Nucleic Acid-Based Sensing Techniques for Diagnostics and Surveillance of Influenza



Abstract
:1. Introduction
2. Amplification-Based Techniques
2.1. Thermal Cycling Amplification Methods
2.1.1. Real-Time Reverse Transcriptase PCR
2.1.2. Microfluidic Automation of RT-PCR
2.2. Isothermal Amplification Methods
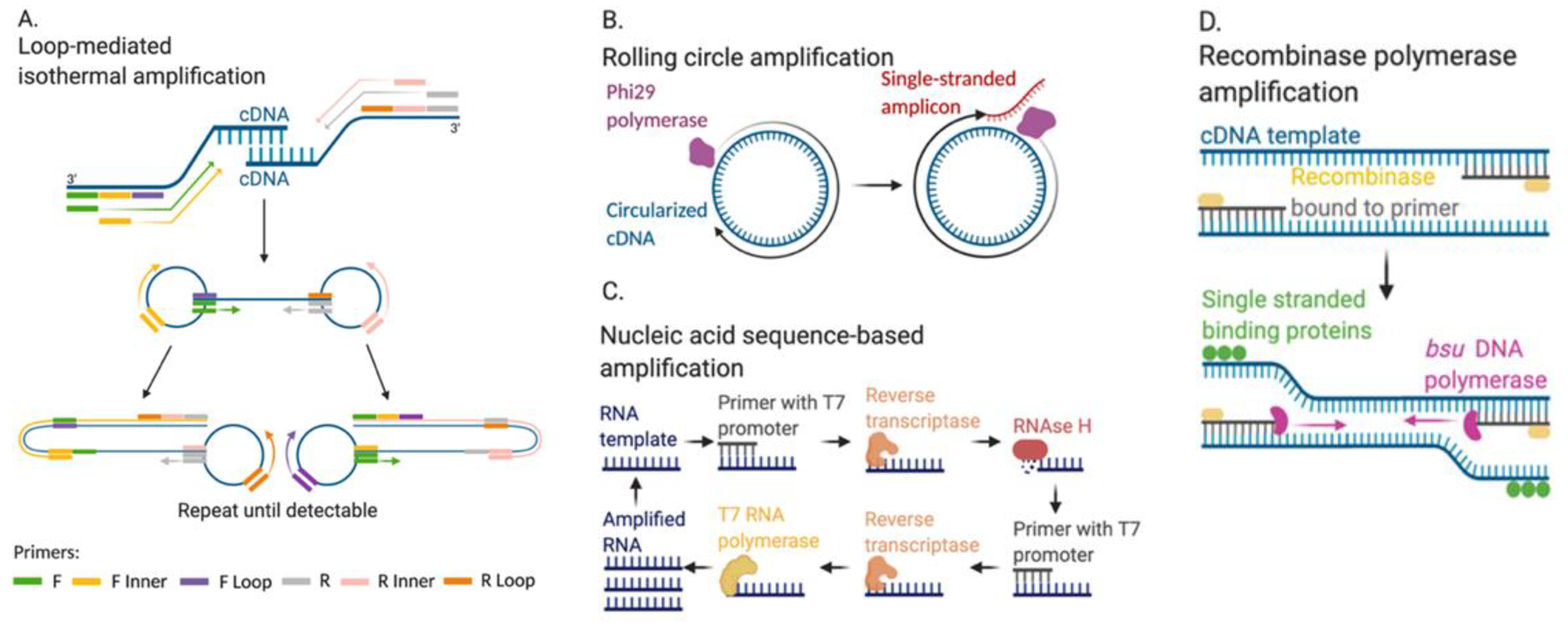
2.2.1. Loop-Mediated Isothermal Amplification (LAMP)
2.2.2. Rolling Circle Amplification
2.2.3. Nucleic Acid Sequence-Based Amplification
2.2.4. Recombinase Polymerase Amplification and CRISPR-Based Diagnostics
3. Direct Nucleic Acid Detection Techniques
3.1. Optical Techniques
3.2. Electrochemical Techniques
3.3. Next-Generation Sequencing
4. CLIA-Waived Nucleic Acid Diagnostics
- Minimal scientific knowledge for performance of test;
- Minimal training and experience for performance and analysis of test;
- Stable and reliable reagents (prepackaged, no special handling, room temperature storage conditions);
- Automated or easily controlled operational steps;
- Stable and readily available materials for calibration, quality control, and external proficiency;
- Automated and/or easily performed troubleshooting and maintenance;
- Minimal interpretation of results.
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lee, V.J.; Ho, Z.J.M.; Goh, E.H.; Campbell, H.; Cohen, C.; Cozza, V.; Fitzner, J.; Jara, J.; Krishnan, A.; Bresee, J.; et al. Advances in measuring influenza burden of disease. Influ. Other Respir. Viruses 2018, 12, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C.R.; Holmes, E.C.; Morens, D.M.; Park, E.-C.; Burke, D.S.; Calisher, C.H.; Laughlin, C.A.; Saif, L.J.; Daszak, P. Cross-Species Virus Transmission and the Emergence of New Epidemic Diseases. Microbiol. Mol. Biol. Rev. 2008, 72, 457–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jester, B.J.; Uyeki, T.M.; Patel, A.; Koonin, L.; Jernigan, D.B. 100 Years of Medical Countermeasures and Pandemic Influenza Preparedness. Am. J. Public Health 2018, 108, 1469–1472. [Google Scholar] [CrossRef]
- Vemula, S.V.; Sayedahmed, E.E.; Sambhara, S.; Mittal, S.K. Vaccine approaches conferring cross-protection against influenza viruses. Expert Rev. Vaccines 2017, 16, 1141–1154. [Google Scholar] [CrossRef]
- Toots, M.; Plemper, R.K. Next-generation direct-acting influenza therapeutics. Transl. Res. 2020, 220, 33–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Galvin, H.D.; Haw, T.Y.; Nutsford, A.N.; Husain, M. Drug resistance in influenza A virus: The epidemiology and management. Infect. Drug Resist. 2017, 10, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Murdock, R.C.; Gallegos, K.M.; Hagen, J.A.; Kelley-Loughnane, N.; Weiss, A.A.; Papautsky, I. Development of a point-of-care diagnostic for influenza detection with antiviral treatment effectiveness indication. Lab Chip 2016, 17, 332–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention. Overview of Influenza Surveillance in the United States. Available online: https://www.cdc.gov/flu/weekly/overview.htm (accessed on 9 January 2021).
