [Google Scholar] [CrossRef]
- Fischer, D.; Li, Y.; Ahlemeyer, B.; Krieglstein, J.; Kissel, T. In Vitro Cytotoxicity Testing of Polycations: Influence of Polymer Structure on Cell Viability and Hemolysis. Biomaterials 2003, 24, 1121–1131. [Google Scholar] [CrossRef]
- Liberatori, G.; Grassi, G.; Guidi, P.; Bernardeschi, M.; Fiorati, A.; Scarcelli, V.; Genovese, M.; Faleri, C.; Protano, G.; Frenzilli, G.; et al. Effect-Based Approach to Assess Nanostructured Cellulose Sponge Removal Efficacy of Zinc Ions from Seawater to Prevent Ecological Risks. Nanomaterials 2020, 10, 1283. [Google Scholar] [CrossRef]
- Woodrow, O.; Plaza, W.; Luoma, S.N.; Hamilton, L.H.; Cook, R.B.; Garcia, D.E.; Gelb, B.S.; Gerber, S.R.; Glazer, C.L.; Hutchison, S.; et al. Project on Emerging Nanotechnologies. Project on Emerging Nanotechnologies is Supported by the PEW Charitable Trusts. Emerging Nanotechnologies Old Problems or New Challenges? 2008. Available online: https://www.pewtrusts.org/en/projects/archived-projects/project-on-emerging-nanotechnologies (accessed on 17 June 2021).
- Levard, C.; Reinsch, B.C.; Michel, F.M.; Oumahi, C.; Lowry, G.V.; Brown, G.E. Sulfidation Processes of PVP-Coated Silver Nanoparticles in Aqueous Solution: Impact on Dissolution Rate. Environ. Sci. Technol. 2011, 45, 5260–5266. [Google Scholar] [CrossRef]
- Hotze, E.M.; Phenrat, T.; Lowry, G.V. Nanoparticle Aggregation: Challenges to Understanding Transport and Reactivity in the Environment. J. Environ. Qual. 2010, 39, 1909–1924. [Google Scholar] [CrossRef] [Green Version]
- Pem, B.; Pongrac, I.M.; Ulm, L.; Pavičić, I.; Vrček, V.; Jurašin, D.D.; Ljubojević, M.; Krivohlavek, A.; Vrček, I.V. Toxicity and Safety Study of Silver and Gold Nanoparticles Functionalized with Cysteine and Glutathione. Beilstein J. Nanotechnol. 2019, 10, 1802–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corsi, I.; Bellingeri, A.; Eliso, M.C.; Grassi, G.; Liberatori, G.; Murano, C.; Sturba, L.; Vannuccini, M.L.; Bergami, E. Eco-Interactions of Engineered Nanomaterials in the Marine Environment: Towards an Eco-Design Framework. Nanomaterials 2021, 11, 1903. https://doi.org/10.3390/nano11081903
Corsi I, Bellingeri A, Eliso MC, Grassi G, Liberatori G, Murano C, Sturba L, Vannuccini ML, Bergami E. Eco-Interactions of Engineered Nanomaterials in the Marine Environment: Towards an Eco-Design Framework. Nanomaterials. 2021; 11(8):1903. https://doi.org/10.3390/nano11081903
Chicago/Turabian StyleCorsi, Ilaria, Arianna Bellingeri, Maria Concetta Eliso, Giacomo Grassi, Giulia Liberatori, Carola Murano, Lucrezia Sturba, Maria Luisa Vannuccini, and Elisa Bergami. 2021. "Eco-Interactions of Engineered Nanomaterials in the Marine Environment: Towards an Eco-Design Framework" Nanomaterials 11, no. 8: 1903. https://doi.org/10.3390/nano11081903
APA StyleCorsi, I., Bellingeri, A., Eliso, M. C., Grassi, G., Liberatori, G., Murano, C., Sturba, L., Vannuccini, M. L., & Bergami, E. (2021). Eco-Interactions of Engineered Nanomaterials in the Marine Environment: Towards an Eco-Design Framework. Nanomaterials, 11(8), 1903. https://doi.org/10.3390/nano11081903