
Ecotoxicity of Heteroaggregates of Polystyrene Nanospheres in Chironomidae and Amphibian

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plastic Materials and Physical-Chemical Analyze
2.1.1. Raw Materials, Preparation of Heteroaggregates, and Physical Characterizations in Water
2.1.2. Physical Characterization in Exposure Media
2.2. Organisms, Breeding, and Toxicity Assessment
2.2.1. Chironomids
2.2.2. Xenopus
3. Results
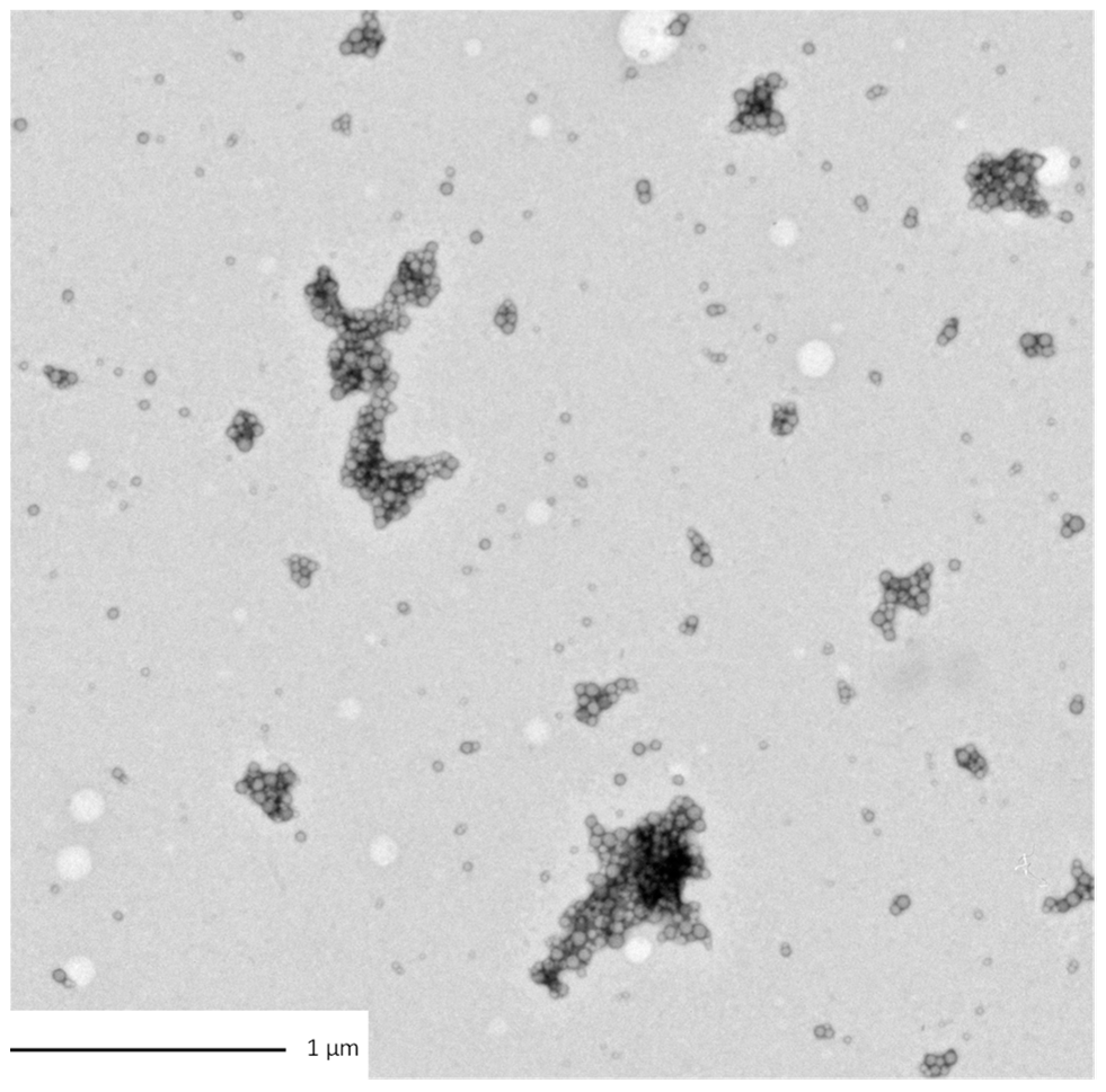
3.1. Morphological Characterization of the Particles and Aggregates
3.2. Toxicity Assessment on Biological Models
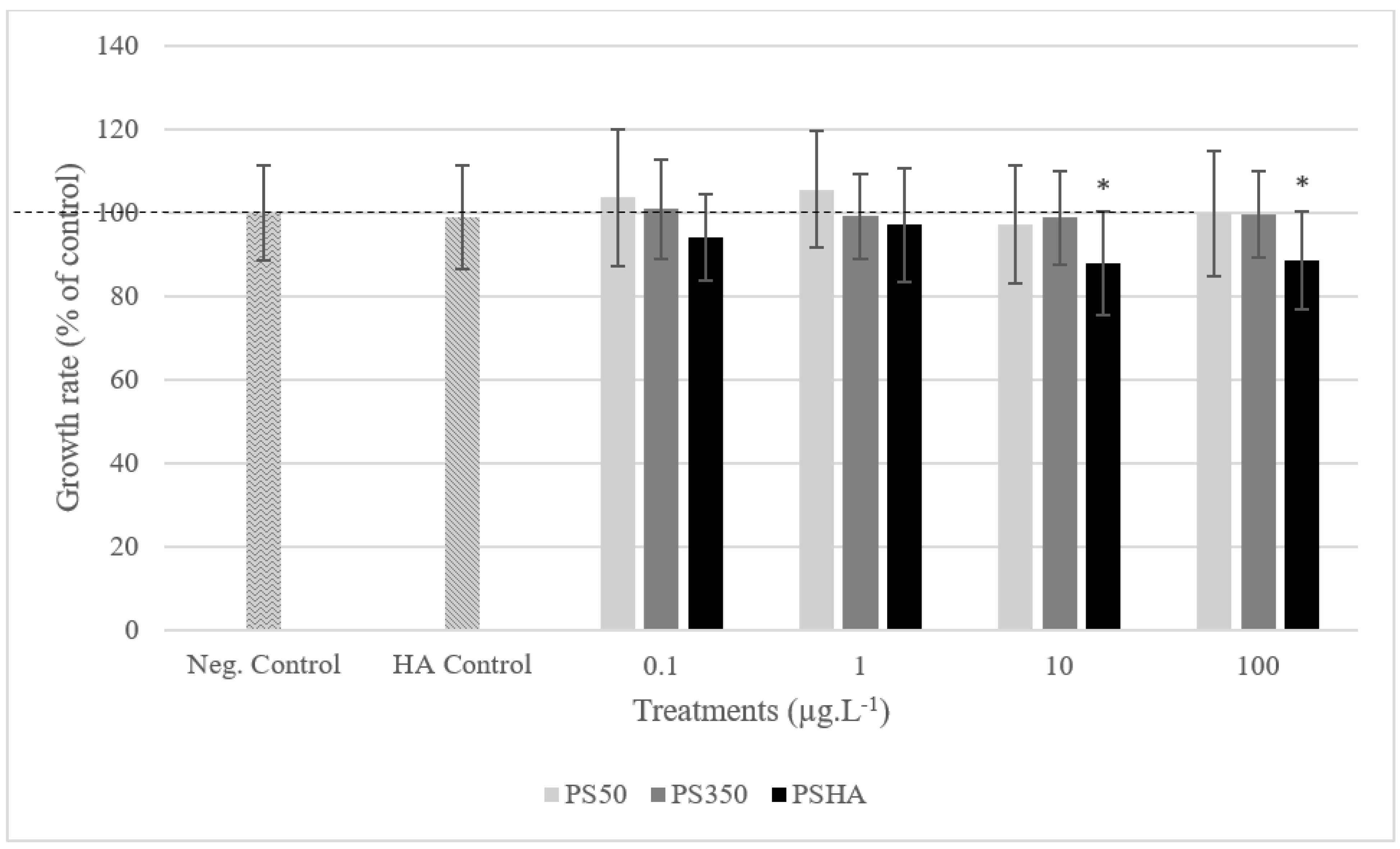
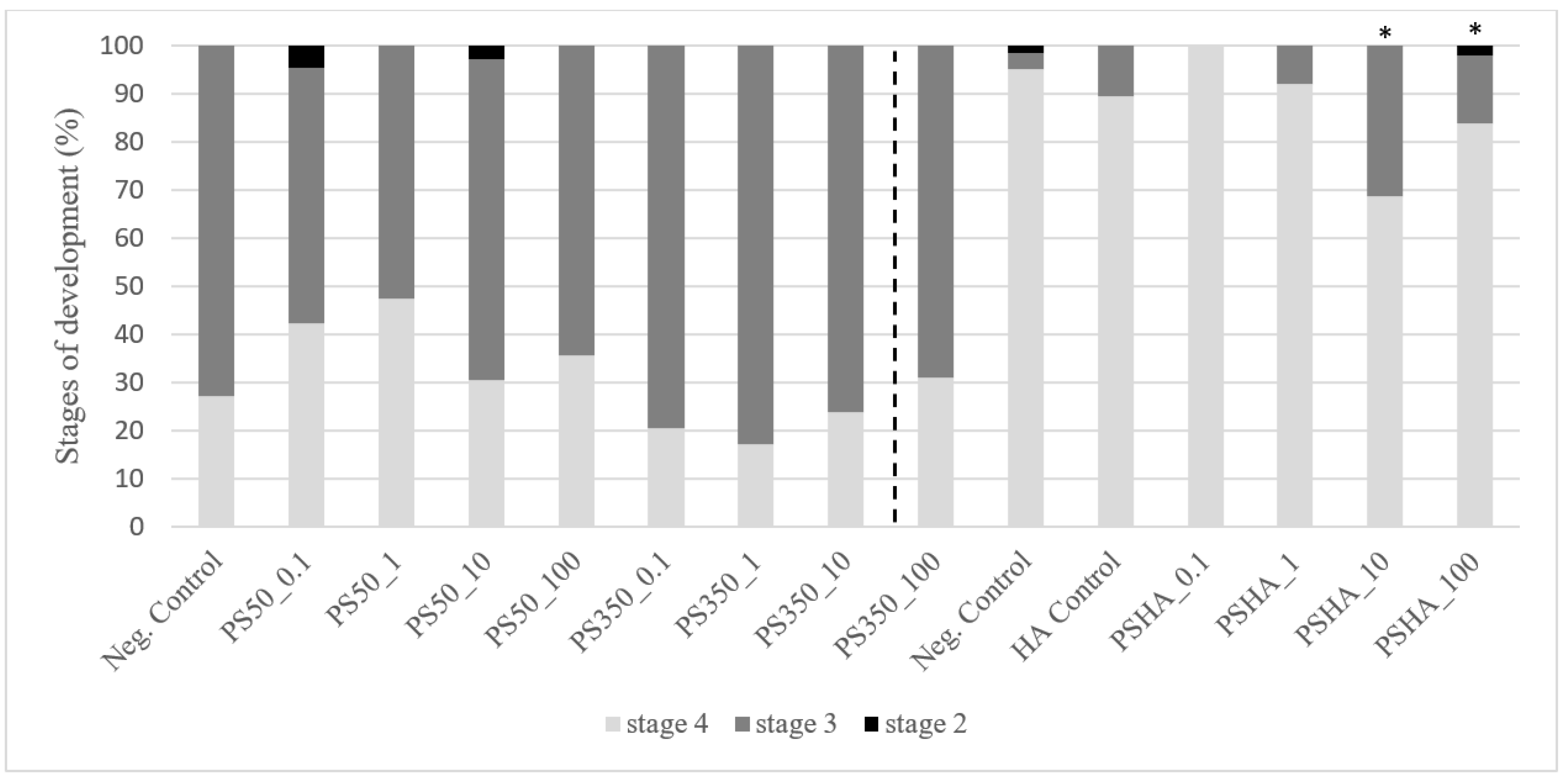
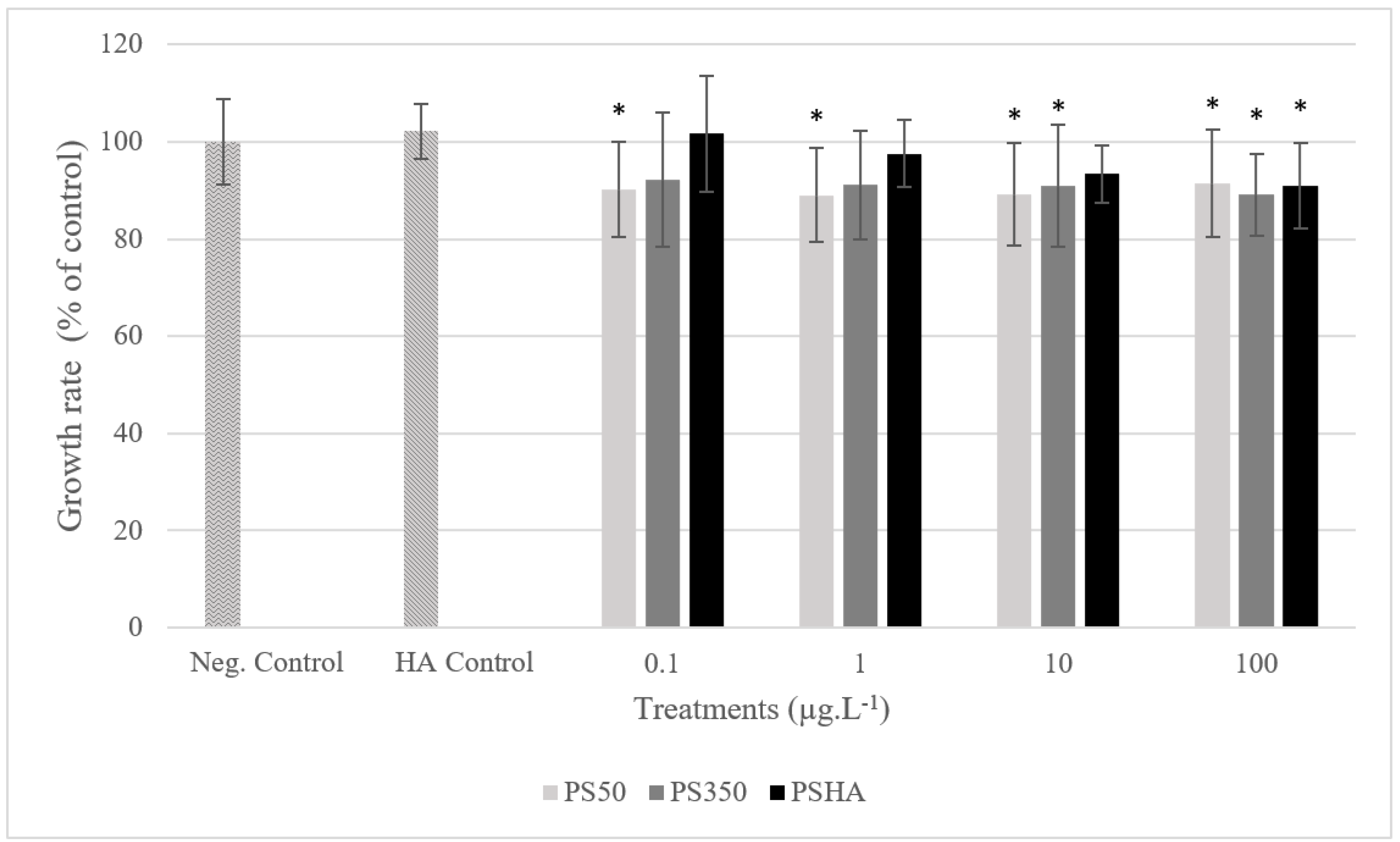
3.2.1. Toxicity on Chironomids
3.2.2. Toxicity on Amphibians
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Plastics Europe. Plastics—The Facts 2018. An Analysis of European Plastics Production, Demand and Waste Data. 2018. Available online: https://plasticseurope.org/wp-content/uploads/2021/10/2018-Plastics-the-facts.pdf (accessed on 1 July 2022).
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, Use, and Fate of All Plastics Ever Made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, T.; Chen, W.Q.; Zhu, B.; Qu, S.; Xu, M. Critical review of global plastics stock and flow data. J. Ind. Ecol. 2021, 25, 1300–1317. [Google Scholar] [CrossRef]
- Plastics Europe. Plastics—The Facts 2016. An Analysis of European Plastics Production, Demand and Waste Data. 2016. Available online: https://plasticseurope.org/wp-content/uploads/2021/10/2016-Plastic-the-facts.pdf (accessed on 1 July 2022).