- Lalkhen, M.C.F.A.G.; McCluskey, B.M.C.F.A. Clinical tests: Sensitivity and specificity. Contin. Educ. Anaesth. Crit. Care Pain 2008, 8, 221–223. [Google Scholar] [CrossRef] [Green Version]
- Vemula, S.V.; Zhao, J.; Liu, J.; Wang, X.; Biswas, S.; Hewlett, I. Current Approaches for Diagnosis of Influenza Virus Infections in Humans. Viruses 2016, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Nakatsu, S.; Murakami, S.; Shindo, K.; Horimoto, T.; Sagara, H.; Noda, T.; Kawaoka, Y. Influenza C and D Viruses Package Eight Organized Ribonucleoprotein Complexes. J. Virol. 2018, 92, e02084-17. [Google Scholar] [CrossRef] [Green Version]
- Asha, K.; Kumar, B. Emerging Influenza D Virus Threat: What We Know so Far! J. Clin. Med. 2019, 8, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, U.; Su, Y.C.F.; Vijaykrishna, D.; Smith, G.J.D. The ecology and adaptive evolution of influenza A interspecies transmission. Influ. Other Respir. Viruses 2016, 11, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Hamming, P.H.; Overeem, N.J.; Huskens, J. Influenza as a molecular walker. Chem. Sci. 2019, 11, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Osterhaus, A.; Rimmelzwaan, G.; Martina, B.; Bestebroer, T.; Fouchier, R. Influenza B virus in seals. Science 2000, 288, 1051–1053. [Google Scholar] [CrossRef]
- Salem, E.; Cook, E.A.; Lbacha, H.A.; Oliva, J.; Awoume, F.; Aplogan, G.; Hymann, E.C.; Muloi, D.; Deem, S.L.; Alali, S.; et al. Serologic Evidence for Influenza C and D Virus among Ruminants and Camelids, Africa, 1991–2015. Emerg. Infect. Dis. 2017, 23, 1556–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohwada, K.; Kitame, F.; Sugawara, K.; Nishimura, H.; Homma, M.; Nakamura, K. Distribution of the Antibody to Influenza C Virus in Dogs and Pigs in Yamagata Prefecture, Japan. Microbiol. Immunol. 1987, 31, 1173–1180. [Google Scholar] [CrossRef]
- Yuanji, G.; Fengen, J.; Ping, W.; Min, W.; Jiming, Z.J. Isolation of Influenza C Virus from Pigs and Experimental Infection of Pigs with Influenza C Virus. J. Gen. Virol. 1983, 64, 177–182. [Google Scholar] [CrossRef]
- Gao, Q.; Chou, Y.-Y.; Doğanay, S.; Vafabakhsh, R.; Ha, T.; Palese, P. The Influenza A Virus PB2, PA, NP, and M Segments Play a Pivotal Role during Genome Packaging. J. Virol. 2012, 86, 7043–7051. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.-H.; Yang, W.; Beineke, A.; Dijkman, R.; Matrosovich, M.; Baumgärtner, W.; Thiel, V.E.; Valentin-Weigand, P.; Meng, F.; Herrler, G. The differentiated airway epithelium infected by influenza viruses maintains the barrier function despite a dramatic loss of ciliated cells. Sci. Rep. 2016, 6, 39668. [Google Scholar] [CrossRef] [Green Version]
- Irving, S.A.; Vandermause, M.F.; Shay, D.K.; Belongia, E.A. Comparison of Nasal and Nasopharyngeal Swabs for Influenza Detection in Adults. Clin. Med. Res. 2012, 10, 215–218. [Google Scholar] [CrossRef] [Green Version]
- Ngaosuwankul, N.; Noisumdaeng, P.; Komolsiri, P.; Pooruk, P.; Chokephaibulkit, K.; Chotpitayasunondh, T.; Sangsajja, C.; Chuchottaworn, C.; Farrar, J.J.; Puthavathana, P. Influenza A viral loads in respiratory samples collected from patients infected with pandemic H1N1, seasonal H1N1 and H3N2 viruses. Virol. J. 2010, 7, 75. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, D.; Shimoni, A.; Keren-Naus, A.; Steinberg, R.; Shemer-Avni, Y.; Lieberman, D.; Lieberman, D. Identification of Respiratory Viruses in Adults: Nasopharyngeal versus Oropharyngeal Sampling. J. Clin. Microbiol. 2009, 47, 3439–3443. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-G.; Yun, S.G.; Kim, M.Y.; Park, K.; Cho, C.H.; Yoon, S.Y.; Nam, M.H.; Lee, C.K.; Cho, Y.-J.; Lim, C.S. Comparison between Saliva and Nasopharyngeal Swab Specimens for Detection of Respiratory Viruses by Multiplex Reverse Transcription-PCR. J. Clin. Microbiol. 2016, 55, 226–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, S.; Thompson, M.G.; Flannery, B.; Fry, A. Comparison of Respiratory Specimen Collection Methods for Detection of Influenza Virus Infection by Reverse Transcription-PCR: A Literature Review. J. Clin. Microbiol. 2019, 57, e00027-19. [Google Scholar] [CrossRef] [Green Version]
- Baccam, P.; Beauchemin, C.; Macken, C.A.; Hayden, F.G.; Perelson, A.S. Kinetics of Influenza A Virus Infection in Humans. J. Virol. 2006, 80, 7590–7599. [Google Scholar] [CrossRef] [Green Version]
- Dare, R.; Zhu, Y.; Williams, J.V.; Griffin, M.; Talbot, H.K. Detection of influenza by real time RT-PCR is not affected by delays in respiratory specimen processing. J. Med. Virol. 2016, 88, 1891–1895. [Google Scholar] [CrossRef] [Green Version]
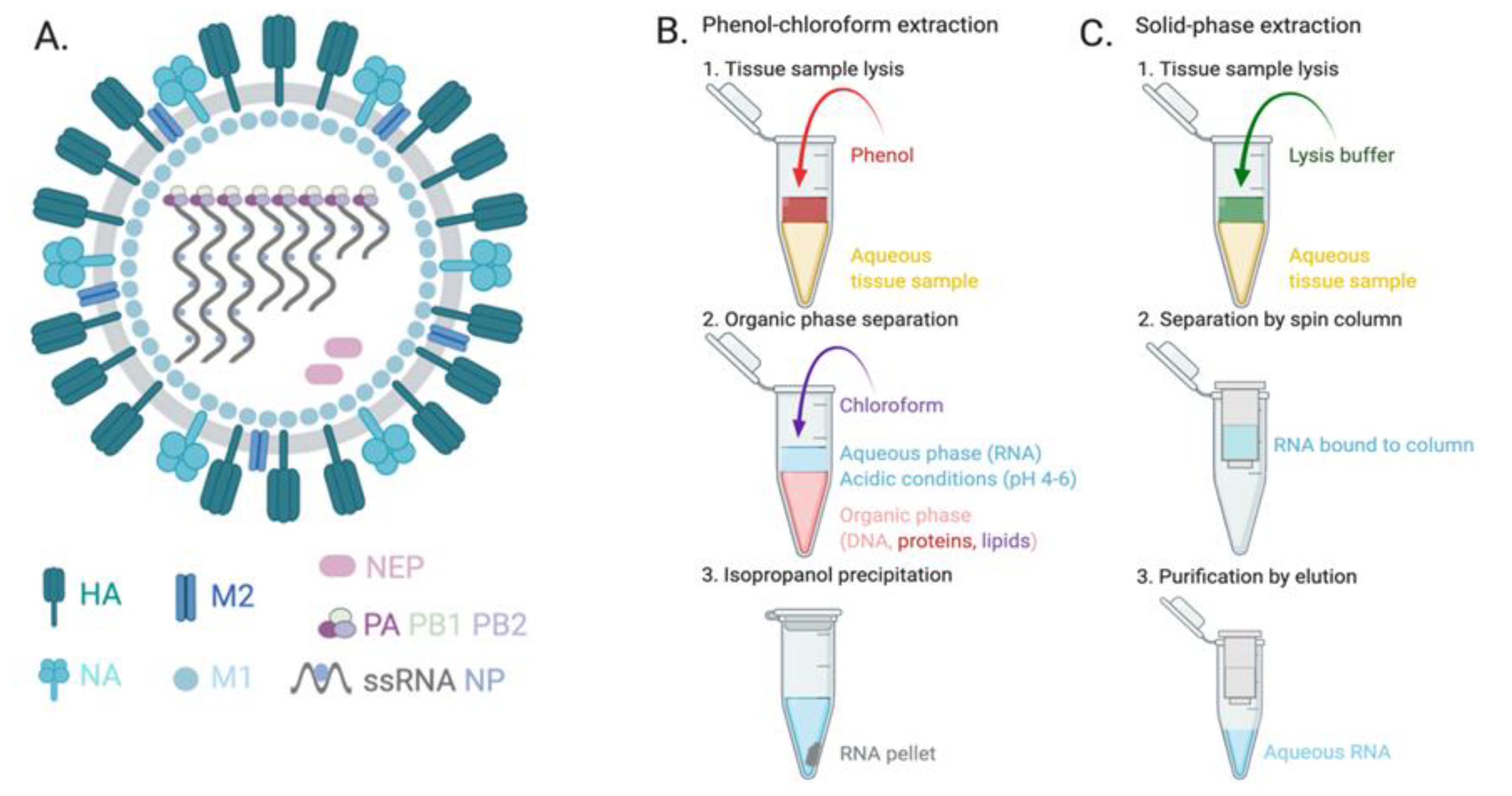
- Tan, S.C.; Yiap, B.C. DNA, RNA, and Protein Extraction: The Past and The Present. J. Biomed. Biotechnol. 2009, 2009, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA Using TRIzol (TRI Reagent). Cold Spring Harb. Protoc. 2010, 2010, 5439. [Google Scholar] [CrossRef]
