Bacteriophage-Based Biosensors: A Platform for Detection of Foodborne Bacterial Pathogens from Food and Environment

 , , , , , ,
, , , , , , 

Abstract
:1. Introduction
2. Phage-Based Biosensors
3. Phage-Mediated Bacterial Detection Approaches
3.1. Bacterial β-D-Galactosidase
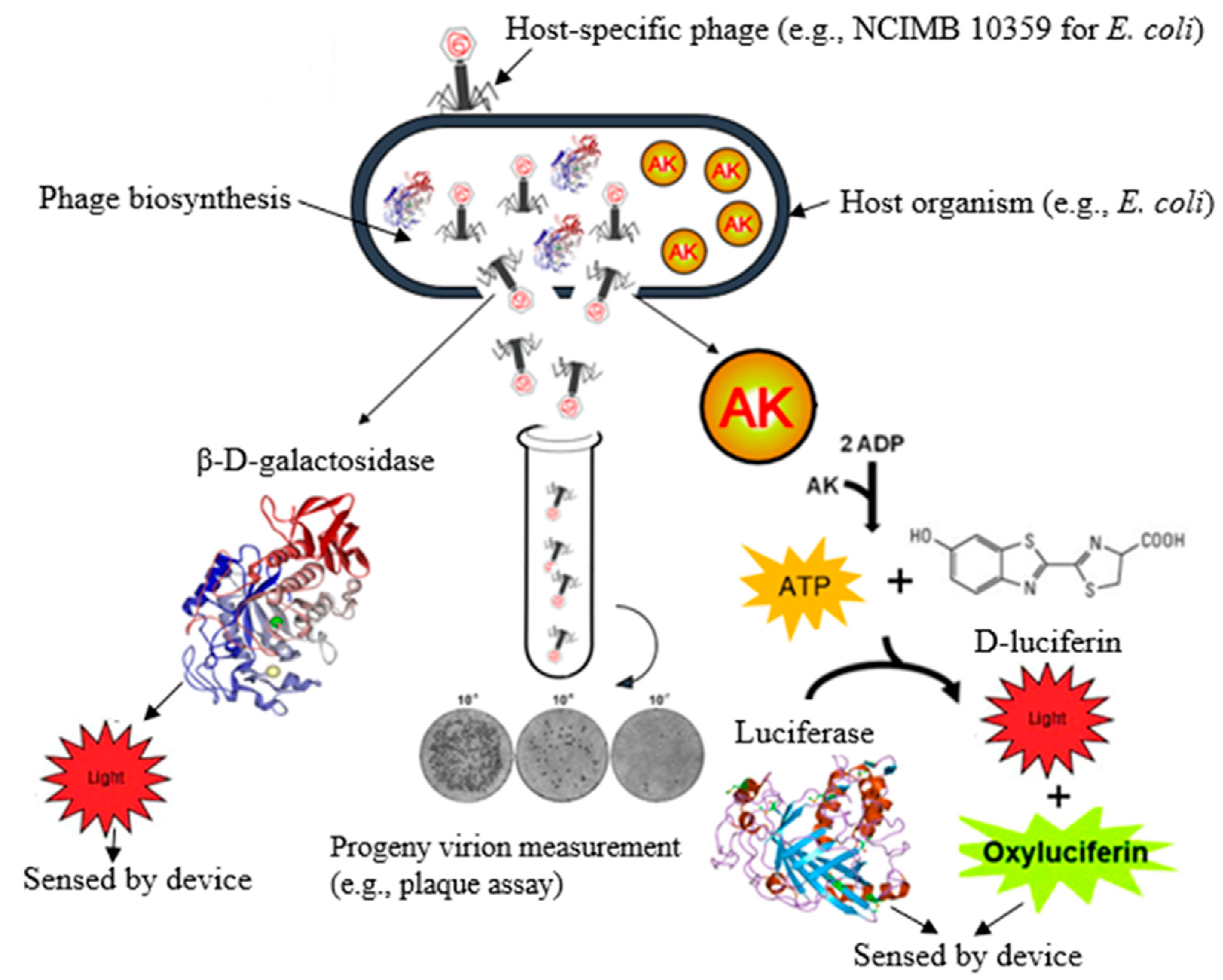
3.2. Adenosine Triphosphate
3.3. Adenylate Kinase
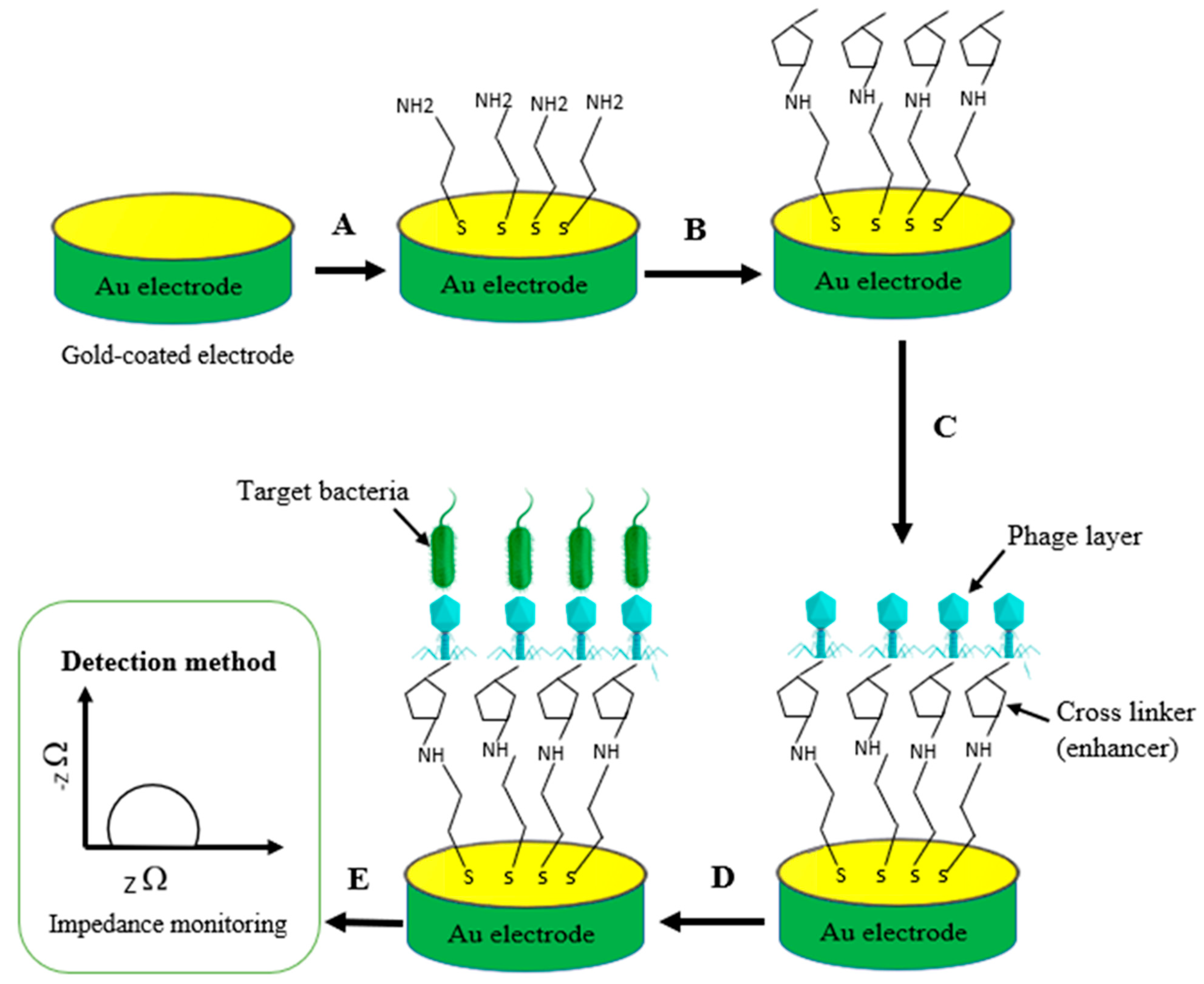
3.4. Conductivity (Impedance)
3.5. Whole-Phage or Progeny Virion Detection
3.6. Reporter Phages
3.7. Phage-Associated Proteins
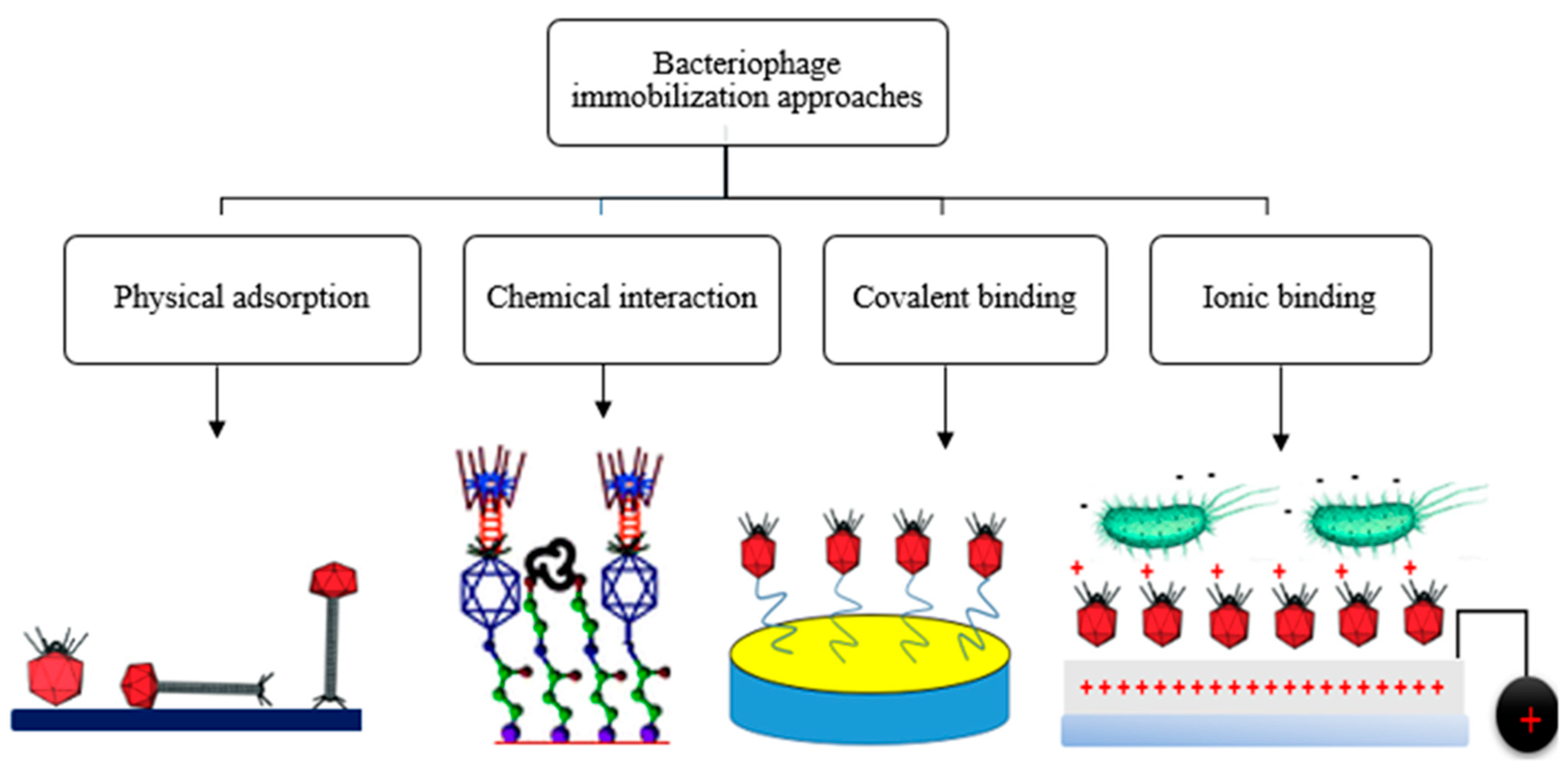
4. Phage Immobilization Strategies
5. Types of Phage-Based Biosensors
5.1. Phage-Based Optical Biosensors
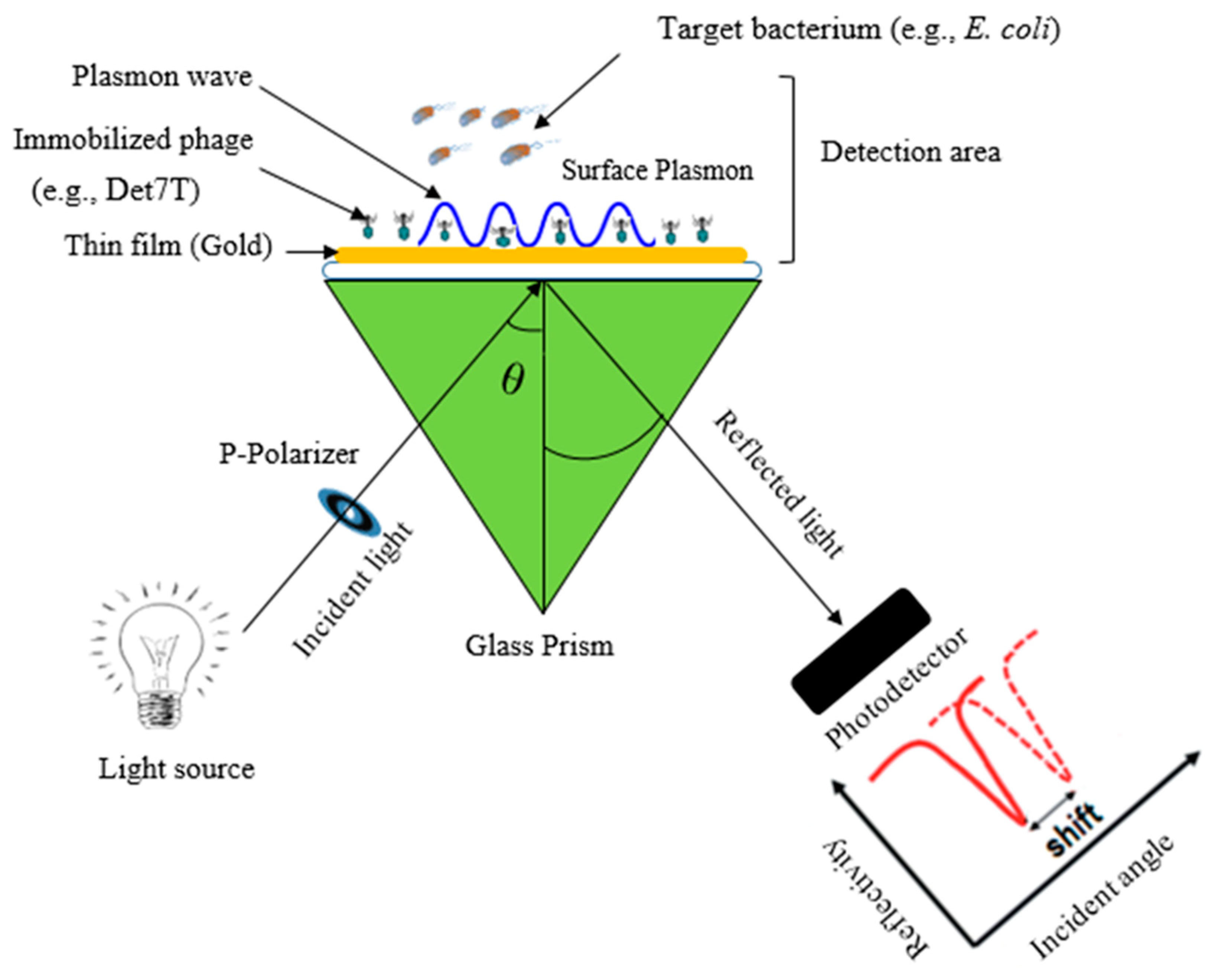
5.1.1. Surface Plasmon Resonance Sensors
5.1.2. Bioluminescence Sensors
5.1.3. Fluorescent Bioassay

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transducer | Host Bacterium | Bio-Receptor (Phage) | LOD CFU/mL | Assay Time | Food Samples | Ref. |
|---|---|---|---|---|---|---|
| SPR sensor | Methicillin-resistant Staphylococcus aureus (MRSA) | BP14 | 103 | NR | NR | [77] |
| Salmonella spp. | P22 | 103 | 3 min | Chicken carcass (wash) | [52] | |
| S. aureus | 12,600 | 104 | NR | NR | [86] | |
| Campylobacter jejuni | NCTC 12,673 TSP | 102 | 45 min | Milk | [87] | |
| E. coli O157:H7 | T4 | 103 | NR | Skim milk | [88] | |
| E. coli K12 | T4 | 7 × 102 | NR | Skim milk | [89] | |
| L. monocytogenes | scFv | 2 × 106 | NR | NR | [90] | |
| S. aureus | 12,600 | 104 | NR | NR | [86] | |
| Bioluminescence sensor | E. coli G2-2 | AT20 | 103 | NR | NR | [42] |
| E. coli | E. coli phage | 103 | 60 min | NR | [91] | |
| Salmonella Newport | Newport (Felix) phage | 103 | NR | NR | [91] | |
| Salmonella Enteritidis | SJ2 | 103 | 120 min | NR | [42] | |
| E. coli | Wild-type and modified T4 | 6 × 103 | NR | NR | [39] | |
| Yersinia pestis | Phage A1122 with lux tag | 102 | NR | NR | [92] | |
| E. coli B | lacZ T4 phage | 10 | NR | Water | [93] | |
| P. aeruginosa | Pap1 | 56 | NR | Milk | [94] | |
| S. flexneri | Shfl25875 | 103 CFU/g | NR | NR | [95] | |
| Fluorescent bioassay | E. coli | T7 | 20 | NR | NR | [30] |
| E. coli | QD-labeled lambda phage | ND | NR | NR | [96] | |
| Staphylococcal enterotoxin B (SEB) | phage-displayed peptides | 1.4 ng | NR | NR | [97] | |
| E. coli O157:H7 | PP01 | 1 | NR | Apple juice | [98] | |
| S. aureus | P-S. aureus-9 | 2.47 × 103 | NR | NR | [99] | |
| S. Typhimurium | P22 | 1 CFU/24 mL | NR | Milk | [100] | |
| B. anthracis | Wβ | 104 CFU/g | NR | NR | [101] | |
| E. coli TD2158 | HK620 | 102–104 | NR | NR | [85] | |
| QCM-based assays | Salmonella Typhimurium | Filamentous phage | 102 | 3 min | NR | [102] |
| M. tuberculosis and M. smegmatis | D29 | 103 | NR | NR | [103] | |
| S. aureus | 12,600 | 104 | NR | NR | [104] | |
| E. coli | T4 | NR | NR | Milk | [16] | |
| E. coli K12 | Wild type | 103 | NR | NR | [105] |
5.2. Phage-Based Electrochemical Biosensors
5.2.1. Amperometric Biosensors
5.2.2. Electrochemical Impedance Spectroscopy (EIS) Biosensors
| Transducer | Phage | Host Bacterium | Food Samples | LOD (CFU/mL) | Ref. |
|---|---|---|---|---|---|
| Impedimetric Sensors | T4 | E. coli K12 | NR | 104 | [117] |
| T4 | E. coli K12 | NR | 104 | [118] | |
| T4 | E. coli K12 | NR | 103 | [105] | |
| Gamma phage | B. anthracis | Water | 103 | [119] | |
| T4 | E. coli B | Water | 8.0 × 102 | [115] | |
