Microfluidic Lab-on-a-Chip Based on UHF-Dielectrophoresis for Stemness Phenotype Characterization and Discrimination among Glioblastoma Cells

 , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
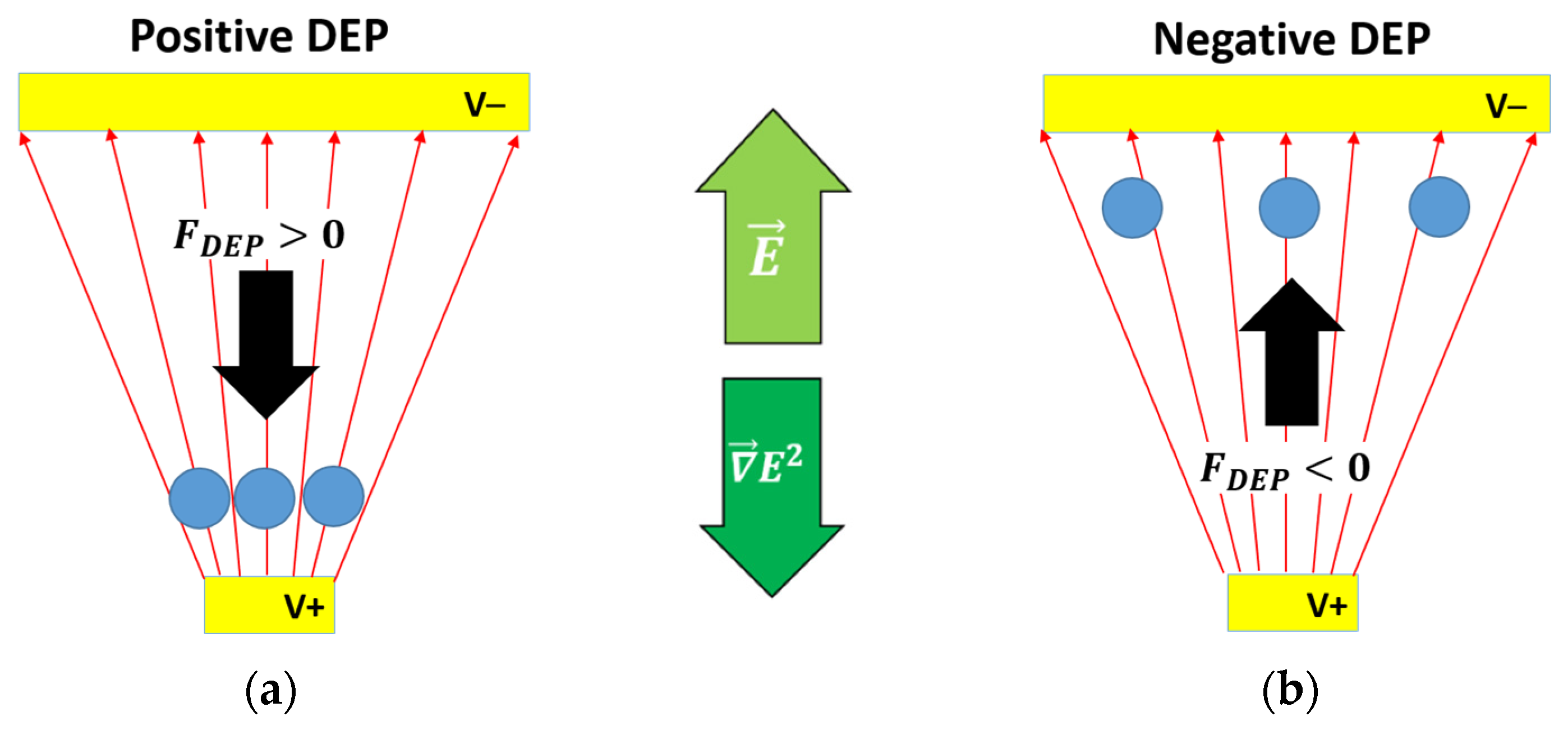
1.1. Basic DEP Theory
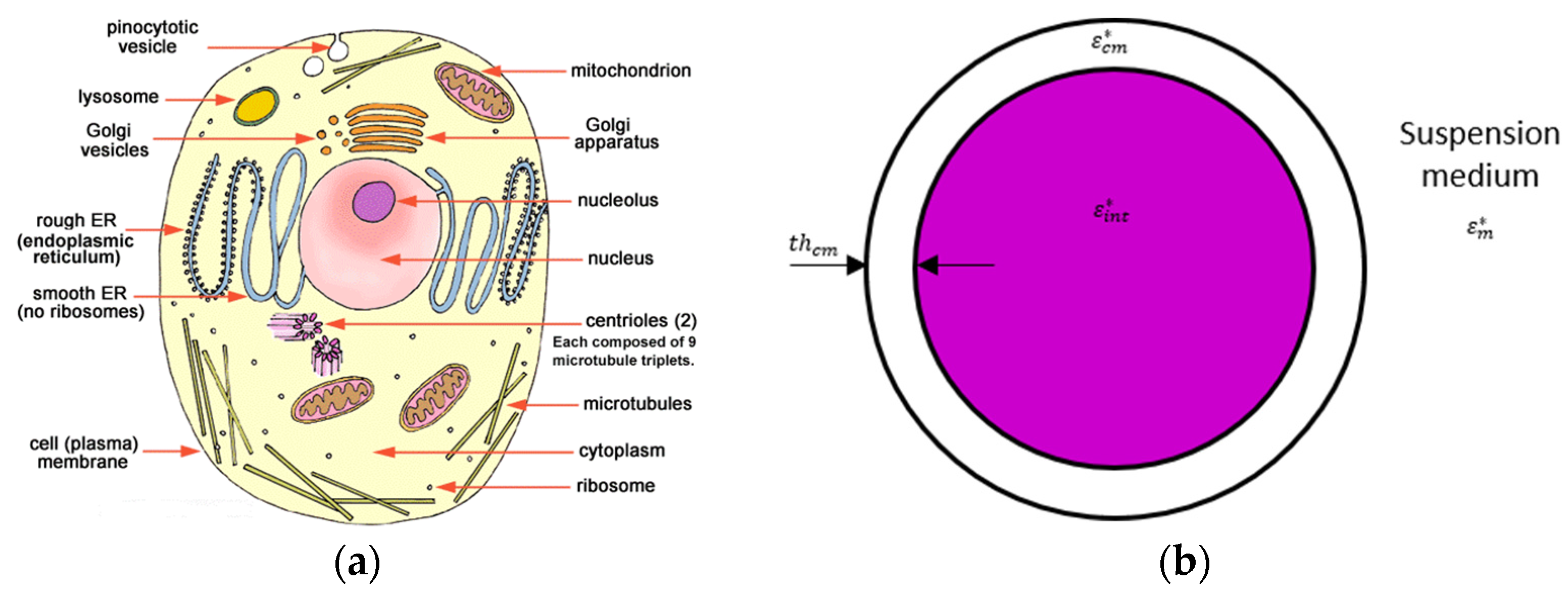
1.2. From a Biological Cell to a Single-Shell Model
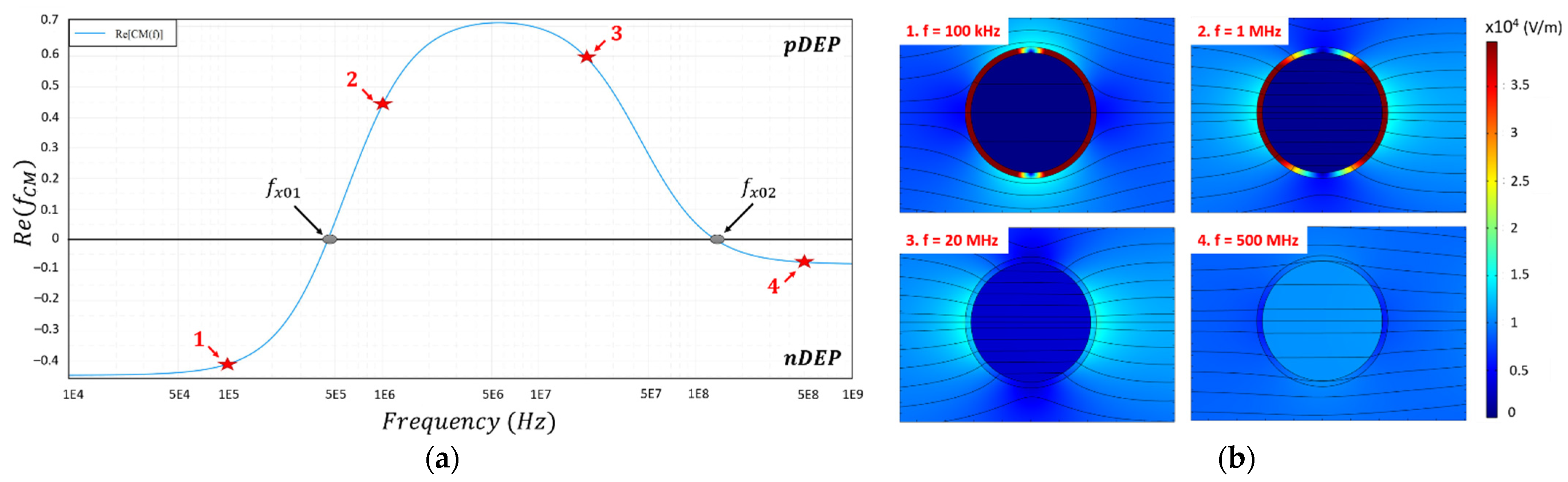
1.3. Effect of the Cellular Dielectric Properties on the Clausius–Mossotti Factor
2. Materials and Methods
2.1. Cell Line Culture
- Normal Normoxia Medium (NM): induces normal differentiation in DMEM supplemented by 10% FBS, 2 mM glutamine and 1% penicillin/streptomycin.
- Define Normoxia Medium (DM): the starvation of 10% Fetal Bovine Serum (FBS) in this medium induces stringent conditions. DM is supplemented in two specific growth factors: EGF (Epidermal Growth Factors) and bFGF2 (basic Fibroblast Growth Factors) required for clonal expansion and the formation of glioma spheres which are composed of several thousand aggregated cells. DM composition consists in DMEM/F12 supplemented by 0.6% glucose, 1% sodium bicarbonate, 1% MEM non-essential amino acids, 5 mM HEPES, 9.6 µg/mL putrescine, 10 µg/mL ITSS, 0.063 µg/mL progesterone, 60 µg/mL N-acetyl-l-cysteine, 2 µg/mL heparin, 0.1 mg/mL penicillin/streptomycin, 50X B-27 supplement without vitamin A, 20 ng/mL EGF, 20 ng/mL bFG.