- Plastics Europe. Plastics—The Facts 2017. An Analysis of European Plastics Production, Demand and Waste Data. 2017, p. 44. Available online: https://plasticseurope.org/wp-content/uploads/2021/10/2017-Plastics-the-facts.pdf (accessed on 1 July 2022).
- Plastics Europe. Plastics—The Facts 2019. An Analysis of European Plastics Production, Demand and Waste Data. 2019. Available online: https://plasticseurope.org/wp-content/uploads/2021/10/2019-Plastics-the-facts.pdf (accessed on 1 July 2022).
- Adyel, T.M. Accumulation of plastic waste during COVID-19. Science 2020, 369, 1314–1315. [Google Scholar] [PubMed]
- Silva, A.L.P.; Prata, J.C.; Walker, T.R.; Duarte, A.C.; Ouyang, W.; Barcelo, D.; Rocha-Santos, T. Increased plastic pollution due to COVID-19 pandemic: Challenges and recommendations. Chem. Eng. J. 2021, 405, 126683. [Google Scholar] [CrossRef]
- Sharma, S.; Chatterjee, S. Microplastic pollution, a threat to marine ecosystem and human health: A short review. Environ. Sci. Pollut. Res. Int. 2017, 24, 21530–21547. [Google Scholar] [CrossRef] [PubMed]
- Guzzetti, E.; Sureda, A.; Tejeda, S.; Faggio, C. Microplastic in marine organism: Environmental and toxicological effects. Environ. Toxicol. Pharmacol. 2018, 64, 164–171. [Google Scholar] [CrossRef]
- Bai, Z.; Wang, N.; Wang, M. Effects of microplastics on marine copepods. Ecotoxicol. Environ. Saf. 2021, 217, 112243. [Google Scholar] [CrossRef]
- Palmer, J.; Herat, S. Ecotoxicity of Microplastic Pollutants to Marine Organisms: A Systematic Review. Water Air Soil Pollut. 2021, 2, 195. [Google Scholar] [CrossRef]