| Specific phage | S. arlettae | NR | 2 | [116] | |
| T4 | E. coli K12 | NR | 102 | [120] | |
| T2 | E. coli B | NR | 103 | [121] | |
| CBD | Listeria | NR | 1.1 × 104 | [122] | |
| Endolysin Ply500 | L. monocytogenes | Milk | 105 | [122] | |
| Lytic phage | Salmonella Newport | NR | 103 | [123] | |
| Amperometric Biosensors | T4 | E. coli | NR | 1 | [124] |
| Phage lambda | E. coli | NR | 1 CFU/100 mL | [33] | |
| M13 | E. coli TG1 | NR | 1 | [125] | |
| B1-7064 | B. cereus | NR | 10 | [34] | |
| D29 | M. smegmatis | NR | 10 | [34] | |
| T7 | E. coli | NR | 102 | [126] |
5.3. Micromechanical Biosensors
Phage-Based Quartz Crystal Microbalance Assays
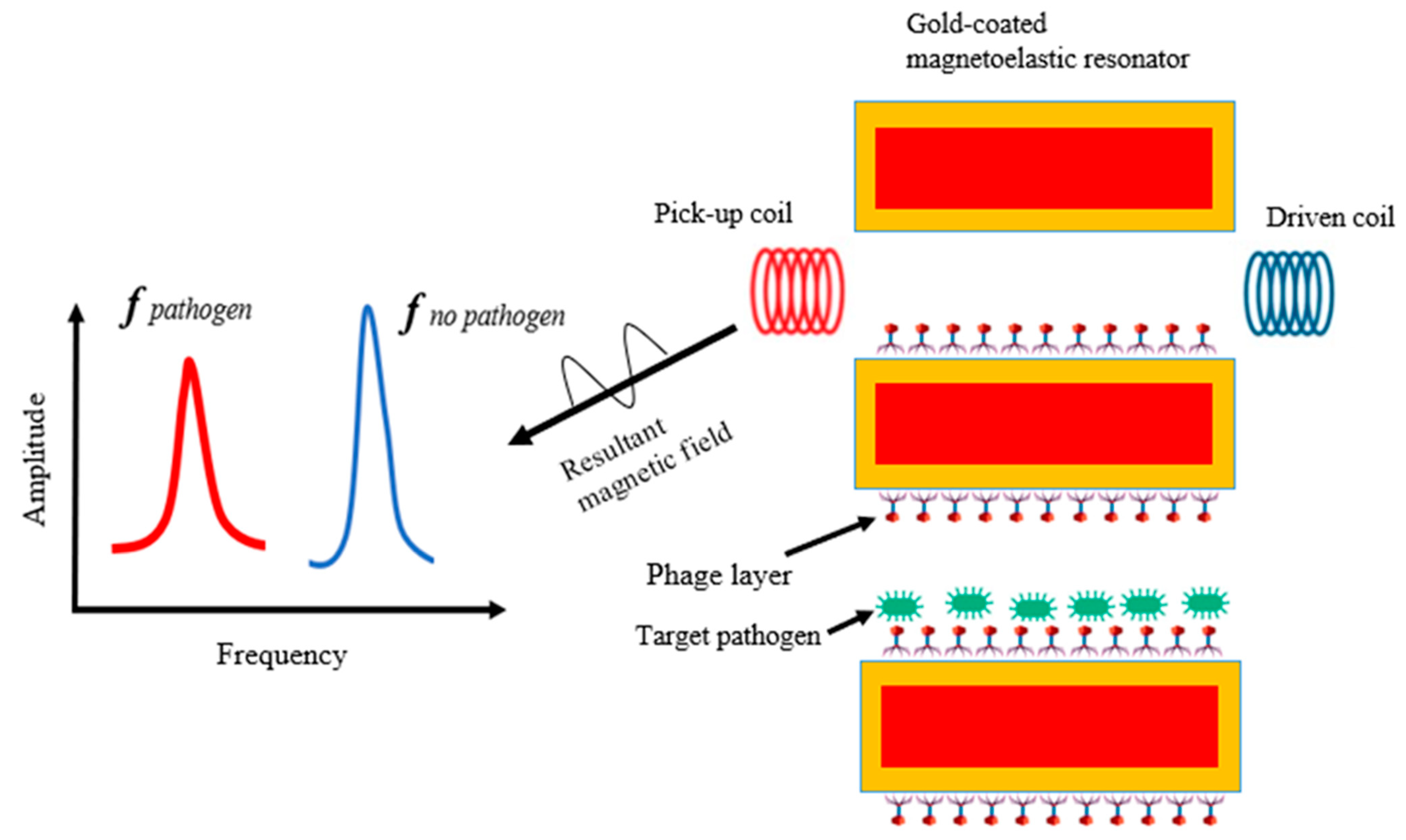
5.4. Phage-Based Magnetoelastic Biosensor
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on major food-borne zoonotic bacterial pathogens. J. Trop. Med. 2020, 2020, 4674235. [Google Scholar] [CrossRef]
- Bintsis, T. Foodborne pathogens. AIMS Microbiol. 2017, 3, 529–563. [Google Scholar] [CrossRef]
- Dhama, K.; Rajagunalan, S.; Chakraborty, S.; Verma, A.K.; Kumar, A.; Tiwari, R.; Kapoor, S. Food-borne pathogens of animal origin-diagnosis, prevention, control and their zoonotic significance: A review. Pak. J. Biol. Sci. 2013, 16, 1076–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellberg, R.S.; Chu, E. Effects of climate change on the persistence and dispersal of foodborne bacterial pathogens in the outdoor environment: A review. Crit. Rev. Microbiol. 2016, 42, 548–572. [Google Scholar] [CrossRef]
- Crim, S.M.; Iwamoto, M.; Huang, J.Y.; Griffin, P.M.; Gilliss, D.; Cronquist, A.B.; Cartter, M.; Tobin-D’Angelo, M.; Blythe, D.; Smith, K. Incidence and trends of infection with pathogens transmitted commonly through food—Foodborne Diseases Active Surveillance Network, 10 US sites, 2006–2013. Morb. Mortal. Wkly. Rep. 2014, 63, 328–332. [Google Scholar]
- Bhunia, A.K. One day to one hour: How quickly can foodborne pathogens be detected? Fut. Microbiol. 2014, 9, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Bai, X.; Bhunia, A.K. Current State of Biosensors Development and their Application in Foodborne Pathogen Detection. J. Food Prot. 2021, 84, 1213–1227. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Lin, C.-W.; Wang, J.; Oh, D.H. Advances in rapid detection methods for foodborne pathogens. J. Microbiol. Biotechnol. 2014, 24, 297–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogaça, M.B.T.; Bhunia, A.K.; Lopes-Luz, L.; de Almeida, E.P.R.P.; Vieira, J.D.G.; Bührer-Sékula, S. Antibody- and nucleic acid–based lateral flow immunoassay for Listeria monocytogenes detection. Anal. Bioanal. Chem. 2021, 413, 4161–4180. [Google Scholar] [CrossRef]
- Lopes-Luz, L.; Mendonça, M.; Bernardes Fogaça, M.; Kipnis, A.; Bhunia, A.K.; Bührer-Sékula, S. Listeria monocytogenes: Review of pathogenesis and virulence determinants-targeted immunological assays. Crit. Rev. Microbiol. 2021, 47, 647–666. [Google Scholar] [CrossRef]
- Ferone, M.; Gowen, A.; Fanning, S.; Scannell, A.G.M. Microbial detection and identification methods: Bench top assays to omics approaches. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3106–3129. [Google Scholar] [CrossRef]
