BODIPY Dyes as Probes and Sensors to Study Amyloid-β-Related Processes

Abstract
:1. Introduction
2. Amyloid-β and Alzheimer’s Disease
General Comments on the Preparation of Aβ Monomers, Oligomers, and Fibrils
- Aβ monomers
- Aβ oligomers
- Aβ fibrils
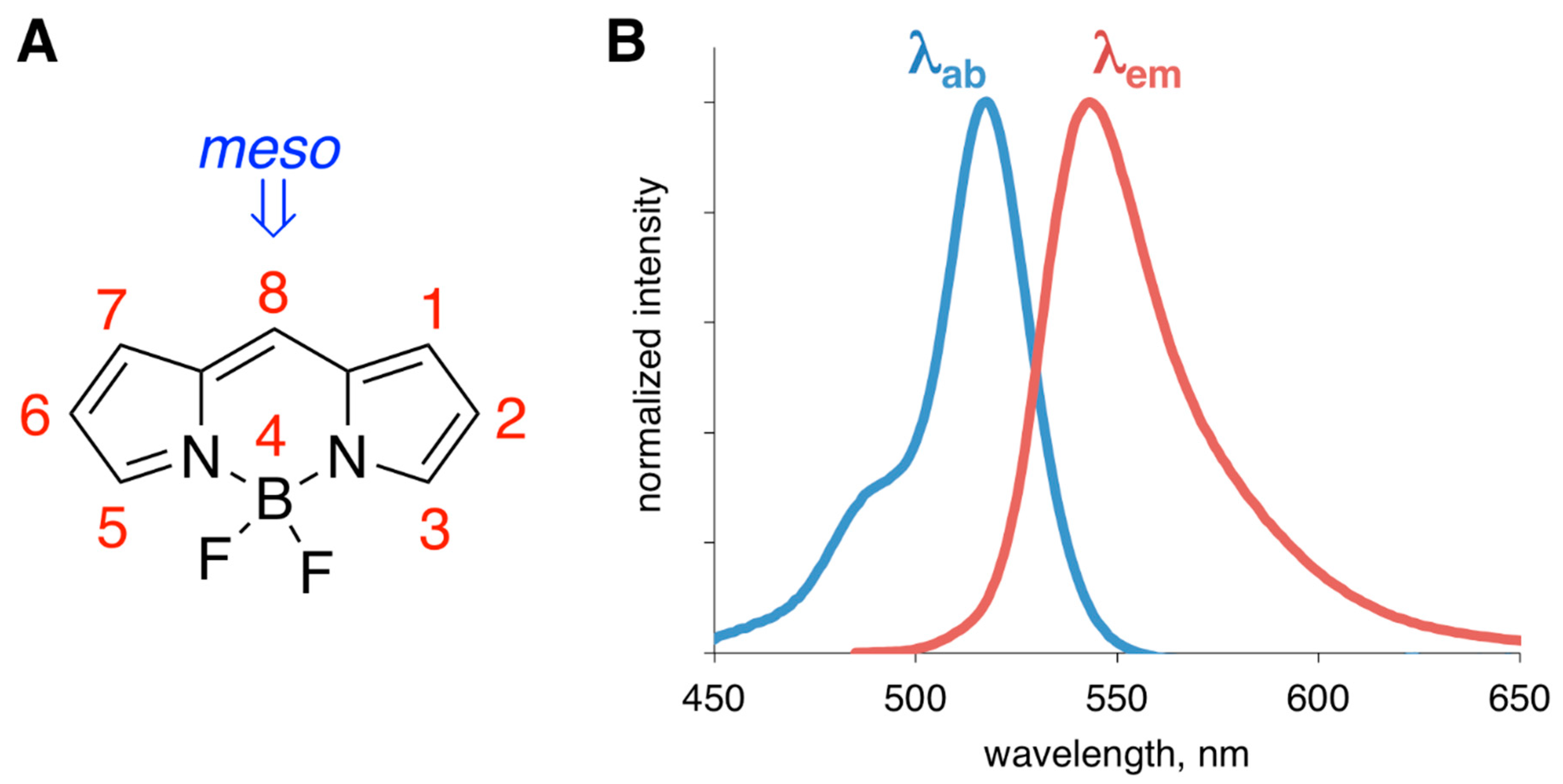
3. BODIPY-Based Sensors for Monitoring Aggregation and Conformational Changes of Amyloids
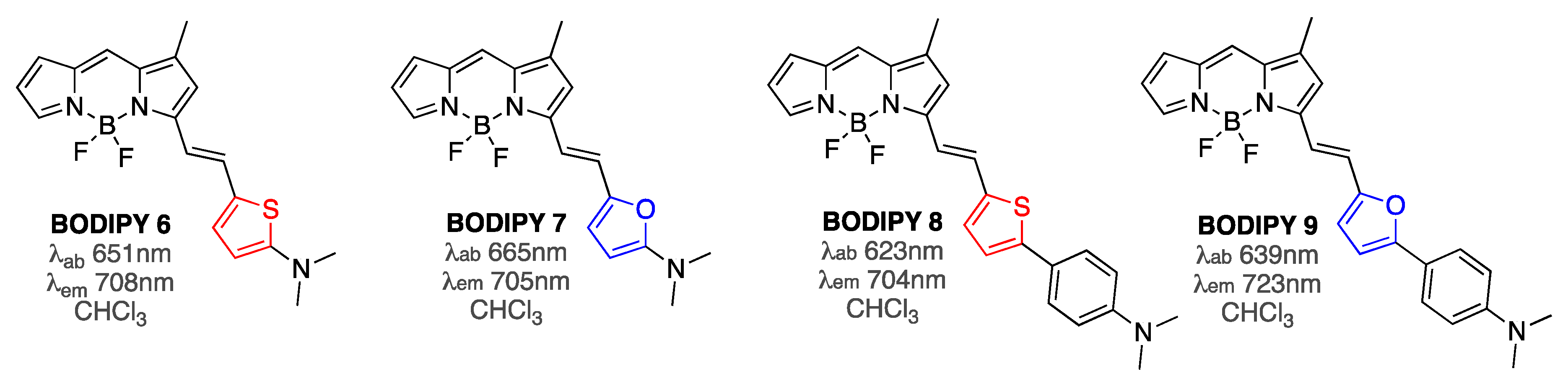
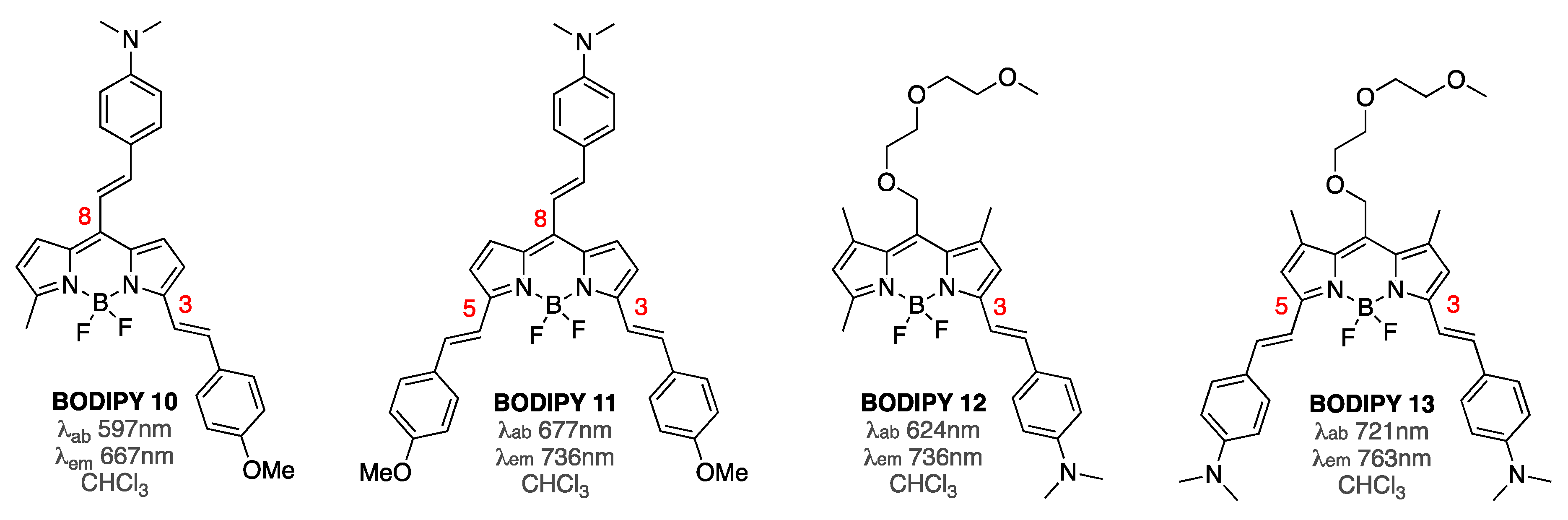
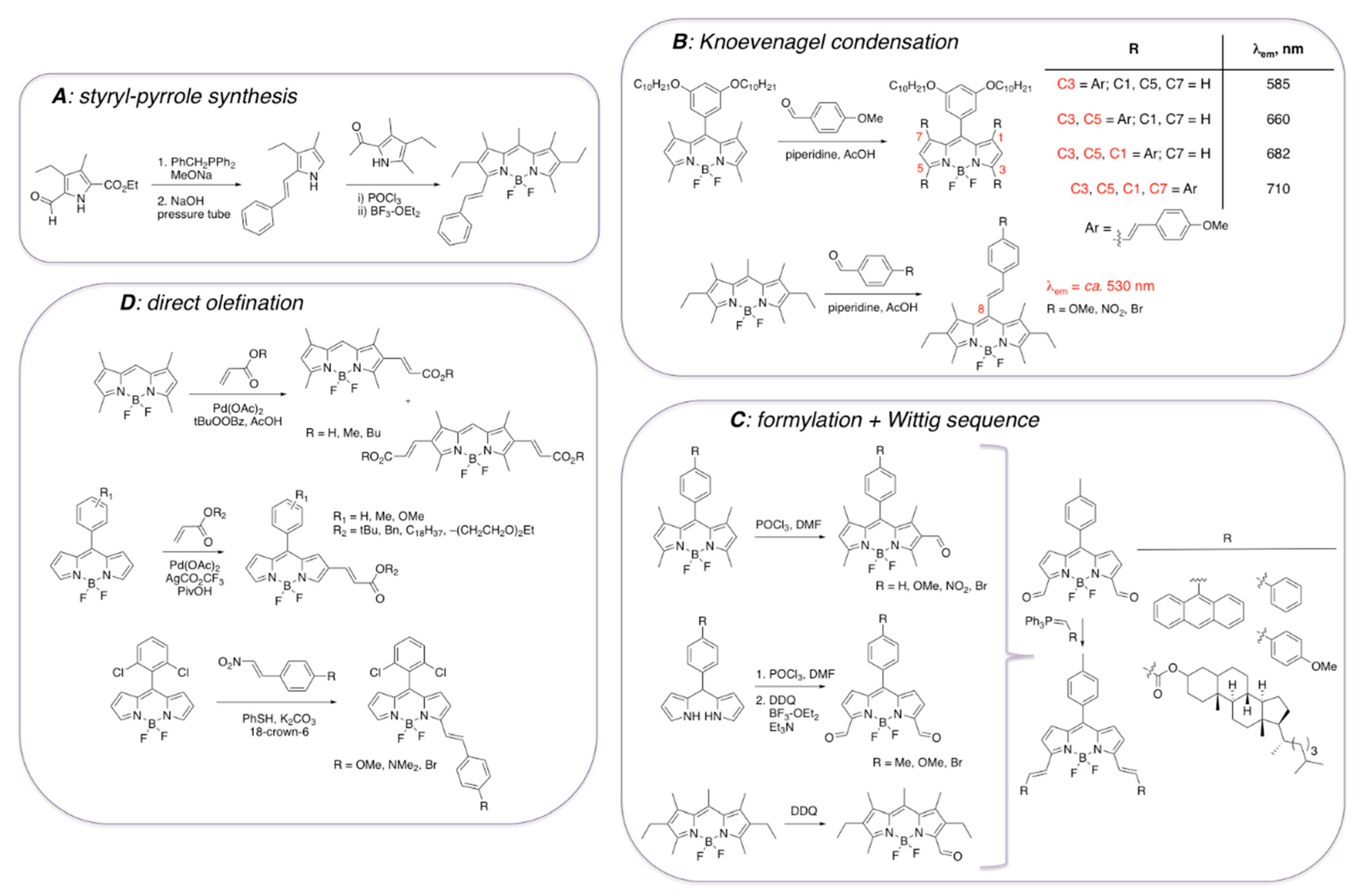
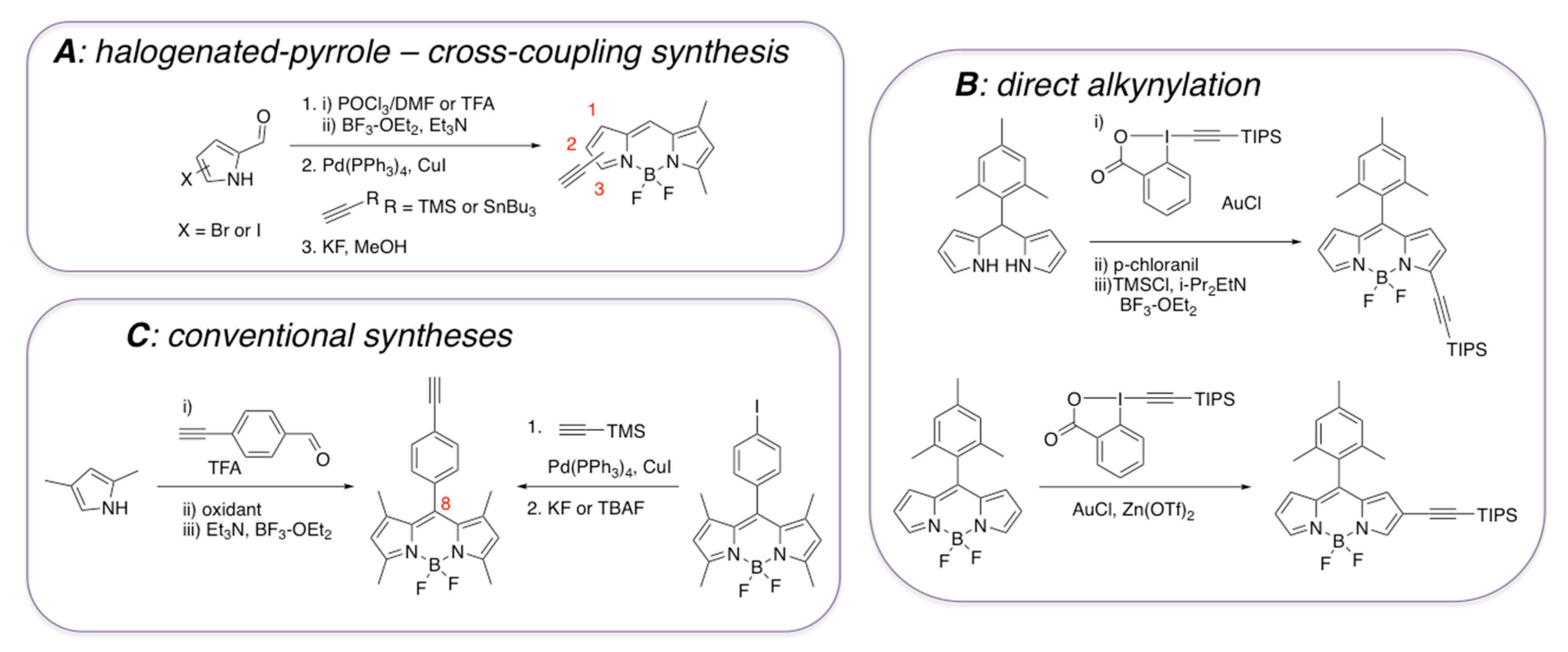
3.1. Non-Covalent Interactions of BODIPY Dyes with Aβ Fibrils
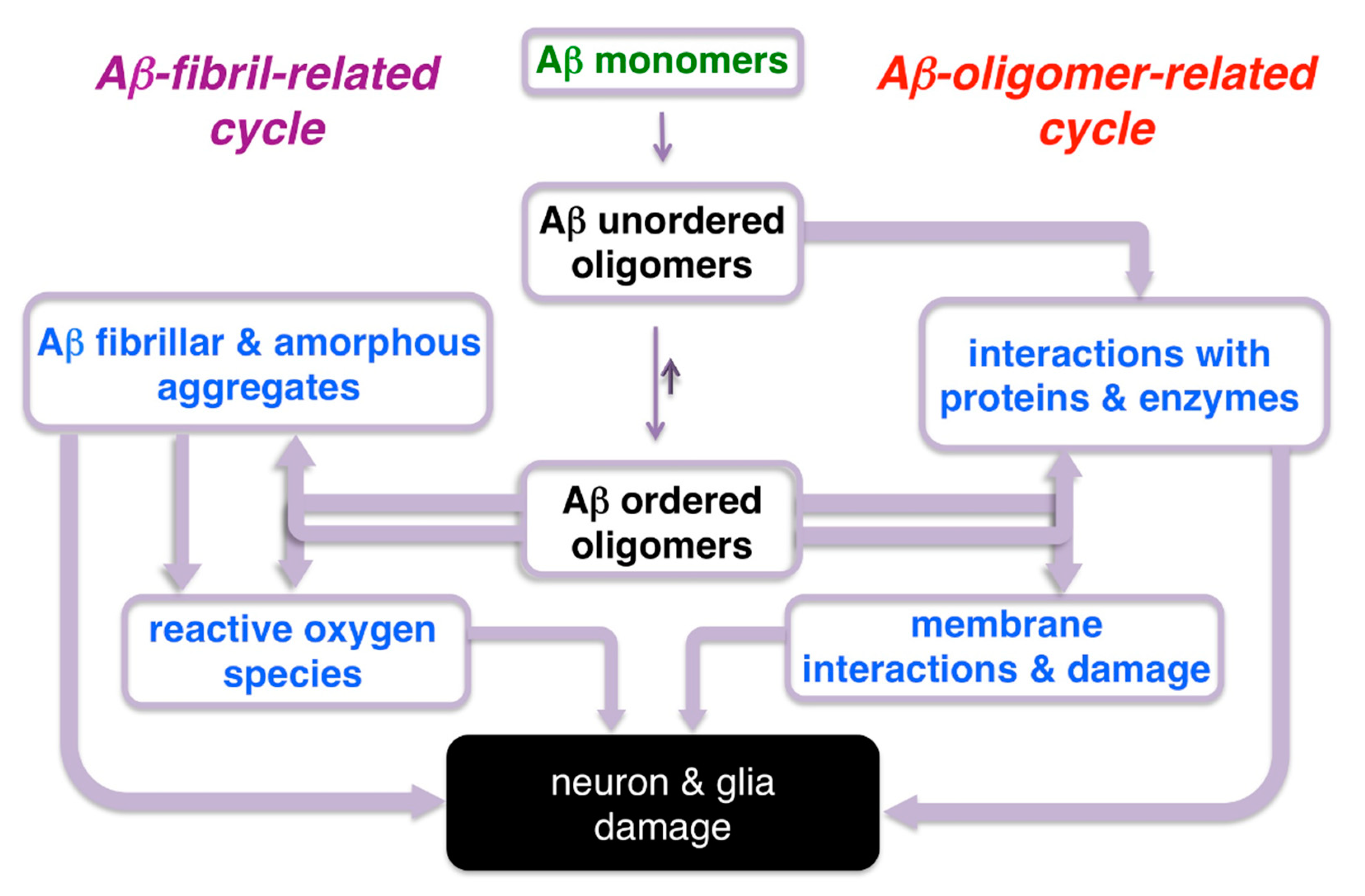
3.2. Non-Covalent Interactions of BODIPY Dyes with Aβ Oligomers
3.3. Photoinduced Electron Transfer-Based BODIPY Sensors
3.4. BODIPY–Amyloid Interactions in Membrane Environments
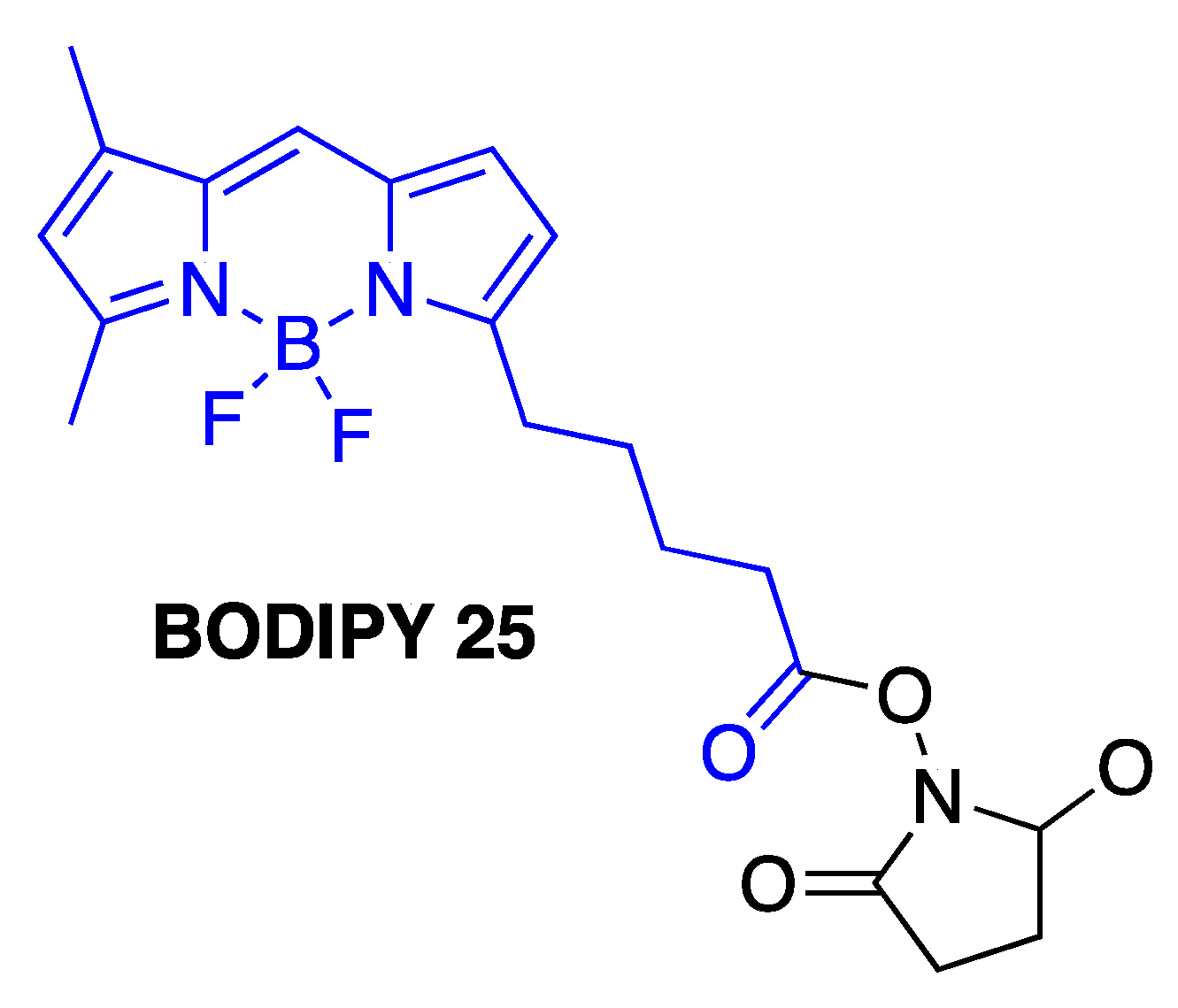
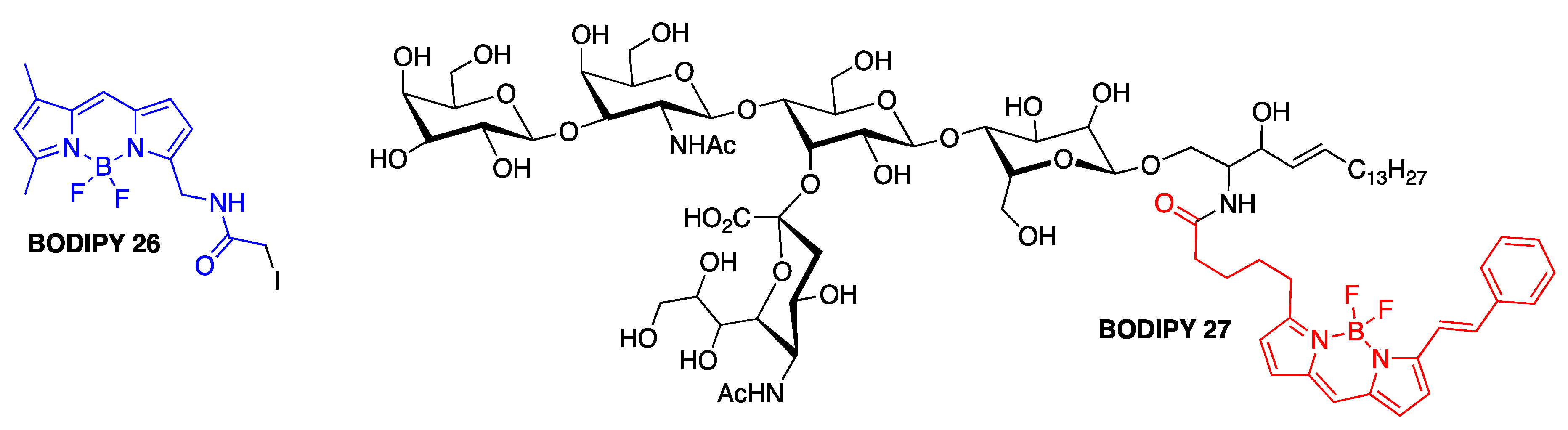
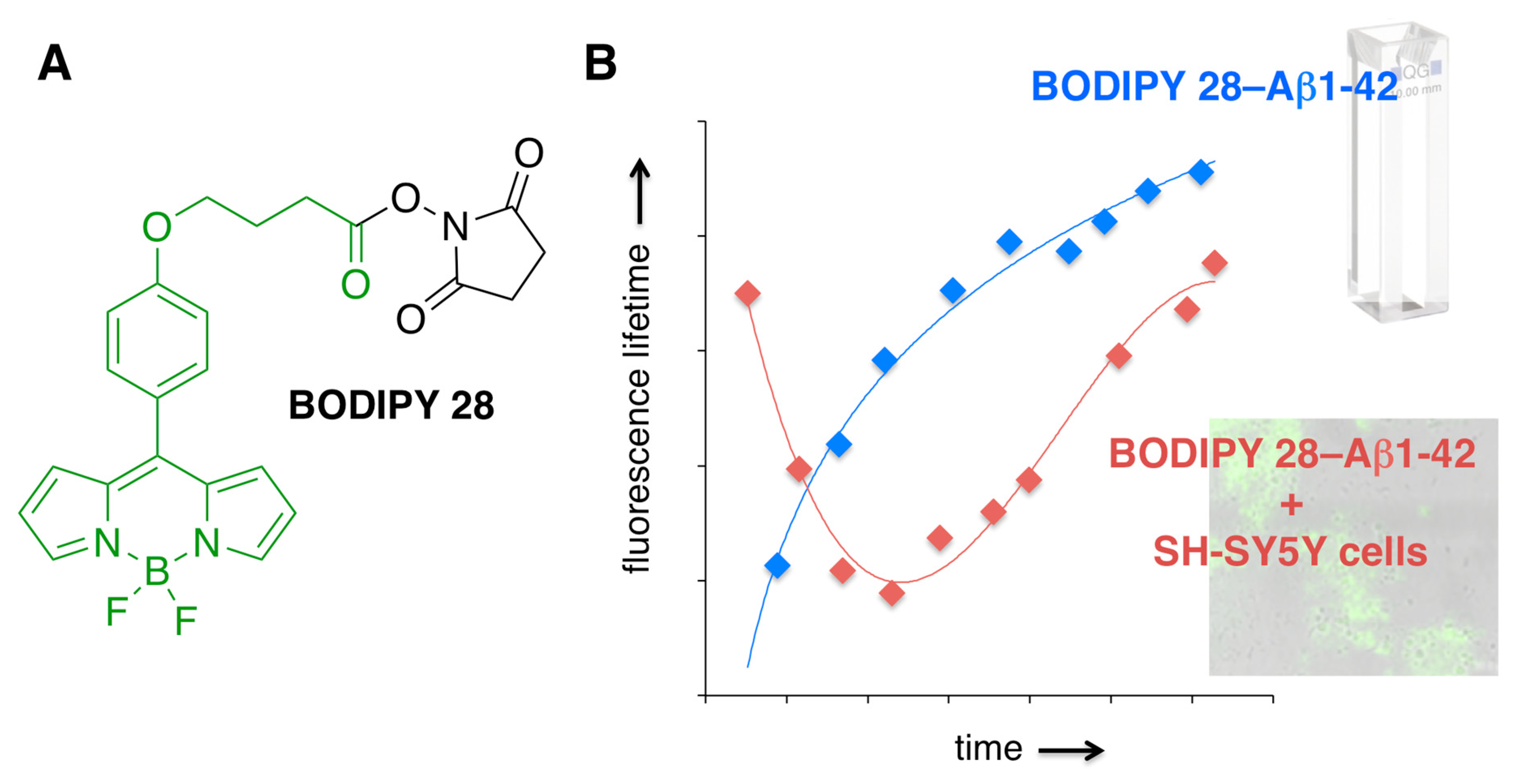
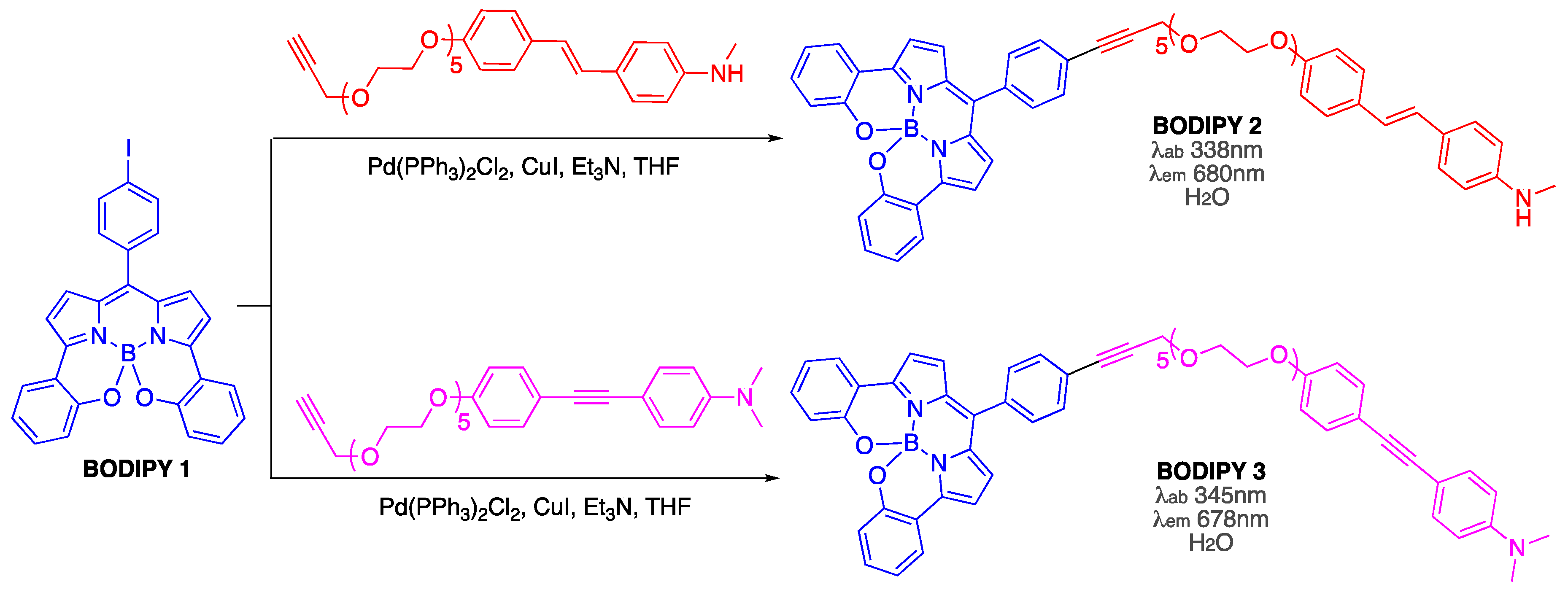
3.5. Covalent BODIPY-Aβ Probes
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Selkoe, D.J. Folding proteins in fatal ways. Nature 2003, 426, 900–904. [Google Scholar] [CrossRef]
- Bacciantini, M.; Giannoni, E.; Chiti, F.; Baroni, F.; Formigli, L.; Zurdo, J.; Taddei, N.; Ramponi, G.; Dobson, C.M.; Stefani, M. Inherent toxicity of aggregates implies a common mechanism for protein misfolding disease. Nature 2002, 416, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Knowles, T.P.J.; Vendruscolo, M.; Dobson, C.M. The amyloid state and its association with protein misfolding diseases. Nat. Mol. Cell Biol. 2014, 15, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Chiti, F.; Dobson, C.M. Amyloid formation by globular proteins under native conditions. Nat. Chem. Biol. 2009, 5, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ding, H.; Ran, X.; Tang, H.; Cao, D. Recent progress on reaction-based BODIPY probes for anion detection. Dye. Pigment. 2020, 172, 107857. [Google Scholar] [CrossRef]
- Kowada, T.; Maeda, H.; Kikuchi, K. BODIPY-based probes for the fluorescence imaging of biomolecules in living cells. Chem. Soc. Rev. 2015, 44, 4953–4972. [Google Scholar] [CrossRef]
- Ni, Y.; Wu, J. Far-red and near infrared BODIPY dyes: Synthesis and applications for fluorescent pH probes and bio-imaging. Org. Biomol. Chem. 2014, 12, 3774–3791. [Google Scholar] [CrossRef]
- Boens, N.; Leen, V.; Dehaen, W. Fluorescent indicators based on BODIPY. Chem. Soc. Rev. 2012, 41, 1130–1172. [Google Scholar] [CrossRef]
- Arora, H.; Ramesh, M.; Rajasekhar, K.; Govindaraju, T. Molecular tools to detect alloforms of Aβ and tau: Implications for multiplexing and multimodal diagnosis of Alzheimer’s disease. Bull. Chem. Soc. Jpn. 2020, 93, 507–546. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.; New, E.J.; Sunde, M. Strategies for the molecular imaging of amyloid and the value of a multimodal approach. ACS Sens. 2020, 5, 2268–2282. [Google Scholar] [CrossRef]
