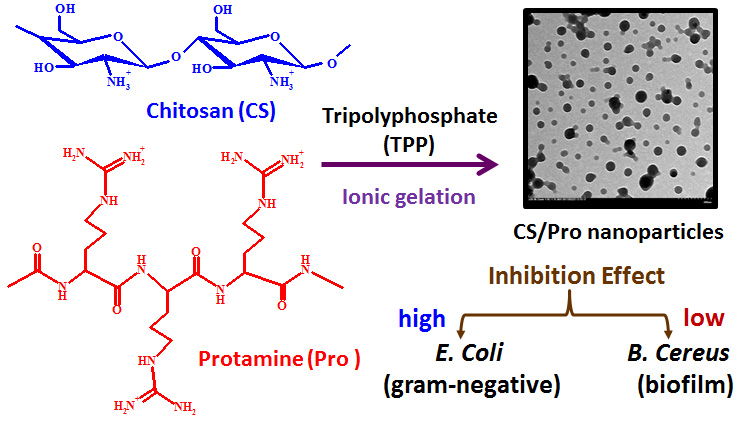
Antibacterial Effects of Chitosan/Cationic Peptide Nanoparticles

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Characterzation of Chitosan and Protamine
2.2. Chemical and Physical Properties of Nanoparticles
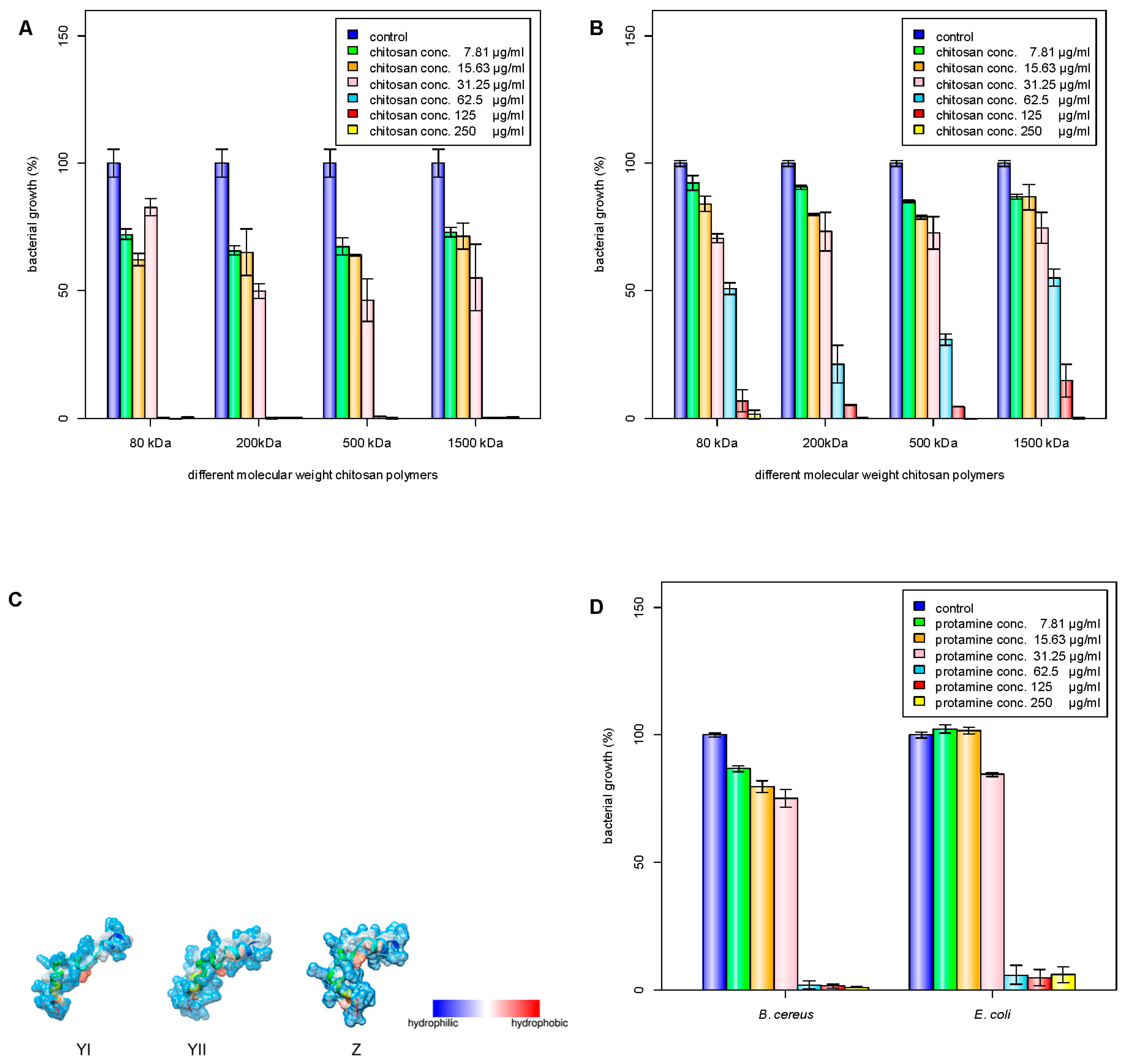
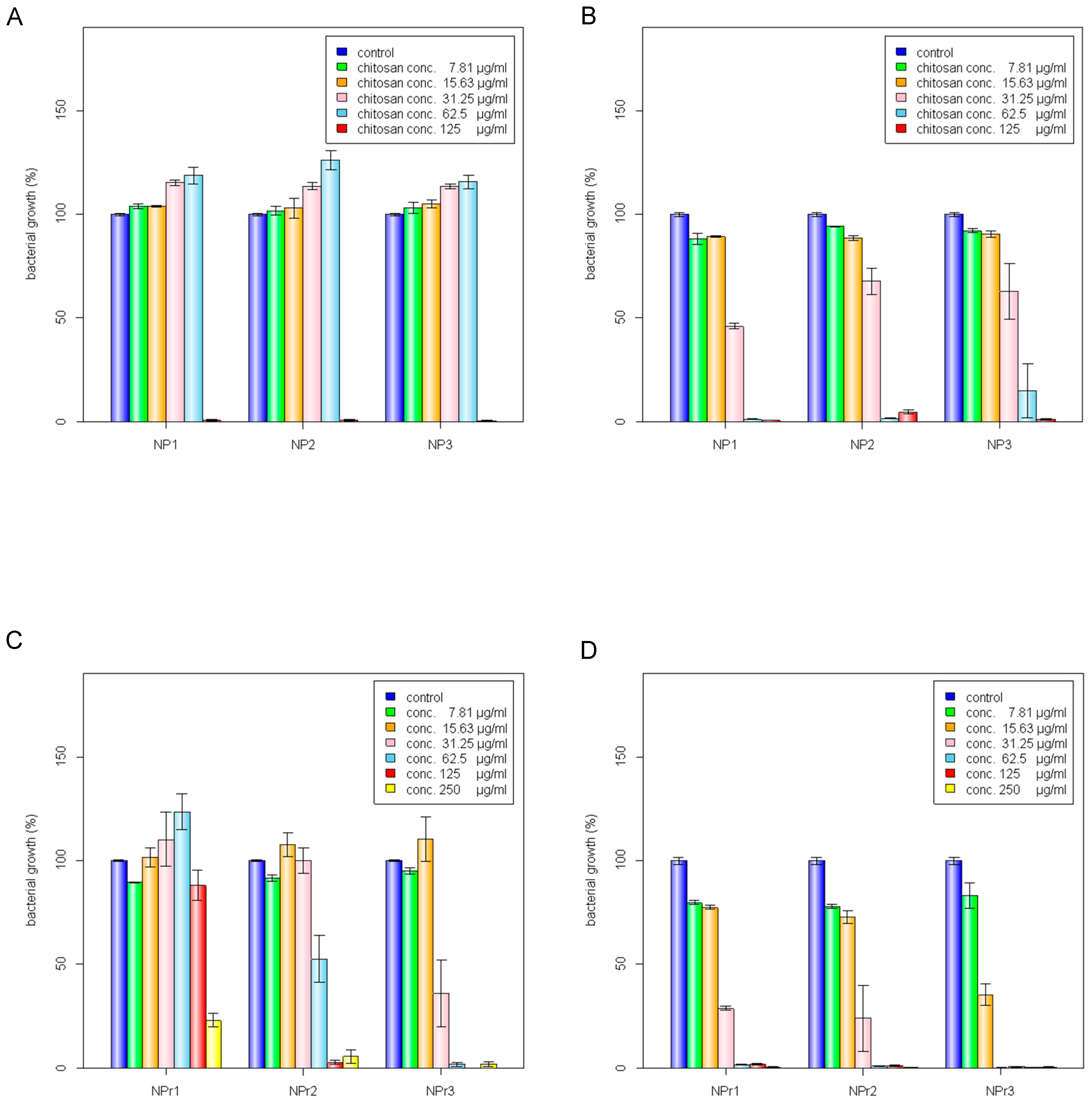
2.3. Antibacterial Effects of Chitosan and Chitosan/Protamine Nanoparticles
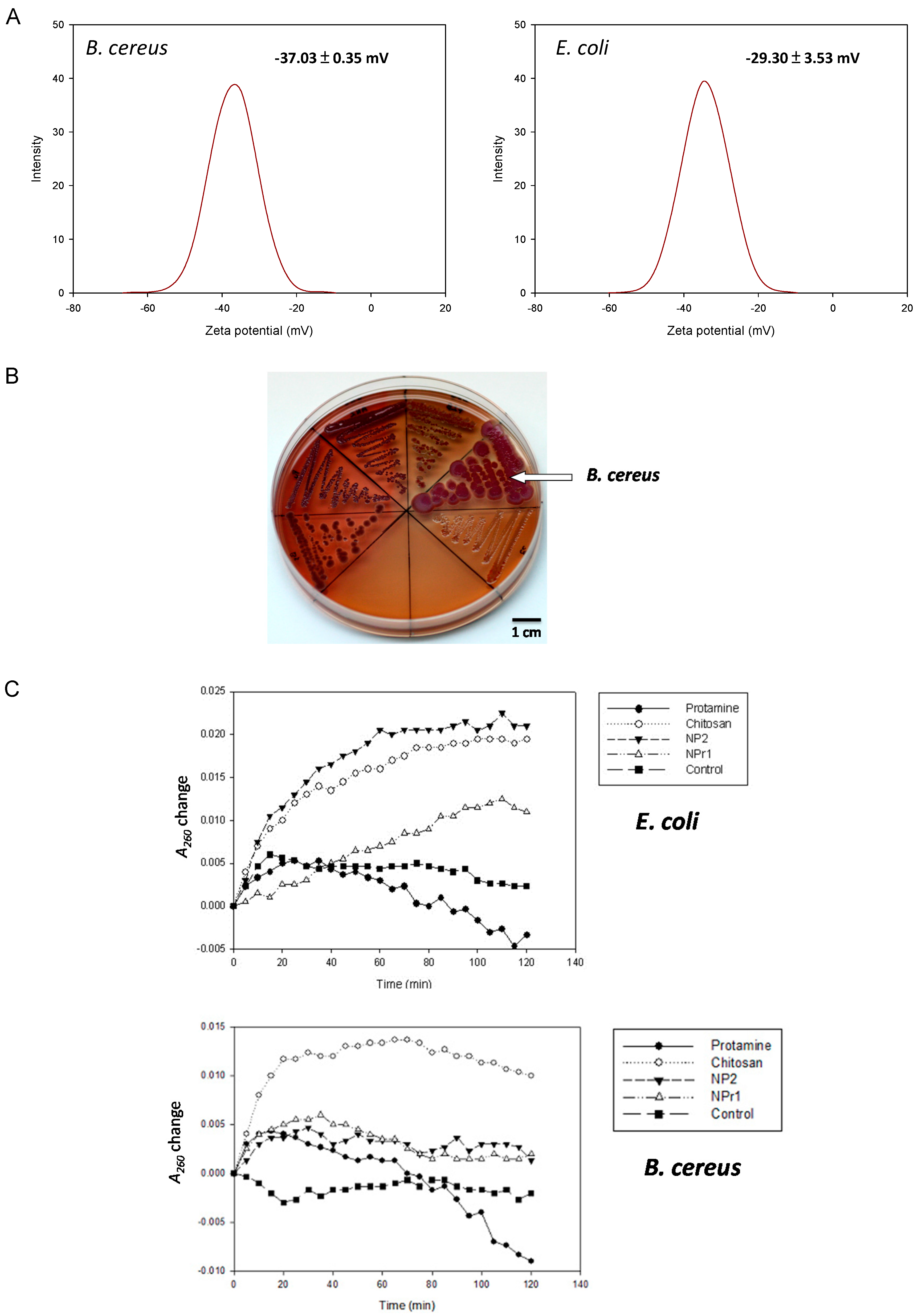
2.4. Surface Charge and Hydrophobicity
2.5. Formation of Biofilm-Like Structure
2.6. Production of Extra Polysaccharides
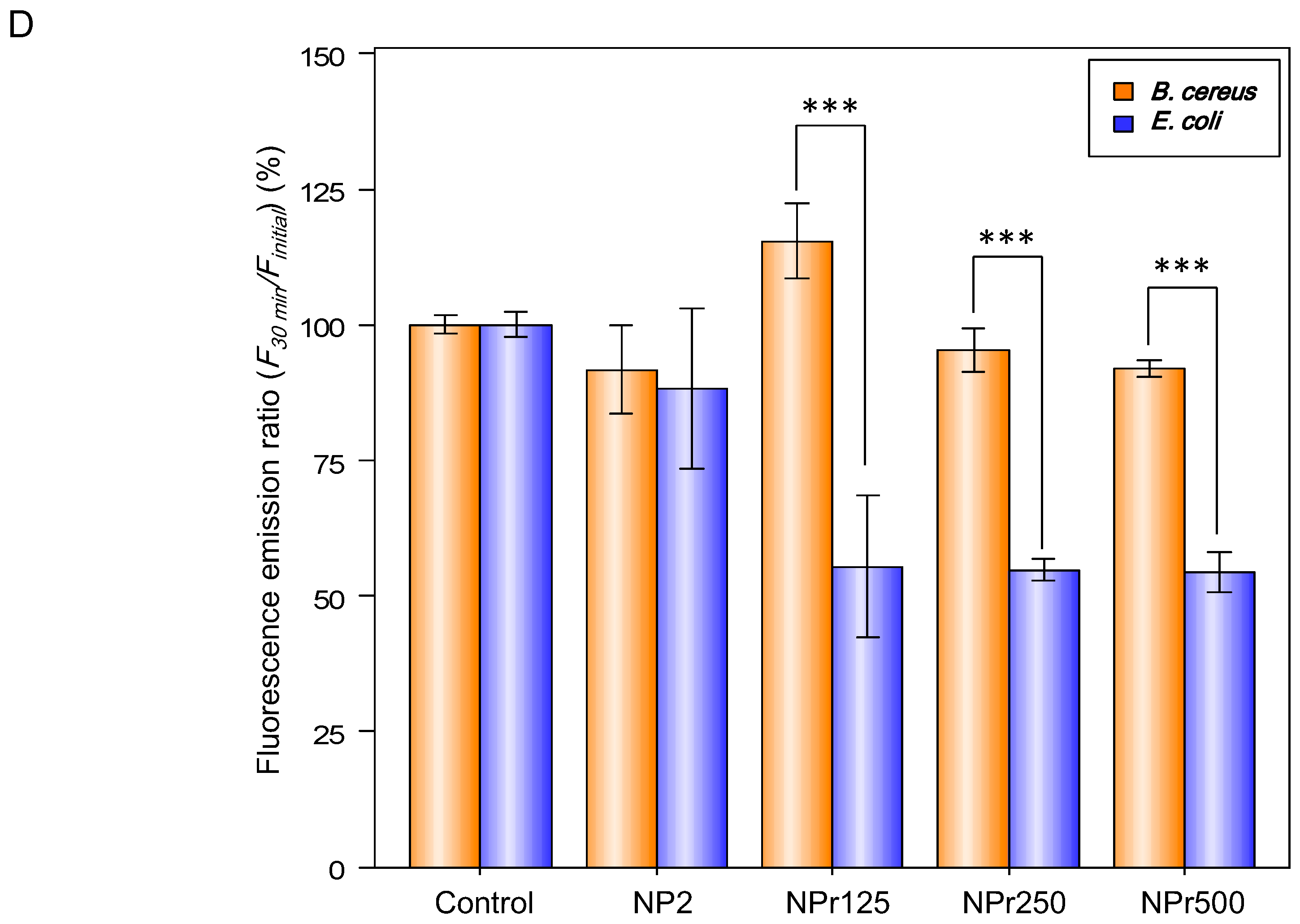
2.7. Membrane Permeability
3. Experimental Section
3.1. Materials
3.2. Preparation of Chitosan-TPP Nanoparticles (NP)
3.3. Preparation of Chitosan/Protamine Nanoparticles (NPrs)
3.4. Chemical and Physical Properties of Nanoparticles
3.5. Measurement of Bacterial Zeta Potential
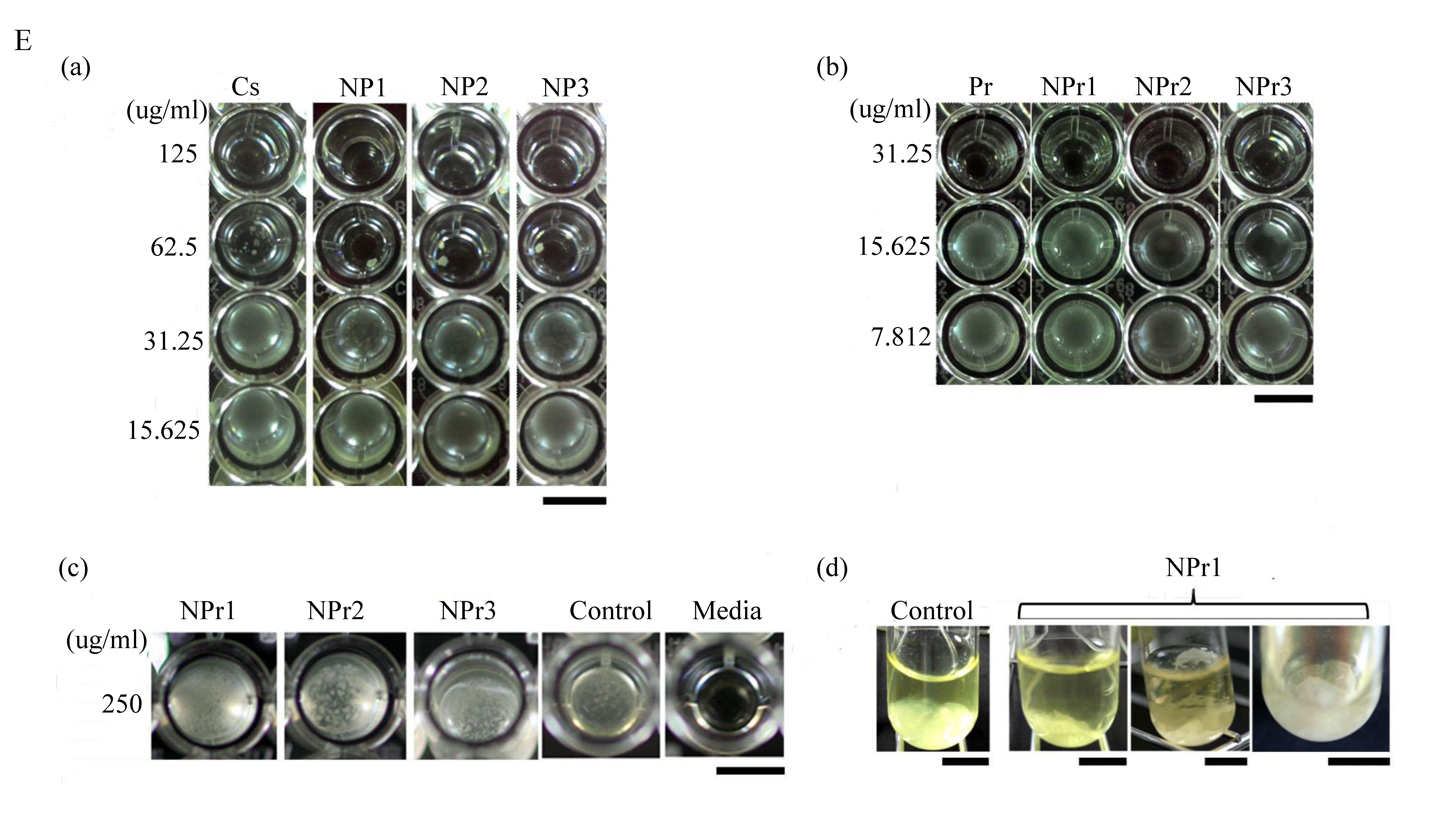
3.6. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
3.7. Measurement of the Hydrophobicity of Cell Surface
3.8. Detection of Membrane Integrity
3.9. Detection of Polysaccharide Formation
3.10. Detection of Biofilm-Like Structure Formation
3.11. In Silico Analysis of Protamine
3.12. Fluorescence Assay for Nanoparticles Binding to Bacteria