- van Emmerik, T.; Schwarz, A. Plastic debris in rivers. Wiley Interdiscip. Rev. Water 2019, 7, e1398. [Google Scholar] [CrossRef] [Green Version]
- Lebreton, L.C.M.; Van der Zwet, J.; Damsteeg, J.W.; Slat, B.; Andrady, A.; Reisser, J. River plastic emissions to the world’s oceans. Nat. Commun. 2017, 8, 15611. [Google Scholar] [CrossRef]
- Weiss, L.; Ludwig, W.; Heussner, S.; Canals, M.; Ghiglione, J.F.; Estournel, C.; Constant, M.; Kerherve, P. The missing ocean plastic sink: Gone with the rivers. Science 2021, 373, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, G.L.; Gallardo, J.D.; Jones, E.W.; Hollliman, P.J.; Watson, T.M.; Sarp, S. Detection of trace sub-micron (nano) plastics in water samples using pyrolysis-gas chromatography time of flight mass spectrometry (PY-GCToF). Chemosphere 2020, 249, 126179. [Google Scholar] [CrossRef] [PubMed]
- Rowenczyk, L.; Cai, H.; Nguyen, B.; Sirois, M.; Côté-Laurin, M.-C.; Toupoint, N.; Ismail, A.; Tufenkji, N. From freshwaters to bivalves: Microplastic distribution along the Saint-Lawrence river-to-sea continuum. J. Hazard. Mater. 2022, 435, 128977. [Google Scholar] [CrossRef]
- ter Halle, A.; Ladirat, L.; Gendre, X.; Goudouneche, D.; Pusineri, C.; Routaboul, C.; Tenailleau, C.; Duployer, B.; Perez, E. Understanding the Fragmentation Pattern of Marine Plastic Debris. Environ. Sci. Technol. 2016, 50, 5668–5675. [Google Scholar] [CrossRef] [Green Version]
- Andrady, A.L. Microplastics in the marine environment. Mar. Pollut. Bull. 2011, 62, 1596–1605. [Google Scholar] [CrossRef] [PubMed]