- Gyasi, Y.I.; Pang, Y.-P.; Li, X.-R.; Gu, J.-X.; Cheng, X.-J.; Liu, J.; Xu, T.; Liu, Y. Biological applications of near infrared fluorescence dye probes in monitoring Alzheimer’s disease. Eur. J. Med. Chem. 2020, 187, 111982. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ren, B.; Zhang, D.; Liu, Y.; Zhang, M.; Zhao, C.; Zheng, J. Design principles and fundamental understanding of biosensors for amyloid-β detection. J. Mater. Chem. B 2020, 8, 6179–6196. [Google Scholar] [CrossRef]
- Aliyan, A.; Cook, N.P.; Martí, A.A. Interrogating amyloid aggregates using fluorescent probes. Chem. Rev. 2019, 119, 11819–11856. [Google Scholar] [CrossRef]
- Jun, Y.W.; Cho, S.W.; Jung, J.; Huh, Y.; Kim, Y.; Kim, D.; Ahn, K.H. Frontiers in probing Alzheimer’s disease biomarkers with fluorescent small molecules. ACS Cent. Sci. 2019, 5, 209–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Kim, S.M.; Kim, H.Y.; Kim, Y. Fluorescence chemicals to detect insoluble and soluble amyloid-β aggregates. ACS Chem. Neurosci. 2019, 10, 2647–2657. [Google Scholar] [CrossRef]
- Cao, K.J.; Yang, J. Translational opportunities for amyloid-targeting fluorophores. Chem. Commun. 2018, 54, 9107–9118. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, L.; Faustino, C. Therapeutic strategies targeting amyloid-β in Alzheimer’s disease. Curr. Alzheimer Res. 2019, 16, 418–452. [Google Scholar] [CrossRef]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef]
- Ciccone, L.; Shi, C.; di Lorenzo, D.; Van Baelen, A.-C.; Tonali, N. The positive side of the Alzheimer’s disease amyloid cross-interactions: The case of the Aβ1-42 peptide with tau, TTR, CysC, and ApoA1. Molecules 2020, 25, 2439. [Google Scholar] [CrossRef]
- Makowski, L. The structural basis of amyloid strains in Alzheimer’s disease. ACS Biomater. Sci. Eng. 2020, 6, 2498–2505. [Google Scholar] [CrossRef]
- Eisenberg, D.S.; Sawaya, M.R. Implications for Alzheimer’s disease of an atomic resolution structure of amyloid-β(1-42) fibrils. Proc. Nat. Acad. Sci. USA 2016, 113, 9398–9400. [Google Scholar] [CrossRef] [Green Version]
- Miller, Y.; Ma, B.; Nussinov, R. Polymorphism in Alzheimer Aβ amyloid organization reflects conformational selection in a rugged energy landscape. Chem. Rev. 2010, 110, 4820–4838. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Selkoe, D.J. A mechanistic hypothesis for the impairment of synaptic plasticity by soluble Aβ oligomers from Alzheimer’s brain. J. Neurochem. 2020, 154, 583–597. [Google Scholar] [CrossRef]
- Fantini, J.; Chahinian, H.; Yahi, N. Progress toward Alzheimer’s disease treatment: Leveraging the Achilles’ heel of Aβ oligomers? Protein Sci. 2020, 29, 1748–1759. [Google Scholar] [CrossRef]
- Picone, P.; Nuzzo, D.; Giacomazza, D.; Di Carlo, M. β-Amyloid peptide: The cell compartment multi-faceted interaction in Alzheimer’s disease. Neurotox. Res. 2020, 37, 250–263. [Google Scholar] [CrossRef]
- Morsy, A.; Tripper, P.C. Current and emerging pharmacological targets for the treatment of Alzheimer’s disease. J. Alzheimers Dis. 2019, 72, S145–S176. [Google Scholar] [CrossRef]