3.13. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Watkins, R.R.; Bonomo, R.A. Overview: Global and local impact of antibiotic resistance. Infect. Dis. Clin. N. Am. 2016, 30, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zuo, J.H.; Wang, Q.; Na, Y.; Gao, L.P. Inhibitory effect of chitosan on growth of the fungal phytopathogen, Sclerotinia sclerotiorum, and sclerotinia rot of carrot. J. Integr. Agric. 2015, 14, 691–697. [Google Scholar] [CrossRef]
- Mellegård, H.; Strand, S.P.; Christensen, B.E.; Granum, P.E.; Hardy, S.P. Antibacterial activity of chemically defined chitosans: Influence of molecular weight, degree of acetylation and test organism. Int. J. Food Microbiol. 2011, 148, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, S.; Mahapatra, S.K.; Chattopadhyay, S.; Das, S.; Dash, S.K.; Majumder, S.; Pramanik, P.; Roy, S. A novel chitosan based antimalarial drug delivery against Plasmodium berghei infection. Acta Trop. 2013, 128, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Sathiyabama, M.; Parthasarathy, R. Biological preparation of chitosan nanoparticles and its in vitro antifungal efficacy against some phytopathogenic fungi. Carbohydr. Polym. 2016, 151, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Ai, H.; Wang, F.R.; Xia, Y.Q.; Chen, X.M.; Lei, C.L. Antioxidant, antifungal and antiviral activities of chitosan from the larvae of housefly, Musca domestica L. Food Chem. 2012, 132, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.H.; Lin, H.T.V.; Wu, G.J.; Tsai, G.J. pH Effects on solubility, zeta potential, and correlation between antibacterial activity and molecular weight of chitosan. Carbohydr. Polym. 