Multisite Simultaneous Neural Recording of Motor Pathway in Free-Moving Rats

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
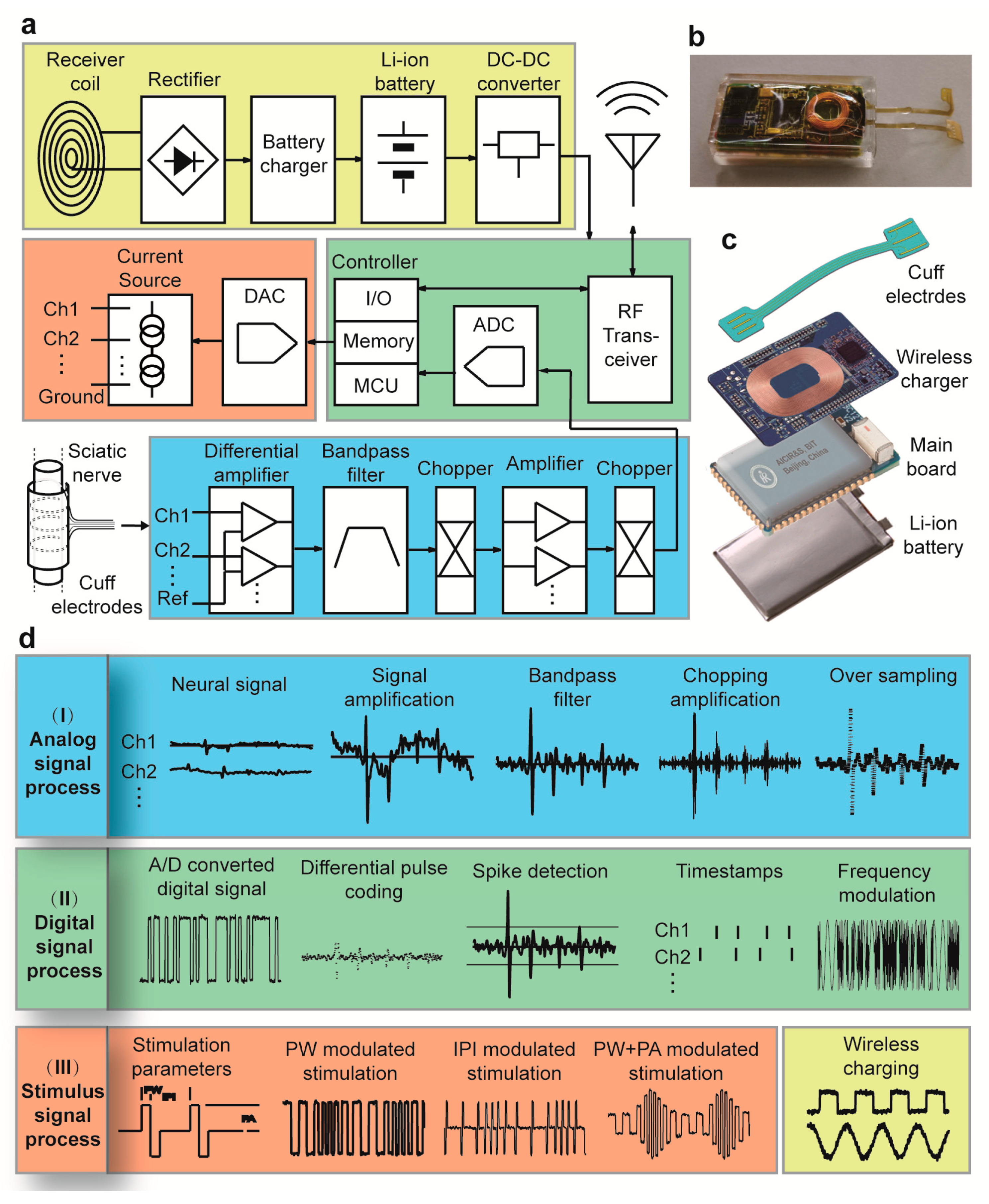
2.2. Design and Function of the Transmitter Module
2.3. Implantation of Electrodes
2.3.1. Motor Cortex Electrodes
2.3.2. Spinal Cord Recording
2.3.3. Sciatic Nerve Interface
2.3.4. EMG Microwires
2.4. Recording during Tasks
2.4.1. Bipedal Treadmill Locomotion
2.4.2. Wheel Running
2.4.3. Open Field Walking
3. Results
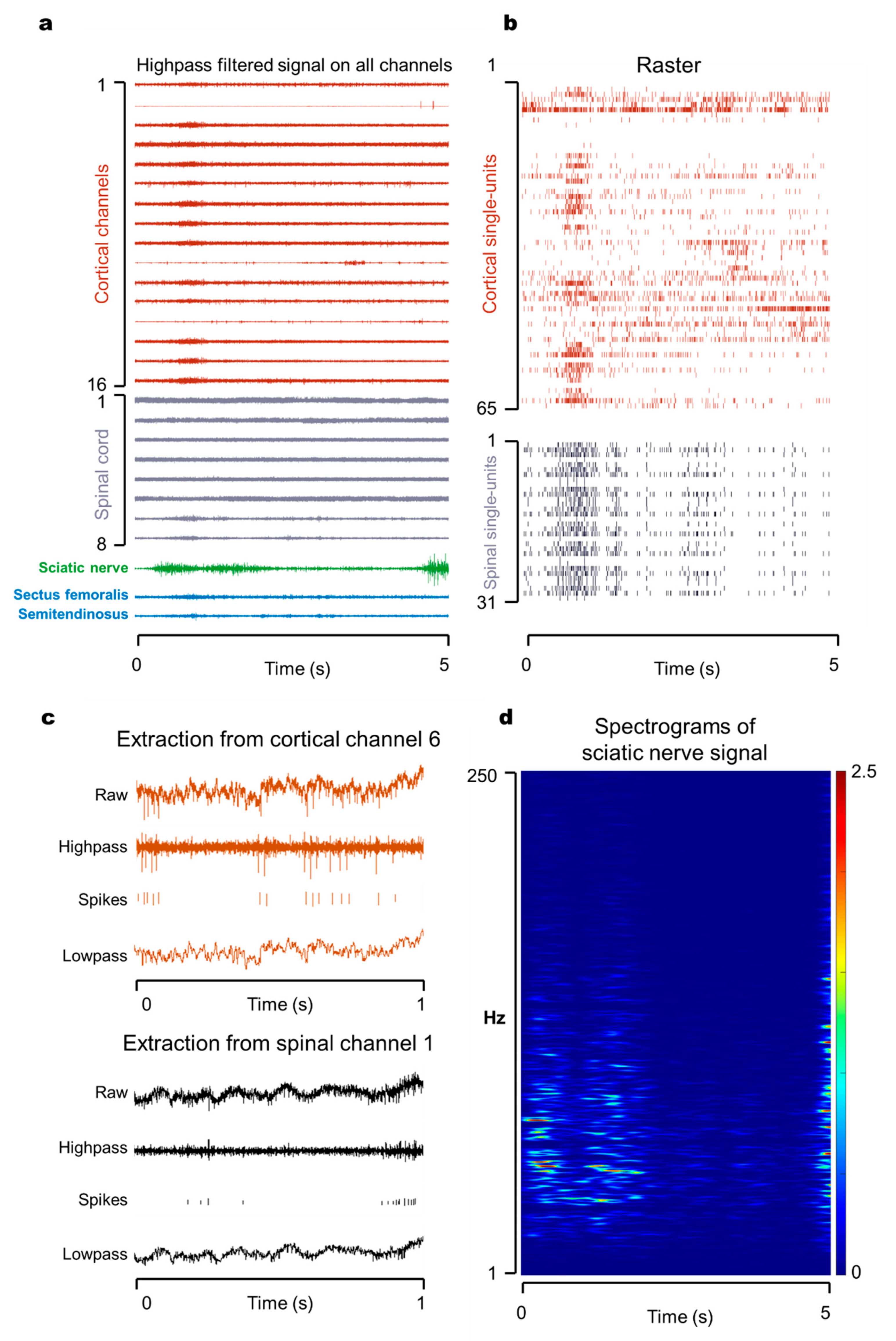
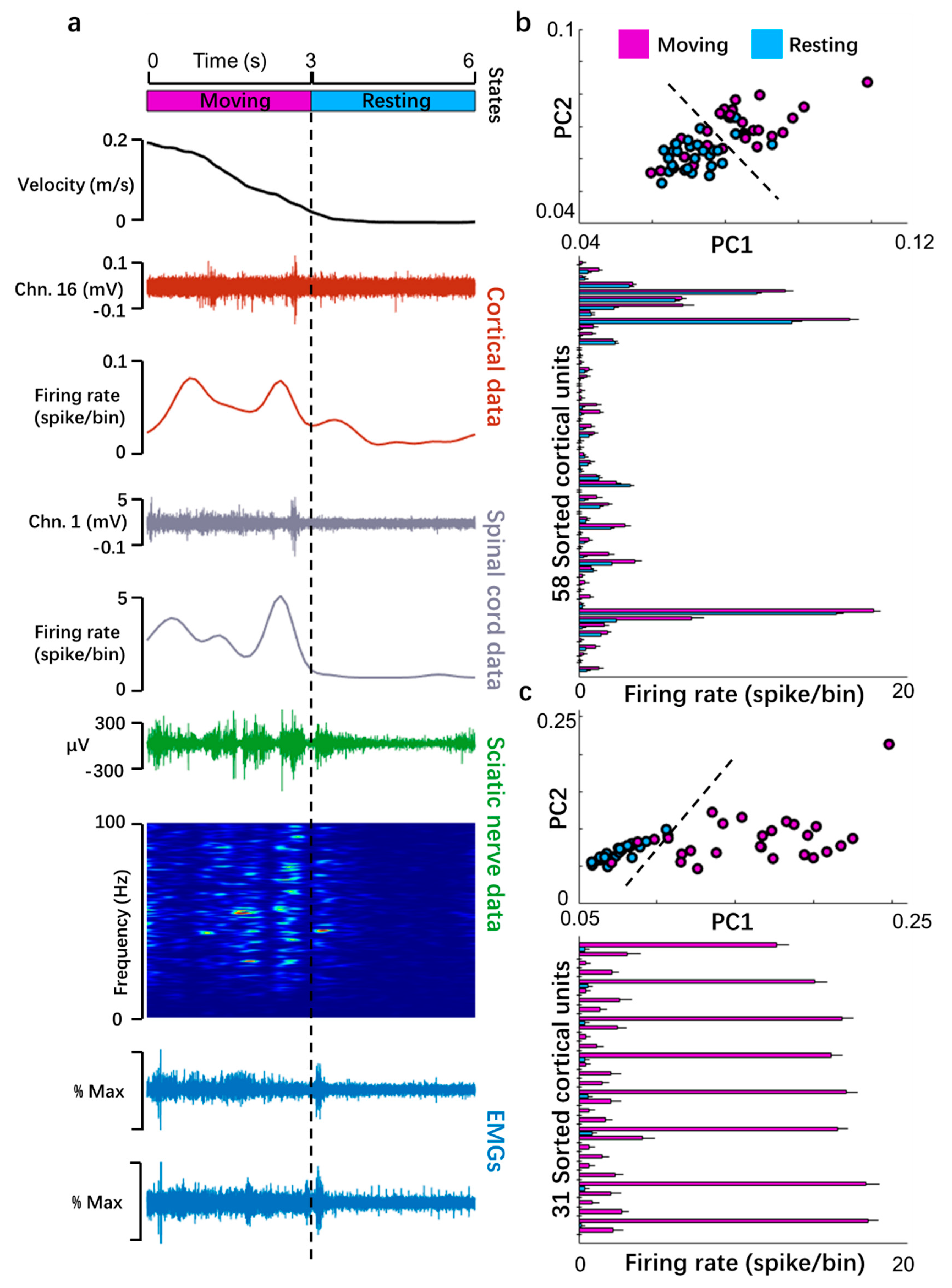
3.1. Multisite Simultaneous Recording in Motor Pathway
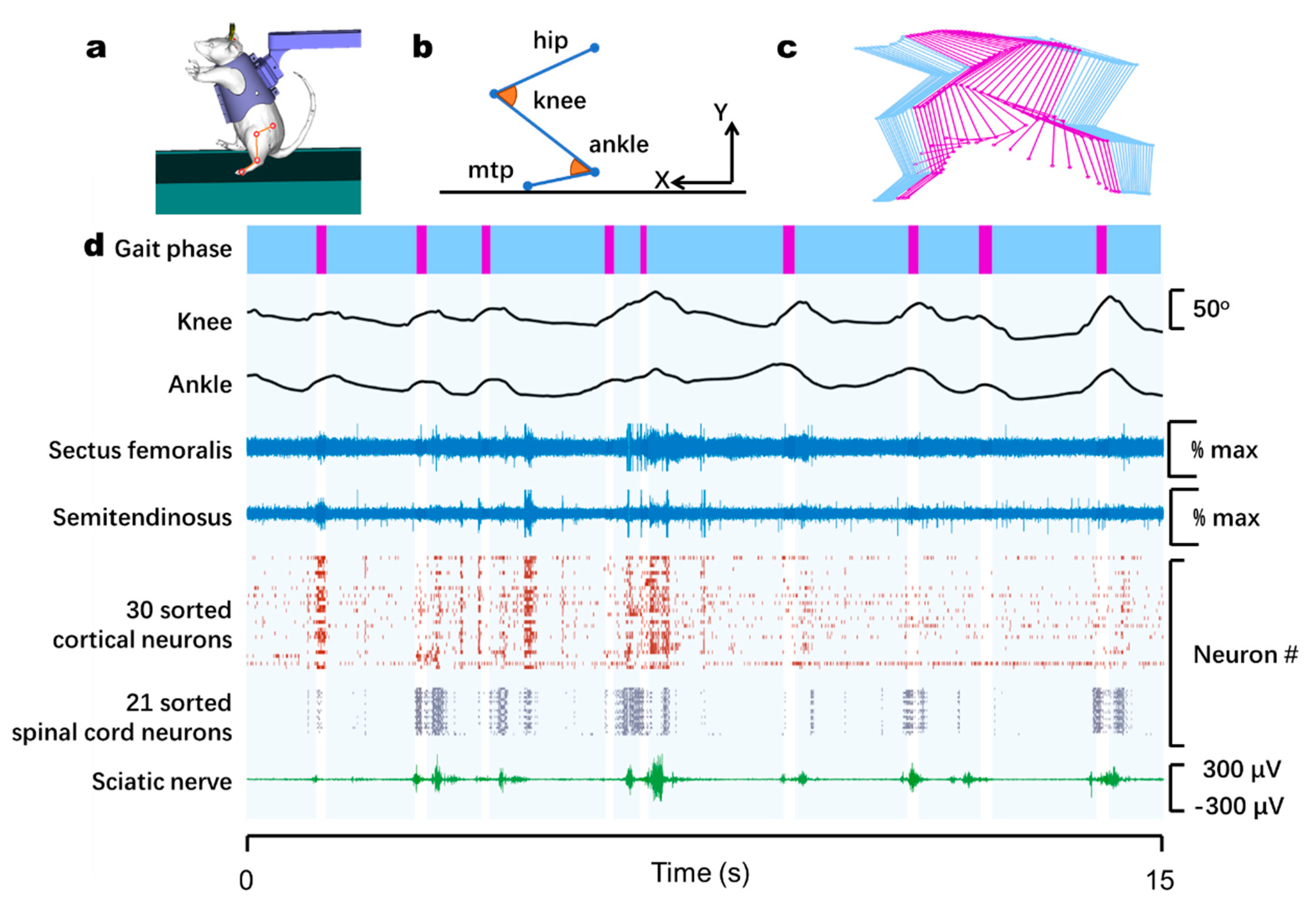
3.2. Monitoring of Neural and Muscular Activity during Bipedal Walking on a Treadmill
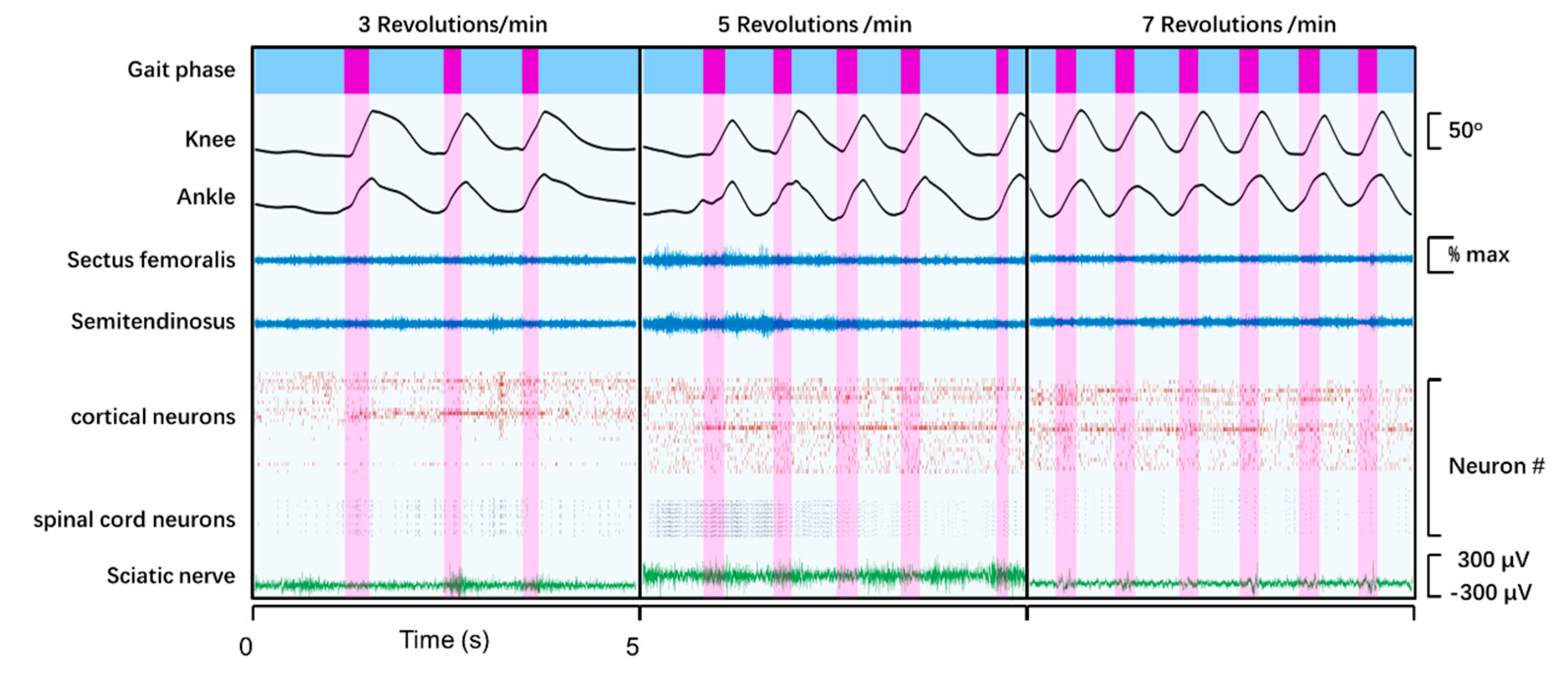
3.3. Recording of Neural and Muscular Activity during Quadrupedal Wheel Running
3.4. Investigation of Signal Transitions in Open Field Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prochazka, A. Neurophysiology and neural engineering: A review. J. Neurophysiol. 2017, 118, 1292–1309. [Google Scholar] [CrossRef]
- Krook-Magnuson, E.; Gelinas, J.N.; Soltesz, I.; Buzsaki, G. Neuroelectronics and Biooptics: Closed-Loop Technologies in Neurological Disorders. JAMA Neurol. 2015, 72, 823–829. [Google Scholar] [CrossRef]
- Beudel, M.; Brown, P. Adaptive deep brain stimulation in Parkinson’s disease. Parkinsonism Relat Disord 2016, 22 (Suppl. 1), S123–S126. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.R.; Lieber, C.M. Precision electronic medicine in the brain. Nat. Biotechnol. 2019, 37, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, D.S.; Guo, L.; Gulati, T.; Davidson, G.; Hishinuma, A.K.; Won, S.J.; Knight, R.T.; Chang, E.F.; Swanson, R.A.; Ganguly, K. Low-frequency cortical activity is a neuromodulatory target that tracks recovery after stroke. Nat. Med. 2018, 24, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, U.; Birbaumer, N.; Curado, M.R. Brain-machine interface (BMI) in paralysis. Ann. Phys. Rehabil. Med. 2015, 58, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; Liu, C.Y.; Apuzzo, M.L. A primer on brain-machine interfaces, concepts, and technology: A key element in the future of functional neurorestoration. World Neurosurg. 