- Aliakbar Ahovan, Z.; Hashemi, A.; De Plano, L.M.; Gholipourmalekabadi, M.; Seifalian, A. Bacteriophage based biosensors: Trends, outcomes and challenges. Nanomaterials 2020, 10, 501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, N.; Bhunia, A.K. Food-Associated Stress Primes Foodborne Pathogens for the Gastrointestinal Phase of Infection. Front. Microbiol. 2018, 9, 1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meile, S.; Kilcher, S.; Loessner, M.J.; Dunne, M. Reporter phage-based detection of bacterial pathogens: Design guidelines and recent developments. Viruses 2020, 12, 944. [Google Scholar] [CrossRef] [PubMed]
- Quintela, I.A.; Wu, V.C.H. A sandwich-type bacteriophage-based amperometric biosensor for the detection of Shiga toxin-producing Escherichia coli serogroups in complex matrices. RSC Adv. 2020, 10, 35765–35775. [Google Scholar] [CrossRef] [PubMed]
- Martelet, A.; L’Hostis, G.; Nevers, M.-C.; Volland, H.; Junot, C.; Becher, F.; Muller, B.H. Phage amplification and immunomagnetic separation combined with targeted mass spectrometry for sensitive detection of viable bacteria in complex food matrices. Anal. Chem. 2015, 87, 5553–5560. [Google Scholar] [CrossRef]
- Choi, I.Y.; Lee, C.; Song, W.K.; Jang, S.J.; Park, M.-K. Lytic KFS-SE2 phage as a novel bio-receptor for Salmonella Enteritidis detection. J. Microbiol. 2019, 57, 170–179. [Google Scholar] [CrossRef]
- Choi, I.Y.; Park, J.H.; Gwak, K.M.; Kim, K.-P.; Oh, J.-H.; Park, M.-K. Studies on lytic, tailed Bacillus cereus-specific phage for use in a ferromagnetoelastic biosensor as a novel recognition element. J. Microbiol. Biotechnol. 2018, 28, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, Y.; Fan, T.; Wang, P.; Zhang, X.; Luo, J.; Zhou, F.; Yao, Y.; Liao, X.; Li, Y.; Gao, F. A novel label-free homogeneous electrochemical immunosensor based on proximity hybridization-triggered isothermal exponential amplification induced G-quadruplex formation. Sens. Actuators B Chem. 2017, 248, 187–194. [Google Scholar] [CrossRef]
- Hinkley, T.C.; Garing, S.; Singh, S.; Le Ny, A.-L.M.; Nichols, K.P.; Peters, J.E.; Talbert, J.N.; Nugen, S.R. Reporter bacteriophage T7 NLC utilizes a novel NanoLuc: CBM fusion for the ultrasensitive detection of Escherichia coli in water. Analyst 2018, 143, 4074–4082. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Kanach, A.; Han, R.; Applegate, B. Application of bacteriophage in rapid detection of Escherichia coli in foods. Curr. Opin. Food Sci. 2021, 39, 43–50. [Google Scholar] [CrossRef]
- Farrokhzad, K.; Rosenfield, C.; Applegate, B. Phage technology in high throughput screening for pathogen detection in food. In High Throughput Screening for Food Safety Assessment; Bhunia, A.K., Kim, M.S., Taitt, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 81–121. [Google Scholar]
- Peltomaa, R.; López-Perolio, I.; Benito-Peña, E.; Barderas, R.; Moreno-Bondi, M.C. Application of bacteriophages in sensor development. Anal. Bioanal. Chem. 2016, 408, 1805–1828. [Google Scholar] [CrossRef] [PubMed]
- Waseh, S.; Hanifi-Moghaddam, P.; Coleman, R.; Masotti, M.; Ryan, S.; Foss, M.; MacKenzie, R.; Henry, M.; Szymanski, C.M.; Tanha, J. Orally administered P22 phage tailspike protein reduces Salmonella colonization in chickens: Prospects of a novel therapy against bacterial infections. PLoS ONE 2010, 5, e13904. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chau, Y.; Lee, Y.-k. Phage-based electrochemical sensors: A review. Micromachines 2019, 10, 855. [Google Scholar] [CrossRef] [Green Version]