2.2. Primary GBM Cell Isolations, Culturing and Separation by Flow Cytometry
2.3. DEP Suspension Medium
2.4. Comparative Transcriptomic Analysis (mRNA Levels) of the Stemness Phenotype
2.5. Crossover Frequency Experiment
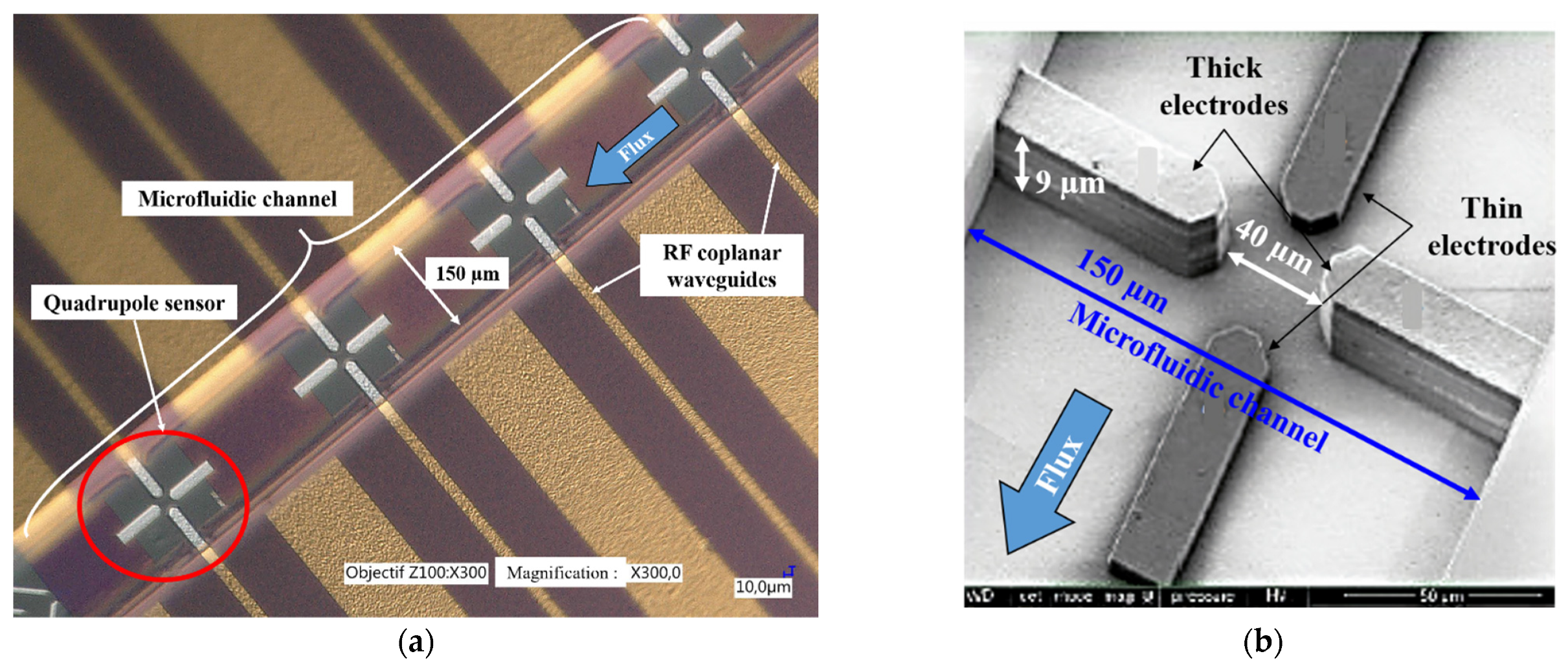
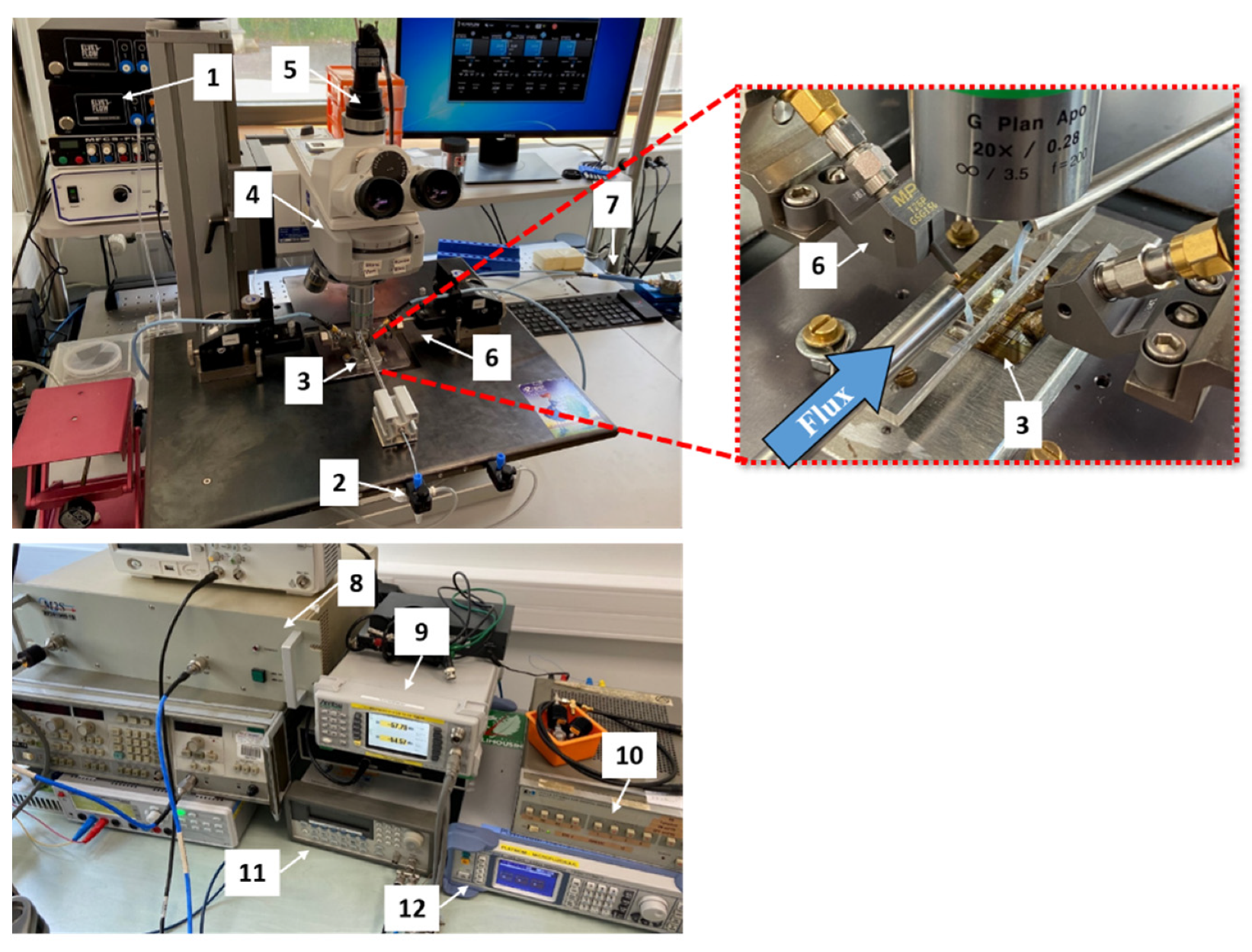
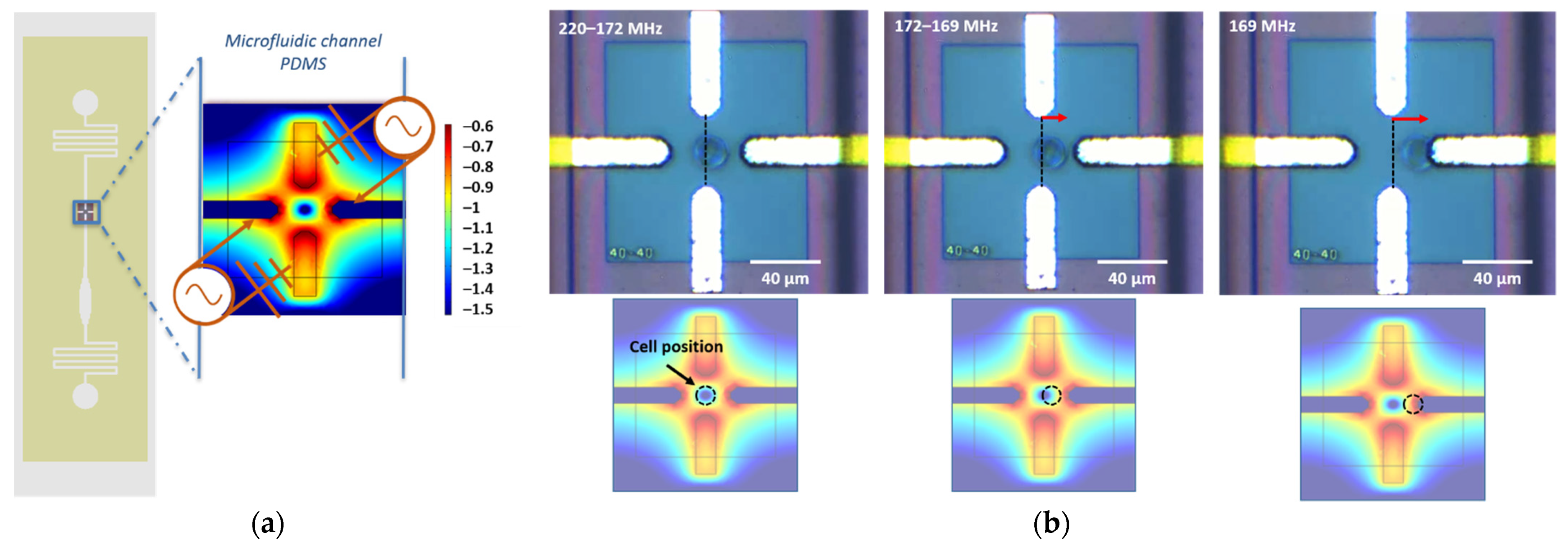
2.5.1. DEP Sensor Design and Experimental Setup
2.5.2. fx01 and fx02 Crossover Frequencies Measurements
2.6. Statistical Analysis
3. Results
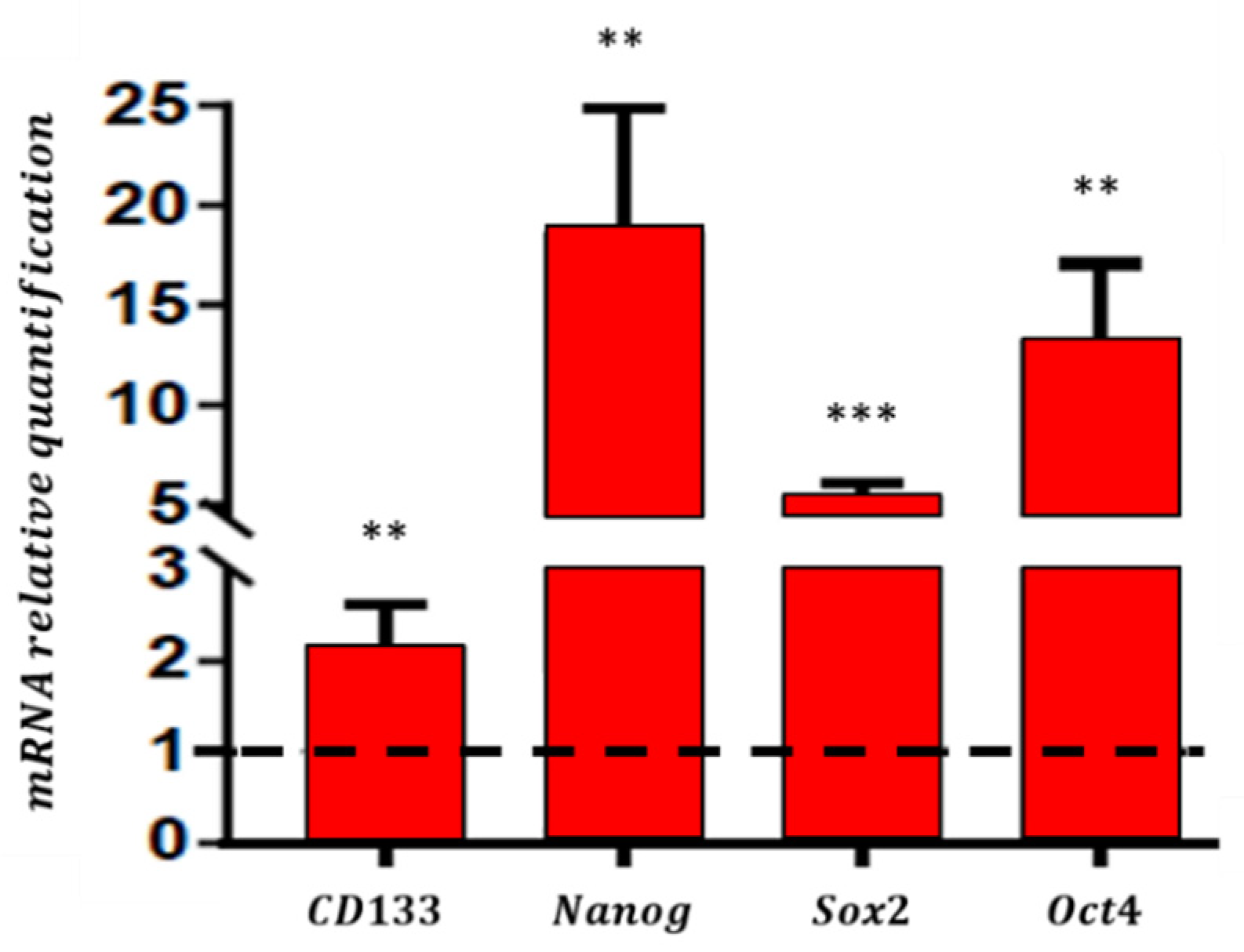
3.1. Enrichment of CSC in the Define Medium
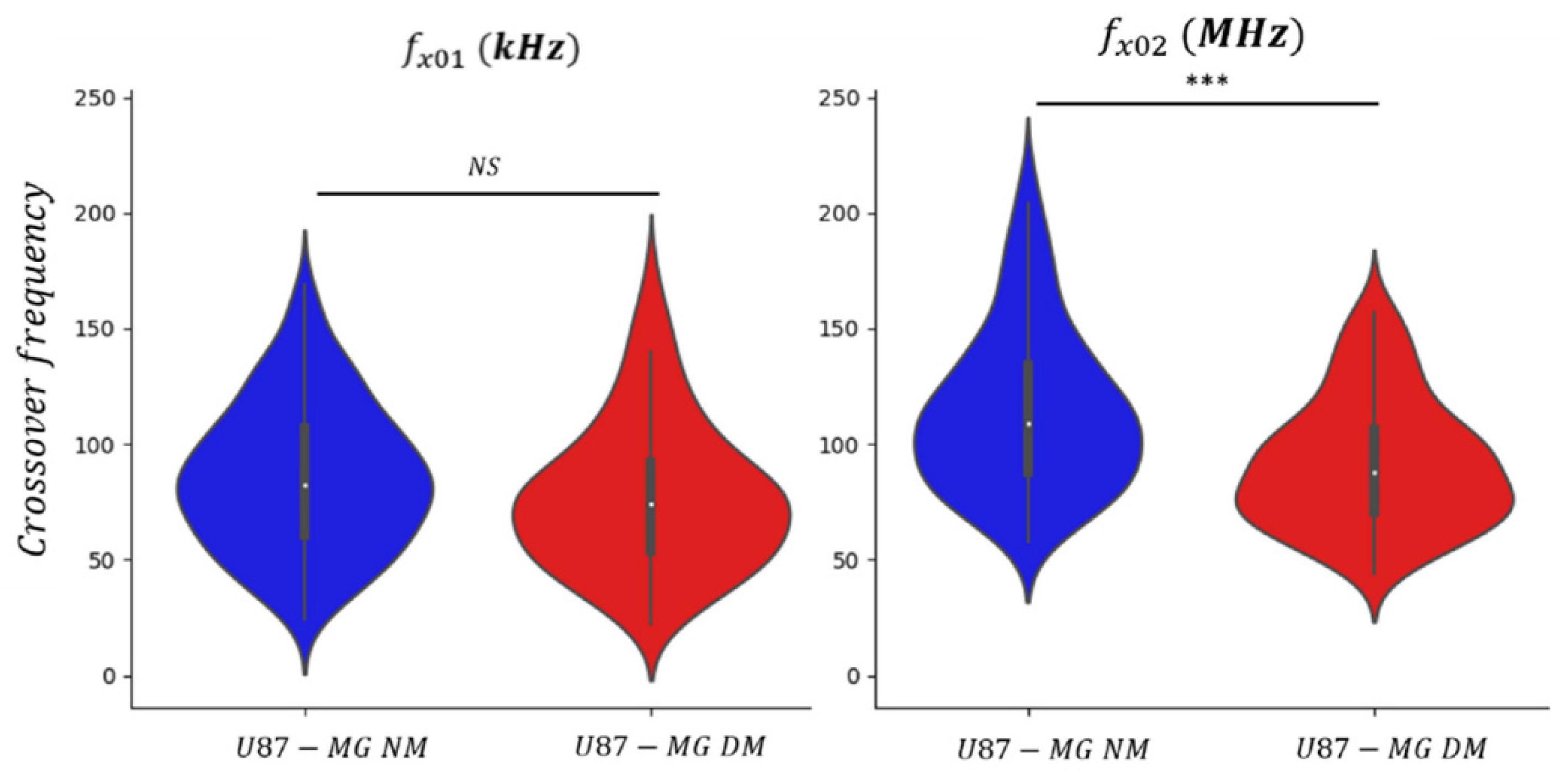
3.2. Dielectrophoretic Signatures fx01 and fx02 of U87-MG Cell Line
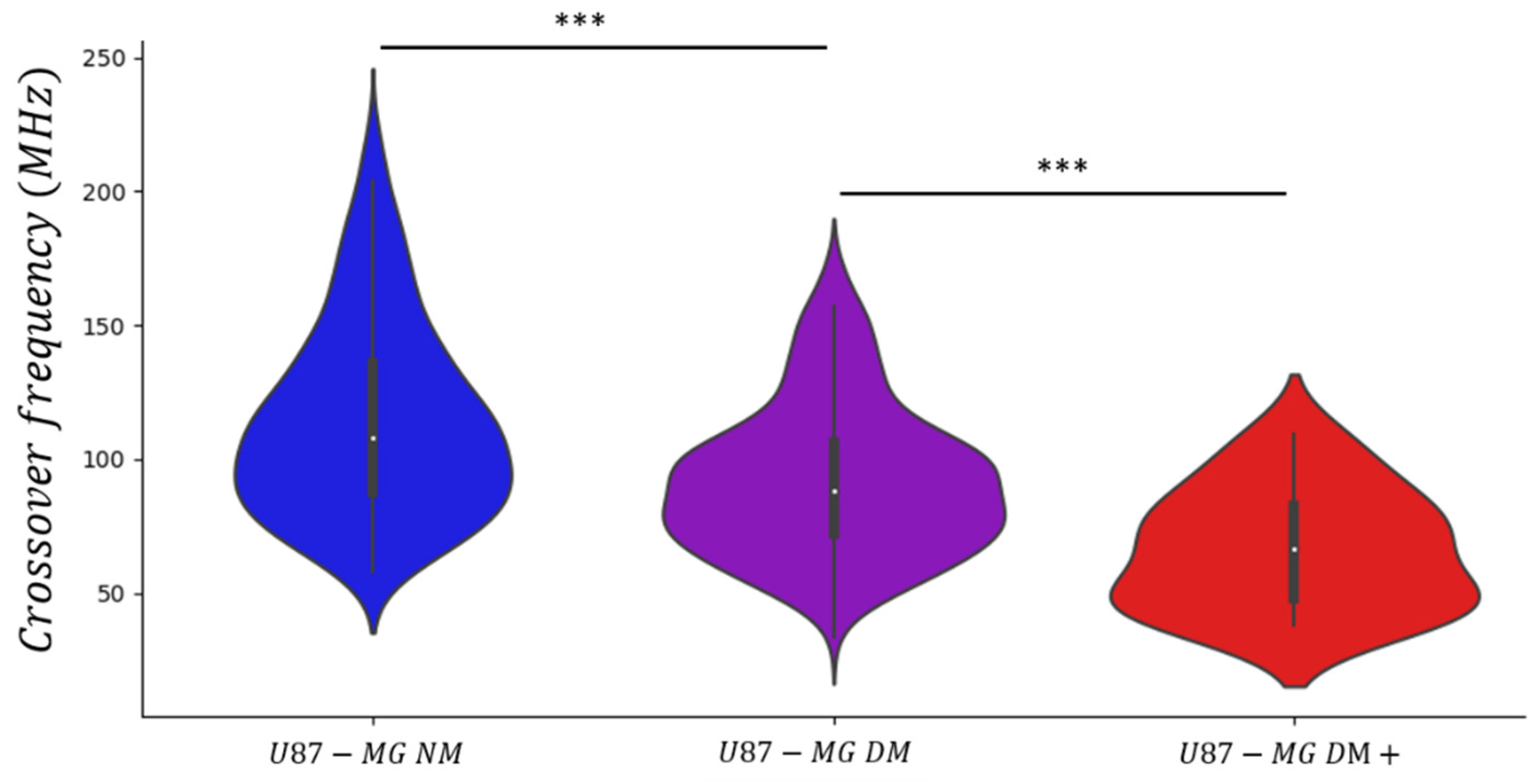
3.3. Kinetic Evolution of the Stemness Phenotype
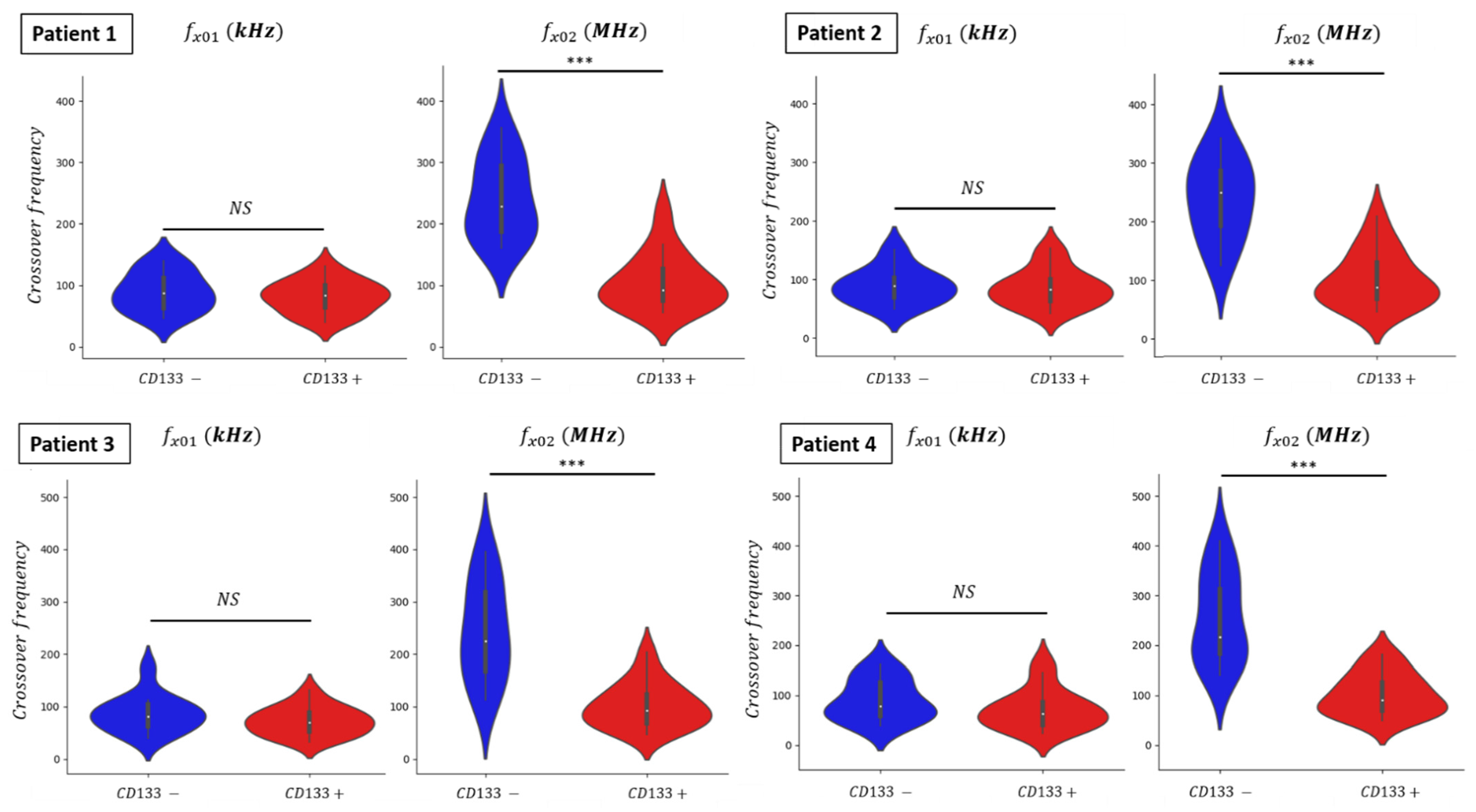
3.4. Dielectrophoretic Signatures fx01 and fx02 of GBM Primary Cultures
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Cheray, M.; Bégaud, G.; Deluche, E.; Nivet, A.; Battu, S.; Lalloué, F.; Verdier, M.; Bessette, B. Cancer Stem-Like Cells in Glioblastoma. Exon. Publ. 2017, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Stupp, R.; Mason, W.P.; Bent, M.V.D.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, H.S. Potential Improvement of Survival Statistics for Glioblastoma Multiforme (WHO IV). Surg. Neurol. Int. 2019, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Tabatabai, G.; Weller, M. Glioblastoma Stem Cells. Cell Tissue Res. 2011, 343, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.; Yu, T.-S.; McKay, R.M.; Burns, D.K.; Kernie, S.G.; Parada, L.F. A Restricted Cell Population Propagates Glioblastoma Growth after Chemotherapy. Nature 2012, 488, 522–526. [Google Scholar] [CrossRef] [Green Version]
- Gossett, D.R.; Weaver, W.M.; Mach, A.J.; Hur, S.C.; Tse, H.T.K.; Lee, W.; Amini, H.; Di Carlo, D. Label-Free Cell Separation and Sorting in Microfluidic Systems. Anal. Bioanal. Chem. 2010, 397, 3249–3267. [Google Scholar] [CrossRef] [Green Version]
