Magnetite Nanoparticles Induce Genotoxicity in the Lungs of Mice via Inflammatory Response

 ,
,
Abstract
:1. Introduction
2. Results
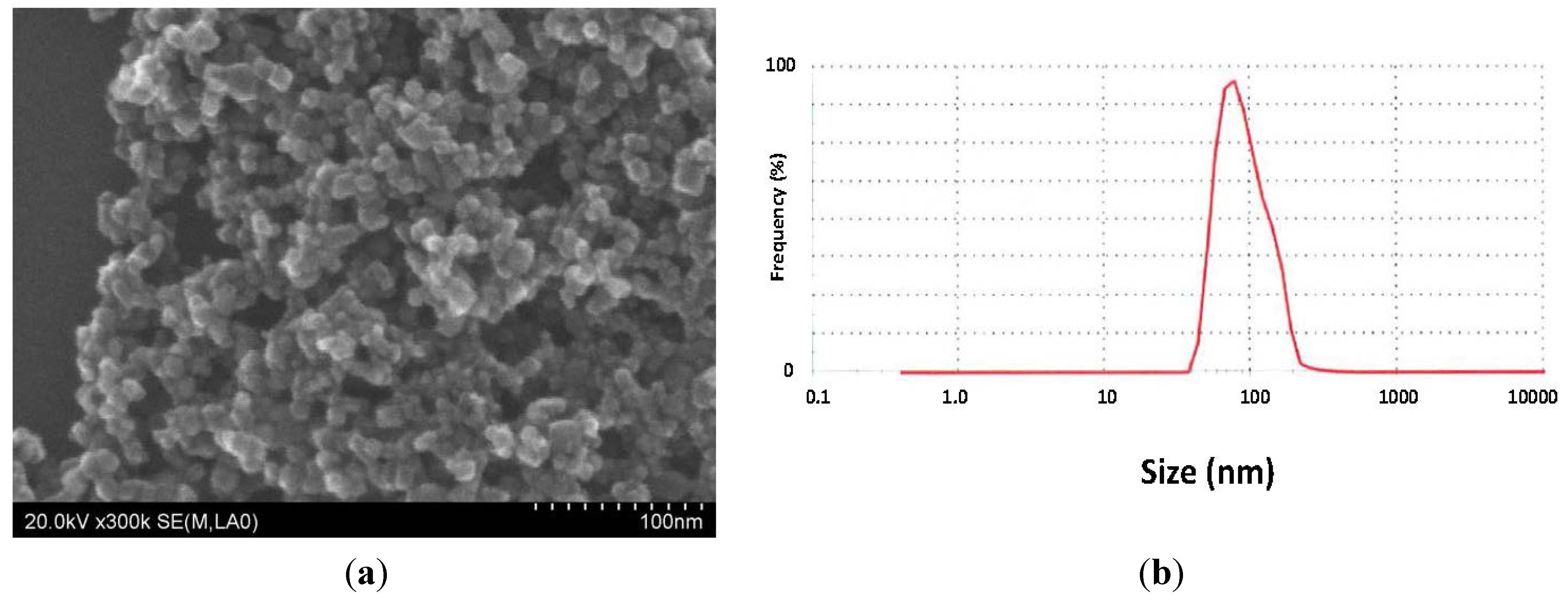
2.1. Characterization of MGT

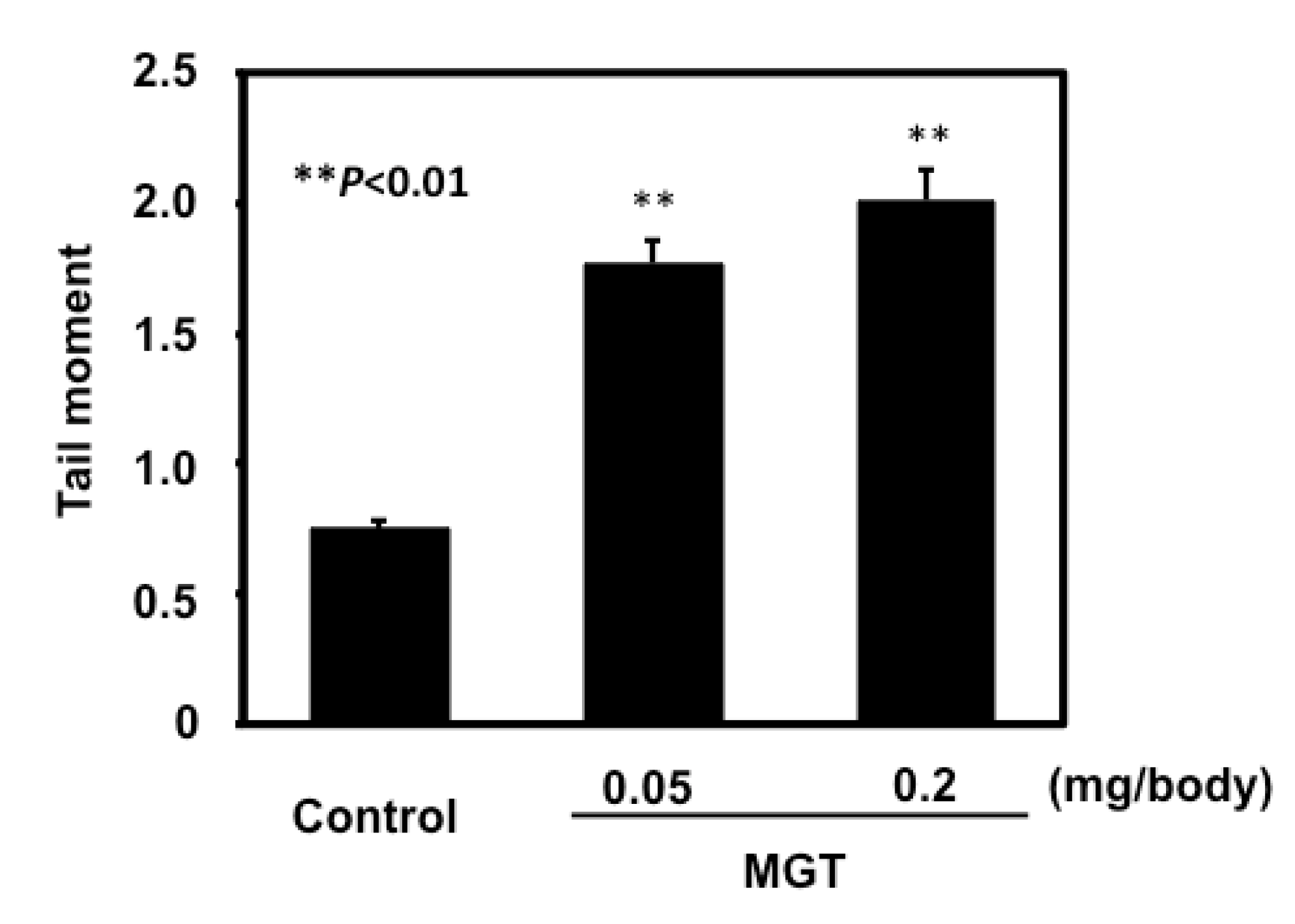
2.2. In Vivo Genotoxicity of MGT

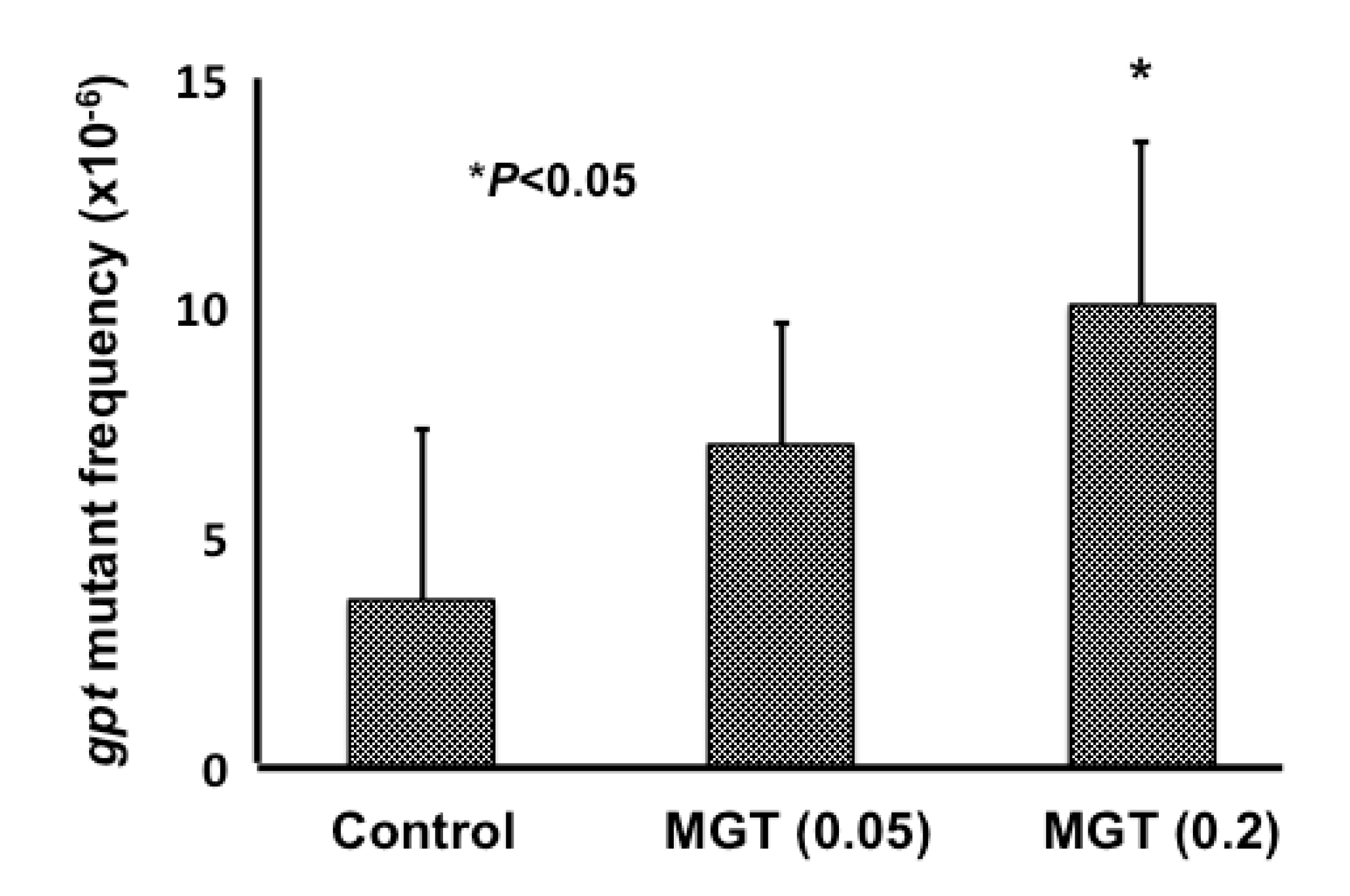
2.3. gpt and Spi− Mutations in the Lungs of gpt Transgenic Mice Treated with MGT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Mouse ID | Number of colonies | MF (×10−6) | Average MF (×10−6) * | ||
|---|---|---|---|---|---|---|
| Mutant | Total | |||||
| Control † | 1 | 6 | 747,000 | 8.03 | - | |
| 2 | 2 | 592,500 | 3.38 | - | ||
| 3 | 14 | 2,998,500 | 4.67 | - | ||
| 4 | 3 | 2,937,000 | 1.02 | - | ||
| 5 | 2 | 1,759,500 | 1.14 | - | ||
| Total | 27 | 9,034,500 | - | 3.65 ± 3.69 | ||
| MGT | 0.05 (mg/body) × 4 | 1 | 2 | 621,900 | 3.22 | - |
| 2 | 7 | 903,000 | 7.75 | - | ||
| 3 | 5 | 666,000 | 7.51 | - | ||
| 4 | 5 | 814,500 | 6.14 | - | ||
| 5 | 7 | 669,000 | 10.46 | - | ||
| Total | 26 | 3,674,400 | - | 7.02 ± 2.64 | ||
| 0.2 (mg/body) × 4 | 1 | 2 | 502,500 | 3.98 | - | |
| 2 | 5 | 475,500 | 10.52 | - | ||
| 3 | 5 | 546,000 | 9.16 | - | ||
| 4 | 12 | 846,000 | 14.18 | - | ||
| 5 | 5 | 522,000 | 9.58 | - | ||
| 6 | 1 | 78,000 | 12.82 | - | ||
| Total | 30 | 2,970,000 | - | 10.04 ± 3.54 ‡ | ||
| Type of mutation | Control | MGT | P value * | ||||
|---|---|---|---|---|---|---|---|
| Number of mutants (%) | Specific MF † (×10−6) | Number of mutants (%) | Specific MF † (×10−6) | ||||
