Carbon Nanotubes’ Effect on Mitochondrial Oxygen Flux Dynamics: Polarography Experimental Study and Machine Learning Models using Star Graph Trace Invariants of Raman Spectra

 ,
,  ,
,  , , , and
, , , and 
Abstract
:
1. Introduction
2. Results
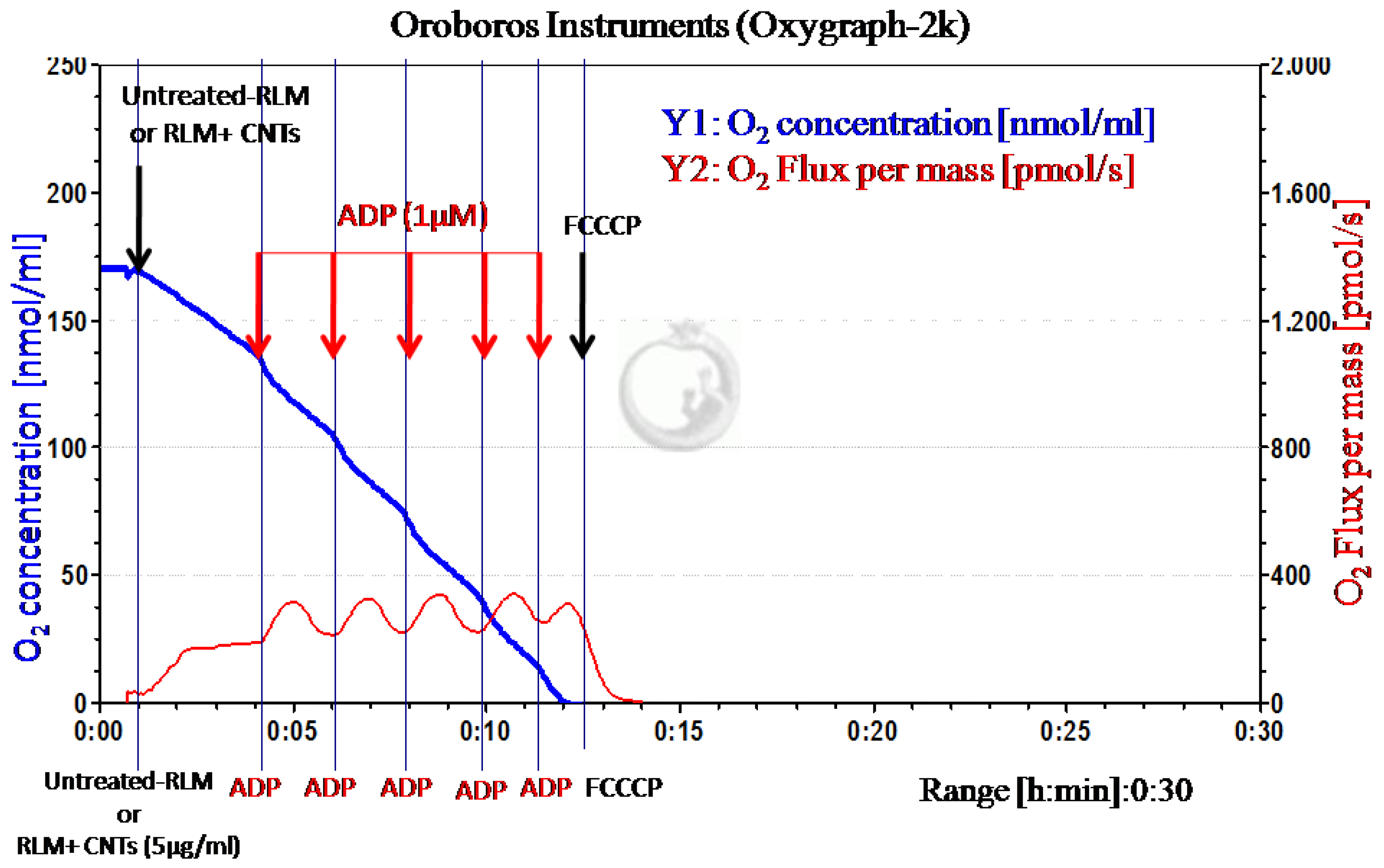
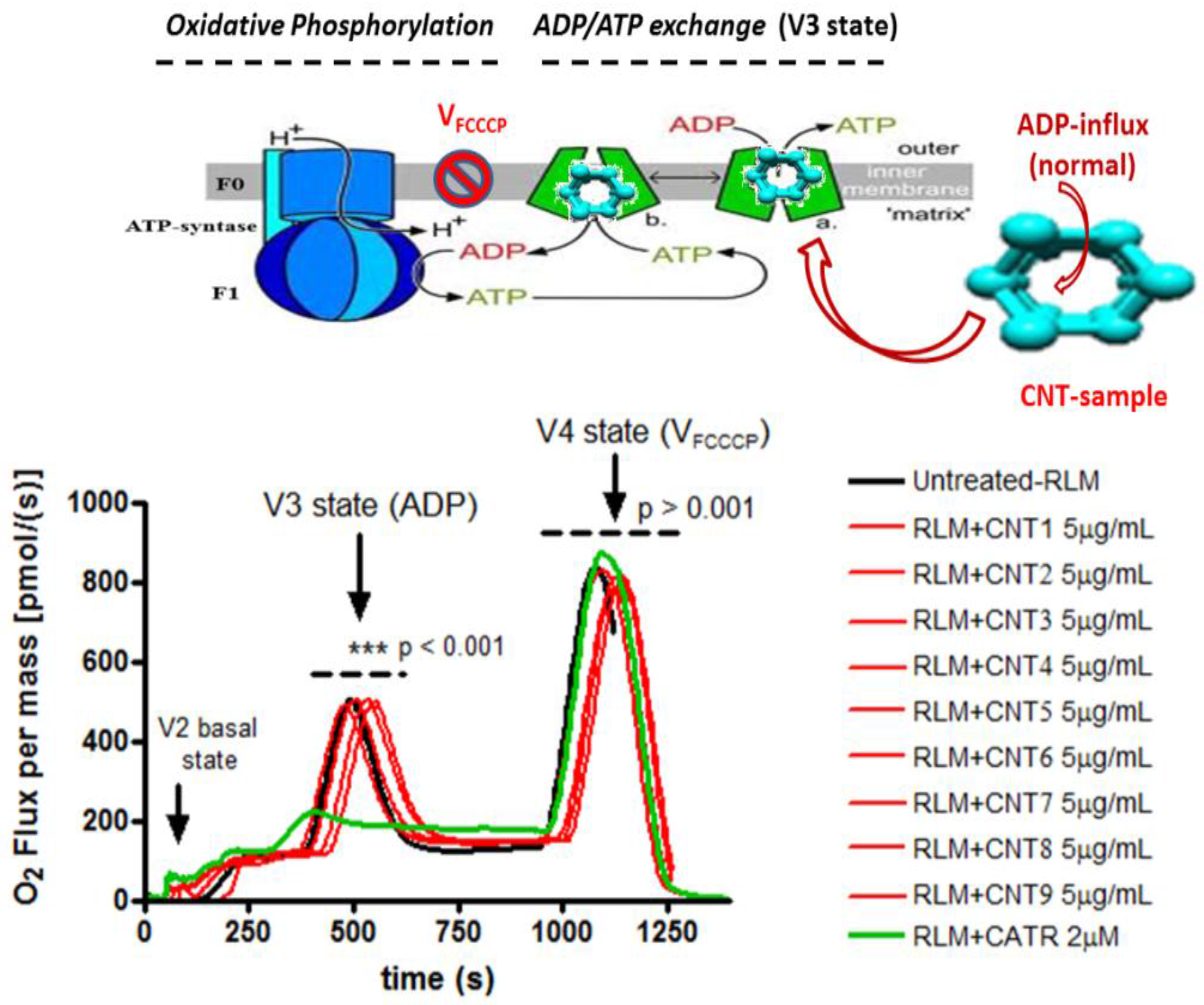
2.1. Experimental Results
2.2. Computational Results
3. Discussion
3.1. Discussion of Experimental Results
3.2. Discussion of Computational Results
4. Materials and Methods
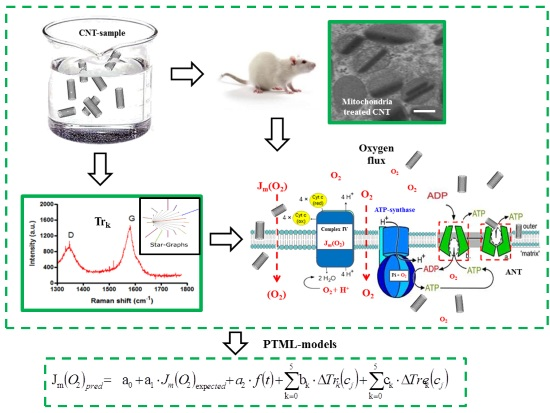
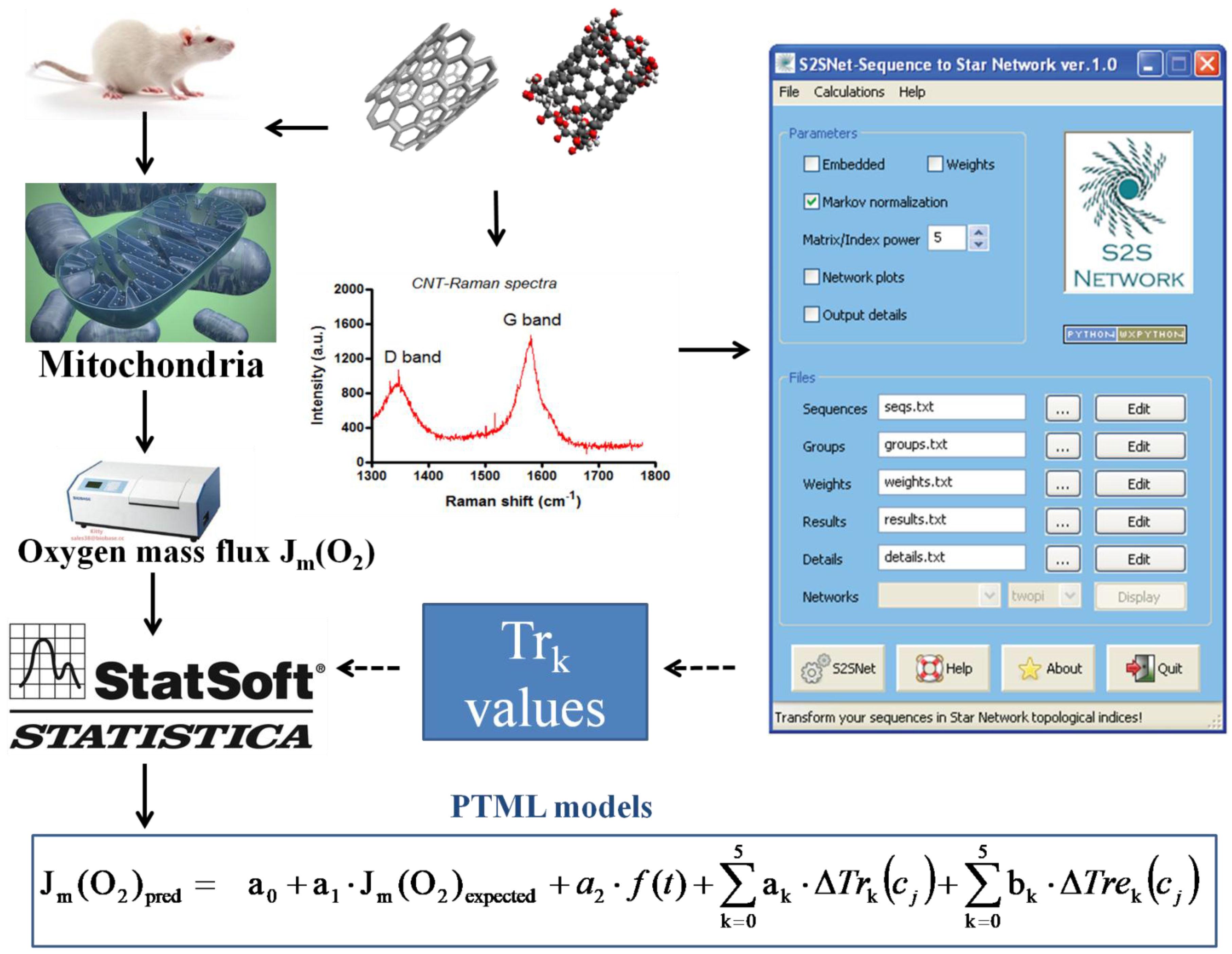
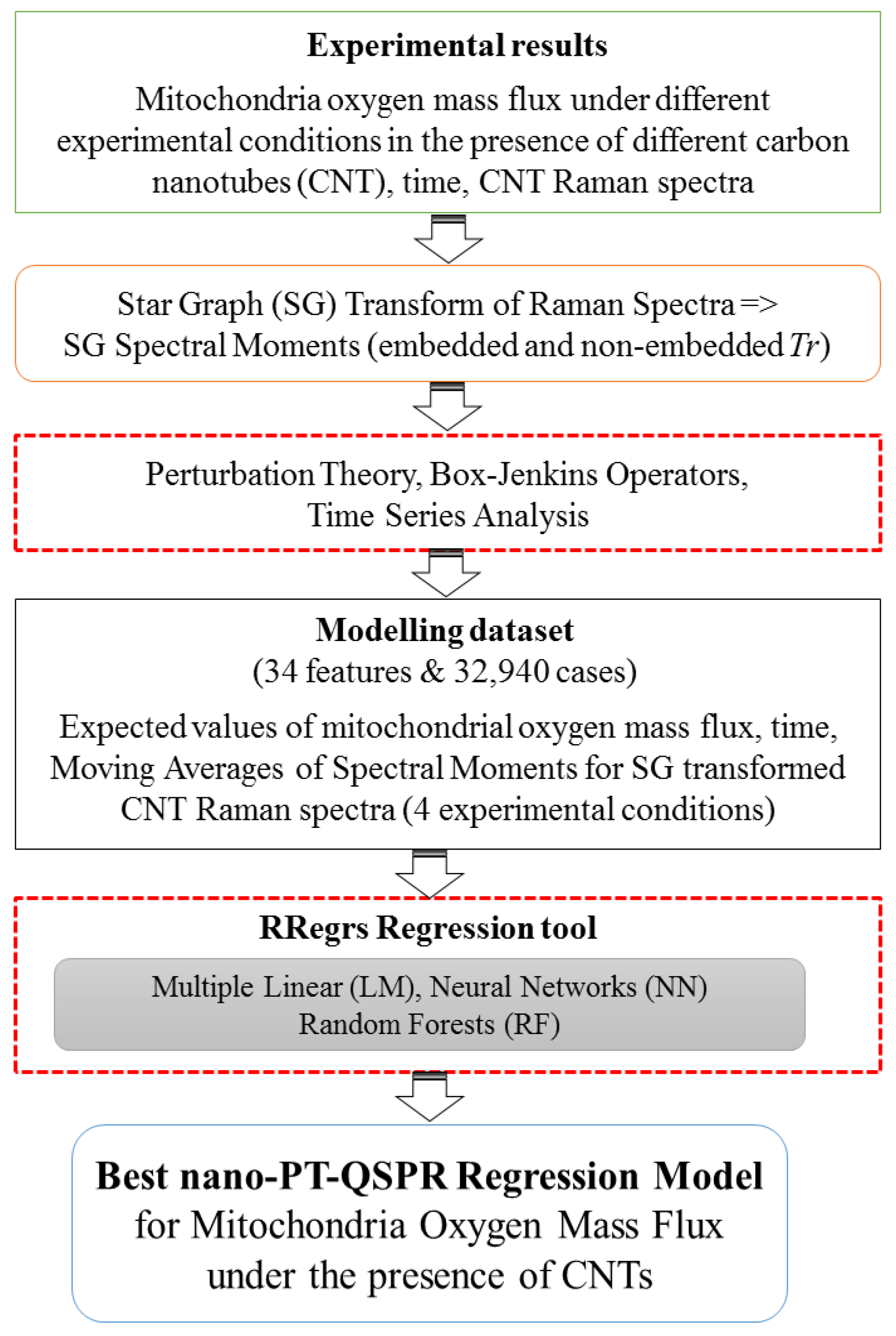
4.1. General Workflow
- (a)
- The measurement of the CNT effect on the mitochondrial oxygen mass flux with polarography;
- (b)
- The definition and calculation of the matrix trace invariants (Trk) of SG transforms of Raman spectra for a series of CNTs for the first time;
- (c)
- The use of the Trk values as input to seek new PTML models able to predict CNTs’ effect on mitochondrial oxygen mass flux.
- (1)
- Experimental measurements of the mitochondria oxygen mass flux in the presence of different CNT types;
- (2)
- Transformation of CNT’s Raman spectra into SG spectral moments;
- (3)
- Calculation of the expected values of the mitochondria oxygen mass flux and the moving averages of the SG spectral moments under different experimental conditions;
- (4)
- Search for the best regression PTML models using the RRegrs package in R (https://github.com/enanomapper/RRegrs/).
4.2. Experimental Methods
4.2.1. General Procedures
4.2.2. Monitoring Mitochondrial Oxygen Mass
4.3. Computational Methods
4.3.1. Trace Invariants of Raman Spectra