- Pethig, R.R. Dielectrophoresis: Theory, Methodology and Biological Applications; John Wiley & Sons: Hoboken, NJ, USA, 2017; ISBN 978-1-118-67145-0. [Google Scholar]
- Cottet, J.; Fabregue, O.; Berger, C.; Buret, F.; Renaud, P.; Frénéa-Robin, M. MyDEP: A New Computational Tool for Dielectric Modeling of Particles and Cells. Biophys. J. 2018, 116, 12–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afshar, S.; Fazelkhah, A.; Braasch, K.; Salimi, E.; Butler, M.; Thomson, D.J.; Bridges, G.E. Full Beta-Dispersion Region Dielectric Spectra and Dielectric Models of Viable and Non-Viable CHO Cells. IEEE J. Electromagn. RF Microw. Med. Biol. 2020, 5, 70–77. [Google Scholar] [CrossRef]
- Fazelkhah, A.; Afshar, S.; Braasch, K.; Butler, M.; Salimi, E.; Bridges, G.; Thomson, D. Cytoplasmic conductivity as a marker for bioprocess monitoring: Study of Chinese hamster ovary cells under nutrient deprivation and reintroduction. Biotechnol. Bioeng. 2019, 116, 2896–2905. [Google Scholar] [CrossRef]
- Cottet, J.; Fabregue, O.; Berger, C.; Buret, F.; Renaud, P.; Frénéa-Robin, M. MyDEP: A New Computational Tool for Dielectric Modeling of Particles and Cells. Zenodo 2019. [Google Scholar] [CrossRef] [Green Version]
- Pethig, R.; Menachery, A.; Pells, S.; De Sousa, P. Dielectrophoresis: A Review of Applications for Stem Cell Research. J. Biomed. Biotechnol. 2010, 2010, 182581. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Yao, J.; Sugawara, M.; Takei, M. Numerical Study of Particle-Fluid Flow Under AC Electrokinetics in Elec-trode-Multilayered Microfluidic Device. IEEE Trans. Biomed. Eng. 2018, 66, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Ma, X.; Li, H.; Li, L.; Cheng, X.; Hwang, J.C.M. Validation of Clausius–Mossotti Function in Wideband Single-Cell Dielec-trophoresis. IEEE J. Electromagn. RF Microw. Med. Biol. 2019, 3, 127–133. [Google Scholar] [CrossRef]