- Bárdy, P.; Pantůček, R.; Benešík, M.; Doškař, J. Genetically modified bacteriophages in applied microbiology. J. Appl. Microbiol. 2016, 121, 618–633. [Google Scholar] [CrossRef] [PubMed]
- Alcaine, S.; Law, K.; Ho, S.; Kinchla, A.; Sela, D.; Nugen, S. Bioengineering bacteriophages to enhance the sensitivity of phage amplification-based paper fluidic detection of bacteria. Biosens. Bioelectron. 2016, 82, 14–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, W.; Ullah, M.W.; Farooq, U.; Aziz, A.; Wang, S. Bacteriophage-based advanced bacterial detection: Concept, mechanisms, and applications. Biosens. Bioelectron. 2021, 177, 112973. [Google Scholar] [CrossRef]
- Casey, A.; Coffey, A.; McAuliffe, O. Genetics and genomics of bacteriophages: The evolution of bacteriophage genomes and genomic research. Bacteriophages Biol. Technol. Ther. 2021, 193–218. [Google Scholar] [CrossRef]
- Edgar, R.; McKinstry, M.; Hwang, J.; Oppenheim, A.B.; Fekete, R.A.; Giulian, G.; Merril, C.; Nagashima, K.; Adhya, S. High-sensitivity bacterial detection using biotin-tagged phage and quantum-dot nanocomplexes. Proc. Natl. Acad. Sci. USA 2006, 103, 4841–4845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakshmanan, R.S.; Guntupalli, R.; Hu, J.; Kim, D.-J.; Petrenko, V.A.; Barbaree, J.M.; Chin, B.A. Phage immobilized magnetoelastic sensor for the detection of Salmonella Typhimurium. J. Microbiol. Methods 2007, 71, 55–60. [Google Scholar] [CrossRef]
- Anany, H.; Brovko, L.; El Dougdoug, N.K.; Sohar, J.; Fenn, H.; Alasiri, N.; Jabrane, T.; Mangin, P.; Ali, M.M.; Kannan, B.; et al. Print to detect: A rapid and ultrasensitive phage-based dipstick assay for foodborne pathogens. Anal. Bioanal. Chem. 2018, 410, 1217–1230. [Google Scholar] [CrossRef]
- Neufeld, T.; Schwartz-Mittelmann, A.; Biran, D.; Ron, E.Z.; Rishpon, J. Combined phage typing and amperometric detection of released enzymatic activity for the specific identification and quantification of bacteria. Anal. Chem. 2003, 75, 580–585. [Google Scholar] [CrossRef]
- Yemini, M.; Levi, Y.; Yagil, E.; Rishpon, J. Specific electrochemical phage sensing for Bacillus cereus and Mycobacterium smegmatis. Bioelectrochemistry 2007, 70, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Hinkley, T.C.; Singh, S.; Garing, S.; Le Ny, A.-L.M.; Nichols, K.P.; Peters, J.E.; Talbert, J.N.; Nugen, S.R. A phage-based assay for the rapid, quantitative, and single CFU visualization of E. coli (ECOR# 13) in drinking water. Sci. Rep. 2018, 8, 14630. [Google Scholar] [PubMed] [Green Version]
- Rees, C.E.; Dodd, C.E. Phage for rapid detection and control of bacterial pathogens in food. Adv. Appl. Microbiol. 2006, 59, 159–186. [Google Scholar] [PubMed]
- Squirrell, D.; Price, R.; Murphy, M. Rapid and specific detection of bacteria using bioluminescence. Anal. Chim. Acta 2002, 457, 109–114. [Google Scholar] [CrossRef]
- Brovko, L.Y.; Anany, H.; Griffiths, M.W. Bacteriophages for detection and control of bacterial pathogens in food and food-processing environment. Adv. Food Nutr. Res. 2012, 67, 241–288. [Google Scholar] [PubMed]
- Minikh, O.; Tolba, M.; Brovko, L.; Griffiths, M. Bacteriophage-based biosorbents coupled with bioluminescent ATP assay for rapid concentration and detection of Escherichia coli. J. Microbiol. Methods 2010, 82, 177–183. [Google Scholar] [CrossRef]