- Lee, S.J.C.; Nam, E.; Lee, H.J.; Savelieff, M.G.; Lim, M.H. Towards an understanding of amyloid-β oligomers: Characterization, toxicity, mechanisms, and inhibitors. Chem. Soc. Rev. 2017, 46, 310–323. [Google Scholar] [CrossRef]
- Jarosz-Griffiths, H.H.; Noble, E.; Rushworth, J.V.; Hooper, N.M. Amyloid-β receptors: The good, the bad, and the prion protein. J. Biol. Chem. 2016, 291, 3174–3183. [Google Scholar] [CrossRef] [Green Version]
- Zou, K.; Gong, J.-S.; Yanagisawa, K.; Michikawa, M. A novel function of monomeric amyloid β-protein serving as an antioxidant molecule against metal-induced oxidative damage. J. Neurosci. 2002, 22, 4833–4841. [Google Scholar] [CrossRef] [Green Version]
- López-Toledano, M.A.; Shelanski, M.L. Neurogenic effect of β−amyloid peptide in the development of neural stem cells. J. Neurosci. 2004, 24, 5439–5444. [Google Scholar] [CrossRef]
- Tamagno, E.; Guglielmotto, M.; Moneleone, D.; Manassero, G.; Vasciaveo, V.; Tabaton, M. The unexpected role of Aβ1-42 monomers in the pathogenesis of Alzheimer’s disease. J. Alzheimers Dis. 2018, 62, 1241–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Roberts, C.J. Protein aggregation—Mechanisms, detection, and control. Int. J. Pharm. 2018, 550, 251–268. [Google Scholar] [CrossRef] [PubMed]
- Giles, U.; Ziessel, R.; Harriman, A. The chemistry of fluorescent Bodipy dyes: Versatility unsurpassed. Angew. Chem. Int. Ed. 2008, 47, 1184–1201. [Google Scholar] [CrossRef]
- Loudet, A.; Burgess, K. BODIPY dyes and their derivatives: Synthesis and spectroscopic properties. Chem. Rev. 2007, 107, 4891–4932. [Google Scholar] [CrossRef]
- Wang, J.; Boens, N.; Jiao, L.; Hao, E. Aromatic [b]-fused BODIPY dyes as promising near-infrared dyes. Org. Biomol. Chem. 2020, 18, 4135–4146. [Google Scholar] [CrossRef]
- Lu, H.; Mack, J.; Yang, Y.; Shen, Z. Structural modification strategies for the rational design of red/NIR region BODIPYs. Chem. Soc. Rev. 2014, 43, 4778–4823. [Google Scholar] [CrossRef] [Green Version]
- Boens, N.; Verbelen, B.; Ortiz, M.; Jiao, L.; Dehaem, W. Synthesis of BODIPY dyes through postfunctionalization of the boron dipyrromethene core. Coord. Chem. Rev. 2019, 399, 213024. [Google Scholar] [CrossRef]
- Parhi, A.K.; Kung, M.-P.; Ploessl, K.; Kung, H.F. Synthesis of fluorescent probes bas on stilbenes and diphenylacetylenes targeting β-amyloid plaques. Tetrahedron Lett. 2008, 49, 3395–3399. [Google Scholar] [CrossRef]
- Zhang, W.; Oya, S.; Kung, M.-P.; Hou, C.; Maier, D.L.; Kung, H.F. F-18 polyethyleneglycol stilbenes as PET imaging agents targeting Aβ aggregates in the brain. Nucl. Med. Biol. 2005, 32, 799–809. [Google Scholar] [CrossRef]
- Chandra, R.; Oya, S.; Kung, M.-P.; Hou, C.; Jin, L.-W.; Kung, H.F. New diphenylacetylenes as probes for positron emission tomographic imaging of amyloid plaques. J. Med. Chem. 2007, 50, 2415–2423. [Google Scholar] [CrossRef]
- Costela, A.; García-Moreno, I.; Pntado-Sierra, M.; Amat-Guerri, F.; Liras, M.; Sastre, R.; López Arbeloa, F.; Bañuelos Prieto, J.; López Arbeloa, I. New laser dye based on the 3-styryl analog of the BODIPY dye PM567. J. Photochem. Photobiol. A 2008, 198, 192–199. [Google Scholar] [CrossRef]