- Alazzam, A.; Stiharu, I.; Bhat, R.; Meguerditchian, A.-N. Interdigitated Comb-like Electrodes for Continuous Separation of Malignant Cells from Blood Using Dielectrophoresis. Electrophoresis 2011, 32, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Park, J.-K. Microfluidic system for dielectrophoretic separation based on a trapezoidal electrode array. Lab Chip 2005, 5, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Vahey, M.D.; Voldman, J. An Equilibrium Method for Continuous-Flow Cell Sorting Using Dielectrophoresis. Anal. Chem. 2008, 80, 3135–3143. [Google Scholar] [CrossRef]
- Chung, C.; Pethig, R.; Smith, S.; Waterfall, M. Intracellular Potassium under Osmotic Stress Determines the Dielectrophoresis Cross-over Frequency of Murine Myeloma Cells in the MHz Range. Electrophoresis 2018, 39, 989–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, C.; Waterfall, M.; Pells, S.; Menachery, A.; Smith, S.; Pethig, R. Dielectrophoretic Characterisation of Mammalian Cells above 100 MHz. J. Electr. Bioimpedance 2011, 2, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Manczak, R.; Saada, S.; Tanori, M.; Casciati, A.; Dalmay, C.; Bessette, B.; Begaud, G.; Battu, S.; Blondy, P.; Jauberteau, M.O.; et al. High-Frequency Dielectrophoresis Characterization of Differentiated vs Undifferentiated Medulloblastoma Cells. In Proceedings of the 2018 EMF-Med 1st World Conference on Biomedical Applications of Electromagnetic Fields (EMF-Med); 2018; pp. 1–2. [Google Scholar] [CrossRef] [Green Version]
- Casciati, A.; Tanori, M.; Manczak, R.; Saada, S.; Tanno, B.; Giardullo, P.; Porcù, E.; Rampazzo, E.; Persano, L.; Viola, G.; et al. Human Medulloblastoma Cell Lines: Investigating on Cancer Stem Cell-Like Phenotype. Cancers 2020, 12, 226. [Google Scholar] [CrossRef] [Green Version]