4.3.2. PTML Model
4.3.3. Model Dataset
4.3.4. PTML Regression Predictors
- -
- NNreg function used a grid for 200, 300 and 400 neurons in the hidden layer (.size) and a decay of 0, 0.01, 0.2, 0.1 (.decay) (method = ‘nnet’);
- -
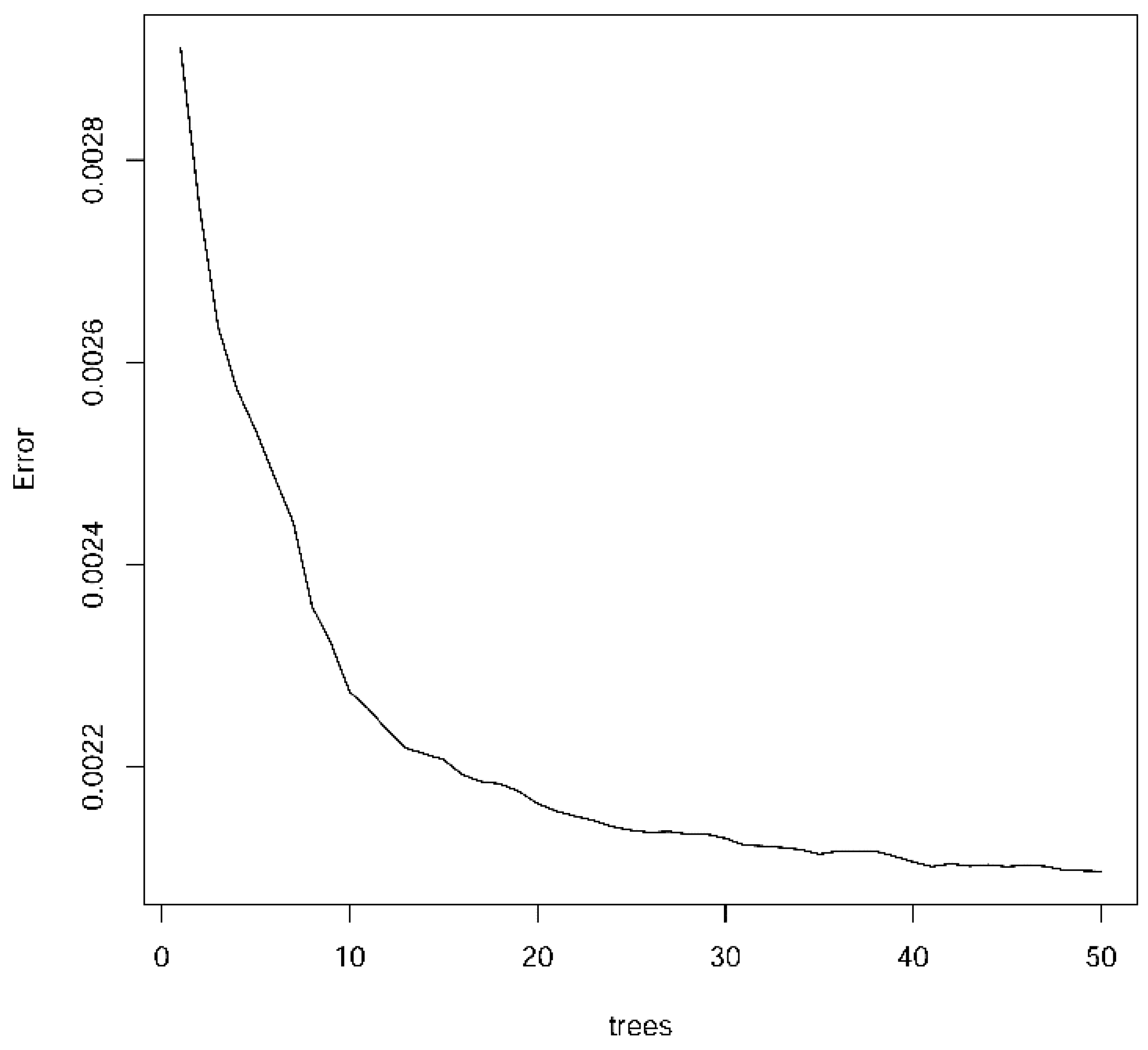
- RFreg function used 1500 trees (ntree = 1500 for method = ‘rf’).
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Prato, M.; Kostarelos, K.; Bianco, A. Functionalized carbon nanotubes in drug design and discovery. Acc. Chem. Res. 2008, 41, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Toropov, A.A.; Leszczynska, D.; Leszczynski, J. Predicting water solubility and octanol water partition coefficient for carbon nanotubes based on the chiral vector. Comput. Biol. Chem. 2007, 31, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.F.; Wu, Y.H.; Hou, Z.Q.; Zhang, Q.Q. Ros and nf-kappab are involved in upregulation of il-8 in a549 cells exposed to multi-walled carbon nanotubes. Biochem. Biophys. Res. Commun. 2009, 379, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Shvedova, A.A.; Castranova, V.; Kisin, E.R.; Schwegler-Berry, D.; Murray, A.R.; Gandelsman, V.Z.; Maynard, A.; Baron, P. Exposure to carbon nanotube material: Assessment of nanotube cytotoxicity using human keratinocyte cells. J. Toxicol. Environ. Health A 2003, 66, 1909–1926. [Google Scholar] [CrossRef] [PubMed]
- Bottini, M.; Bruckner, S.; Nika, K.; Bottini, N.; Bellucci, S.; Magrini, A.; Bergamaschi, A.; Mustelin, T. Multi-walled carbon nanotubes induce t lymphocyte apoptosis. Toxicol. Lett. 2006, 160, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Tian, F.; Ozkan, C.S.; Wang, M.; Gao, H. Effect