- Buyukcakir, O.; Bozdemir, O.A.; Kolemen, S.; Erbas, S.; Akkaya, E.U. Tetrastyryl-Bodipy dyes: Convenient synthesis and characterization of elusive near IR fluorophores. Org. Lett. 2009, 11, 4644–4647. [Google Scholar] [CrossRef] [PubMed]
- Shivran, N.; Mula, S.; Ghanty, T.K.; Chattopadhyay, S. Steric strain release-directed regioselective functionalization of meso-methyl Bodipy dyes. Org. Lett. 2011, 13, 5870–5873. [Google Scholar] [CrossRef] [PubMed]
- Madhu, S.; Rajeswara Rao, M.; Shaikh, M.S.; Ravikanth, M. 3,5-diformylboron dipyrromethenes as fluorescent pH sensors. Inorg. Chem. 2011, 50, 4392–4400. [Google Scholar] [CrossRef]
- Sathyamoorthi, G.; Wolford, L.T.; Haag, A.M.; Boyer, J.H. Selective side-chain oxidation of peralkylated pyrromethene-BF2 complexes. Heteroat. Chem. 1994, 5, 245–249. [Google Scholar] [CrossRef]
- Jiao, L.; Yu, C.; Li, J.; Wang, Z.; Wu, M.; Hao, E. β-Formyl-BODIPYs from the Vilsmeier-Haack reaction. J. Org. Chem. 2009, 74, 7525–7528. [Google Scholar] [CrossRef]
- Lakshmi, V.; Ravikanth, M. Synthesis of conjugated BODIPYs via the Wittig reaction. J. Org. Chem. 2013, 78, 4993–5000. [Google Scholar] [CrossRef]
- Leen, V.; Van der Auweraer, M.; Boens, N.; Dehaen, W. Vicarious nucleophilic substitution of α-hydrogen on BODIPY and its extension to direct ethenylation. Org. Lett. 2011, 13, 1470–1473. [Google Scholar] [CrossRef]
- Thivierge, C.; Bandichhor, R.; Burgess, K. Spectral dispersion and water solubilization of BODIPY dyes via palladium-catalyzed C-H functionalization. Org. Lett. 2007, 9, 2135–2138. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Gong, Q.; Wang, H.; Hao, E.; Lo, P.-C.; Jiao, L. β-Alkenyl BODIPY dyes: Regioselective synthesis via oxidative C-H olefination, photophysical properties, and bioimaging studies. J. Org. Chem. 2019, 84, 5078–5090. [Google Scholar] [CrossRef]
- Wirtz, M.; Grüter, A.; Rebmann, P.; Dier, T.; Volmer, D.A.; Huch, V.; Jung, G. Two-color emissive probes for click reactions. Chem. Commun. 2014, 50, 12694–12697. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.-Y.; Guan, Y.-S.; Chen, Y.-Z.; Wu, L.-Z.; Tung, C.-H.; Yang, Q.-Z. BODIPY-based ratiometric fluorescent sensor for highly selective detection of glutathione over cysteine and homocysteine. J. Am. Chem. Soc. 2012, 134, 18928–18931. [Google Scholar] [CrossRef]
- Shimada, T.; Mori, S.; Ishida, M.; Furuta, H. Regioselectively α- and β-alkynylated BODIPY dyes via gold(I)-catalyzed direct C-H functionalization and their photophysical properties. Beilstein J. Org. Chem. 2020, 16, 587–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leen, V.; Yuan, P.; Wang, L.; Boens, N.; Dehaen, W. Synthesis of meso-halogenated BODIPYs and access to meso-substituted analogues. Org. Lett. 2012, 14, 6150–6153. [Google Scholar] [CrossRef]
- Dhokale, B.; Jadhav, T.; Mobin, S.M.; Misra, R. Meso alkynylated tetraphenylethylene (TPE) and 2,3,3-triphenylacrylonitrile (TPAN) substituted BODIPYs. J. Org. Chem. 2015, 80, 8018–8025. [Google Scholar] [CrossRef]