- Alcaine, S.D.; Pacitto, D.; Sela, D.A.; Nugen, S.R. Phage & phosphatase: A novel phage-based probe for rapid, multi-platform detection of bacteria. Analyst 2015, 140, 7629–7636. [Google Scholar]
- Corbitt, A.; Bennion, N.; Forsythe, S. Adenylate kinase amplification of ATP bioluminescence for hygiene monitoring in the food and beverage industry. Lett. Appl. Microbiol. 2000, 30, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Brovko, L.; Griffiths, M. Influence of phage population on the phage-mediated bioluminescent adenylate kinase (AK) assay for detection of bacteria. Lett. Appl. Microbiol. 2001, 33, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Kannan, P.; Yong, H.Y.; Reiman, L.; Cleaver, C.; Patel, P.; Bhagwat, A.A. Bacteriophage-based rapid and sensitive detection of Escherichia coli O157: H7 isolates from ground beef. Foodborne Pathog. Dis. 2010, 7, 1551–1558. [Google Scholar] [CrossRef]
- Chang, T.C.; Ding, H.C.; Chen, S. A conductance method for the identification of Escherichia coli O157: H7 using bacteriophage AR1. J. Food Prot. 2002, 65, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Guła, G.; Szymanowska, P.; Piasecki, T.; Góras, S.; Gotszalk, T.; Drulis-Kawa, Z. The application of impedance spectroscopy for Pseudomonas biofilm monitoring during phage infection. Viruses 2020, 12, 407. [Google Scholar] [CrossRef] [Green Version]
- Falahee, M.B.; Park, S.F.; Adams, M.R. Detection and enumeration of Campylobacter jejuni and Campylobacter coli by indirect impedimetry with an oxygen scavenging system. J. Food Prot. 2003, 66, 1724–1726. [Google Scholar] [CrossRef]
- Stewart, G.S.; Jassim, S.A.; Denyer, S.P.; Newby, P.; Linley, K.; Dhir, V.K. The specific and sensitive detection of bacterial pathogens within 4 h using bacteriophage amplification. J. Appl. Microbiol. 1998, 84, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.R.; Jensen, K.R.; Mondesire, R.R.; Voorhees, K.J. Rapid detection of Bacillus anthracis by γ phage amplification and lateral flow immunochromatography. J. Microbiol. Methods 2015, 118, 51–56. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, L.; Bolton, D.; McAuliffe, O.; Coffey, A. Bacteriophages in food applications: From foe to friend. Annu. Rev. Food Sci. Technol. 2019, 10, 151–172. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Maestu, A.; Fuciños, P.; Azinheiro, S.; Carvalho, C.; Carvalho, J.; Prado, M. Specific detection of viable Salmonella Enteritidis by phage amplification combined with qPCR (PAA-qPCR) in spiked chicken meat samples. Food Control. 2019, 99, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, A.A.; Wollin, R.; Gemski, P.; Wohlhieter, J.A. Interaction between bacteriophage Sf6 and Shigella flexneri. J. Virol. 1978, 27, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Arya, S.K.; Glass, N.; Hanifi-Moghaddam, P.; Naidoo, R.; Szymanski, C.M.; Tanha, J.; Evoy, S. Bacteriophage tailspike proteins as molecular probes for sensitive and selective bacterial detection. Biosens. Bioelectron. 2010, 26, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Stone, E.; Campbell, K.; Grant, I.; McAuliffe, O. Understanding and exploiting phage–host interactions. Viruses 2019, 11, 567. [Google Scholar] [CrossRef] [PubMed]