of single wall carbon nanotubes on human HEK293 cells. Toxicol. Lett. 2005, 155, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.; Huaux, F.; Moreau, N.; Misson, P.; Heilier, J.F.; Delos, M.; Arras, M.; Fonseca, A.; Nagy, J.B.; Lison, D. Respiratory toxicity of multi-wall carbon nanotubes. Toxicol. Appl. Pharmacol. 2005, 207, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Heller, D.A.; Baik, S.; Eurell, T.E.; Strano, M.S. Single-walled carbon nanotube spectroscopy in live cells: Towards long-term labels and optical sensors. Adv. Mater. 2005, 17, 2793–2799. [Google Scholar] [CrossRef]
- Smart, S.K.; Cassady, A.I.; Lu, G.Q.; Martin, D.J. The biocompatibility of carbon nanotubes. Carbon 2006, 44, 1034–1047. [Google Scholar] [CrossRef]
- Toropov, A.A.; Toropova, A.P.; Pizzo, F.; Lombardo, A.; Gadaleta, D.; Benfenati, E. Coral: Model for no observed adverse effect level (noael). Mol. Divers. 2015, 19, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Urabe, H.; Sugawara, Y.; Ataka, M.; Rupprecht, A. Low-frequency raman spectra of lysozyme crystals and oriented DNA films: Dynamics of crystal water. Biophys. J. 1998, 74, 1533–1540. [Google Scholar] [CrossRef]
- Chou, K.C. Identification of low-frequency modes in protein molecules. Biochem. J. 1983, 215, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Piacenza, M.; Grimme, S. Van der waals interactions in aromatic systems: Structure and energetics of dimers and trimers of pyridine. ChemPhysChem 2005, 6, 1554–1558. [Google Scholar] [CrossRef] [PubMed]