- Jameson, L.P.; Dzyuba, S.V. Expeditious, mechanochemical synthesis of BODIPY dyes. Beilstein J. Org. Chem. 2013, 9, 786–790. [Google Scholar] [CrossRef]
- Ogle, M.M.; Smith McWilliams, A.D.; Ware, M.J.; Curley, S.A.; Corr, S.J.; Martí, A.A. Sensing temperature in vitro and in cells using BODIPY molecular probe. J. Phys. Chem. B 2019, 123, 7282–7289. [Google Scholar] [CrossRef]
- Raut, S.; Kimball, J.; Fudala, R.; Doan, H.; Maliwal, B.; Sabnis, N.; Lacko, A.; Gryczynski, I.; Dzyuba, S.V.; Gryczynski, Z. A homodimeric BODIPY rotor as a fluorescent viscosity sensor for membrane-mimicking and cellular environments. Phys. Chem. Chem. Phys. 2014, 16, 27037–27042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raut, S.L.; Kimball, J.D.; Fudala, R.; Bora, I.; Chib, R.; Jaafari, H.; Castillo, M.K.; Smith, N.W.; Gryczynski, I.; Dzyuba, S.V.; et al. A triazine-based BODIPY trimer as a molecular viscometer. Phys. Chem. Chem. Phys. 2016, 18, 4535–4540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, M.; Ishikawa, M.; Kimura, H.; Hayashi, S.; Matsumura, K.; Watanabe, H.; Shimizu, Y.; Cheng, Y.; Cui, M.; Kawashima, H.; et al. Development of dual functional SPECT/fluorescent probes for imaging cerebral β-amyloid plaques. Bioorg. Med. Chem. Lett. 2010, 20, 3885–3888. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Watanabe, H.; Kimura, H.; Saji, H. BODIPY-based molecular probe for imaging of cerebral β-amyloid plaques. ACS Chem. Neurosci. 2012, 3, 319–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, H.; Ono, M.; Matsumura, K.; Yoshimura, M.; Kimura, H.; Saji, H. Molecular imaging of β-amyloid plaques with near-infrared boron dipyrromethane (BODIPY)-based fluorescent probes. Mol. Imaging 2013, 12, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Sozmen, F.; Kolemen, S.; Kumada, H.-O.; Ono, M.; Saji, H.; Akkaya, E.U. Designing BODIPY-based probes for fluorescence imaging of β-amyloid plaques. RSC Adv. 2014, 4, 51032–51037. [Google Scholar] [CrossRef] [Green Version]
- Smith, N.W.; Alonso, A.; Brown, C.M.; Dzyuba, S.V. Triazole-containing BODIPY dyes as novel fluorescent probes for soluble oligomers of amyloid Aβ1-42 peptide. Biochem. Biophys. Res. Commun. 2010, 391, 1455–1458. [Google Scholar] [CrossRef]
- Jameson, L.P.; Smith, N.W.; Dzyuba, S.V. Dye-binding assays for evaluation of the effects of small molecule inhibitor on amyloid (Aβ) self-assembly. ACS Chem. Neurosci. 2012, 3, 807–819. [Google Scholar] [CrossRef] [Green Version]
- Smith, N.W.; Annunziata, O.; Dzyuba, S.V. Amphotericin B interactions with soluble oligomers of amyloid Aβ1-42 peptide. Bioorg. Med. Chem. 2009, 17, 2366–2370. [Google Scholar] [CrossRef]
- Tonali, N.; Dodero, V.I.; Kaffy, J.; Herics, L.; Ongeri, S.; Sewald, N. Real-time BODIPY-binding assay to screen inhibitors of the early oligomerization process of Aβ1-42 peptide. ChemBioChem 2020, 21, 1129–1135. [Google Scholar] [CrossRef]