| Base substitution | Transition | G:C to A:T | 8 (29.6) | 0.89 | 14 (46.7) | 4.71 | 0.00002 |
| A:T to G:C | 4 (14.8) | 0.44 | 2 (6.7) | 0.67 | 0.62571 | ||
| Transversion | G:C to T:A | 6 (22.2) | 0.66 | 6 (20) | 2.02 | 0.04258 | |
| G:C to C:G | 0 (0) | 0.00 | 0 (0) | 0.00 | - | ||
| A:T to T:A | 0 (0) | 0.00 | 0 (0) | 0.00 | - | ||
| A:T to C:G | 2 (7.4) | 0.22 | 2 (6.7) | 0.67 | 0.2417 | ||
| Insertion | 2 (7.4) | 0.22 | 1 (3.3) | 0.34 | 0.73017 | ||
| Deletion | 5 (18.5) | 0.55 | 4 (13.3) | 1.35 | 0.17072 | ||
| Others | 0 (0) | 0.00 | 1 (3.3) | 0.34 | 0.08114 | ||
| Total | 27(100) | - | 30 (100) | - | - | ||
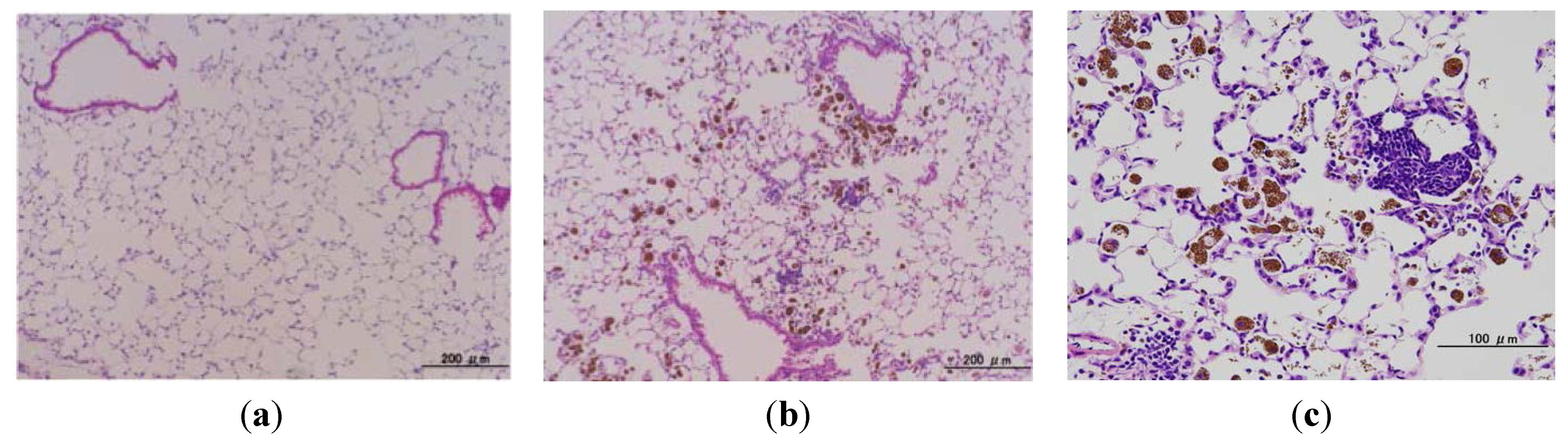
2.4. Microscopic Findings in the Lungs of gpt Delta Transgenic Mice Administered MGT

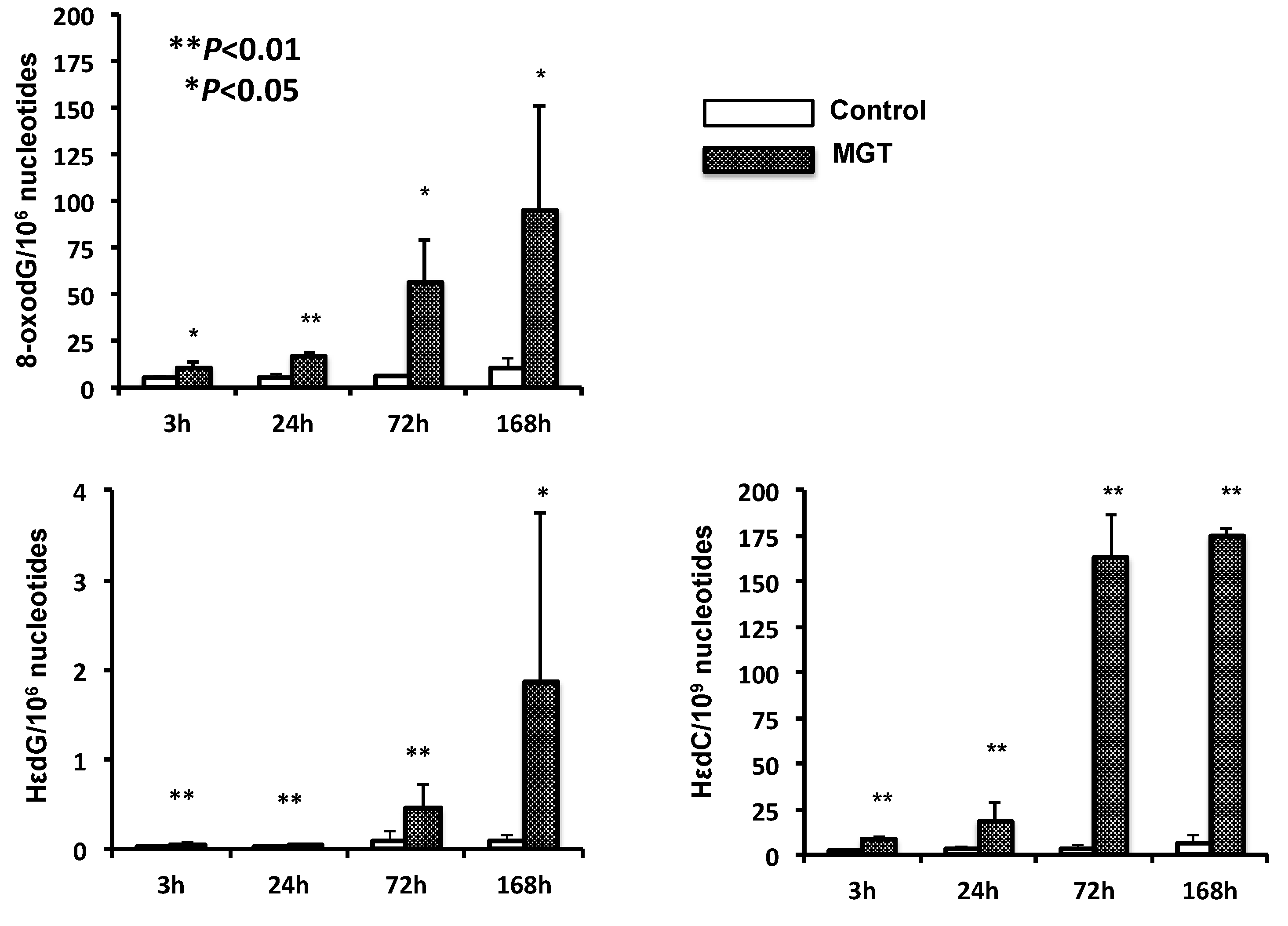
2.5. Quantification of Oxidative and Lipid Peroxide-Related DNA Adducts

3. Discussion
4. Experimental Section
4.1. Materials and Chemicals
4.2. Animals
4.3. Preparation and Characterization of MGT
4.4. Comet Assay
4.5. gpt and Spi− Mutation Assay
4.6. Histopathological Evaluation
4.7. Oxidative and Lipid Peroxide-Related DNA Adduct Formation
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cabreraa, L.; Gutierreza, S.; Menendezb, N.; Moralesc, M.P.; Herrasti, P. Magnetite nanoparticles: Electrochemical synthesis and characterization. Electrochim. Acta 2008, 53, 3436–3441. [Google Scholar] [CrossRef]
- Parveen, S.; Misra, R.; Sahoo, S.K. Nanoparticles: A boon to drug delivery, therapeutics, diagnostics and imaging. Nanomedicine 2012, 8, 147–166. [Google Scholar] [CrossRef]