- Hennrich, F.; Krupke, R.; Lebedkin, S.; Arnold, K.; Fischer, R.; Resasco, D.E.; Kappes, M.M. Raman spectroscopy of individual single-walled carbon nanotubes from various sources. J. Phys. Chem. B 2005, 109, 10567–10573. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhang, L.H.; Wang, L.R.; Xue, X.; Sun, J.H.; Wu, Y.; Zou, G.; Wu, X.; Wang, P.C.; Wamer, W.G.; et al. Single-walled carbon nanotubes alter cytochrome c electron transfer and modulate mitochondrial function. ACS Nano 2012, 6, 10486–10496. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, Y.; Yang, Y.; Sun, L.; Han, D.; Li, H.; Wang, C. Pharmacological and toxicological target organelles and safe use of single-walled carbon nanotubes as drug carriers in treating alzheimer disease. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Durruthy, M.; Alberici, L.C.; Curti, C.; Naal, Z.; Atique-Sawazaki, D.T.; Vazquez-Naya, J.M.; Gonzalez-Diaz, H.; Munteanu, C.R. Experimental-computational study of carbon nanotube effects on mitochondrial respiration: In silico nano-qspr machine learning models based on new raman spectra transform with markov-shannon entropy invariants. J. Chem. Inf. Model. 2017, 57, 1029–1044. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lozano, C.; Cuinas, R.F.; Seoane, J.A.; Fernandez-Blanco, E.; Dorado, J.; Munteanu, C.R. Classification of signaling proteins based on molecular star graph descriptors using machine learning models. J. Theor. Biol. 2015, 384, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lozano, C.; Gestal, M.; Pedreira-Souto, N.; Postelnicu, L.; Dorado, J.; Munteanu, C.R. Kernel-based feature selection techniques for transport proteins based on star graph topological indices. Curr. Top. Med. Chem. 