- Teoh, C.L.; Su, D.; Sahu, S.; Yun, S.-W.; Drummond, E.; Prelli, F.; Lim, S.; Cho, S.; Ham, S.; Wisniewski, T.; et al. Chemical fluorescent probe for detection of Aβ oligomers. J. Am. Chem. Soc. 2015, 137, 13503–13509. [Google Scholar] [CrossRef] [Green Version]
- Cuvas-Yañez, E.; Muchowski, J.M.; Cruz-Almanza, R. Rhodium(II) catalyzed intermolecular insertion of carbenoids derived from 2-pyrrolyl and 3-indolyl α-diazo-β-ketoesters and α-diazoketones. Tetrahedron 2004, 60, 1505–1511. [Google Scholar] [CrossRef]
- Malan, S.F.; van Marle, A.; Menge, W.M.; Zukiana, V.; Hoffman, M.; Timmerman, H.; Leurs, R. Fluorescent ligands for the histamine H2 receptor: Synthesis and preliminary characterization. Bioorg. Med. Chem. 2004, 12, 6495–6503. [Google Scholar] [CrossRef]
- Yun, S.-W.; Kang, N.-Y.; Park, S.-J.; Ha, H.-H.; Kim, Y.K.; Lee, J.-S.; Chang, Y.-T. Diversity oriented fluorescence library approach (DOFLA) for live cell imaging probe development. Acc. Chem. Res. 2014, 47, 1277–1286. [Google Scholar] [CrossRef]
- Shi, Z.; Han, X.; Hu, W.; Bai, H.; Peng, B.; Ji, L.; Fan, Q.; Li, L.; Huang, W. Bioapplications of small molecule aza-BODIPY: From rational structural design to in vivo investigations. Chem. Soc. Rev. 2020, 49, 7533–7567. [Google Scholar] [CrossRef] [PubMed]
- Jameson, L.P.; Dzyuba, S.V. Aza-BODIPY: Improved synthesis and interaction with soluble Aβ1-42. Bioorg. Med. Chem. Lett. 2013, 23, 1732–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Li, M.; Fan, J.; Peng, X. Activity-based sensing and theranostic probes based on protoinduced electron transfer. Acc. Chem. Res. 2019, 52, 2818–2831. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, E.; Kazan, H.H.; Çosut, B. Recent chem-/biosensor and bioimaging studies based on indole-decorated BODIPYs. Luminescence 2020, 35, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Farione, M.; Urbanska, K.; Lawlicki, M. BODIPY- and porphyrin-based sensors for recognition of amino acids and their derivatives. Molecules 2020, 25, 4523. [Google Scholar] [CrossRef]
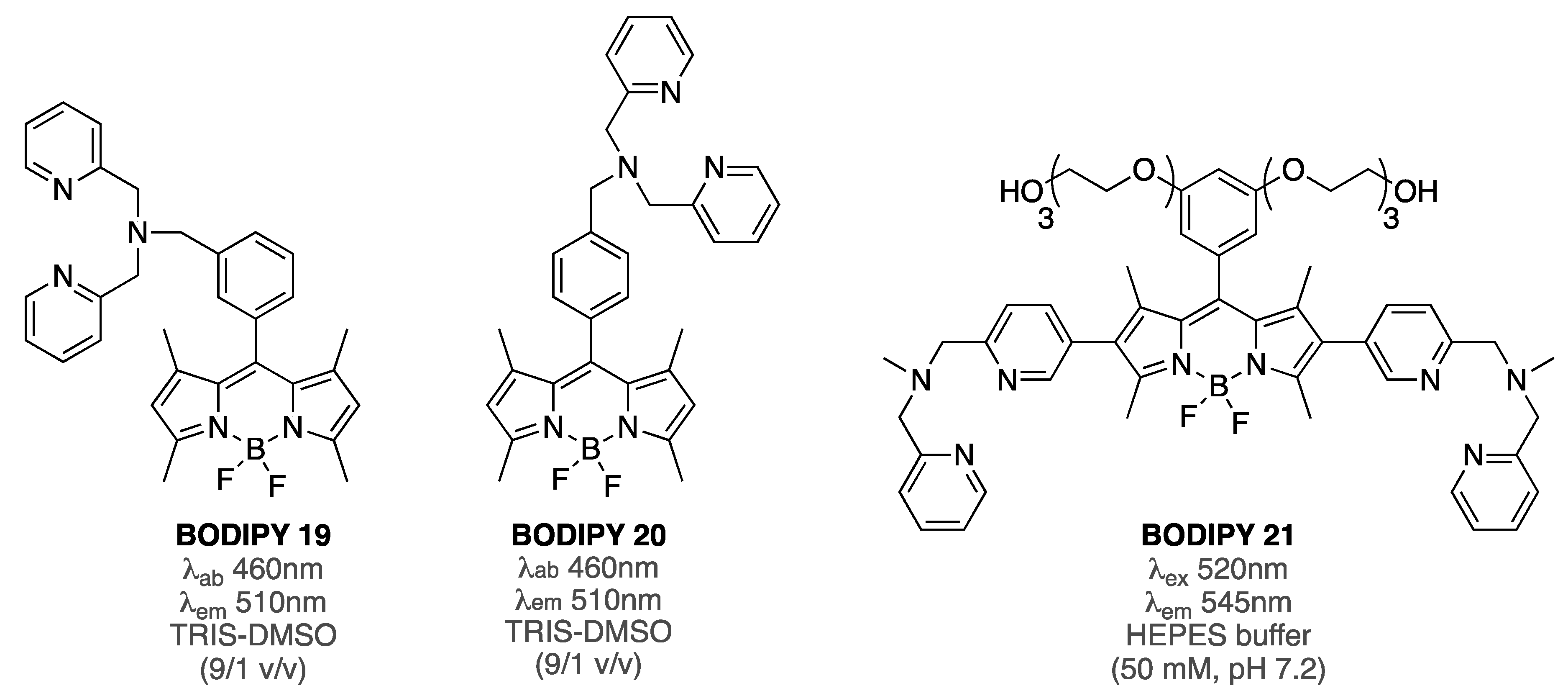
- Kong, M.-Y.; Chen, Q.-Y.; Yao, L.; Wang, Y.-B. Spectroscopic study on the interaction of Aβ42 with di(picolyl)amine derivatives and the toxicity of SH-S5Y5 cells. Spectrochim. Acta A 2015, 138, 225–228. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Q.-Y.; Wang, P.-D.; Wu, Y. Multifunctional BODIPY derivatives to image cancer cells and sense copper(II) ions in living cells. RSC Adv. 2013, 3, 5524–5528. [Google Scholar] [CrossRef]
- Zhao, T.-T.; Chen, Q.-Y.; Wang, P.-D.; Chen, Z.-P. A DNA-Ag cluster as a sensor for BODIPY isomers and HepG-2 cells. RSC Adv. 2014, 4, 10390–10394. [Google Scholar] [CrossRef]
- Ojida, A.; Sakamoto, T.; Inoe, M.-a.; Fujishima, S.-h.; Lippens, G.; Hamachi, I. Probe for selective detection of neurofibrillary tangles in the brains of Alzheimer’s disease patients. J. Am. Chem. Soc. 2009, 131, 6543–6548. [Google Scholar] [CrossRef]