- Gigault, J.; Pedrono, B.; Maxit, B.; Ter Halle, A. Marine plastic litter: The unanalyzed nano-fraction. Environ. Sci. Nano 2016, 3, 346–350. [Google Scholar] [CrossRef]
- Wright, S.L.; Thompson, R.C.; Galloway, T.S. The physical impacts of microplastics on marine organisms: A review. Environ. Pollut. 2013, 178, 483–492. [Google Scholar] [CrossRef]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 2015, 75, 63–82. [Google Scholar] [CrossRef]
- Galloway, T.S.; Cole, M.; Lewis, C. Interactions of microplastic debris throughout the marine ecosystem. Nat. Ecol. Evol. 2017, 1, 116. [Google Scholar] [CrossRef]
- Nel, A.E.; Madler, L.; Velegol, D.; Xia, T.; Hoek, E.M.V.; Somasundaran, P.; Klaessig, F.; Castranova, V.; Thompson, M. Understanding biophysicochemical interactions at the nano-bio interface. Nat. Mater. 2009, 8, 543–557. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, C.; Diaz Banos, F.G.; Esteban, M.A.; Cuesta, A. Functionalized Nanoplastics (NPs) Increase the Toxicity of Metals in Fish Cell Lines. Int. J. Mol. Sci. 2021, 22, 7141. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.; Galloway, T.S. Ingestion of Nanoplastics and Microplastics by Pacific Oyster Larvae. Environ. Sci. Technol. 2015, 49, 14625–14632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattsson, K.; Johnson, E.V.; Malmendal, A.; Linse, S.; Hansson, L.A.; Cedervall, T. Brain damage and behavioural disorders in fish induced by plastic nanoparticles delivered through the food chain. Sci. Rep. 2017, 7, 11452. [Google Scholar] [CrossRef] [PubMed]