2013, 13, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Blanco, E.; Aguiar-Pulido, V.; Munteanu, C.R.; Dorado, J. Random forest classification based on star graph topological indices for antioxidant proteins. J. Theor. Biol. 2012, 317, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Perez-Bello, A.; Munteanu, C.R.; Ubeira, F.M.; De Magalhaes, A.L.; Uriarte, E.; Gonzalez-Diaz, H. Alignment-free prediction of mycobacterial DNA promoters based on pseudo-folding lattice network or star-graph topological indices. J. Theor. Biol. 2009, 256, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.M.; Aguiar, V.; Seoane, J.A.; Freire, A.; Serantes, J.A.; Dorado, J.; Pazos, A.; Munteanu, C.R. Star graphs of protein sequences and proteome mass spectra in cancer prediction. Curr. Proteom. 2009, 6, 275–288. [Google Scholar] [CrossRef]
- Fernandez-Blanco, E.; Rivero, D.; Rabunal, J.; Dorado, J.; Pazos, A.; Munteanu, C.R. Automatic seizure detection based on star graph topological indices. J. Neurosci. Methods 2012, 209, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Guo, J.; Chen, T.; Nie, H.; Wang, H.; Zang, J.; Cui, X.; Jia, G. Multi-walled carbon nanotubes induce apoptosis via mitochondrial pathway and scavenger receptor. Toxicol. In Vitro 2012, 26, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Knobeloch, L.M.; Blondin, G.A.; Harkin, J.M. Use of submitochondrial particles for prediction of chemical toxicity in man. Bull. Environ. Contam. Toxicol. 1990, 44, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Horan, M.P.; Pichaud, N.; Ballard, J.W. Review: Quantifying mitochondrial dysfunction in complex diseases of aging. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 1022–1035. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Diaz, H.; Arrasate, S.; Juan, A.G.; Sotomayor, N.; Lete, E.; Speck-Planche, A.; Ruso, J.M.; Luan, F.; Cordeiro, M.N. Matrix trace operators: From spectral moments of molecular graphs and complex networks to perturbations in synthetic reactions, micelle nanoparticles, and drug adme processes. Curr. Drug Metab. 2014, 15, 470–488. [Google Scholar] [CrossRef] [PubMed]
- Vergara-Galicia, J.; Prado-Prado, F.J.; Gonzalez-Diaz, H. Galvez-markov network transferability indices: Review of classic theory and new model for perturbations in metabolic reactions. Curr. Drug Metab. 2014, 15, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Diaz, H.; Perez-Montoto, L.G.; Ubeira, F.M. Model for vaccine design by prediction of B-epitopes of iedb given perturbations in peptide sequence, in vivo process, experimental techniques, and source or host organisms. J. Immunol. Res. 2014, 2014, 768515. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Diaz, H.; Arrasate, S.; Gomez-SanJuan, A.; Sotomayor, N.; Lete, E.; Besada-Porto, L.; Ruso, J.M. General theory for multiple input-output perturbations in complex molecular systems. 1. Linear qspr electronegativity models in physical, organic, and medicinal chemistry. Curr. Top. Med. Chem. 2013, 13, 1713–1741. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, C.R. Regression Models to Predict Mitochondria Oxygen Mass Flux in the preseNce of the Cnts. Figshare, 2016. Available online: Https://dx.Doi.Org/10.6084/m9.Figshare.3545561 (accessed on 7 August 2016).