2013, 79, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Aflalo, T.; Kellis, S.; Klaes, C.; Lee, B.; Shi, Y.; Pejsa, K.; Shanfield, K.; Hayes-Jackson, S.; Aisen, M.; Heck, C.; et al. Neurophysiology. Decoding motor imagery from the posterior parietal cortex of a tetraplegic human. Science 2015, 348, 906–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moses, D.A.; Metzger, S.L.; Liu, J.R.; Anumanchipalli, G.K.; Makin, J.G.; Sun, P.F.; Chartier, J.; Dougherty, M.E.; Liu, P.M.; Abrams, G.M.; et al. Neuroprosthesis for Decoding Speech in a Paralyzed Person with Anarthria. N. Engl. J. Med. 2021, 385, 217–227. [Google Scholar] [CrossRef]
- Simeral, J.D.; Hosman, T.; Saab, J.; Flesher, S.N.; Vilela, M.; Franco, B.; Kelemen, J.N.; Brandman, D.M.; Ciancibello, J.G.; Rezaii, P.G.; et al. Home Use of a Percutaneous Wireless Intracortical Brain-Computer Interface by Individuals with Tetraplegia. IEEE Trans. Biomed. Eng. 2021, 68, 2313–2325. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, J.V.; Wong, Y.T. Neurobionics and the brain-computer interface: Current applications and future horizons. Med. J. Aust. 2017, 206, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Lemke, S.M.; Ramanathan, D.S.; Guo, L.; Won, S.J.; Ganguly, K. Emergent modular neural control drives coordinated motor actions. Nat. Neurosci. 2019, 22, 1122–1131. [Google Scholar] [CrossRef]
- Ejaz, N.; Hamada, M.; Diedrichsen, J. Hand use predicts the structure of representations in sensorimotor cortex. Nat. Neurosci. 2015, 18, 1034–1040. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.L.; Carta, S.; Soldado-Magraner, J.; Schneider, B.L.; Helmchen, F. Behaviour-dependent recruitment of long-range projection neurons in somatosensory cortex. Nature 2013, 499, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Rangroo Thrane, V.; Thrane, A.S.; Wang, F.; Cotrina, M.L.; Smith, N.A.; Chen, M.; Xu, Q.; Kang, N.; Fujita, T.; Nagelhus, E.A.; et al. Ammonia triggers neuronal disinhibition and seizures by impairing astrocyte