- Yang, G.; Erdman, D.E.; Kodani, M.; Kools, J.; Bowen, M.D.; Fields, B.S. Comparison of commercial systems for extraction of nucleic acids from DNA/RNA respiratory pathogens. J. Virol. Methods 2011, 171, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.R.; Wang, J.; Opal, S.M.; Tripathi, A. Isolating Influenza RNA from Clinical Samples Using Microfluidic Oil-Water Interfaces. PLoS ONE 2016, 11, e0149522. [Google Scholar] [CrossRef] [PubMed]
- Monis, P.; Giglio, S. Nucleic acid amplification-based techniques for pathogen detection and identification. Infect. Genet. Evol. 2006, 6, 2–12. [Google Scholar] [CrossRef]
- Souf, S. Recent advances in diagnostic testing for viral infections. Bioscience Horizons. Int. J. Stud. Res. 2016, 9. [Google Scholar] [CrossRef]
- Lee, N.; Chan, P.K.; Hui, D.S.C.; Rainer, T.H.; Wong, E.; Choi, K.; Lui, G.C.Y.; Wong, B.C.K.; Wong, R.Y.K.; Lam, W.; et al. Viral Loads and Duration of Viral Shedding in Adult Patients Hospitalized with Influenza. J. Infect. Dis. 2009, 200, 492–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Henrickson, K.J. Update on Influenza Diagnostics: Lessons from the Novel H1N1 Influenza A Pandemic. Clin. Microbiol. Rev. 2012, 25, 344–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahrberg, C.D.; Lee, J.M.; Chung, B.G. Microwell Array-based Digital PCR for Influenza Virus Detection. BioChip J. 2019, 13, 269–276. [Google Scholar] [CrossRef]
- Dong, H.; Zhang, Y.; Xiong, H.; Yan, A.; Ding, G.; Chen, Y.; Xie, L.; Chen, J.; Zhang, G.; Hao, P.; et al. Detection of human novel influenza A (H1N1) viruses using multi-fluorescent real-time RT-PCR. Virus Res. 2010, 147, 85–90. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Porter, E.; Lu, N.; Li, Y.; Yuan, F.; Lohman, M.; Noll, L.; Zheng, W.; Stoy, C.; et al. Development of a multiplex real-time RT-PCR assay for simultaneous detection and differentiation of influenza A, B, C, and D viruses. Diagn. Microbiol. Infect. Dis. 2019, 95, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Saito, M.; Kondoh, K.; Hossain, M.M.; Koketsu, R.; Sasaki, T.; Nagatani, N.; Ikuta, K.; Tamiya, E. Rapid detection for primary screening of influenza A virus: Microfluidic RT-PCR chip and electrochemical DNA sensor. Analyst 2011, 136, 2064–2068. [Google Scholar] [CrossRef]
- Kim, Y.; Abafogi, A.T.; Tran, B.M.; Kim, J.; Lee, J.; Chen, Z.; Bae, P.K.; Park, K.; Shin, Y.-B.; Van Noort, D.; et al. Integrated Microfluidic Preconcentration and Nucleic Amplification System for Detection of Influenza A Virus H1N1 in Saliva. Micromachines 2020, 11, 203. [Google Scholar] [CrossRef] [Green Version]
- Cao, Q.; Mahalanabis, M.; Chang, J.; Carey, B.; Hsieh, C.; Stanley, A.; Odell, C.A.; Mitchell, P.; Feldman, J.; Pollock, N.R.; et al. Microfluidic Chip for Molecular Amplification of Influenza A RNA in Human Respiratory Specimens. PLoS ONE 2012, 7, e33176. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.M.; Sabbavarapu, N.M.; Fu, C.Y.; Jan, J.T.; Wang, J.R.; Hung, S.C. Lee, G.-B. An integrated microfluidic system for rapid detection and multiple subtyping of influenza A viruses by using glycan-coated magnetic beads and RT-PCR. Lab Chip 2019, 19, 1277–1286. [Google Scholar] [CrossRef]
- Kubo, T.; Agoh, M.; Mai, L.Q.; Fukushima, K.; Nishimura, H.; Yamaguchi, A.; Hirano, M.; Yoshikawa, A.; Hasebe, F.; Kohno, S.; et al. Development of a Reverse Transcription-Loop-Mediated Isothermal Amplification Assay for Detection of Pandemic (H1N1) 2009 Virus as a Novel Molecular Method for Diagnosis of Pandemic Influenza in Resource-Limited Settings. J. Clin. Microbiol. 2010, 48, 728–735. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.J.; Baek, Y.H.; Lloren, K.K.S.; Choi, W.-S.; Jeong, J.H.; Antigua, K.J.C.; Kwon, H.-I.; Park, S.-J.; Kim, E.-H.; Kim, Y.-I.; et al. Rapid and simple colorimetric detection of multiple influenza viruses infecting humans using a reverse transcriptional loop-mediated isothermal amplification (RT-LAMP) diagnostic platform. BMC Infect. Dis. 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahony, J.; Chong, S.; Bulir, D.; Ruyter, A.; Mwawasi, K.; Waltho, D. Multiplex loop-mediated isothermal amplification (M-LAMP) assay for the detection of influenza A/H1, A/H3 and influenza B can provide a specimen-to-result diagnosis in 40min with single genome copy sensitivity. J. Clin. Virol. 2013, 58, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Segawa, Y.; Watanabe, H.; Yotoriyama, T.; Kai, S.; Yasuda, A.; Shimizu, N.; Tojo, N. Point-of-care testing system enabling 30 min detection of influenza genes. Lab Chip 2011, 11, 1166–1167. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Wang, P.; Zang, Y.; Ge, Y.; Jin, Q.; Zhao, J.; Xu, X.; Zhao, G.; Mao, H. A colorimetric method for H1N1 DNA detection using rolling circle amplification. Analyst 2013, 138, 3457–3462. [Google Scholar] [CrossRef] [PubMed]