- Tallec, K.; Huvet, A.; Di Poi, C.; Gonzalez-Fernandez, C.; Lambert, C.; Petton, B.; Le Goic, N.; Berchel, M.; Soudant, P.; Paul-Pont, I. Nanoplastics impaired oyster free living stages, gametes and embryos. Environ. Pollut. 2018, 242, 1226–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ter Halle, A.; Ghiglione, J.F. Nanoplastics: A Complex, Polluting Terra Incognita. Environ. Sci. Technol. 2021, 55, 14466–14469. [Google Scholar] [CrossRef]
- Besseling, E.; Quik, J.T.K.; Sun, M.; Koelmans, A.A. Fate of nano- and microplastic in freshwater systems: A modeling study. Environ. Pollut. 2017, 220, 540–548. [Google Scholar] [CrossRef]
- Pradel, A.; Ferreres, S.; Veclin, C.; Hadri, H.E.; Gautier, M.; Grassl, B.; Gigault, J. Stabilization of Fragmental Polystyrene Nanoplastic by Natural Organic Matter: Insight into Mechanisms. ACS EST Water 2021, 1, 1198–1208. [Google Scholar] [CrossRef]
- Rowenczyk, L.; Leflaive, J.; Clergeaud, F.; Minet, A.; Feriol, J.; Gauthier, L.; Gigault, J.; Mouchet, F.; Ory, D.; Pinelli, E.; et al. Heteroaggregates of polystyrene nanospheres and organic matter: Preparation, characterization and evaluation of their toxicity on algae in environmentally relevant conditions. Nanomat 2021, 11, 482. [Google Scholar] [CrossRef]
- ISO 21427-1; Qualité de l’eau—Évaluation de la génotoxicité par le mesurage de l’induction de micronoyaux—Partie 1: Évaluation de la génotoxicité à l’aide de larves d’amphibiens. Evaluation of genotoxicity by measurement of their induction of micronuclei—Part 1: Evaluation of genotoxicity using amphibian larvae. ICS: 13.060.70; International Organization for Standardization: Geneva, Switzerland, 2006; p. 15.