- Hilger, I.; Kaiser, W.A. Iron oxide-based nanostructures for MRI and magnetic hyperthermia. Nanomedicine 2012, 7, 1443–1459. [Google Scholar] [CrossRef]
- Caruthers, S.D.; Wickline, S.A.; Lanza, G.M. Nanotechnological applications in medicine. Curr. Opin. Biotechnol. 2007, 18, 26–30. [Google Scholar] [CrossRef]
- Simberg, D.; Duza, T.; Park, J.H.; Essler, M.; Pilch, J.; Zhang, L.; Derfus, A.M.; Yang, M.; Hoffman, R.M.; Bhatia, S.; et al. Biomimetic amplification of nanoparticle homing to tumors. Proc. Natl. Acad. Sci. USA 2007, 104, 932–936. [Google Scholar] [CrossRef]
- Xu, C.J.; Sun, S.H. Monodisperse magnetic nanoparticles for biomedical applications. Polym. Int. 2007, 56, 821–826. [Google Scholar] [CrossRef]
- Guichard, Y.; Schmit, J.; Darne, C.; Gaté, L.; Goutet, M.; Rousset, D.; Rastoix, O.; Wrobel, R.; Witschger, O.; Martin, A.; et al. Cytotoxicity and genotoxicity of nanosized and microsized titanium dioxide and iron oxide particles in Syrian hamster embryo cells. Ann. Occup. Hyg. 2012, 56, 631–644. [Google Scholar]
- Aranda, A.; Sequedo, L.; Tolosa, L.; Quintas, G.; Burello, E.; Castell, J.V.; Gombau, L. Dichloro-dihydro-fluorescein diacetate (DCFH-DA) assay: A quantitative method for oxidative stress assessment of nanoparticle-treated cells. Toxicol. Vitro 2013, 27, 954–963. [Google Scholar] [CrossRef]
- Singh, N.; Jenkins, G.J.; Asadi, R.; Doak, S.H. Potential toxicity of superparamagnetic iron oxide nanoparticles (SPION). Nano Rev. 2010, 1, 5358–5373. [Google Scholar]