potassium buffering. Nat. Med. 2013, 19, 1643–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchgessner, M.A.; Franklin, A.D.; Callaway, E.M. Context-dependent and dynamic functional influence of corticothalamic pathways to first- and higher-order visual thalamus. Proc. Natl. Acad. Sci. USA 2020, 117, 13066–13077. [Google Scholar] [CrossRef]
- Chang, C.H.; Grace, A.A. Inhibitory Modulation of Orbitofrontal Cortex on Medial Prefrontal Cortex-Amygdala Information Flow. Cereb. Cortex 2018, 28, 1–8. [Google Scholar] [CrossRef]
- Nurmikko, A. Challenges for Large-Scale Cortical Interfaces. Neuron 2020, 108, 259–269. [Google Scholar] [CrossRef]
- Luan, L.; Robinson, J.T.; Aazhang, B.; Chi, T.; Yang, K.; Li, X.; Rathore, H.; Singer, A.; Yellapantula, S.; Fan, Y.; et al. Recent Advances in Electrical Neural Interface Engineering: Minimal Invasiveness, Longevity, and Scalability. Neuron 2020, 108, 302–321. [Google Scholar] [CrossRef] [PubMed]
- Obidin, N.; Tasnim, F.; Dagdeviren, C. The Future of Neuroimplantable Devices: A Materials Science and Regulatory Perspective. Adv. Mater. 2020, 32, e1901482. [Google Scholar] [CrossRef] [PubMed]
- Lang, Y.; Du, P.; Shin, H.C. Encoding-based brain-computer interface controlled by non-motor area of rat brain. Sci. China Life Sci. 2011, 54, 841–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, J.J.; Steinmetz, N.A.; Siegle, J.H.; Denman, D.J.; Bauza, M.; Barbarits, B.; Lee, A.K.; Anastassiou, C.A.; Andrei, A.; Aydın, Ç.; et al. Fully integrated silicon probes for high-density recording of neural activity. Nature 2017, 551, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Colachis, S.C.t.; Dunlap, C.F.; Annetta, N.V.; Tamrakar, S.M.; Bockbrader, M.A.; Friedenberg, D.A. Long-term intracortical microelectrode array performance in a human: A 5 year retrospective analysis. J. Neural. Eng. 2021, 18, 0460d7. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, M.; Sharma, D.; Ross, D.; Zhao, F. A Critical Review of Microelectrode Arrays and Strategies for Improving Neural Interfaces. Adv. Healthc. Mater. 2019, 8, e1900558. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Borton, D.A.; Komar, J.; Agha, N.; Lu, Y.; Li, H.; Laurens, J.; Lang, Y.; Li, Q.; Bull, C.; et al. Wireless neurosensor for full-spectrum electrophysiology recordings during free behavior. Neuron 2014, 84, 1170–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kale, R.P.; Kouzani, A.Z.; Walder, K.; Berk, M.; Tye, S.J. Evolution of optogenetic microdevices. Neurophotonics 2015, 2, 031206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warden, M.R.; Cardin, J.A.; Deisseroth, K. Optical neural interfaces. Annu. Rev. Biomed. Eng. 2014, 16, 103–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, S.B.; Ribeiro, J.F.; Silva, A.F.; Costa, R.M.; Correia, J.H. Design and manufacturing challenges of optogenetic neural interfaces: A review. J. Neural Eng. 2017, 14, 041001. [Google Scholar] [CrossRef]
- Rudmann, L.; Alt, M.T.; Ashouri Vajari, D.; Stieglitz, T. Integrated optoelectronic microprobes. Curr. Opin. Neurobiol. 2018, 50, 72–82. [Google Scholar] [CrossRef]
- Li, B.; Liu, S.; Hu, D.; Li, G.; Tang, R.; Song, D.; Lang, Y.; He, J. Electrocortical activity in freely walking rats varies with environmental conditions. Brain Res. 2021, 1751, 147188. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, B.; Lv, X.; Liu, S.; Liu, Y.; Tang, R.; Lang, Y.; Huang, Q.; He, J. Low-Intensity Focused Ultrasound-Mediated Attenuation of Acute Seizure Activity Based on EEG Brain Functional Connectivity. Brain Sci. 2021, 11, 711. [Google Scholar] [CrossRef] [PubMed]
- Uta, D.; Oti, T.; Sakamoto, T.; Sakamoto, H. In Vivo Electrophysiology of Peptidergic Neurons in Deep Layers of the Lumbar Spinal Cord after Optogenetic Stimulation of Hypothalamic Paraventricular Oxytocin Neurons in Rats. Int. J. Mol. Sci. 2021, 22, 3400. [Google Scholar] [CrossRef] [PubMed]
- Inada, Y.; Funai, Y.; Yamasaki, H.; Mori, T.; Nishikawa, K. Effects of sevoflurane and desflurane on the nociceptive responses of substantia gelatinosa neurons in the rat spinal cord dorsal horn: An in vivo patch-clamp analysis. Mol. Pain 2020, 16, 1744806920903149. [Google Scholar] [CrossRef] [Green Version]