- Martínez, A.; Francisco-Marquez, M.; Galano, A. Effect of different functional groups on the free radical scavenging capability of single-walled carbon nanotubes. J. Phys. Chem. C 2010, 114, 14734–14739. [Google Scholar] [CrossRef]
- Rivera Gil, P.; Oberdorster, G.; Elder, A.; Puntes, V.; Parak, W.J. Correlating physico-chemical with toxicological properties of nanoparticles: The present and the future. ACS Nano 2010, 4, 5527–5531. [Google Scholar] [CrossRef] [PubMed]
- Hinderliter, P.M.; Minard, K.R.; Orr, G.; Chrisler, W.B.; Thrall, B.D.; Pounds, J.G.; Teeguarden, J.G. Isdd: A computational model of particle sedimentation, diffusion and target cell dosimetry for in vitro toxicity studies. Part. Fibre Toxicol. 2010, 7, 36. [Google Scholar] [CrossRef] [PubMed]
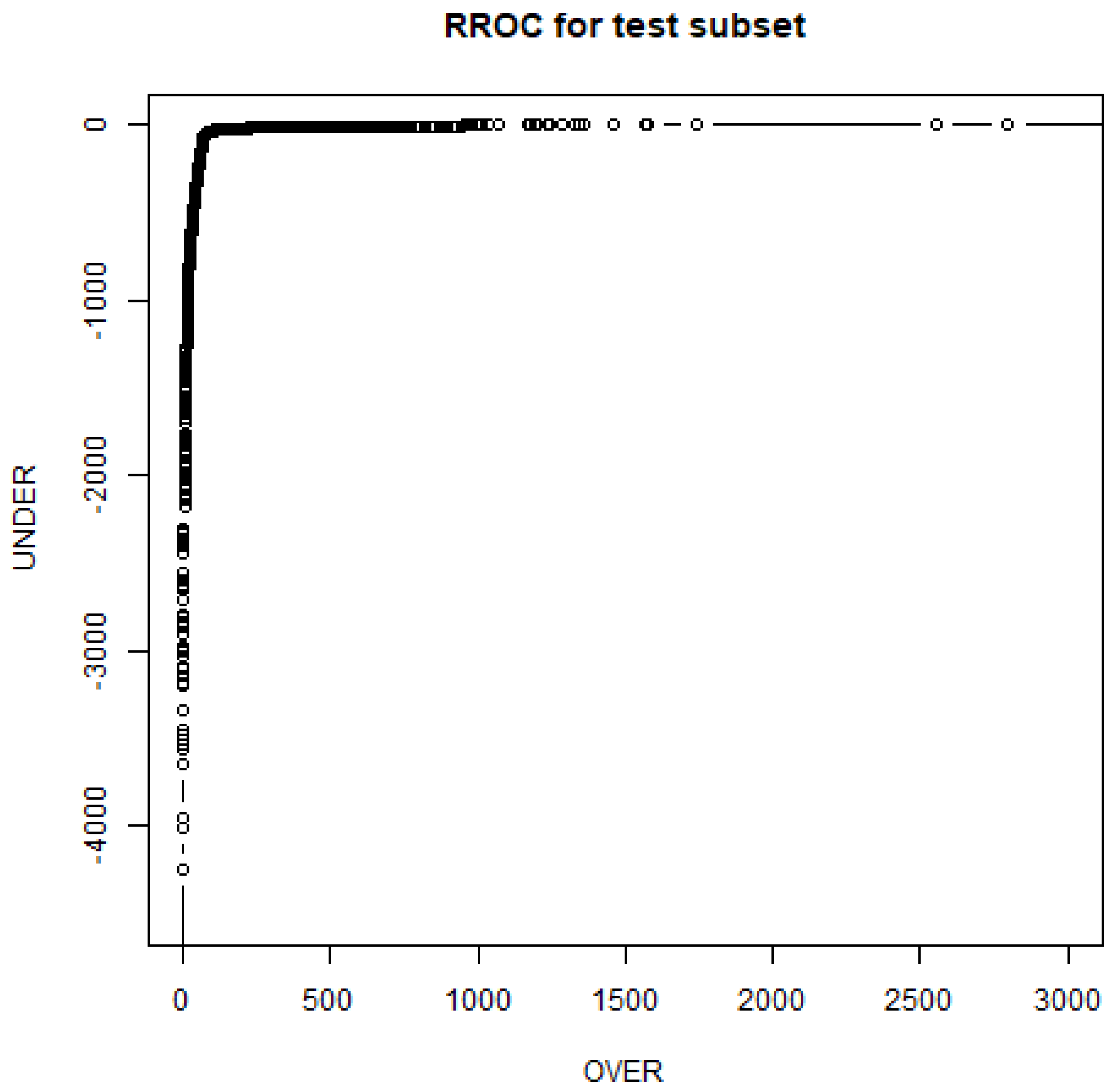
- Hernández-Orallo, J. Roc curves for regression. Pattern Recogn. 2013, 46, 3395–3411. [Google Scholar] [CrossRef]
- Pedersen, P.L.; Greenawalt, J.W.; Reynafarje, B.; Hullihen, J.; Decker, G.L.; Soper, J.W.; Bustamente, E. Preparation and characterization of mitochondria and submitochondrial particles of rat liver and liver-derived tissues. Methods Cell Biol. 1978, 20, 411–481. [Google Scholar] [PubMed]
- Lanza, I.R.; Nair, K.S. Functional assessment of isolated mitochondria in vitro. Methods Enzymol. 2009, 457, 349–372. [Google Scholar] [PubMed]
- Gnaiger, E. Bioenergetics at low oxygen: Dependence of respiration and phosphorylation on oxygen and adenosine diphosphate supply. Respir. Physiol. 2001, 128, 277–297. [Google Scholar] [CrossRef]
- Munteanu, C.R.; Magalhaes, A.L.; Duardo-Sanchez, A.; Pazos, A.; Gonzalez-Diaz, H. S2Snet: A tool for transforming characters and numeric sequences into star network topological indices in chemoinformatics, bioinformatics, biomedical, and social-legal sciences. Curr. Bioinf. 2013, 8, 429–437. [Google Scholar] [CrossRef]
- Munteanu, C.R. Cnt Raman for Mitocondrial Oxygen Consumption Data Set and Regression Models. Figshare. Available online: Https://dx.Doi.Org/10.6084/m9.Figshare.3472349 (accessed on 31 January 2017).