- Wisuthiphaet, N.; Yang, X.; Young, G.M.; Nitin, N. Rapid detection of Escherichia coli in beverages using genetically engineered bacteriophage T7. AMB Express 2019, 9, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, M.; Kim, S.; Ryu, S. Sensitive detection of viable Escherichia coli O157:H7 from foods using a luciferase-reporter phage phiV10lux. Int. J. Food Microbiol. 2017, 254, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Coronel-Aguilera, C.P.; Romero, P.L.; Perry, L.; Minocha, U.; Rosenfield, C.; Gehring, A.G.; Paoli, G.C.; Bhunia, A.K.; Applegate, B. The use of a novel nanoLuc -based reporter phage for the detection of Escherichia coli O157:H7. Sci. Rep. 2016, 6, 33235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinkley, T.C.; Garing, S.; Jain, P.; Williford, J.; Le Ny, A.-L.M.; Nichols, K.P.; Peters, J.E.; Talbert, J.N.; Nugen, S.R. A Syringe-based biosensor to rapidly detect low levels of Escherichia coli (ECOR13) in drinking water using engineered bacteriophages. Sensors 2020, 20, 1953. [Google Scholar] [CrossRef] [Green Version]
- Zurier, H.S.; Duong, M.M.; Goddard, J.M.; Nugen, S.R. Engineering Biorthogonal Phage-Based Nanobots for Ultrasensitive, In Situ Bacteria Detection. ACS Appl. Bio Mater. 2020, 3, 5824–5831. [Google Scholar] [CrossRef]
- Chen, A.; Wang, D.; Nugen, S.R.; Chen, J. An Engineered Reporter Phage for the Fluorometric Detection of Escherichia coli in Ground Beef. Microorganisms 2021, 9, 436. [Google Scholar] [CrossRef]
- Santos, S.B.; Cunha, A.P.; Macedo, M.; Nogueira, C.L.; Brandão, A.; Costa, S.P.; Melo, L.D.R.; Azeredo, J.; Carvalho, C.M. Bacteriophage-receptor binding proteins for multiplex detection of Staphylococcus and Enterococcus in blood. Biotechnol. Bioeng. 2020, 117, 3286–3298. [Google Scholar] [CrossRef]
- Sumrall, E.T.; Keller, A.P.; Shen, Y.; Loessner, M.J. Structure and function of Listeria teichoic acids and their implications. Mol. Microbiol. 2020, 113, 627–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunstmann, S.; Scheidt, T.; Buchwald, S.; Helm, A.; Mulard, L.A.; Fruth, A.; Barbirz, S. Bacteriophage Sf6 tailspike protein for detection of Shigella flexneri pathogens. Viruses 2018, 10, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denyes, J.M.; Dunne, M.; Steiner, S.; Mittelviefhaus, M.; Weiss, A.; Schmidt, H.; Klumpp, J.; Loessner, M.J. Modified bacteriophage S16 long tail fiber proteins for rapid and specific immobilization and detection of Salmonella cells. Appl. Environ. Microbiol. 2017, 83, e00277-17. [Google Scholar] [CrossRef]
- He, Y.; Shi, Y.; Liu, M.; Wang, Y.; Wang, L.; Lu, S.; Fu, Z. Nonlytic recombinant phage tail fiber protein for specific recognition of Pseudomonas aeruginosa. Anal. Chem. 2018, 90, 14462–14468. [Google Scholar] [CrossRef]
- Poshtiban, S.; Javed, M.A.; Arutyunov, D.; Singh, A.; Banting, G.; Szymanski, C.M.; Evoy, S. Phage receptor binding protein-based magnetic enrichment method as an aid for real time PCR detection of foodborne bacteria. Analyst 2013, 138, 5619–5626. [Google Scholar] [CrossRef]
- Sumrall, E.T.; Röhrig, C.; Hupfeld, M.; Selvakumar, L.; Du, J.; Dunne, M.; Schmelcher, M.; Shen, Y.; Loessner, M.J. Glycotyping and specific separation of Listeria monocytogenes with a novel bacteriophage protein tool kit. Appl. Environ. Microbiol. 2020, 86, e00612–e00620. [Google Scholar] [CrossRef]
- Kong, M.; Sim, J.; Kang, T.; Nguyen, H.H.; Park, H.k.; Chung, B.H.; Ryu, S. A novel and highly specific phage endolysin cell wall binding domain for detection of Bacillus cereus. Eur. Biophys. J. 2015, 44, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Torres, N.; Ávila, M.; Narbad, A.; Mayer, M.J.; Garde, S. Use of fluorescent CTP1L endolysin cell wall-binding domain to study the evolution of Clostridium tyrobutyricum during cheese ripening. Food Microbiol. 2019, 78, 11–17. [Google Scholar] [CrossRef] [PubMed]