- Ramesh, V.; Ravichandran, P.; Copeland, C.L.; Gopikrishnan, R.; Biradar, S.; Goornavar, V.; Ramesh, G.T.; Hall, J.C. Magnetite induces oxidative stress and apoptosis in lung epithelial cells. Mol. Cell Biochem. 2012, 363, 225–234. [Google Scholar] [CrossRef]
- Könczöl, M.; Ebeling, S.; Goldenberg, E.; Treude, F.; Gminski, R.; Gieré, R.; Grobéty, B.; Rothen-Rutishauser, B.; Merfort, I.; Mersch-Sundermann, V. Cytotoxicity and genotoxicity of size-fractionated iron oxide (magnetite) in A549 human lung epithelial cells: Role of ROS, JNK, and NF-κB. Chem. Res. Toxicol. 2011, 24, 1460–1475. [Google Scholar] [CrossRef]
- Karlsson, H.L.; Gustafsson, J.; Cronholm, P.; Möller, L. Size-dependent toxicity of metal oxide particles—A comparison between nano- and micrometer size. Toxicol. Lett. 2009, 188, 112–118. [Google Scholar] [CrossRef]
- Ma, P.; Luo, Q.; Chen, J.; Gan, Y.; Du, J.; Ding, S.; Xi, Z.; Yang, X. Intraperitoneal injection of magnetic Fe3O4-nanoparticle induces hepatic and renal tissue injury via oxidative stress in mice. Int. J. Nanomed. 2012, 7, 4809–4818. [Google Scholar]
- Weissleder, R.; Stark, D.D.; Engelstad, B.L.; Bacon, B.R.; Compton, C.C.; White, D.L.; Jacobs, P.; Lewis, J. Superparamagnetic iron oxide: Pharmacokinetics and toxicity. Am. J. Roentgenol. 1989, 152, 167–173. [Google Scholar] [CrossRef]
- Singh, N.; Jenkins, G.J.; Nelson, B.C.; Marquis, B.J.; Maffeis, T.G.; Brown, A.P.; Williams, P.M.; Wright, C.J.; Doak, S.H. The role of iron redox state in the genotoxicity of ultrafine superparamagnetic iron oxide nanoparticles. Biomaterials 2012, 33, 163–170. [Google Scholar] [CrossRef]