- Garbarino, F.; Minero, G.A.S.; Rizzi, G.; Fock, J.; Hansen, M.F. Integration of rolling circle amplification and optomagnetic detection on a polymer chip. Biosens. Bioelectron. 2019, 142, 111485. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.; Telles, J.-N.; Corden, S.; Gao, R.; Vernet, G.; Van Aarle, P.; Shu, Y.-L. Development and validation of a commercial real-time NASBA assay for the rapid confirmation of influenza A H5N1 virus in clinical samples. J. Virol. Methods 2010, 170, 173–176. [Google Scholar] [CrossRef]
- Ge, Y.; Cui, L.; Qi, X.; Shan, J.; Shan, Y.; Qi, Y.; Wu, B.; Wang, H.; Shi, Z. Detection of novel swine origin influenza A virus (H1N1) by real-time nucleic acid sequence-based amplification. J. Virol. Methods 2010, 163, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.-T.; Curran, M.D.; Ellis, J.S.; Parmar, S.; Ritchie, A.V.; Sharma, P.I.; Allain, J.-P.; Jalal, H.; Zambon, M.; Lee, H.H. Nucleic Acid Dipstick Test for Molecular Diagnosis of Pandemic H1N1. J. Clin. Microbiol. 2010, 48, 3608–3613. [Google Scholar] [CrossRef] [Green Version]
- Freije, C.A.; Myhrvold, C.; Boehm, C.K.; Lin, A.E.; Welch, N.L.; Carter, A.; Metsky, H.C.; Luo, C.Y.; Abudayyeh, O.O.; Gootenberg, J.S.; et al. Programmable Inhibition and Detection of RNA Viruses Using Cas13. Mol. Cell 2019, 76, 826–837.e11. [Google Scholar] [CrossRef] [Green Version]
- Mayuramart, O.; Nimsamer, P.; Rattanaburi, S.; Chantaravisoot, N.; Khongnomnan, K.; Chansaenroj, J.; Puenpa, J.; Suntronwong, N.; Vichaiwattana, P.; Poovorawan, Y.; et al. Detection of severe acute respiratory syndrome coronavirus 2 and influenza viruses based on CRISPR-Cas12a. Exp. Biol. Med. 2020, 10. [Google Scholar] [CrossRef]
- Ackerman, C.M.; Myhrvold, C.; Thakku, S.G.; Freije, C.A.; Metsky, H.C.; Yang, D.K.; Ye, S.H.; Boehm, C.K.; Kosoko-Thoroddsen, T.-S.F.; Kehe, J.; et al. Massively multiplexed nucleic acid detection with Cas13. Nat. Cell Biol. 2020, 582, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.J.; Selvarangan, R. Evaluation of the Alere i Influenza A&B Nucleic Acid Amplification Test by Use of Respiratory Specimens Collected in Viral Transport Medium. J. Clin. Microbiol. 2014, 52, 3992–3995. [Google Scholar] [CrossRef] [Green Version]
- Walper, S.A.; Aragonés, G.L.; Sapsford, K.E.; Brown, C.W.; Rowland, C.E.; Breger, J.C.; Medintz, I.L. Detecting Biothreat Agents: From Current Diagnostics to Developing Sensor Technologies. ACS Sens. 2018, 3, 1894–2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angione, S.L.; Inde, Z.; Beck, C.M.; Artenstein, A.W.; Opal, S.M.; Tripathi, A. Microdroplet Sandwich Real-Time RT-PCR for Detection of Pandemic and Seasonal Influenza Subtypes. PLoS ONE 2013, 8, e73497. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.L.; Hu, P.; Shafer, D.A. Seven Novel Probe Systems for Real-Time PCR Provide Absolute Single-Base Discrimination, Higher Signaling, and Generic Components. J. Mol. Diagn. 2014, 16, 627–638. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.J.; Osborn, A.M. Advantages and limitations of quantitative PCR (Q-PCR)-based approaches in microbial ecology. FEMS Microbiol. Ecol. 2009, 67, 6–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z. Design, Synthesis, Purification and Characterization of Molecular Beacons; Chaoyong, J., Yang, W.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; p. 194. [Google Scholar]
- Bustin, S.A. Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): Trends and problems. J. Mol. Endocrinol. 2002, 29, 23–39. [Google Scholar] [CrossRef]
- Tahir, M.S.; Mehmood, D.; Sultan, A.U.; Saeed, M.H.; Khan, A.R.; Ansari, F.; Salman, M.M.; Majeed, K.A. A modified strategy of multiplex RT-PCR for simultaneous detection of H5, H7, and H9 subtypes of avian influenza virus based on common forward oligo. J. Appl. Poult. Res. 2016, 25, 322–327. [Google Scholar] [CrossRef]
- Spurgeon, S.L.; Jones, R.C.; Ramakrishnan, R. High Throughput Gene Expression Measurement with Real Time PCR in a Microfluidic Dynamic Array. PLoS ONE 2008, 3, e1662. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chen, F.; Li, Q.; Wang, L.; Fan, C. Isothermal Amplification of Nucleic Acids. Chem. Rev. 2015, 115, 12491–12545. [Google Scholar] [CrossRef] [PubMed]