- Turner, K.; O’Connell, P.R.; Jones, J.F.X. The projection of anorectal afferents to spinal cord and effect of sacral neuromodulation on dorsal horn neurons which receive such input in the rat. Neurogastroenterol. Motil. 2019, 31, e13536. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T.; Merrill, A.W.; Carstens, M.I.; Carstens, E. Activation of superficial dorsal horn neurons in the mouse by a PAR-2 agonist and 5-HT: Potential role in itch. J. Neurosci. 2009, 29, 6691–6699. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Yamada, A.; Kim, W.; Kim, S.K.; Furue, H. Noradrenergic inhibition of spinal hyperexcitation elicited by cutaneous cold stimuli in rats with oxaliplatin-induced allodynia: Electrophysiological and behavioral assessments. J. Physiol. Sci. 2017, 67, 431–438. [Google Scholar] [CrossRef]
- van den Hoogen, N.J.; Patijn, J.; Tibboel, D.; Joosten, B.A.; Fitzgerald, M.; Kwok, C.H.T. Repeated touch and needle-prick stimulation in the neonatal period increases the baseline mechanical sensitivity and postinjury hypersensitivity of adult spinal sensory neurons. Pain 2018, 159, 1166–1175. [Google Scholar] [CrossRef]
- Choi, S.; Chae, H.K.; Heo, H.; Hahm, D.H.; Kim, W.; Kim, S.K. Analgesic Effect of Melittin on Oxaliplatin-Induced Peripheral Neuropathy in Rats. Toxins (Basel) 2019, 11, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, T.M.; Lee, D.; Bradley, K.; McMahon, S.B. Methodology for quantifying excitability of identified projection neurons in the dorsal horn of the spinal cord, specifically to study spinal cord stimulation paradigms. J. Neurosci. Methods 2020, 330, 108479. [Google Scholar] [CrossRef]
- Lu, C.; Park, S.; Richner, T.J.; Derry, A.; Brown, I.; Hou, C.; Rao, S.; Kang, J.; Mortiz, C.T.; Fink, Y.; et al. Flexible and stretchable nanowire-coated fibers for optoelectronic probing of spinal cord circuits. Sci. Adv. 2017, 3, e1600955. [Google Scholar] [CrossRef] [Green Version]