- Pistollato, F.; Persano, L.; Della Puppa, A.; Rampazzo, E.; Basso, G. Isolation and Expansion of Regionally Defined Human Glio-blastoma Cells In Vitro. Curr. Protoc. Stem Cell Biol. 2011, 17, 3.4.1–3.4.10. [Google Scholar] [CrossRef]
- Pistollato, F.; Abbadi, S.; Rampazzo, E.; Persano, L.; Della Puppa, A.; Frasson, C.; Sarto, E.; Scienza, R.; D’Avella, D.; Basso, G. Intratumoral Hypoxic Gradient Drives Stem Cells Distribution and MGMT Expression in Glioblastoma. Stem Cells 2010, 28, 851–862. [Google Scholar] [CrossRef]
- Frasson, C.; Rampazzo, E.; Accordi, B.; Beggio, G.; Pistollato, F.; Basso, G.; Persano, L. Inhibition of PI3K Signalling Selectively Affects Medulloblastoma Cancer Stem Cells. BioMed Res. Int. 2015, 2015, 973912. [Google Scholar] [CrossRef] [Green Version]
- Hjeij, F.; Dalmay, C.; Bessaudou, A.; Blondy, P.; Pothier, A.; Bessette, B.; Bégaud, G.; Jauberteau, M.O.; Lalloué, F.; Kaynak, C.B.; et al. UHF Dielectrophoretic Handling of Individual Biological Cells Using BiCMOS Microfluidic RF-Sensors. In Proceedings of the 2016 46th European Microwave Conference (EuMC), London, UK, 4–6 October 2016; pp. 265–268. [Google Scholar] [CrossRef]
- Suslov, O.N.; Kukekov, V.G.; Ignatova, T.N.; Steindler, D.A. Neural stem cell heterogeneity demonstrated by molecular pheno-typing of clonal neurospheres. Proc. Natl. Acad. Sci. USA 2002, 99, 14506–14511. [Google Scholar] [CrossRef] [Green Version]
- Pethig, R.; Bressler, V.; Carswell-Crumpton, C.; Chen, Y.; Foster-Haje, L.; García-Ojeda, M.E.; Lee, R.S.; Lock, G.M.; Talary, M.S.; Tate, K.M. Dielectrophoretic Studies of the Activation of Human T Lymphocytes Using a Newly Developed Cell Pro-filing System. Electrophoresis 2002, 23, 2057–2063. [Google Scholar] [CrossRef]