- Ren, W.; Xu, M.; Liang, S.H.; Xiang, H.; Tang, L.; Zhang, M.; Ding, M.; Ding, D.; Li, X.; Zhang, H.; et al. Discovery of novel fluorescent probe for the sensitive detection of β-amyloid deposits. Biosens. Bioelectron. 2016, 75, 136–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, W.; Zhang, J.; Peng, C.; Xiang, H.; Chen, J.; Peng, C.; Zhu, W.; Huang, R.; Zhang, H.; Hu, Y. Fluorescent imaging of β-amyloid using BODIPY based near-infrared off-on fluorescent probe. Bioconjugate Chem. 2018, 29, 3459–3466. [Google Scholar] [CrossRef]
- Bharadwaj, P.; Solomon, T.; Malajczuk, C.J.; Mancera, R.L.; Howard, M.; Arrigan, D.W.M.; Newsholme, P.; Martins, R.N. Role of the cell membrane interface in modulating production and uptake of Alzheimer’s beta amyloid protein. Biochim. Biophys. Acta BBA–Biomembr. 2018, 1860, 1639–1651. [Google Scholar] [CrossRef] [PubMed]
- Relini, A.; Marano, N.; Gliozzi, A. Misfolding of amyloidogenic proteins and their interactions with membranes. Biomolecules 2014, 4, 20–55. [Google Scholar] [CrossRef] [Green Version]
- López-Duarte, I.; Vu, T.T.; Izquierdo, M.A.; Bull, J.A.; Kuimova, M.K. A molecular rotor for measuring viscosity in plasma membranes of live cells. Chem. Commun. 2014, 50, 5282–5284. [Google Scholar] [CrossRef]
- Kubánková, M.; López-Duarte, I.; Kiryushko, D.; Kuimova, M.K. Molecular rotors report on changes in live cell plasma membrane microviscosity upon interaction with beta-amyloid aggregates. Soft Matter 2018, 14, 9466–9474. [Google Scholar] [CrossRef]
- Dresser, L.; Hunter, P.; Yendybayeva, F.; Hargreaves, A.L.; Howard, J.A.L.; Evans, G.J.O.; Leake, M.C.; Quinn, S.D. Amyloid-β oligomerization monitored by single-molecule stepwise photobleaching. Methods 2020, in press. [Google Scholar] [CrossRef]
- Sharda, N.; Pengo, T.; Wang, Z.; Kandimalla, K.K. Amyloid-β peptides disrupt interactions between VAMP-2 and SNAP-25 in neuronal cells as determined by FRET/FLIM. J. Alzheimer’s Dis. 2020, 77, 1–13. [Google Scholar] [CrossRef]
- Schneider, M.; Walta, S.; Cadek, C.; Richtering, W.; Willbold, D. Fluorescence correlation spectroscopy reveals a cooperative unfolding of monomeric amyloid-β 42 with a low Gibbs free energy. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gouwens, L.K.; Makoni, N.; Rogers, V.A.; Nicols, M.R. Amyloid-β42 protofibrils are internalized by microglia more extensively than monomers. Brain Res. 2016, 1648, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Kedia, N.; Illes-Togh, E.; Haralampiev, I.; Prisner, S.; Herrmann, A.; Wanker, E.E.; Bieschke, J. Amyloid-β(1-42) aggregation initiates its cellular uptake and cytotoxicity. J. Biol. Chem. 2016, 291, 19590–19606. [Google Scholar] [CrossRef] [Green Version]
- Garai, K.; Frieden, C. Quantitative analysis of the time course of Aβ oligomerization and subsequent growth steps using tetramethylrhodamine-labeled Aβ. Proc. Natl. Acad. Sci. USA 2013, 110, 3321–3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stine, W.B.; Jungbauer, L.; Yu, C.; LaDu, M.J. Preparing synthetic Aβ in different aggregation states. Methods Mol. Biol. 2011, 670, 13–32. [Google Scholar] [CrossRef] [Green Version]