- Szalay, B.; Tátrai, E.; Nyírő, G.; Vezér, T.; Dura, G. Potential toxic effects of iron oxide nanoparticles in in vivo and in vitro experiments. J. Appl. Toxicol. 2012, 32, 446–453. [Google Scholar] [CrossRef]
- Watanabe, M.; Yoneda, M.; Morohashi, A.; Okamoto, D.; Sato, A.; Kurioka, D.; Hirokawa, H.; Shiraishi, T.; Kawai, K.; Kasai, K.; et al. Effects of Fe3O4-based magnetic nanoparticles on A549 cells. Int. J. Mol. Sci. 2013, 14, 15548–15560. [Google Scholar]
- Kawanishi, M.; Ogo, S.; Ikemoto, M.; Totsuka, Y.; Ishino, K.; Wakabayahsi, K.; Yagi, T. Genotoxicity and reactive oxygen species production induced by magnetite nanoparticles in mammalian cells. J. Toxicol. Sci. 2013, 38, 503–511. [Google Scholar] [CrossRef]
- Park, E.J.; Kim, H.; Kim, Y.; Yi, J.; Choi, K.; Park, K. Inflammatory responses may be induced by a single intratracheal instillation of iron nanoparticles in mice. Toxicology 2010, 275, 65–71. [Google Scholar] [CrossRef]
- Carr, G.J.; Gorelick, N.J. Mutational spectra in transgenic animal research: Data analysis and study design based upon the mutant or mutation frequency. Environ. Mol. Mutagen. 1996, 28, 405–413. [Google Scholar] [CrossRef]
- Totsuka, Y.; Higuchi, T.; Imai, T.; Nishikawa, A.; Nohmi, T.; Kato, T.; Masuda, S.; Kinae, N.; Hiyoshi, K.; Ogo, S.; et al. Genotoxicity of nano/microparticles in in vitro micronuclei, in vivo comet and mutation assay systems. Part. Fibre Toxicol. 2009, 6. [Google Scholar] [CrossRef]