- OECD. Test Guideline 219 Sediment—Water Chironomid Toxicity Test Using Spiked Water; OECD: Paris, France, 2004. [Google Scholar]
- Mouchet, F.; Gauthier, L. Genotoxicity of contaminants: Amphibian micronucleus assay. In Encyclopedia of Aquatic Toxicology. Comprehensive Handbook (or Practical Guide) of Ecotoxicological Terms; Blaise, C., Férard, J.F., Eds.; Springer Publishers: Dordrecht, The Netherlands, 2013; Volume 2, p. 1221. [Google Scholar]
- Environment Canada. Biological Test Method. Test for Survival and Growth in Sediment Using Larvae of Freshwater Midges (Chironomus tentans or Chironomus riparius); SPE 1/RM/32; Environment Canada: Otawa, ON, Canada, 1997.
- AFNOR. Détermination de la Toxicité des Sédiments vis-à-vis de Chironomus Riparius; XP T 90-339-1; Association Française de NORmalisation (AFNOR): Paris, France, 2004. [Google Scholar]
- U.S. Environmental Protection Agenc. Methods for Measuring the Toxicity and Bioaccumulation of Sediment-Associated Contaminants with Freshwater Invertebrates, 2nd ed.; EPA 600/R-99/064; U.S. Environmental Protection Agency: Washington, DC, USA, 2000.
- Dias, V.; Vasseur, C.; Bonzom, J.-M. Exposure of Chironomus riparius larvae to uranium: Effects on survival, development time, growth, and mouthpart deformities. Chemosphere 2008, 71, 574–581. [Google Scholar] [CrossRef]
- Warwick, W.F.; Tisdale, N.A. Morphological deformities in Chironomus, Cryptochironomus, and Procladius larvae (Diptera: Chironomidae) from two differentially stressed sites in Tobin Lake, Saskatchewan. Can. J. Fish. Aquat. Sci. 1988, 45, 1123–1144. [Google Scholar] [CrossRef]
- Vermeulen, A.C.; Dall, P.C.; Lindegaard, C.; Ollevier, F.; Goddeeris, B. Improving the methodology of chironomid, deformation analysis for sediment toxicity assessment: A case study in three Danish lowland streams. Arch. Für Hydrobiol. 1998, 144, 103–125. [Google Scholar] [CrossRef]
- Mouchet, F.; Gauthier, L.; Baudrimont, M.; Gonzalez, P.; Mailhes, C.; Ferrier, V.; Devaux, A. Comparative evaluation of the toxicity and genotoxicity of cadmium in amphibian larvae (Xenopus laevis and Pleurodeles waltl) using the comet assay and the micronucleus test. Environ. Toxicol. 2007, 22, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Mouchet, F.; Landois, P.; Datsyuk, V.; Puech, P.; Pinelli, E.; Flahaut, E.; Gauthier, L. International amphibian micronucleus standardized procedure (ISO 21427-1) for in vivo evaluation of double-walled carbon nanotubes toxicity and genotoxicity in water. Environ. Toxicol. 2011, 26, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Nieuwkoop, P.D.; Faber, J. Normal Table of Xenopus laevis (Daudin). In A Systematical and Chronological Survey of the Development from the Fertilized Egg till the End of Metamorphosis; Garland Publishing: Amsterdam, The Netherlands, 1956. [Google Scholar]
- Madre, J.F. Logiciel Mesurim; Académie d’Amiens: Amiens, France. Available online: http://www.ac-amiens.fr/pedagogie/svt/info/logiciels/Mesurim2/Index.htm (accessed on 1 July 2022).