- Minev, I.R.; Musienko, P.; Hirsch, A.; Barraud, Q.; Wenger, N.; Moraud, E.M.; Gandar, J.; Capogrosso, M.; Milekovic, T.; Asboth, L.; et al. Biomaterials. Electronic dura mater for long-term multimodal neural interfaces. Science 2015, 347, 159–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minev, I.R.; Chew, D.J.; Delivopoulos, E.; Fawcett, J.W.; Lacour, S.P. High sensitivity recording of afferent nerve activity using ultra-compliant microchannel electrodes: An acute in vivo validation. J. Neural Eng. 2012, 9, 026005. [Google Scholar] [CrossRef] [PubMed]
- Prins, N.W.; Mylavarapu, R.; Shoup, A.M.; Debnath, S.; Prasad, A. Spinal cord neural interfacing in common marmosets (Callithrix jacchus). J. Neural Eng. 2020, 17, 016031. [Google Scholar] [CrossRef]
- Hong, K.S.; Aziz, N.; Ghafoor, U. Motor-commands decoding using peripheral nerve signals: A review. J. Neural Eng. 2018, 15, 031004. [Google Scholar] [CrossRef] [Green Version]
- Rongyu Tang, X.L.; Sun, W.; Dai, C.; He, J. A fully implantable wireless neural interface for simultaneous recording from multiple sites of peripheral nerves in free moving animal. In Proceedings of the 2017 IEEE International Conference on Cyborg and Bionic Systems (CBS), Beijing, China, 17–19 October 2017. [Google Scholar]
- Shahriari, D.; Rosenfeld, D.; Anikeeva, P. Emerging Frontier of Peripheral Nerve and Organ Interfaces. Neuron 2020, 108, 270–285. [Google Scholar] [CrossRef] [PubMed]
- Tyler, D.J.; Durand, D.M. Functionally selective peripheral nerve stimulation with a flat interface nerve electrode. IEEE Trans. Neural Syst. Rehabil. Eng. 2002, 10, 294–303. [Google Scholar] [CrossRef]
- Yoshida, K.; Horch, K. Selective stimulation of peripheral nerve fibers using dual intrafascicular electrodes. IEEE Trans. Biomed. Eng. 1993, 40, 492–494. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, A.; Tahilramani, M.; Bentley, J.T.; Gore, R.K.; Millard, D.C.; Mukhatyar, V.J.; Joseph, A.; Haque, A.S.; Stanley, G.B.; English, A.W.; et al. Microchannel-based regenerative scaffold for chronic peripheral nerve interfacing in amputees. Biomaterials 2015, 41, 151–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gore, R.K.; Choi, Y.; Bellamkonda, R.; English, A. Functional recordings from awake, behaving rodents through a microchannel based regenerative neural interface. J. Neural Eng. 2015, 12, 016017. [Google Scholar] [CrossRef] [Green Version]