- Arrigo, N.C.; Watts, D.M.; Frolov, I.; Weaver, S.C. Experimental infection of Aedes sollicitans and Aedes taeniorhynchus with two chimeric Sindbis/Eastern equine encephalitis virus vaccine candidates. Am. J. Trop. Med. Hyg. 2008, 78, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Shojaei, T.R.; Tabatabaei, M.; Shawky, S.; Salleh, M.A.M.; Bald, D. A review on emerging diagnostic assay for viral detection: The case of avian influenza virus. Mol. Biol. Rep. 2014, 42, 187–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, A.; Shibasaki, F. Updated Values for Molecular Diagnosis for Highly Pathogenic Avian Influenza Virus. Viruses 2012, 4, 1235–1257. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.; Hibbitts, S.; Owen, N.; Corden, S.A.; Harrison, G.; Fox, J.; Gelder, C.; Westmoreland, D. Development and evaluation of a real-time nucleic acid sequence based amplification assay for rapid detection of influenza A. J. Med Virol. 2004, 74, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [Green Version]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Servellita, V.; Singh, J.; Miao, X.; Streithorst, J.A.; Granados, A.; Sotomayor-Gonzalez, A.; et al. CRISPR–Cas12-based detection of SARS-CoV-2. Nat. Biotechnol. 2020, 38, 870–874. [Google Scholar] [CrossRef] [Green Version]
- Kellner, M.J.; Koob, J.G.; Gootenberg, J.S.; Abudayyeh, O.O.; Zhang, F. SHERLOCK: Nucleic acid detection with CRISPR nucleases. Nat. Protoc. 2019, 14, 2986–3012. [Google Scholar] [CrossRef]
- Ducheyne, P.; Grainger, D.W.; Healy, K.E.; Hutmacher, D.W.; Kirkpack, C.J. 3.30 Biosensors based on sol–gel derived materials In Comprehensive Biomaterials, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 3, pp. 657–689. [Google Scholar]
- Pan, Q.; Luo, F.; Liu, M.; Zhang, X.-L. Oligonucleotide aptamers: Promising and powerful diagnostic and therapeutic tools for infectious diseases. J. Infect. 2018, 77, 83–98. [Google Scholar] [CrossRef]
- Wei, F.; Lillehoj, P.B.; Ho, C.-M. DNA Diagnostics: Nanotechnology-Enhanced Electrochemical Detection of Nucleic Acids. Pediatr. Res. 2010, 67, 458–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinsella, J.M.; Ivanisevic, A. Taking charge of biomolecules. Nat. Nanotechnol. 2007, 2, 596–597. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.M.; Kim, K.R.; Kim, C.S. Biosensor for Rapid and Sensitive Detection of Influenza Virus. Biotechnol. Bioprocess Eng. 2018, 23, 371–382. [Google Scholar] [CrossRef]
- Adegoke, O.; Kato, T.; Park, E.Y. An ultrasensitive alloyed near-infrared quinternary quantum dot-molecular beacon nanodiagnostic bioprobe for influenza virus RNA. Biosens. Bioelectron. 2016, 80, 483–490. [Google Scholar] [CrossRef]
- Liu, G.; Li, J.; Feng, D.-Q.; Zhu, J.-J.; Wang, W. Silver Nanoclusters Beacon as Stimuli-Responsive Versatile Platform for Multiplex DNAs Detection and Aptamer–Substrate Complexes Sensing. Anal. Chem. 2016, 89, 1002–1008. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-C.D.; Liao, Y.-C.; Lai, Y.-H.; Chuang, M.-C. Recognition of Dual Targets by a Molecular Beacon-Based Sensor: Subtyping of Influenza A Virus. Anal. Chem. 2015, 87, 5410–5416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Tian, J.; He, Y.; Chen, S.; Jiang, Y.; Zhao, Y.; Zhao, S. Protein-binding aptamer assisted signal amplification for the detection of influenza A (H1N1) DNA sequences based on quantum dot fluorescence polarization analysis. Analyst 2013, 138, 4722. [Google Scholar] [CrossRef]
- Tian, J.; Zhao, H.; Liu, M.; Chen, Y.; Quan, X. Detection of influenza A virus based on fluorescence resonance energy transfer from quantum dots to carbon nanotubes. Anal. Chim. Acta 2012, 723, 83–87. [Google Scholar] [CrossRef]
- Lim, S.H.; Buchy, P.; Mardy, S.; Kang, M.S.; Yu, A.D.C. Specific Nucleic Acid Detection Using Photophysical Properties of Quantum Dot Probes. Anal. Chem. 2010, 82, 886–891. [Google Scholar] [CrossRef]
- Bai, H.; Wang, R.; Hargis, B.; Lu, H.; Li, Y. A SPR Aptasensor for Detection of Avian Influenza Virus H5N1. Sensors 2012, 12, 12506–12518. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Bhatia, S.; Pateriya, A.K.; Sood, R.; Nagarajan, S.; Murugkar, H.V.; Kumar, S.; Singh, P.; Singh, V.P. Label-free peptide nucleic acid biosensor for visual detection of multiple strains of influenza A virus suitable for field applications. Anal. Chim. Acta 2020, 1093, 123–130. [Google Scholar] [CrossRef]
- Tran, T.L.; Nguyen, T.T.; Chu, V.T.; Tran, Q.T.; Mai, A.T. Detection of influenza A virus using carbon nanotubes field effect transistor based DNA sensor. Phys. E Low Dimens. Syst. Nanostruct. 2017, 93, 83–86. [Google Scholar] [CrossRef]
- Bonanni, A.; Pividori, M.I.; Del Valle, M. Impedimetric detection of influenza A (H1N1) DNA sequence using carbon nanotubes platform and gold nanoparticles amplification. Analyst 2010, 135, 1765–1772. [Google Scholar] [CrossRef] [PubMed]
- Tam, P.D.; Van Hieu, N.; Chien, N.D.; Le, A.-T.; Tuan, M.A. DNA sensor development based on multi-wall carbon nanotubes for label-free influenza virus (type A) detection. J. Immunol. Methods 2009, 350, 118–124. [Google Scholar] [CrossRef]
- Lee, J.; Morita, M.; Takemura, K.; Park, E.Y. A multi-functional gold/iron-oxide nanoparticle-CNT hybrid nanomaterial as virus DNA sensing platform. Biosens. Bioelectron. 2018, 102, 425–431. [Google Scholar] [CrossRef] [PubMed]