- O’connell, L.; Marcoux, P.R.; Roupioz, Y. Strategies for surface immobilization of whole bacteriophages: A review. ACS Biomater. Sci. Eng. 2021, 7, 1987–2014. [Google Scholar] [CrossRef]
- Richter, Ł.; Księżarczyk, K.; Paszkowska, K.; Janczuk-Richter, M.; Niedziółka-Jönsson, J.; Gapiński, J.; Łoś, M.; Hołyst, R.; Paczesny, J. Adsorption of bacteriophages on polypropylene labware affects the reproducibility of phage research. Sci. Rep. 2021, 11, 7387. [Google Scholar] [CrossRef]
- Tawil, N.; Sacher, E.; Mandeville, R.; Meunier, M. Strategies for the immobilization of bacteriophages on gold surfaces monitored by surface plasmon resonance and surface morphology. J. Phys. Chem. C 2013, 117, 6686–6691. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, C.; Chau, Y.; Lee, Y.-K. The synergy of chemical immobilization and electrical orientation of T4 bacteriophage on a micro electrochemical sensor for low-level viable bacteria detection via differential pulse voltammetry. Biosens. Bioelectron. 2020, 151, 111914. [Google Scholar] [CrossRef] [PubMed]
- Karoonuthaisiri, N.; Charlermroj, R.; Morton, M.J.; Oplatowska-Stachowiak, M.; Grant, I.R.; Elliott, C.T. Development of a M13 bacteriophage-based SPR detection using Salmonella as a case study. Sens. Actuators B Chem. 2014, 190, 214–220. [Google Scholar] [CrossRef]
- Huertas, C.S.; Calvo-Lozano, O.; Mitchell, A.; Lechuga, L.M. Advanced evanescent-wave optical biosensors for the detection of nucleic acids: An analytic perspective. Front. Chem. 2019, 7, 724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vachali, P.P.; Li, B.; Bartschi, A.; Bernstein, P.S. Surface plasmon resonance (SPR)-based biosensor technology for the quantitative characterization of protein–carotenoid interactions. Arch. Biochem. Biophys. 2015, 572, 66–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.J.; Lim, W.K. Rapid label-free detection of E. coli using a novel SPR biosensor containing a fragment of tail protein from phage lambda. Prep. Biochem. Biotechnol. 2018, 48, 498–505. [Google Scholar] [CrossRef]
- Tawil, N.; Sacher, E.; Mandeville, R.; Meunier, M. Surface plasmon resonance detection of E. coli and methicillin-resistant S. aureus using bacteriophages. Biosens. Bioelectron. 2012, 37, 24–29. [Google Scholar] [CrossRef]
- Singh, A.; Arutyunov, D.; Szymanski, C.M.; Evoy, S. Bacteriophage based probes for pathogen detection. Analyst 2012, 137, 3405–3421. [Google Scholar] [CrossRef]
- Chuang, T.-L.; Wei, S.-C.; Lee, S.-Y.; Lin, C.-W. A polycarbonate based surface plasmon resonance sensing cartridge for high sensitivity HBV loop-mediated isothermal amplification. Biosens. Bioelectron. 2012, 32, 89–95. [Google Scholar] [CrossRef]
- Hyeon, S.H.; Lim, W.K.; Shin, H.J. Novel surface plasmon resonance biosensor that uses full-length Det7 phage tail protein for rapid and selective detection of Salmonella enterica serovar Typhimurium. Biotechnol. Appl. Biochem. 2021, 68, 5–12. [Google Scholar] [CrossRef]
- Kozak, S.; Alcaine, S.D. Phage-based forensic tool for spatial visualization of bacterial contaminants in cheese. J. Dairy Sci. 2020, 103, 5964–5971. [Google Scholar] [CrossRef]