2015, 134, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Sahariah, P.; Oskarsson, B.M.; Hjalmarsdottir, M.A.; Masson, M. Synthesis of guanidinylated chitosan with the aid of multiple protecting groups and investigation of antibacterial activity. Carbohydr. Polym. 2015, 127, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.X.; Li, J.N.; Li, Q.Q.; Shen, Y.Y.; Ge, Z.C.; Zhang, W.W.; Chen, S.G. Enhanced water-solubility, antibacterial activity and biocompatibility upon introducing sulfobetaine and quaternary ammonium to chitosan. Carbohydr. Polym. 2016, 143, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Wu, C.; Fu, S.; Wang, L.; Yuan, C.; Chen, S.; Hu, Y. Integration of lysozyme into chitosan nanoparticles for improving antibacterial activity. Carbohydr. Polym. 2017, 155, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.T.; Austin, J.W.; Gill, T.A. Antibacterial effect of protamine in combination with EDTA and refrigeration. Int. J. Food Microbiol. 2001, 66, 149–161. [Google Scholar] [CrossRef]
- Potter, R.; Hansen, L.T.; Gill, T.A. Inhibition of foodborne bacteria by native and modified protamine: Importance of electrostatic interactions. Int. J. Food Microbiol. 2005, 103, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Lesmes, L.P.; Bohorquez, M.Y.; Carreno, L.F.; Patarroyo, M.E.; Lozano, J.M. A C-terminal cationic fragment derived from an arginine-rich peptide exhibits in vitro antibacterial and anti-plasmodial activities governed by its secondary structure properties. Peptides 2009, 30, 2150–2160. [Google Scholar] [CrossRef] [PubMed]
- Darouiche, R.O.; Mansouri, M.D.; Gawande, P.V.; Madhyastha, S. Efficacy of combination of chlorhexidine and protamine sulphate against device-associated pathogens. J. Antimicrob. Chemother. 2008, 61, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Aspedon, A.; Groisman, E.A. The antibacterial action of protamine: Evidence for disruption of cytoplasmic membrane energization in Salmonella typhimurium. Microbiology-UK 1996, 142, 3389–3397. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.; Verheul, A.; Gram, L.; Gill, T.; Abee, T. Protamine-induced permeabilization of cell envelopes of gram-positive and gram-negative bacteria. Appl. Environ. Microbiol. 1997, 63, 1155–1159. [Google Scholar] [PubMed]
- Willcox, M.D.P.; Hume, E.B.H.; Aliwarga, Y.; Kumar, N.; Cole, N. A novel cationic-peptide coating for the prevention of microbial colonization on contact lenses. J. Appl. Microbiol. 2008, 105, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Burton, E.; Gawande, P.V.; Yakandawala, N.; LoVetri, K.; Zhanel, G.G.; Romeo, T.; Friesen, A.D.; Madhyastha, S. Antibiofilm activity of GlmU enzyme inhibitors against catheter-associated uropathogens. Antimicrob. Agents Chemother. 2006, 50, 1835–1840. [Google Scholar] [CrossRef] [PubMed]