- Kato, T.; Totsuka, Y.; Ishino, K.; Matsumoto, Y.; Tada, Y.; Nakae, D.; Goto, S.; Masuda, S.; Ogo, S.; Kawanishi, M.; et al. Genotoxicity of multi-walled carbon nanotubes in both in vitro and in vivo assay systems. Nanotoxicology 2013, 7, 452–461. [Google Scholar]
- Totsuka, Y.; Kato, T.; Masuda, S.; Ishino, K.; Matsumoto, Y.; Goto, S.; Kawanishi, M.; Yagi, T.; Wakabayashi, K. In vitro and in vivo genotoxicity induced by fullerene (C60) and kaolin. Genes Environ. 2011, 33, 14–20. [Google Scholar] [CrossRef]
- Kreutzer, D.A.; Essigmann, J.M. Oxidized, deaminated cytosines are a source of C → T transitions in vivo. Proc. Natl. Acad. Sci. USA 1998, 95, 3578–3582. [Google Scholar] [CrossRef]
- Bird, A.P. CpG-rich islands and the function of DNA methylation. Nature 1986, 321, 209–213. [Google Scholar] [CrossRef]
- Shibutani, S.; Takeshita, M.; Grollman, A.P. Insertion of specific bases during DNA synthesis past the oxidation-damaged base 8-oxodG. Nature 1991, 349, 431–434. [Google Scholar] [CrossRef]
- Moriya, M. Single-stranded shuttle phagemid for mutagenesis studies in mammalian cells: 8-Oxoguanine in DNA induces targeted G.C-->T.A transversions in simian kidney cells. Proc. Natl. Acad. Sci. USA 1993, 90, 1122–1126. [Google Scholar] [CrossRef]
- Pollack, M.; Yang, I.Y.; Kim, H.Y.; Blair, I.A.; Moriya, M. Translesion DNA Synthesis across the heptanone-etheno-2′-deoxycytidine adduct in cells. Chem. Res. Toxicol. 2006, 19, 1074–1079. [Google Scholar] [CrossRef]