- Li, Z. CD133: A Stem Cell Biomarker and Beyond. Exp. Hematol. Oncol. 2013, 2, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, F. CD133: A cancer stem cells marker, is used in colorectal cancers. World J. Gastroenterol. 2013, 19, 2603–2611. [Google Scholar] [CrossRef] [PubMed]
- Kemper, K.; Sprick, M.; De Bree, M.; Scopelliti, A.; Vermeulen, L.; Hoek, M.; Zeilstra, J.; Pals, S.T.; Mehmet, H.; Stassi, G.; et al. The AC133 Epitope, but not the CD133 Protein, Is Lost upon Cancer Stem Cell Differentiation. Cancer Res. 2010, 70, 719–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provent, T.; Mauvy, A.; Manczak, R.; Saada, S.; Dalmay, C.; Bessette, B.; Lalloue, F.; Pothier, A. A High Frequency Dielectrophoresis Cytometer for Continuous Flow Biological Cells Refinement. In Proceedings of the 2020 50th European Microwave Conference (EuMC), Utrecht, The Netherlands, 12–14 January 2021; pp. 921–924. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| Particle radius | 11.5 µm |
| Membrane thickness | 700 nm 1 |
| Intracellular relative permittivity | 50 |
| Intracellular conductivity | 0.5 S/m |
| Membrane relative permittivity | 1002 |
| Membrane conductivity | 1.43 × 104 S/m 2 |
| Medium relative permittivity | 78 |
| Medium conductivity | 0.02 S/m |
| Cell Culture Conditions | Crossover Frequency | Number of Cells Measured | Median Value | SD |
|---|---|---|---|---|
| Normal Medium (NN) | fx01 | 139 | 82 kHz | 31.5 kHz |
| Define medium (DN) | 134 | 74 kHz | 32.1 kHz | |
| Normal Medium (NN) | fx02 | 139 | 109 MHz | 35.2 MHz |
| Define medium (DN) | 134 | 88 MHz | 27.9 MHz |
| Culture Condition | Median Value | SD |
|---|---|---|
| Normal Medium (NM) | 108 MHz | 36.2 MHz |
| Define Medium (DM) | 88 MHz | 27.9 MHz |
| Define Medium (DM+) | 67 MHz | 22.1 MHz |
| Cell Population | Crossover Frequency | Median Value | |
|---|---|---|---|
| Patient 1 | CD133− | fx01 | 88 kHz |
| CD133+ | 83 kHz | ||