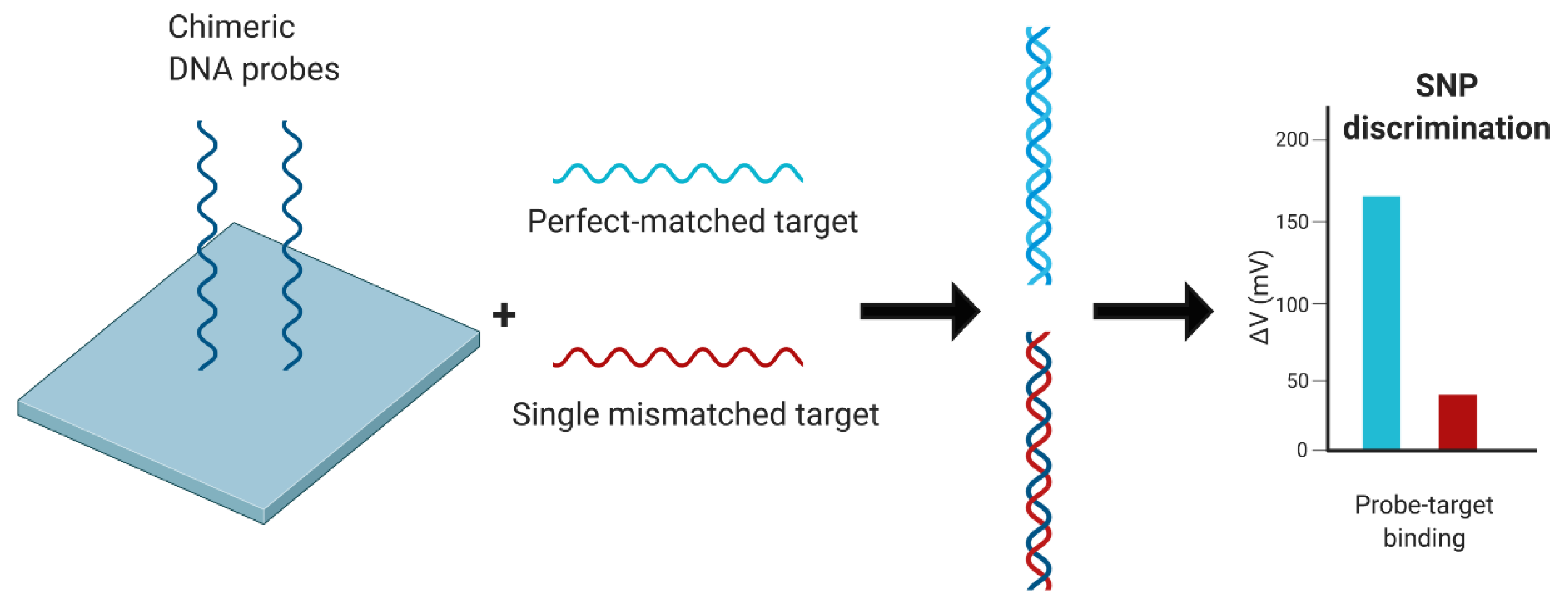
- Huang, C.-J.; Lin, Z.-E.; Yang, Y.-S.; Chan, H.W.-H.; Chen, W.-Y. Neutralized chimeric DNA probe for detection of single nucleotide polymorphism on surface plasmon resonance biosensor. Biosens. Bioelectron. 2018, 99, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Loo, J.F.C.; Wang, S.S.; Peng, F.; He, J.A.; He, L.; Guo, Y.C.; Gu, D.; Kwok, H.C.; Wu, S.Y.; Ho, H.P.; et al. A non-PCR SPR platform using RNase H to detect MicroRNA 29a-3p from throat swabs of human subjects with influenza A virus H1N1 infection. Analyst 2015, 140, 4566–4575. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, I.; Malecka, K.; Stachyra, A.; Góra-Sochacka, A.; Sirko, A.; Zagórski-Ostoja, W.; Radecka, H.; Radecki, J. Single Electrode Genosensor for Simultaneous Determination of Sequences Encoding Hemagglutinin and Neuraminidase of Avian Influenza Virus Type H5N1. Anal. Chem. 2013, 85, 10167–10173. [Google Scholar] [CrossRef]
- Dong, S.; Zhao, R.; Zhu, J.; Lu, X.; Li, Y.; Qiu, S.; Jia, L.; Jiao, X.; Song, S.; Fan, C.; et al. Electrochemical DNA Biosensor Based on a Tetrahedral Nanostructure Probe for the Detection of Avian Influenza A (H7N9) Virus. ACS Appl. Mater. Interfaces 2015, 7, 8834–8842. [Google Scholar] [CrossRef]
- Chan, C.; Shi, J.; Fan, Y.; Yang, M. A microfluidic flow-through chip integrated with reduced graphene oxide transistor for influenza virus gene detection. Sens. Actuators B Chem. 2017, 251, 927–933. [Google Scholar] [CrossRef]
- Malecka, K.; Stachyra, A.; Góra-Sochacka, A.; Sirko, A.; Zagórski-Ostoja, W.; Dehaen, W.; Radecka, H.; Radecki, J. New redox-active layer create via epoxy–amine reaction—The base of genosensor for the detection of specific DNA and RNA sequences of avian influenza virus H5N1. Biosens. Bioelectron. 2015, 65, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Liu, J.; Vemula, S.V.; Lin, C.; Tan, J.; Ragupathy, V.; Wang, X.; Mbondji-Wonje, C.; Ye, Z.; Landry, M.L.; et al. Sensitive Detection and Simultaneous Discrimination of Influenza A and B Viruses in Nasopharyngeal Swabs in a Single Assay Using Next-Generation Sequencing-Based Diagnostics. PLoS ONE 2016, 11, e0163175. [Google Scholar] [CrossRef]
- Lewandowski, K.; Xu, Y.; Pullan, S.T.; Lumley, S.F.; Foster, D.; Sanderson, N.; Vaughan, A.; Morgan, M.; Bright, N.; Kavanagh, J.; et al. Metagenomic Nanopore Sequencing of Influenza Virus Direct from Clinical Respiratory Samples. J. Clin. Microbiol. 2019, 58, e00963-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziąbowska, K.; Czaczyk, E.; Nidzworski, D. Detection Methods of Human and Animal Influenza Virus—Current Trends. Biosensors 2018, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, Z.; Yu, Y.; Wang, M.; Li, J.; Zhang, Z.; Liu, J.; Wu, X.; Lu, A.; Zhang, G.; Zhang, B. Recent Advances in SELEX Technology and Aptamer Applications in Biomedicine. Int. J. Mol. Sci. 2017, 18, 2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kustin, T.; Ling, G.; Sharabi, S.; Ram, D.; Friedman, N.; Zuckerman, N.; Bucris, E.D.; Glatman-Freedman, A.; Stern, A.; Mandelboim, M. A method to identify respiratory virus infections in clinical samples using next-generation sequencing. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- O’Flaherty, B.M.; Li, Y.; Tao, Y.; Paden, C.R.; Queen, K.; Zhang, J.; Dinwiddie, D.L.; Gross, S.M.; Schroth, G.P.; Tong, S. Comprehensive viral enrichment enables sensitive respiratory virus genomic identification and analysis by next generation sequencing. Genome Res. 2018, 28, 869–877. [Google Scholar] [CrossRef] [Green Version]
- Fischer, N.; Indenbirken, D.; Meyer, T.; Lutgehetmann, M.; Lellek, H.; Spohn, M.; Aepfelbacher, M.; Alawi, M.; Grundhoff, A. Evaluation of Unbiased Next-Generation Sequencing of RNA (RNA-seq) as a Diagnostic Method in Influenza Virus-Positive Respiratory Samples. J. Clin. Microbiol. 2015, 53, 2238–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prachayangprecha, S.; Schapendonk, C.; Koopmans, M.P.; Osterhaus, A.; Schürch, A.; Pas, S.; Van Der Eijk, A.A.; Poovorawan, Y.; Haagmans, B.; Smits, S. Exploring the Potential of Next-Generation Sequencing in Detection of Respiratory Viruses. J. Clin. Microbiol. 2014, 52, 3722–3730. [Google Scholar] [CrossRef] [Green Version]
- Graf, E.H.; Simmon, K.E.; Tardif, K.D.; Hymas, W.; Flygare, S.; Eilbeck, K.; Yandell, M.; Schlaberg, R. Unbiased Detection of Respiratory Viruses by Use of RNA Sequencing-Based Metagenomics: A Systematic Comparison to a Commercial PCR Panel. J. Clin. Microbiol. 2016, 54, 1000–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United States Food and Drug Administration. CLIA Categorizations. 2020. Available online: https://www.fda.gov/medical-devices/ivd-regulatory-assistance/clia-categorizations (accessed on 12 January 2020).