- Yang, I.Y.; Hashimoto, K.; de Wind, N.; Blair, I.A.; Moriya, M. Two distinct translesion synthesis pathways across a lipid peroxidation-derived DNA adduct in mammalian cells. J. Biol. Chem. 2009, 284, 191–198. [Google Scholar] [CrossRef]
- Dostert, C.; Pétrilli, V.; van Bruggen, R.; Steele, C.; Mossman, B.T.; Tschopp, J. Innate immune activation through Nalp3 inflammasome sensing of asbestos and silica. Science 2008, 320, 674–677. [Google Scholar] [CrossRef]
- Cassel, S.L.; Eisenbarth, S.C.; Iyer, S.S.; Sadler, J.J.; Colegio, O.R.; Tephly, L.A.; Carter, A.B.; Rothman, P.B.; Flavell, R.A.; Sutterwala, F.S. The Nalp3 inflammasome is essential for the development of silicosis. Proc. Natl. Acad. Sci. USA 2008, 105, 9035–9040. [Google Scholar] [CrossRef]
- Kim, J.E.; Shin, J.Y.; Cho, M.H. Magnetic nanoparticles: An update of application for drug delivery and possible toxic effects. Arch. Toxicol. 2012, 86, 685–700. [Google Scholar] [CrossRef]
- Simberg, D.; Park, J.H.; Karmali, P.P.; Zhang, W.M.; Merkulov, S.; McCrae, K.; Bhatia, S.N.; Sailor, M.; Ruoslahti, E. Differential proteomics analysis of the surface heterogeneity of dextran iron oxide nanoparticles and the implications for their in vivo clearance. Biomaterials 2009, 30, 3926–3933. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.C.; Lee, J.H.; Lee, J.; Kim, H.Y.; Park, J.Y.; Cho, J.; Lee, J.; Han, D.W. Subtle cytotoxicity and genotoxicity differences in superparamagnetic iron oxide nanoparticles coated with various functional groups. Int. J. Nanomed. 2011, 6, 3219–3231. [Google Scholar]