- Mandrekar, S.; Jiang, Q.; Lee, C.Y.D.; Koenigsknecht-Talboo, J.; Holtzman, D.M.; Landreth, G.E. Microglia mediate the clearance of soluble Aβ through fluid phase macropinocytosis. J. Neurosci. 2009, 29, 4252–4262. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Xu, L.; Yang, J.; Peng, X.; Wang, H.; Yu, N.; Hua, Y.; Zhao, J.; He, J.; Hong, T. The effect of fluorescent labels on Aβ42 aggregation detected by fluorescence correlation spectroscopy. Biopolymers 2018, 109, e23237. [Google Scholar] [CrossRef]
- Wägele, J.; De Sio, S.; Voigt, B.; Balnach, J.; Ott, M. How fluorescent tags modify oligomers size distributions of the Alzheimer peptide. Biophys. J. 2019, 116, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, A.; Kinjo, M. State-of-the-art fluorescence fluctuation-based spectroscopic techniques for the study of protein aggregation. Int. J. Mol. Sci. 2018, 19, 964. [Google Scholar] [CrossRef] [Green Version]
- Yanagisawa, K. GM1 ganglioside and Alzheimer’s disease. Glycoconj. J. 2015, 32, 87–91. [Google Scholar] [CrossRef]
- Matsuzaki, K. How do membranes initiate Alzheimer’s disease? Formation of toxic amyloid fibrils by the amyloid β-protein on ganglioside clusters. Acc. Chem. Res. 2014, 47, 2397–2404. [Google Scholar] [CrossRef]
- Mikhalyov, I.; Olofsson, A.; Gröbner, G.; Johansson, L.B.-Å. Designed fluorescent probes reveal interactions between amyloid-β(1-40) peptides and GM1 gangliosides in micelles and lipid vesicles. Biophys. J. 2010, 99, 1510–1519. [Google Scholar] [CrossRef] [Green Version]
- Kubankova, M.; López-Duarte, I.; Bull, J.A.; Vudukul, D.M.; Serpell, L.S.; de Saint Victor, M.; Stride, E.; Kuimova, M.K. Probing supramolecular protein assembly using covalently attached fluorescent molecular rotors. Biomaterials 2017, 139, 195–201. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aβ | BODIPY | Reference |
|---|---|---|
| Fibrils/aggregates/AD homogenates | ||
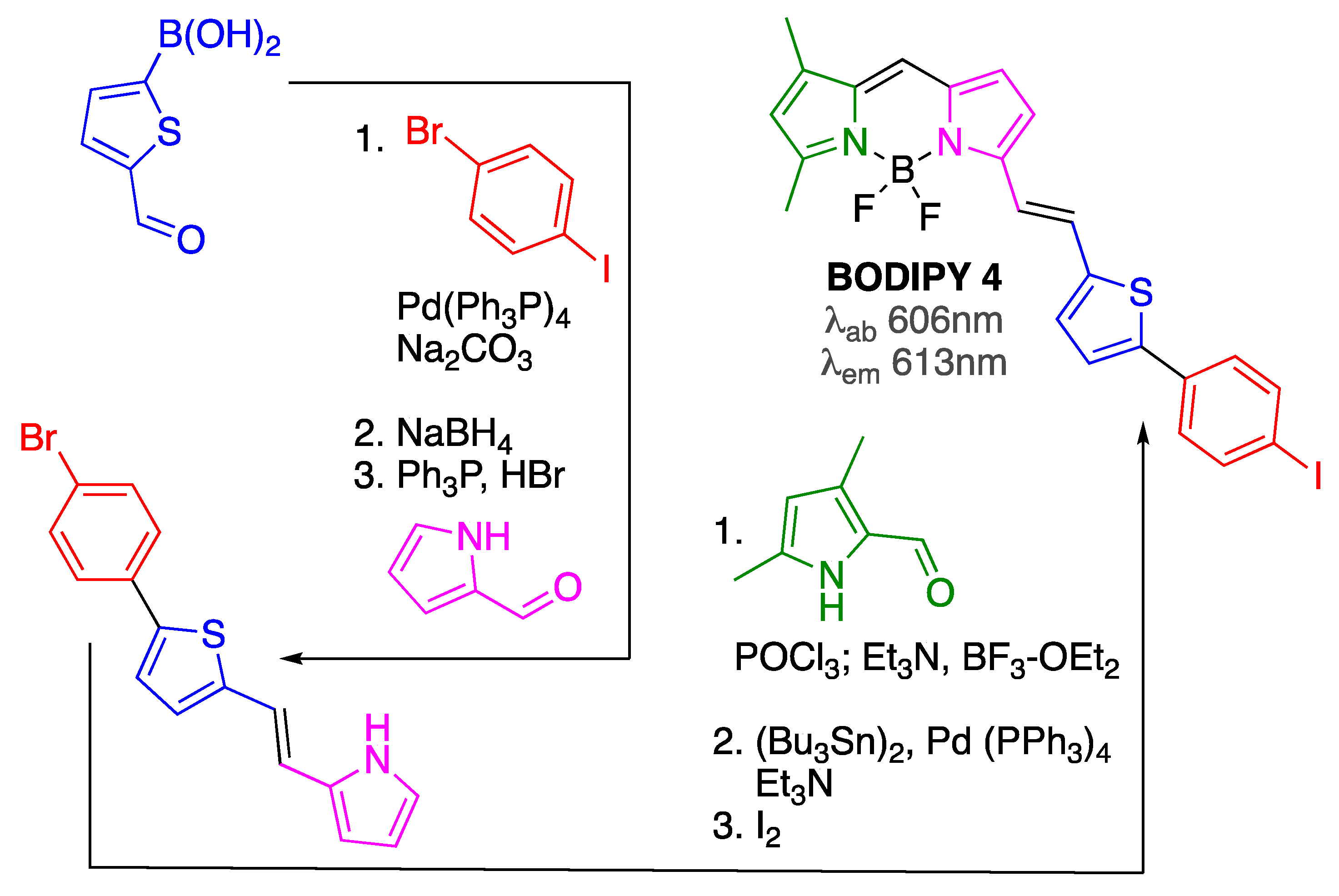
| BODIPY 4 | [60] | |
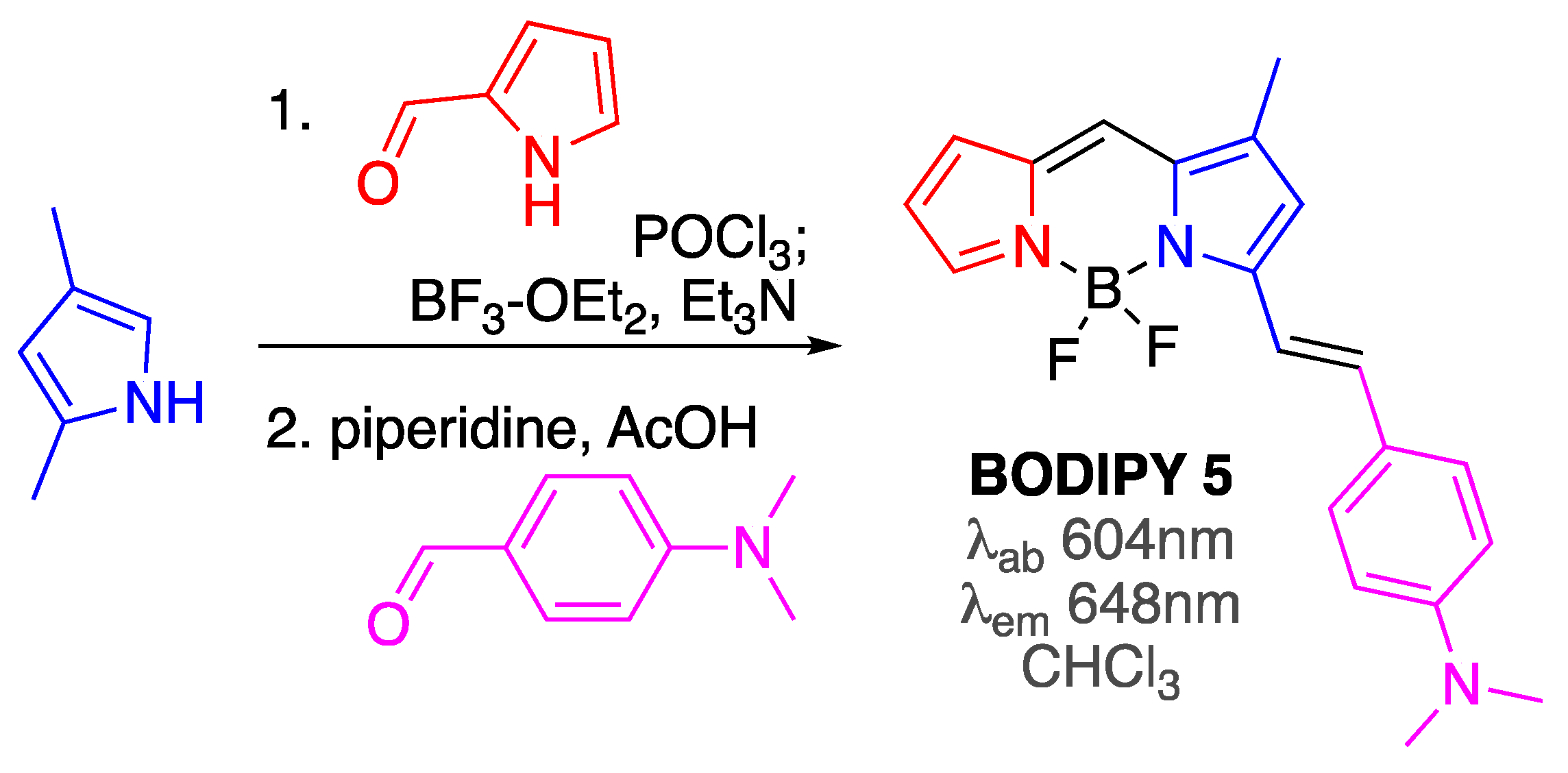
| BODIPY 5 | [61] | |
| BODIPY 6–9 | [62] | |
| BODIPY 10–13 | [63] | |
| BODIPY 19 | [77] | |
| BODIPY 21 | [80] | |
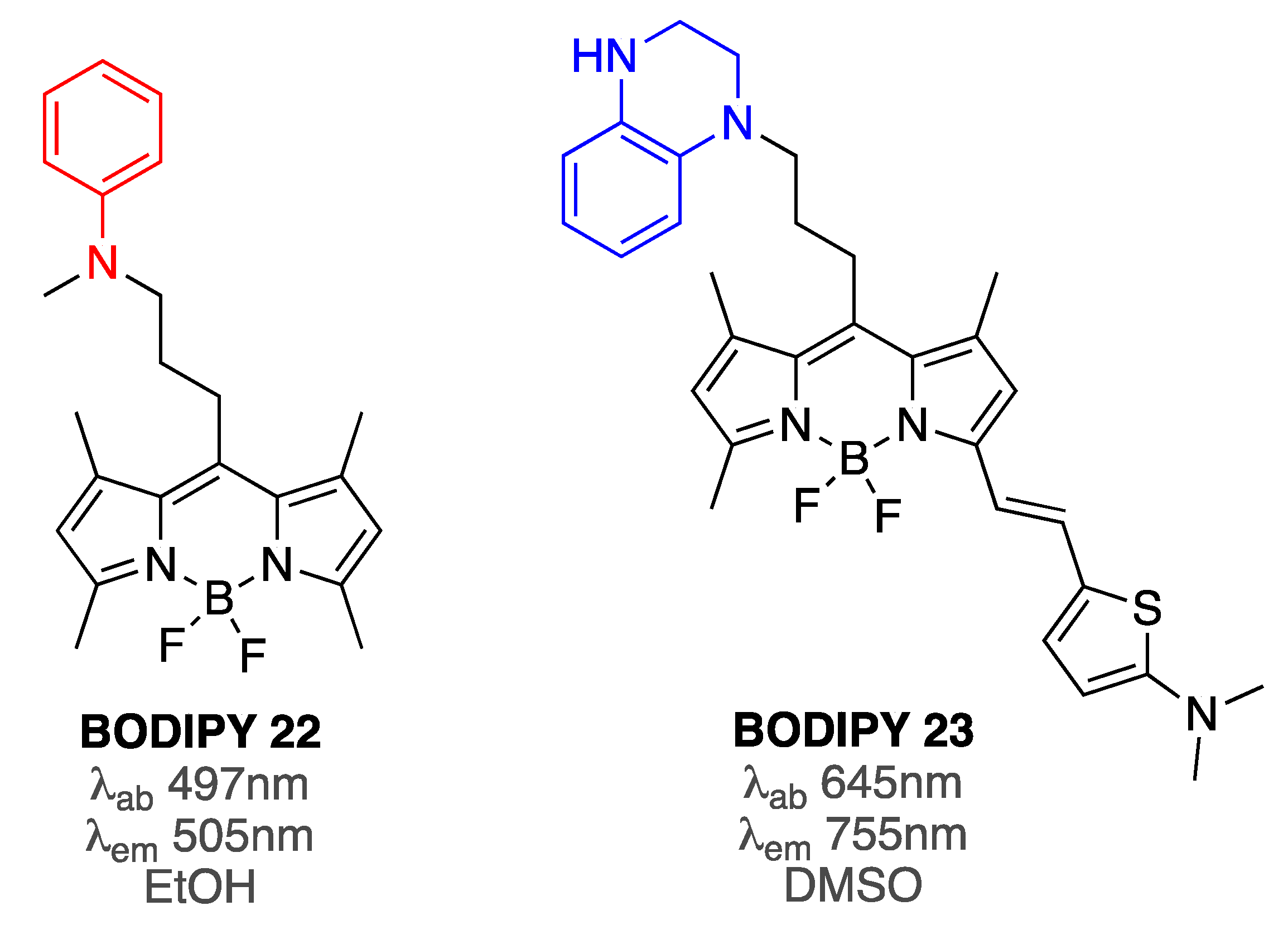
| BODIPY 22 | [81] | |
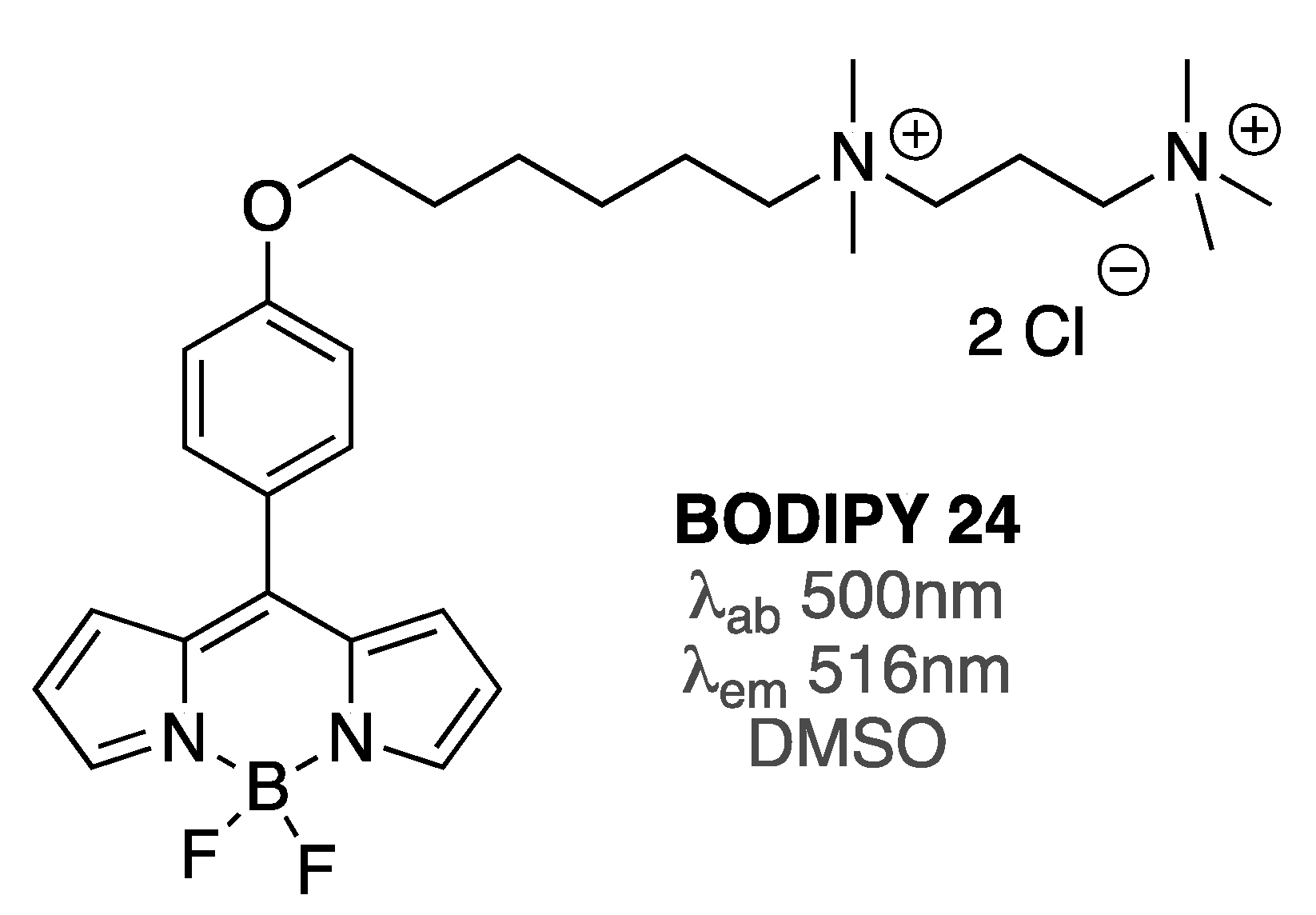
| BODIPY 24 | [85] | |
| Oligomers | ||
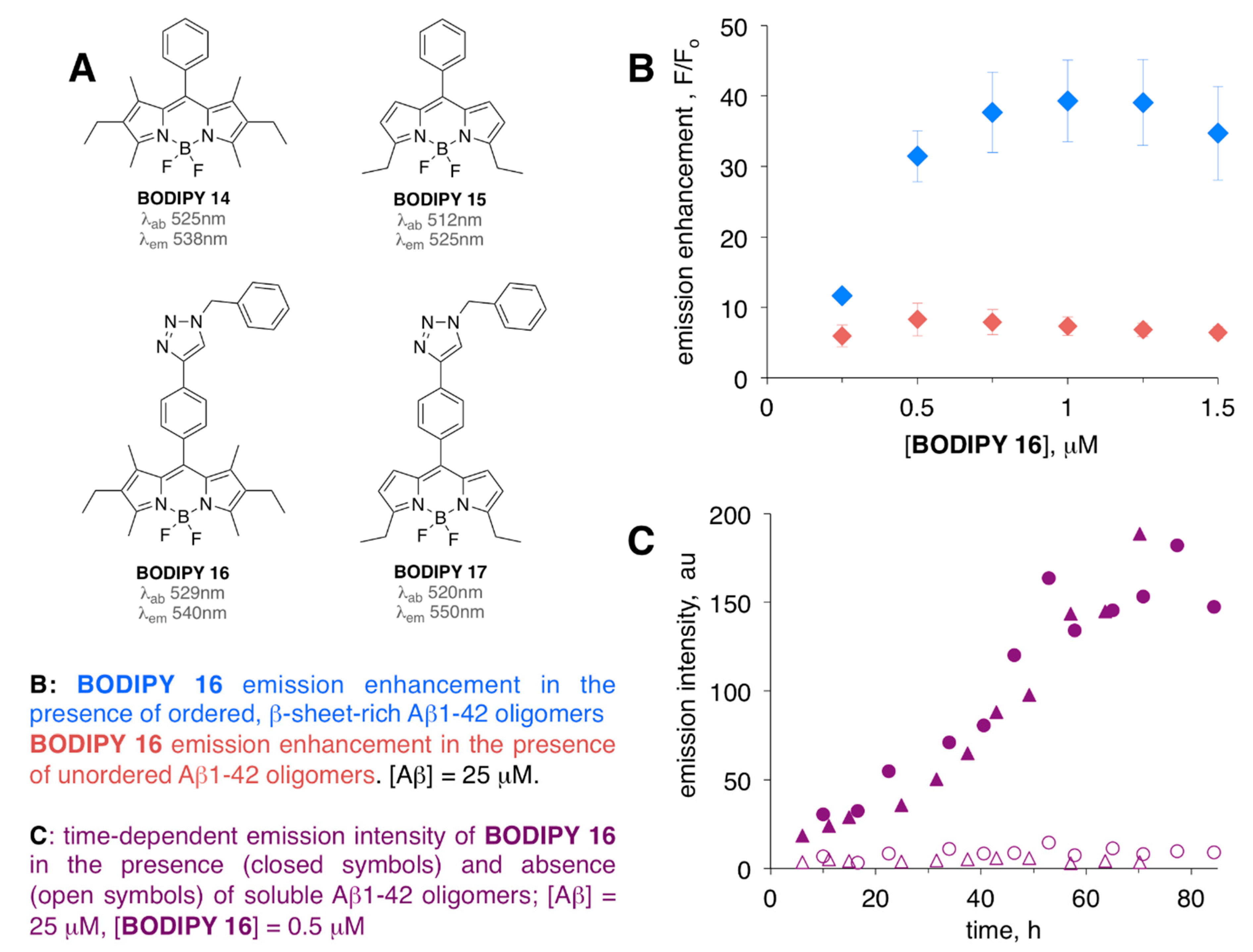
| BODIPY 16 | [64,67] | |
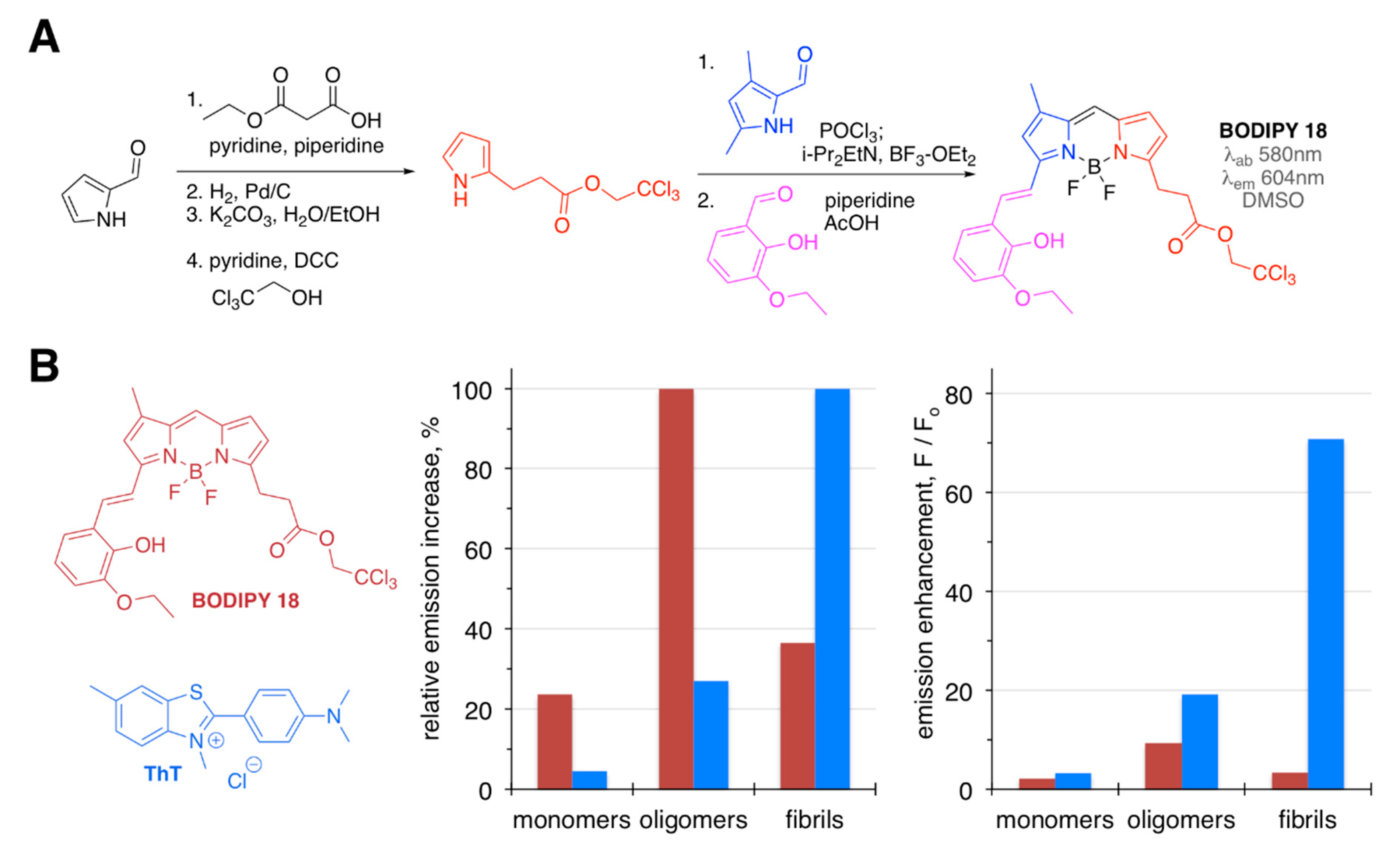
| BODIPY 18 | [68] | |
| BODIPY 23 | [82] | |
| BODIPY 24 | [85] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dzyuba, S.V. BODIPY Dyes as Probes and Sensors to Study Amyloid-β-Related Processes. Biosensors 2020, 10, 192. https://doi.org/10.3390/bios10120192
Dzyuba SV. BODIPY Dyes as Probes and Sensors to Study Amyloid-β-Related Processes. Biosensors. 2020; 10(12):192. https://doi.org/10.3390/bios10120192
Chicago/Turabian StyleDzyuba, Sergei V. 2020. "BODIPY Dyes as Probes and Sensors to Study Amyloid-β-Related Processes" Biosensors 10, no. 12: 192. https://doi.org/10.3390/bios10120192
APA StyleDzyuba, S. V. (2020). BODIPY Dyes as Probes and Sensors to Study Amyloid-β-Related Processes. Biosensors, 10(12), 192. https://doi.org/10.3390/bios10120192