- Young, S.; Phillips, J.; Griego-Fullbright, C.; Wagner, A.; Jim, P.; Chaudhuri, S.; Tang, S.; Sickler, J. Molecular point-of-care testing for influenza A/B and respiratory syncytial virus: Comparison of workflow parameters for the ID Now and cobas Liat systems. J. Clin. Pathol. 2020, 73, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Azar, M.M.; Landry, M.L. Detection of Influenza A and B Viruses and Respiratory Syncytial Virus by Use of Clinical Laboratory Improvement Amendments of 1988 (CLIA)-Waived Point-of-Care Assays: A Paradigm Shift to Molecular Tests. J. Clin. Microbiol. 2018, 56, e00367-18. [Google Scholar] [CrossRef]
- Kanwar, N.; Michael, J.; Doran, K.; Montgomery, E.; Selvarangan, R. Comparison of the ID Now Influenza A & B 2, Cobas Influenza A/B, and Xpert Xpress Flu Point-of-Care Nucleic Acid Amplification Tests for Influenza A/B Virus Detection in Children. J. Clin. Microbiol. 2020, 58, 1611–1619. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.H.; Lam, H.Y.; Yip, C.C.; Cheng, V.C.; Chan, J.F.; Leung, T.H.; Sridhar, S.; Chan, K.H.; Tang, B.S.; Yuen, K.Y. Evaluation of the molecular Xpert Xpress Flu/RSV assay vs. Alere i Influenza A & B assay for rapid detection of influenza viruses. Diagn. Microbiol. Infect. Dis. 2018, 90, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Kaplan, S.E.; Lopez, J.C.; Stiles, J.; Lu, X.; Tang, Y.-W. Parallel Validation of Three Molecular Devices for Simultaneous Detection and Identification of Influenza A and B and Respiratory Syncytial Viruses. J. Clin. Microbiol. 2017, 56, e01691-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, D.; Kanwar, N.; Hassan, F.; Essmyer, C.; Selvarangan, R. Comparison of Six Sample-to-Answer Influenza A/B and Respiratory Syncytial Virus Nucleic Acid Amplification Assays Using Respiratory Specimens from Children. J. Clin. Microbiol. 2018, 56, e00930-18. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Method | Influenza Type (Subtype) | Target Gene | Time to Result (min) | No. of Clinical Samples Tested | RNA Detection Limit | Advantages | Current Limitations | Ref. |
|---|---|---|---|---|---|---|---|---|
| Thermal Cycling | ||||||||
| CDC human influenza virus real-time RT-PCR diagnostic panel influenza A/B typing kit | A, B | M1, M2, NS1 | ~240 | n.s. | n.s. | Several kit configurations to detect emerging strains, works on a variety of sample types | Available only to qualified DoD laboratories, U.S. public health laboratories, and NREVSS collaborating laboratories | [35] |
| Microwell array-based digital PCR | A (H7N9) | HA | 60 | None | 700 copies/μL | Simple fabrication, user friendly operation, low device cost (<USD 1/unit) | High background signal caused by reporter dye, dust and other contaminants associated with PDMS material | [36] |
| Multi-fluorescent RT-PCR | A (H1N1) | HA, M1, M2 | 100 | 27 human | n.s. | High sequence coverage, improved multiplex performance | High specificity can lead to primers/probe failure upon viral mutation | [37] |
| Multiplex RT-PCR | A, B, C, D | M, PA, PB | 60 | 758 swine and 1525 bovine | 30 copies per 20 µL reaction (1.5 copies/µL) | Supports surveillance and diagnostics, high sequence coverage (98–100%), simultaneous detection of all 4 influenza types | IBV assays not yet validated in clinical samples | [38] |
| Microfluidic RT-PCR chip and electrochemical DNA sensor | A (H1N1) | M1 | 15 | None | 5 × 103 copies/µL | Rapid, potentially portable and low cost | Poor sensitivity, requires validation in clinical samples | [39] |
| Microfluidic preconcentration and nucleic amplification system | A (H1N1) | M1 | >120 | None | 100 TCID50 | Pre-concentration and amplification in the same device, portable, low cost | Difficult fabrication process, requires validation in clinical samples | [40] |
| Microfluidic RT-PCR chip | A | M1 | >120 | 146 human | 1 copy/µL | Nucleic acid extraction and RT-PCR performed in a single chip, could support POC testing | Long time to result, requires further automation and optimization to achieve POC use | [41] |
| Microfluidic HA x NA subtyping array | A | HA, NA | 100 | None | ~40 copies per 10 µL reaction (4 copies/µL) | Supports simultaneous diagnosis and subtyping, fully automated for POC testing | Variability in various subtyping assays, requires validation in clinical samples | [42] |
| Isothermal | ||||||||
| Reverse transcription-loop-mediated isothermal amplification (RT-LAMP) | A (H1N1) | HA | 40 | 260 human | 10 copies/25 μL (0.4 copies/µL) | Low-cost equipment and minimally trained personnel, well suited for resource-limited settings | 2.2% false negative rate | [43] |
| Colorimetric RT-LAMP | A, B | HA, NA | 60 | 135 human | 0.1–100 genome copies | Reduced detection time compared to conventional PCR, does not require trained personnel | Not as accurate as qRT-PCR, complex primer design | [44] |
| Multiplex RT-LAMP | A (H1N3), B | HA, NS1 | 30 | 202 human | 1 genome equivalent/25 µL reaction (0.04 copies/µL) | Ultra-sensitive, rapid 10 min mechanical sample processing method, rapid 12 min assay run time | 3.6% false negative rate | [45] |
| Microfluidic LAMP | A | HA | 30 | 77 human | n.s. | Rapid, low reaction volume of 1 µL, incorporates solid state reagents | Poor sensitivity (90.9%) | [46] |
| Rolling circle amplification (RCA) | A (H1N1) | M1, M2 | >120 | None | n.s. | Low-cost equipment, colorimetric result for ease of use | No reverse-transcriptase component, assay only tested with DNA template | [47] |
| RCA on polymer chip | B | n.s. | 60 | None | 10 pM | Automated multi-step assay, potential POC application | Requires validation in clinical samples, complex chip design | [48] |
| Real time-nucleic acid sequence-based amplification (NASBA) | A (H5N1) | HA, NA | >120 | 19 human | 10 copies/µL | Supports limited subtyping | Long time to result, recommended for use as a second line test | [49] |
| RT-NASBA | A (H1N1) | HA | 90 | 67 human | 3 copies/µL | No cross-reactivity, 100% sensitivity, speificity, and positive predictive value | Designed for detection of only one viral strain, limited sample comparison | [50] |
| Simple amplification-based assay (SAMBA) | A (H1N1) | HA, M1, M2 | 85 | 262 human | 0.25 PFU per test | Low-cost equipment, easy to read result | Poor sensitivity (95.3%) | [51] |
| Cas13-assisted restriction of viral expression readout (CARVER) | A | NP, M1 | <120 | None | n.s. | End to end platform, multiplexed, Cas13 targeting does not lead to mutations | Needs to be validated on clinical samples | [52] |
| Real-time recombinase polymerase amplifcation CRISPR-Cas12a | A, B | M1 | 60 | 83 human | ~102 copies/µL | Multiplexed assay that can detect COVID-19 and influenza, | Detection of DNA | [53] |
| Combinatorial arrayed reactions for multiplexed evaluation of nucleic acids-Cas13 (CARMEN) | A | HA, NA | 60 | HIV clinical samples | aM level | Highly multiplexed, high sensitivity, low reagent cost (comparative to other SHERLOCK-based methods) | Complex primer and assay design, needs to be validated on influenza clinical samples | [54] |
| Method | Influenza Type (Subtype) | Target Gene | Time to Result (min) | RNA Detection Limit | Advantages | Current Limitations | Ref. |
|---|---|---|---|---|---|---|---|