- Ki, M.H.; Kim, J.E.; Lee, Y.N.; Noh, S.M.; An, S.W.; Cho, H.J.; Kim, D.D. Chitosan-based hybrid nanocomplex for siRNA delivery and its application for cancer therapy. Pharm. Res. 2014, 31, 3323–3334. [Google Scholar] [CrossRef] [PubMed]
- Moran, M.C.; Jorge, A.F.; Vinardell, M.P. Sustainable DNA release from chitosan/protein based-DNA gel particles. Biomacromolecules 2014, 15, 3953–3964. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.Y.; He, H.N.; Han, L.M.; Qin, J.; Chen, S.H.; Ru, G.; Li, R.X.; Yang, P.; Wang, J.X.; Yang, V.C. Enhancing insulin oral absorption by using mucoadhesive nanoparticles loaded with LMWP-linked insulin conjugates. J. Control. Release 2016, 233, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Hosseinnejad, M.; Jafari, S.M. Evaluation of different factors affecting antimicrobial properties of chitosan. Int. J. Biol. Macromol. 2016, 85, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Goy, R.C.; de Britto, D.; Assis, O.B.G. A review of the antimicrobial activity of chitosan. Polim.-Cienc. E Tecnol. 2009, 19, 241–247. [Google Scholar] [CrossRef]
- Li, J.H.; Wu, Y.G.; Zhao, L.Q. Antibacterial activity and mechanism of chitosan with ultra high molecular weight. Carbohydr. Polym. 2016, 148, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Younes, I.; Sellimi, S.; Rinaudo, M.; Jellouli, K.; Nasri, M. Influence of acetylation degree and molecular weight of homogeneous chitosans on antibacterial and antifungal activities. Int. J. Food Microbiol. 