- Musick, K.M.; Rigosa, J.; Narasimhan, S.; Wurth, S.; Capogrosso, M.; Chew, D.J.; Fawcett, J.W.; Micera, S.; Lacour, S.P. Chronic multichannel neural recordings from soft regenerative microchannel electrodes during gait. Sci. Rep. 2015, 5, 14363. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, N.; Cao, Y.; Wang, F.; Wang, P.; Ma, Y.; Lu, B.; Hou, G.; Fang, Z.; Liang, Z.; et al. Climbing-inspired twining electrodes using shape memory for peripheral nerve stimulation and recording. Sci. Adv. 2019, 5, eaaw1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, A.K.; Truccolo, W.; Vargas-Irwin, C.E.; Donoghue, J.P. Decoding 3D reach and grasp from hybrid signals in motor and premotor cortices: Spikes, multiunit activity, and local field potentials. J. Neurophysiol. 2012, 107, 1337–1355. [Google Scholar] [CrossRef] [Green Version]
- Michaels, J.A.; Scherberger, H. Population coding of grasp and laterality-related information in the macaque fronto-parietal network. Sci. Rep. 2018, 8, 1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zippo, A.G.; Storchi, R.; Nencini, S.; Caramenti, G.C.; Valente, M.; Biella, G.E. Neuronal functional connection graphs among multiple areas of the rat somatosensory system during spontaneous and evoked activities. PLoS Comput. Biol. 2013, 9, e1003104. [Google Scholar] [CrossRef] [PubMed]
- van den Brand, R.; Heutschi, J.; Barraud, Q.; DiGiovanna, J.; Bartholdi, K.; Huerlimann, M.; Friedli, L.; Vollenweider, I.; Moraud, E.M.; Duis, S.; et al. Restoring voluntary control of locomotion after paralyzing spinal cord injury. Science 2012, 336, 1182–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassar, I.R.; Yu, C.; Sambangi, J.; Lee, C.D.; Whalen, J.J., 3rd; Petrossians, A.; Grill, W.M. Electrodeposited platinum-iridium coating improves in vivo recording performance of chronically implanted microelectrode arrays. Biomaterials 2019, 205, 120–132. [Google Scholar] [CrossRef]
- Citi, L.; Carpaneto, J.; Yoshida, K.; Hoffmann, K.P.; Koch, K.P.; Dario, P.; Micera, S. On the use of wavelet denoising and spike sorting techniques to process electroneurographic signals recorded using intraneural electrodes. J. Neurosci. Methods 2008, 172, 294–302. [Google Scholar] [CrossRef] [Green Version]