| Optical | |||||||
| NIR QD-molecular beacon bioprobe | A (H1N1) | NA | 3 | 1.9 copies/mL | Works in complex biological samples | Hazards associated with Cd-containing QDs, requires validation in clinical samples | [79] |
| Activatable silver nanoclusters molecular beacon | A (H1N1, H5N1) | HA, NA | 30 | 2 nM | One-pot detection of multiplex DNA, signal detection without fluorophore cleavage from quencher | Complex multiplexed beacon design, requires validation in clinical samples | [80] |
| Dual target molecular beacon sensor | A (H5N2) | HA, NA | 5 | 120 pM | Highly specific concurrent multisequence detection, works in complex biological samples | Complex molecular beacon and assistant strand design, requires validation in clinical samples | [81] |
| QD fluorescence polarization probes with protein-binding aptamer signal amplification | A (H1N1) | n.s. | 95 | 3.45 nM | Long shelf life, low-cost assay | Hazards associated with Cd-containing QDs, time-consuming, requires validation in clinical samples | [82] |
| QD to CNT FRET-based DNA | A (H5N1) | PB2 | >120 | 9.39 nM | Low-cost assay | Long time to result, hazards associated with Cd-containing QDs, assay only tested with DNA template, requires validation in clinical samples | [83] |
| QD probe with custom-made portable sensor | A (H5N1) | HA | >120 | 12.5 µM | Simple design, portable, does not require wash steps | Result reading device does not have high sensitivity | [84] |
| SPR aptasensor | A (H5N1) | HA | 95 | 0.128 HAU | Portable, validated by poultry swab samples | Expensive and complex assay design | [85] |
| Peptide nucleic acid biosensor | A | M1 | 11 | 2.3 ng | High specificity, label free optical detection, low cost, validated by RNA extracted from clinical samples | Complex design and characterization of peptide nucleic acids | [86] |
| Electrochemical | |||||||
| CNT field effect transistor-based DNA sensor | A | M1 | 1 | 1 pM | Long shelf life | Assay only tested with DNA template, requires validation in clinical samples | [87] |
| Impedimetric DNA sensor using CNT and gold nanoparticle amplification | A (H1N1) | n.s. | 35 | 557 pM | High sensitivity | Equipment operation and data analysis requires extensive training, assay only tested with DNA template, requires validation in clinical samples | [88] |
| Multi-wall CNT DNA sensor | A | M1, M2 | 4 | 0.5 nM | Low cost, interdigitated array microelectrode simplicity | Data analysis requires extensive training, assay only tested with DNA template, requires validation in clinical samples | [89] |
| Gold/iron-oxide CNT-hybrid nanomaterial DNA sensor | A (H1N1) | NA | n.s. | 8.4 pM | High selectivity, simple apparatus | Hybrid nanomaterials are relatively uncommon for sensors and thus need further investigation, assay only tested with DNA template, requires validation in clinical samples | [90] |
| SPR biosensor for SNP DNA probe | A (H1) | HA | >120 | n.s. | Highly stable probe, SNP descrimination | Sensor acquisition is time consuming, instrument operation and data analysis requires extensive training and costs, requires validation in clinical samples | [91] |
| RNase facilitated SPR detection of microRNA | A (H1N1) | MicroRNA 29a–3p | 60 | 1 nM | Does not require thermal cycling, rapid reporter assay for microRNA, tested in clinical samples | Expensive instrumentation, low abundance of microRNA in diagnostic samples | [92] |
| Duo-genosensor with DNA probes | A (H5N1) | HA, NA | >120 | 8–100 nM | Two DNA probes lowers possibility of false positive readings, low cost to handle genosensor | Electrode preparation is time consuming (>18 h), expensive instrumentation, requires validation in clinical samples | [93] |
| DNA tetrahedral nanostructure-based biosensor | A (H7N9) | HA | n.s. | 100 fM | Inexpensive equipment and operation, apparatus portable | High complexity tetrahedral probe design, requires validation in clinical samples | [94] |
| Microfluidic chip integrated with reduced graphene oxide transistor | A (H5N1) | HA | n.s. | 5 pM | Flow through strategy provides sensitivity and stability | rGO transistors are not well studied, requires validation in clinical samples, time consuming impedance spectrum measurements | [95] |
| Redox-active genosensor | A (H5N1) | HA | n.s. | 73 pM | High selectivity for RNA sequences | Not as sensitive to DNA sequences compared to RNA, requires validation in clinical samples | [96] |
| Next-Generation Sequencing | |||||||
| Next-generation sequencing-based diagnostic | A, B | Full genome | >120 | n.s. | Supports surveillance, diagnostics as well as simultaneous virulence and drug resistance profiling | Long time to result, extensive data processing required | [97] |
| Metagenomic Nanopore sequencing | A, B | Full genome | >120 | 102–103 copies/mL | Tested in 27 clinical samples, high speicificity (100%) | Low sensitvity (83%), long time to results, extensive data processing required | [98] |
| Kit | Manufacturer | Instrument | Method | Specimens 1 | Assay Time (min) | Cost per Test | Sensitivity (Influenza A, B) | Specificity (Influenza A, B) | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| BioFire FilmArray RP EZ | BioFire Diagnostics, Salt Lake City, UT, USA | FilmArray 2.0 | Multiplex nested PCR | NPS | 60 | n.s. | N/A | N/A | Kit instructions |
| cobas Liat influenza A/B | Roche, Branchburg, NJ, USA | cobas Liat system | Real-time RT-PCR | NPS | 20 | USD 72.10 | 100%, 94.4% | 98.3%, 100% | [107] |
| cobas Liat influenza A/B & RSV | Roche, Branchburg, NJ, USA | cobas Liat system | Real-time RT-PCR | NPS | 20 | USD 77.25 | 99.6%, 99.3% | 97.5%, 99.7% | [108] |
| ID NOW Influenza A & B 2 | Abbott, Lake Bluff, IL, USA | ID NOW Platform | Nicking enzyme isothermal NAAT | NPS, NS direct or in VTM | 15 | USD 105 | 93.2%, 97.2% | 97% | [109] |
| Silaris Influenza A & B | Mesa Biotech, Inc., San Diego, CA, USA | Silaris Dock | RT-PCR | NS direct | 30 | USD 30 | 97%, 94% | 94%, 99% | Kit instructions |
| Xpert Xpress Flu | Cepheid, Sunnyvale, CA, USA | GeneXpert systems GXII and GXIV | Real-time RT-PCR | NPS, NA, NW | 30 | USD 54.60 | 100%, 97.8% | 99.3–100% | [108] |
| Xpert Xpress Flu/RSV | Cepheid, Sunnyvale, CA, USA | GeneXpert systems GXII and GXIV | Real-time RT-PCR | NPS, NA, NW | 30 | USD 86.50 | 100%, 96.3% | 100% | [110] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Courtney, S.J.; Stromberg, Z.R.; Kubicek-Sutherland, J.Z. Nucleic Acid-Based Sensing Techniques for Diagnostics and Surveillance of Influenza. Biosensors 2021, 11, 47. https://doi.org/10.3390/bios11020047
Courtney SJ, Stromberg ZR, Kubicek-Sutherland JZ. Nucleic Acid-Based Sensing Techniques for Diagnostics and Surveillance of Influenza. Biosensors. 2021; 11(2):47. https://doi.org/10.3390/bios11020047
Chicago/Turabian StyleCourtney, Samantha J., Zachary R. Stromberg, and Jessica Z. Kubicek-Sutherland. 2021. "Nucleic Acid-Based Sensing Techniques for Diagnostics and Surveillance of Influenza" Biosensors 11, no. 2: 47. https://doi.org/10.3390/bios11020047
APA StyleCourtney, S. J., Stromberg, Z. R., & Kubicek-Sutherland, J. Z. (2021). Nucleic Acid-Based Sensing Techniques for Diagnostics and Surveillance of Influenza. Biosensors, 11(2), 47. https://doi.org/10.3390/bios11020047