| CD133− | fx02 | 229 MHz | |
| CD133+ | 92 MHz | ||
| Patient 2 | CD133− | fx01 | 89 kHz |
| CD133+ | 83 kHz | ||
| CD133− | fx02 | 248 MHz | |
| CD133+ | 86 MHz | ||
| Patient 3 | CD133− | fx01 | 81 kHz |
| CD133+ | 70 kHz | ||
| CD133− | fx02 | 225 MHz | |
| CD133+ | 92 MHz | ||
| Patient 4 | CD133− | fx01 | 78 kHz |
| CD133+ | 63 kHz | ||
| CD133− | fx02 | 216 MHz | |
| CD133+ | 91 MHz |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambert, E.; Manczak, R.; Barthout, E.; Saada, S.; Porcù, E.; Maule, F.; Bessette, B.; Viola, G.; Persano, L.; Dalmay, C.; et al. Microfluidic Lab-on-a-Chip Based on UHF-Dielectrophoresis for Stemness Phenotype Characterization and Discrimination among Glioblastoma Cells. Biosensors 2021, 11, 388. https://doi.org/10.3390/bios11100388
Lambert E, Manczak R, Barthout E, Saada S, Porcù E, Maule F, Bessette B, Viola G, Persano L, Dalmay C, et al. Microfluidic Lab-on-a-Chip Based on UHF-Dielectrophoresis for Stemness Phenotype Characterization and Discrimination among Glioblastoma Cells. Biosensors. 2021; 11(10):388. https://doi.org/10.3390/bios11100388
Chicago/Turabian StyleLambert, Elisa, Rémi Manczak, Elodie Barthout, Sofiane Saada, Elena Porcù, Francesca Maule, Barbara Bessette, Giampietro Viola, Luca Persano, Claire Dalmay, and et al. 2021. "Microfluidic Lab-on-a-Chip Based on UHF-Dielectrophoresis for Stemness Phenotype Characterization and Discrimination among Glioblastoma Cells" Biosensors 11, no. 10: 388. https://doi.org/10.3390/bios11100388
APA StyleLambert, E., Manczak, R., Barthout, E., Saada, S., Porcù, E., Maule, F., Bessette, B., Viola, G., Persano, L., Dalmay, C., Lalloué, F., & Pothier, A. (2021). Microfluidic Lab-on-a-Chip Based on UHF-Dielectrophoresis for Stemness Phenotype Characterization and Discrimination among Glioblastoma Cells. Biosensors, 11(10), 388. https://doi.org/10.3390/bios11100388