- Tsiliki, G.; Munteanu, C.R.; Seoane, J.A.; Fernandez-Lozano, C.; Sarimveis, H.; Willighagen, E.L. Rregrs: An r package for computer-aided model selection with multiple regression models. J. Cheminform. 2015, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Tsiliki, G.; Munteanu, C.R.; Seoane, J.; Fernandez-Lozano, C.; Sarimveis, H.; Willighagen, E. Using the rregrs R package for automating predictive modelling. In Proceedings of the MOL2NET International Conference on Multidisciplinary Sciences, Bizkaia, Spain, 5–15 December 2015; Volume 1, p. F009. [Google Scholar]
- Bishop, C.M. Neural Networks for Pattern Recognition; Oxford University Press: New York, NY, USA, 1995. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regression Method | Statistics | Training | Test | ||
|---|---|---|---|---|---|
| R2tr | RMSEtr | R2ts | RMSEts | ||
| Linear Multi-regression (LM) | Mean | 0.358 | 0.0959 | 0.356 | 0.0954 |
| Min | 0.349 | 0.0954 | 0.340 | 0.0932 | |
| Max | 0.363 | 0.0966 | 0.384 | 0.0969 | |
| Neural Network (NN) | Mean | 0.645 | 0.0709 | 0.672 | 0.0681 |
| Min | 0.626 | 0.0697 | 0.620 | 0.0613 | |
| Max | 0.659 | 0.0727 | 0.739 | 0.0738 | |
| Random Forest (RF) | Mean | 0.855 | 0.0455 | 0.856 | 0.0452 |
| Min | 0.851 | 0.0451 | 0.853 | 0.0431 | |
| Max | 0.858 | 0.0462 | 0.863 | 0.0461 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Durruthy, M.; Monserrat, J.M.; Rasulev, B.; Casañola-Martín, G.M.; Barreiro Sorrivas, J.M.; Paraíso-Medina, S.; Maojo, V.; González-Díaz, H.; Pazos, A.; Munteanu, C.R. Carbon Nanotubes’ Effect on Mitochondrial Oxygen Flux Dynamics: Polarography Experimental Study and Machine Learning Models using Star Graph Trace Invariants of Raman Spectra. Nanomaterials 2017, 7, 386. https://doi.org/10.3390/nano7110386
González-Durruthy M, Monserrat JM, Rasulev B, Casañola-Martín GM, Barreiro Sorrivas JM, Paraíso-Medina S, Maojo V, González-Díaz H, Pazos A, Munteanu CR. Carbon Nanotubes’ Effect on Mitochondrial Oxygen Flux Dynamics: Polarography Experimental Study and Machine Learning Models using Star Graph Trace Invariants of Raman Spectra. Nanomaterials. 2017; 7(11):386. https://doi.org/10.3390/nano7110386
Chicago/Turabian StyleGonzález-Durruthy, Michael, Jose M. Monserrat, Bakhtiyor Rasulev, Gerardo M. Casañola-Martín, José María Barreiro Sorrivas, Sergio Paraíso-Medina, Víctor Maojo, Humberto González-Díaz, Alejandro Pazos, and Cristian R. Munteanu. 2017. "Carbon Nanotubes’ Effect on Mitochondrial Oxygen Flux Dynamics: Polarography Experimental Study and Machine Learning Models using Star Graph Trace Invariants of Raman Spectra" Nanomaterials 7, no. 11: 386. https://doi.org/10.3390/nano7110386
APA StyleGonzález-Durruthy, M., Monserrat, J. M., Rasulev, B., Casañola-Martín, G. M., Barreiro Sorrivas, J. M., Paraíso-Medina, S., Maojo, V., González-Díaz, H., Pazos, A., & Munteanu, C. R. (2017). Carbon Nanotubes’ Effect on Mitochondrial Oxygen Flux Dynamics: Polarography Experimental Study and Machine Learning Models using Star Graph Trace Invariants of Raman Spectra. Nanomaterials, 7(11), 386. https://doi.org/10.3390/nano7110386