- Estevanato, L.; Cintra, D.; Baldini, N.; Portilho, F.; Barbosa, L.; Martins, O.; Lacava, B.; Miranda-Vilela, A.L.; Tedesco, A.C.; Báo, S.; et al. Preliminary biocompatibility investigation of magnetic albumin nanosphere designed as a potential versatile drug delivery system. Int. J. Nanomed. 2011, 6, 1709–1717. [Google Scholar] [CrossRef]
- Nohmi, T.; Suzuki, T.; Masumura, K. Recent advances in the protocols of transgenic mouse mutation assays. Mutat. Res. 2000, 455, 191–215. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Totsuka, Y.; Ishino, K.; Kato, T.; Goto, S.; Tada, Y.; Nakae, D.; Watanabe, M.; Wakabayashi, K. Magnetite Nanoparticles Induce Genotoxicity in the Lungs of Mice via Inflammatory Response. Nanomaterials 2014, 4, 175-188. https://doi.org/10.3390/nano4010175
Totsuka Y, Ishino K, Kato T, Goto S, Tada Y, Nakae D, Watanabe M, Wakabayashi K. Magnetite Nanoparticles Induce Genotoxicity in the Lungs of Mice via Inflammatory Response. Nanomaterials. 2014; 4(1):175-188. https://doi.org/10.3390/nano4010175
Chicago/Turabian StyleTotsuka, Yukari, Kousuke Ishino, Tatsuya Kato, Sumio Goto, Yukie Tada, Dai Nakae, Masatoshi Watanabe, and Keiji Wakabayashi. 2014. "Magnetite Nanoparticles Induce Genotoxicity in the Lungs of Mice via Inflammatory Response" Nanomaterials 4, no. 1: 175-188. https://doi.org/10.3390/nano4010175
APA StyleTotsuka, Y., Ishino, K., Kato, T., Goto, S., Tada, Y., Nakae, D., Watanabe, M., & Wakabayashi, K. (2014). Magnetite Nanoparticles Induce Genotoxicity in the Lungs of Mice via Inflammatory Response. Nanomaterials, 4(1), 175-188. https://doi.org/10.3390/nano4010175