- McGill, R.; Tuckey, J.; Larsen, W. Variations of box plots. Am. Statist. 1978, 32, 12–16. [Google Scholar]
- Khan, F.R. Prevalence, Fate and Effects of Plastic in Freshwater Environments: New Findings and Next Steps. Toxics 2020, 8, 72. [Google Scholar] [CrossRef]
- Mouchet, F.; Landois, P.; Puech, P.; Pinelli, E.; Flahaut, E.; Gauthier, L. CNT ecotoxicity in amphibians: Assessment of Multi Walled Carbon Nanotubes (MWNT) and Comparison with Double Walled Carbon Nanotubes (DWNT). Special focus Environmental Toxicity of Nanoparticles. Nanomedicine 2010, 5, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Mouchet, F.; Gancet, C.; Flahaut, E.; Pinelli, E.; Boutonnet, J.C.; Gauthier, L. International standardized procedures for in vivo evaluation of multi-walled carbon nanotube toxicity in water. Toxicol. Environ. Chem. 2016, 98, 829–847. [Google Scholar] [CrossRef] [Green Version]
- Besseling, E.; Wang, B.; Lürling, M.; Koelmans, A.A. Nanoplastic affects growth of S. obliquus and reproduction of D. magna. Environ. Sci. Technol. 2014, 48, 12336–12343. [Google Scholar] [CrossRef]
- Tang, J.; Wang, X.; Yin, J.; Han, Y.; Yang, J.; Lu, X.; Xie, T.; Akbar, S.; Lyu, K.; Yang, Z. Molecular characterization of thioredoxin reductase in water flea Daphnia magna and its expression regulation by polystyrene microplastics. Aquat. Toxicol. 2019, 208, 90–97. [Google Scholar] [CrossRef]
- De Felice, B.; Bacchetta, R.; Santo, N.; Ortenzi, M.A.; Antenucci, S.; Tremolada, P.; Parolin, M. Polystyrene microplastics did not affect body growth and swimming activity in Xenopus laevis tadpoles. Environ. Sci. Pollut. Res. Int. 2018, 25, 34644–34651. [Google Scholar] [CrossRef]
- Matthews, S.; Genbo, X.E.; Dumont, E.R.; Meola, V.; Pikuda, O.; Cheong, R.S.; Guo, M.; Tahara, R.; Larsson, H.C.E.; Tufenkji, N. Polystyrene micro- and nanoplastics affect locomotion and daily activity of Drosophila melanogaster. Environ. Sci. Nano 2021, 8, 110–121. [Google Scholar] [CrossRef]
- Li, X.; He, E.; Jiang, K.; Peijnenburg, W.J.G.M.; Qiu, H. The crucial role of a protein corona in determining the aggregation kinetics and colloidal stability of polystyrene nanoplastics. Water Res. 2021, 190, 116742. [Google Scholar] [CrossRef] [PubMed]
- Musino, D.; Genix, A.C.; Chausee, T.; Guy, L.; Meissnet, N.; Kozak, R.; Bizien, T.; Oberdisse, J. Aggregate Formation of Surface-Modified Nanoparticles in Solvents and Polymer Nanocomposites. Langmuir Am. Chem. Soc. 2018, 34, 3010–3020. [Google Scholar] [CrossRef]
- Ding, J.; Zhang, S.; Razanajatovo, R.M.; Zou, H.; Zhu, W. Accumulation, Tissue Distribution, and Biochemical Effects of Polystyrene Microplastics in the Freshwater Fish Red Tilapia (Oreochromis niloticus). Environ. Pollut. 2018, 238, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Feng, C.; Pang, W.; Tian, C.; Zaho, Y. Nanoplastic-Induced Genotoxicity and Intestinal Damage in Freshwater Benthic Clams (Corbicula f luminea): Comparison with Microplastics. ACS Nano 2021, 15, 9469–9481. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Shelver, W.L. Micro- and nanoplastic-mediated pathophysiological changes in rodents, rabbits, and chickens: A review. J. Food Prot. 2021, 84, 1480–1495. [Google Scholar] [CrossRef]
- Pan, Y.; Neuss, S.; Leifert, A.; Fischler, M.; Simon, U.; Schmid, G.; Brandau, W.; Jahnen-Dechent, W. Size-dependent cytotoxicity of gold nanoparticles. Small 2017, 3, 1941–1949. [Google Scholar] [CrossRef]
- Shang, L.; Nienhaus, K.; Nienhaus, G.U. Engineered nanoparticles interacting with cells: Size matters. J. Nanobiotechnol. 2014, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- Jeong, C.B.; Won., E.J.; Kang, H.M.; Lee, M.C.; Hwang, D.S.; Hwang, U.K.; Zhou, B.; Souissi, S.; Lee, S.J.; Lee, J.S. Microplastic size-dependent toxicity, oxidative stress induction, and p-JNK and p-P38 activation in the monogonont rotifer (Brachionus koreanus). Environ. Sci. Technol. 2016, 50, 8849–8857. [Google Scholar] [CrossRef]
- Zettler, E.R.; Mincer, T.J.; Amaral-Zettler, L. Life in the « plastisphere: Microbial Comunities on Plastic Marine Debris. Environ. Sci. Technol. 2013, 47, 7137–7146. [Google Scholar] [CrossRef]