- Bonizzato, M.; Pidpruzhnykova, G.; DiGiovanna, J.; Shkorbatova, P.; Pavlova, N.; Micera, S.; Courtine, G. Brain-controlled modulation of spinal circuits improves recovery from spinal cord injury. Nat. Commun. 2018, 9, 3015. [Google Scholar] [CrossRef] [PubMed]
- Mian, S.Y.; Honey, J.R.; Carnicer-Lombarte, A.; Barone, D.G. Large Animal Studies to Reduce the Foreign Body Reaction in Brain-Computer Interfaces: A Systematic Review. Biosensors 2021, 11, 275. [Google Scholar] [CrossRef] [PubMed]
- Gulino, M.; Kim, D.; Pane, S.; Santos, S.D.; Pego, A.P. Tissue Response to Neural Implants: The Use of Model Systems Toward New Design Solutions of Implantable Microelectrodes. Front. Neurosci. 2019, 13, 689. [Google Scholar] [CrossRef]
- Black, B.J.; Kanneganti, A.; Joshi-Imre, A.; Rihani, R.; Chakraborty, B.; Abbott, J.; Pancrazio, J.J.; Cogan, S.F. Chronic recording and electrochemical performance of Utah microelectrode arrays implanted in rat motor cortex. J. Neurophysiol. 2018, 120, 2083–2090. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, Y.; Tang, R.; Liu, Y.; Xi, P.; Liu, H.; Quan, Z.; Song, D.; Lv, X.; Huang, Q.; He, J. Multisite Simultaneous Neural Recording of Motor Pathway in Free-Moving Rats. Biosensors 2021, 11, 503. https://doi.org/10.3390/bios11120503
Lang Y, Tang R, Liu Y, Xi P, Liu H, Quan Z, Song D, Lv X, Huang Q, He J. Multisite Simultaneous Neural Recording of Motor Pathway in Free-Moving Rats. Biosensors. 2021; 11(12):503. https://doi.org/10.3390/bios11120503
Chicago/Turabian StyleLang, Yiran, Rongyu Tang, Yafei Liu, Pengcheng Xi, Honghao Liu, Zhenzhen Quan, Da Song, Xiaodong Lv, Qiang Huang, and Jiping He. 2021. "Multisite Simultaneous Neural Recording of Motor Pathway in Free-Moving Rats" Biosensors 11, no. 12: 503. https://doi.org/10.3390/bios11120503
APA StyleLang, Y., Tang, R., Liu, Y., Xi, P., Liu, H., Quan, Z., Song, D., Lv, X., Huang, Q., & He, J. (2021). Multisite Simultaneous Neural Recording of Motor Pathway in Free-Moving Rats. Biosensors, 11(12), 503. https://doi.org/10.3390/bios11120503