2014, 185, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F.; Feng, X.Q.; Yang, S.; Fu, G.Q.; Wang, T.P.; Su, Z.X. Chitosan kills Escherichia coli through damage to be of cell membrane mechanism. Carbohydr. Polym. 2010, 79, 493–499. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Pink, D.A.; Hansen, L.T.; Gill, T.A.; Quinn, B.E.; Jericho, M.H.; Beveridge, T.J. Divalent calcium ions inhibit the penetration of protamine through the polysaccharide brush of the outer membrane of Gram-negative bacteria. Langmuir 2003, 19, 8852–8858. [Google Scholar] [CrossRef]
- Hansen, L.T.; Gill, T.A. Solubility and antimicrobial efficacy of protamine on Listeria monocytogenes and Escherichia coli as influenced by pH. J. Appl. Microbiol. 2000, 88, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Pink, D.A.; Hasan, F.M.; Quinn, B.E.; Winterhalter, M.; Mohan, M.; Gill, T.A. Interaction of protamine with gram-negative bacteria membranes: Possible alternative mechanisms of internalization in Escherichia coli, Salmonella typhimurium and Pseudomonas aeruginosa. J. Pept. Sci. 2014, 20, 240–250. [Google Scholar] [CrossRef] [PubMed]
- McDougald, D.; Rice, S.A.; Barraud, N.; Steinberg, P.D.; Kjelleberg, S. Should we stay or should we go: Mechanisms and ecological consequences for biofilm dispersal. Nat. Rev. Microbiol. 2012, 10, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Czaczyk, K.; Myszka, K. Biosynthesis of extracellular polymeric substances (EPS) and its role in microbial biofilm formation. Pol. J. Environ. Stud. 2007, 16, 799–806. [Google Scholar]
- Stewart, P.S. Mechanisms of antibiotic resistance in bacterial biofilms. Int. J. Med. Microbiol. 2002, 292, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Z.S.; Cooper, S.L. Interactions between dendrimer biocides and bacterial membranes. Biomaterials 2002, 23, 3359–3368. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Araújo, A.M.M.; de Olivera, I.C.M.; de Mattos, M.C.; Benchetrit, L.C. Cell surface hydrophobicity and adherence of a strain of group B streptococci during the post-antibiotic effect of penicillin. Revista do Instituto de Medicina Tropical de São Paulo 2008, 50, 203–207. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chitosan | MIC (µg/mL) | MBC (µg/mL) | |||
|---|---|---|---|---|---|
| E. coli | B. cereus | E. coli | B. cereus | ||
| M1 (80 kDa) | 125 | 62.5–125 | 250 | 62.5–125 | |
| M2 (200 kDa) | 62.5–125 | 62.5–125 | 125 | 62.5–125 | |
| M3 (500 kDa) | 125 | 62.5–125 | 125–250 | 62.5–125 | |
| M4 (1500 kDa) | 125–250 | 62.5–125 | 125–250 | 62.5–125 | |
| Type of Nanoparticles | Size (nm) | Zeta Potential (mV) |
|---|---|---|
| CS NPs | ||
| NP1 | 78.4 ± 4.01 | 33.77 ± 1.30 |
| NP2 | 150.67 ± 3.05 | 33.63 ± 0.32 |
| NP3 | 201 ± 3.60 | 32 ± 1.11 |