- Fadare, O.O.; Wan, B.; Guo, L.H.; Xin, Y.; Qin, W.; Yang, Y. Humic acid alleviates the toxicity of polystyrene nanoplastic particles to Daphnia magna. Environ. Sci. Nano 2019, 6, 1466–1477. [Google Scholar] [CrossRef]
- Ikuzawa, M.; Yasumasu, S.; Kobayashi, K.I.; Inokuchi, T.; Iuchi, I. Stomach Remodeling-Associated Changes of Hb/Kb-ATPase Subunit Expression in Xenopus laevis and Hb/Kb-ATPase-dependant Acide Secretion in tadpole Stomach. J. Exp. Zoll. A Comp. Exp. Biol. 2004, 301A, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Browne, M.A.; Galloway, T.S.; Thompson, R. Microplastic—An emerging contaminant of potential concern? Integr. Environ. Assess. Manag. 2007, 3, 559–561. [Google Scholar] [CrossRef] [PubMed]
- Phuong, N.N.; Zalouk-Vergnoux, A.; Poirier, L.; Kamari, A.; Châtel, A.; Mouneyrac, C.; Lagarde, F. Is there any consistency between the microplastics found in the field and those used in laboratory experiments? Environ. Pollut. 2016, 211, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Carlos de Sa, L.; Oliveira, M.; Ribeiro, F.; Lopes, T.; Martyn, R.; Futter, N. Studies of the effects of microplastics on aquatic organisms: What do we know and where should we focus our efforts in the future? Sci. Total Environ. 2018, 645, 1029–1039. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Particles | PS50 | PS350 | PSHA |
|---|---|---|---|
| Primary size (nm) | 50 | 350 | 300–500 in the medium in which they were designed (I = 700 mmol·L−1) |
| Transmission Electron Microscopy observation |  |  |  |
| Polydispersity | Slightly polydisperse, made of particles from 10 to 35 nm | Monodisperse with a diameter between 370 and 410 nm | Polydisperse |
| Shape | Spherical | Spherical | Non-spherical |
| Nature of surface | –COOH | –COOH | –COOH/NH2 |
| Surface charge | Negative | Negative | Negative |
| Functionalization | Less carboxylic moieties | More carboxylic moieties |
| Size Type | Medium | pH | PS50 dzH (nm) | PS350 dzH (nm) | PSHA dzH (nm) |
|---|---|---|---|---|---|
| Size in water | MilliQ | 7.00 | 45 ± 1 | 349 ± 3 | 300–500 ** |
| Size in media | Chironomus | 8.23 | 2356 ± 294 * | 401 ± 8 | 528 ± 22 |
| Size in media | Xenopus | 8.05 | 7255 ± 386 * | 403 ± 10 | 484 ± 64 |
| Size in media after exposure | Xenopus + (HA) | 6.75 | 740 ± 124 * | 414 ± 6.8 | 1053 ± 22 * |
| Particles | Zeta Potential in Chironomus Exposition Medium (mV) | Zeta Potential in Xenopus Exposition Medium (mV) |
|---|---|---|
| PS50 | −16.4 (±0.3) | −17.8 (±0.9) |
| PS350 | −30.5 (±1.1) | −24.8 (±0.7) |
| PSHA | −23.0 (±0.5) | −25.5 (±2.8) |
| Polymer | Treatment (µg/L−1) | Deformity Frequency (%) |
|---|---|---|
| PS50 | 0 | 8.3 |
| 0.1 | 11.1 | |
| 1 | 7.5 | |
| 10 | 11.1 | |
| 100 | 7.1 | |
| PS350 | 0 | 8.3 |
| 0.1 | 4.5 | |
| 1 | 8.6 | |
| 10 | 10.9 | |
| 100 | 7.1 | |
| PSHA | 0 | 0 |
| HA Control | 3.4 | |
| 0.1 | 8.3 | |
| 1 | 0 | |
| 10 | 0 | |
| 100 | 6.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mouchet, F.; Rowenczyk, L.; Minet, A.; Clergeaud, F.; Silvestre, J.; Pinelli, E.; Ferriol, J.; Leflaive, J.; Ten-Hage, L.; Gigault, J.; et al. Ecotoxicity of Heteroaggregates of Polystyrene Nanospheres in Chironomidae and Amphibian. Nanomaterials 2022, 12, 2730. https://doi.org/10.3390/nano12152730
Mouchet F, Rowenczyk L, Minet A, Clergeaud F, Silvestre J, Pinelli E, Ferriol J, Leflaive J, Ten-Hage L, Gigault J, et al. Ecotoxicity of Heteroaggregates of Polystyrene Nanospheres in Chironomidae and Amphibian. Nanomaterials. 2022; 12(15):2730. https://doi.org/10.3390/nano12152730
Chicago/Turabian StyleMouchet, Florence, Laura Rowenczyk, Antoine Minet, Fanny Clergeaud, Jérôme Silvestre, Eric Pinelli, Jessica Ferriol, Joséphine Leflaive, Loïc Ten-Hage, Julien Gigault, and et al. 2022. "Ecotoxicity of Heteroaggregates of Polystyrene Nanospheres in Chironomidae and Amphibian" Nanomaterials 12, no. 15: 2730. https://doi.org/10.3390/nano12152730
APA StyleMouchet, F., Rowenczyk, L., Minet, A., Clergeaud, F., Silvestre, J., Pinelli, E., Ferriol, J., Leflaive, J., Ten-Hage, L., Gigault, J., Ter Halle, A., & Gauthier, L. (2022). Ecotoxicity of Heteroaggregates of Polystyrene Nanospheres in Chironomidae and Amphibian. Nanomaterials, 12(15), 2730. https://doi.org/10.3390/nano12152730