| CS/Protamine NPs | ||
| NPr1 | 114.33 ± 4.16 | 32.23 ± 0.76 |
| NPr2 | 84.8 ± 2.07 | 30.27 ± 0.72 |
| NPr3 | 79.4 ± 1.90 | 27.67 ± 1.45 |
| Antimicrobials | MIC (µg/mL) | MBC (µg/mL) | ||
|---|---|---|---|---|
| E. coli | B. cereus | E. coli | B. cereus | |
| Cs only (200 kDa) | 62.5–125 | 62.5–125 | 125 | 62.5–125 |
| NP1 | 31.25–125 | 125 | 125 | ≥125 |
| NP2 | 31.25–125 | 125 | >250 | ≥250 |
| NP3 | 31.25–125 | 125 | 125 | ≥250 |
| Protamine | 31.25–62.5 | 62.5 | 31.25–62.5 | 125 |
| NPr1 | 31.25 | >250 | 31.25–62.5 | >250 |
| NPr2 | 31.25 | >250 | 31.25–62.5 | >250 |
| NPr3 | 31.25 | 31.25 | 31.25–62.5 | >250 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamara, F.R.; Lin, C.; Mi, F.-L.; Ho, Y.-C. Antibacterial Effects of Chitosan/Cationic Peptide Nanoparticles. Nanomaterials 2018, 8, 88. https://doi.org/10.3390/nano8020088
Tamara FR, Lin C, Mi F-L, Ho Y-C. Antibacterial Effects of Chitosan/Cationic Peptide Nanoparticles. Nanomaterials. 2018; 8(2):88. https://doi.org/10.3390/nano8020088
Chicago/Turabian StyleTamara, Frans Ricardo, Chi Lin, Fwu-Long Mi, and Yi-Cheng Ho. 2018. "Antibacterial Effects of Chitosan/Cationic Peptide Nanoparticles" Nanomaterials 8, no. 2: 88. https://doi.org/10.3390/nano8020088
APA StyleTamara, F. R., Lin, C., Mi, F. -L., & Ho, Y. -C. (2018). Antibacterial Effects of Chitosan/Cationic Peptide Nanoparticles. Nanomaterials, 8(2), 88. https://doi.org/10.3390/nano8020088