- Kim, S.; Kim, M.; Ryu, S. Development of an engineered bioluminescent reporter phage for the sensitive detection of viable Salmonella Typhimurium. Anal. Chem. 2014, 86, 5858–5864. [Google Scholar] [CrossRef] [PubMed]
- Franche, N.; Vinay, M.; Ansaldi, M. Substrate-independent luminescent phage-based biosensor to specifically detect enteric bacteria such as E. coli. Environ. Sci. Pollut. Res. 2017, 24, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Onuki, M.; Satoh, H.; Mino, T. Isolation, characterization of bacteriophages specific to Microlunatus phosphovorus and their application for rapid host detection. Lett. Appl. Microbiol. 2006, 42, 259–264. [Google Scholar] [CrossRef]
- Vinay, M.; Franche, N.; Grégori, G.; Fantino, J.-R.; Pouillot, F.; Ansaldi, M. Phage-based fluorescent biosensor prototypes to specifically detect enteric bacteria such as E. coli and Salmonella enterica Typhimurium. PLoS ONE 2015, 10, e0131466. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Sorokulova, I.B.; Vodyanoy, V.J.; Simonian, A.L. Lytic phage as a specific and selective probe for detection of Staphylococcus aureus—A surface plasmon resonance spectroscopic study. Biosens. Bioelectron. 2007, 22, 948–955. [Google Scholar] [CrossRef]
- Singh, A.; Arutyunov, D.; McDermott, M.T.; Szymanski, C.M.; Evoy, S. Specific detection of Campylobacter jejuni using the bacteriophage NCTC 12673 receptor binding protein as a probe. Analyst 2011, 136, 4780–4786. [Google Scholar] [CrossRef]
- Wang, L.; Shi, L.; Su, J.; Ye, Y.; Zhong, Q. Detection of Vibrio parahaemolyticus in food samples using in situ loop-mediated isothermal amplification method. Gene 2013, 515, 421–425. [Google Scholar] [CrossRef]
- Arya, S.K.; Singh, A.; Naidoo, R.; Wu, P.; McDermott, M.T.; Evoy, S. Chemically immobilized T4-bacteriophage for specific Escherichia coli detection using surface plasmon resonance. Analyst 2011, 136, 486–492. [Google Scholar] [CrossRef]
- Nanduri, V.; Bhunia, A.K.; Tu, S.-I.; Paoli, G.C.; Brewster, J.D. SPR biosensor for the detection of L. monocytogenes using phage-displayed antibody. Biosens. Bioelectron. 2007, 23, 248–252. [Google Scholar] [CrossRef]
- Blasco, R.; Murphy, M.; Sanders, M.; Squirrell, D. Specific assays for bacteria using phage mediated release of adenylate kinase. J. Appl. Microbiol. 1998, 84, 661–666. [Google Scholar] [CrossRef]
- Schofield, D.A.; Molineux, I.J.; Westwater, C. Diagnostic bioluminescent phage for detection of Yersinia pestis. J. Clin. Microbiol. 2009, 47, 3887–3894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnham, S.; Hu, J.; Anany, H.; Brovko, L.; Deiss, F.; Derda, R.; Griffiths, M.W. Towards rapid on-site phage-mediated detection of generic Escherichia coli in water using luminescent and visual readout. Anal. Bioanal. Chem. 2014, 406, 5685–5693. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; He, Y.; Fan, E.; Wang, L.; Lu, S.; Fu, Z. Label-free electrochemiluminescent biosensor for rapid and sensitive detection of Pseudomonas aeruginosa using phage as highly specific recognition agent. Biosens. Bioelectron. 2017, 94, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Schofield, D.A.; Wray, D.J.; Molineux, I.J. Isolation and development of bioluminescent reporter phages for bacterial dysentery. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Yim, P.B.; Clarke, M.L.; McKinstry, M.; De Paoli Lacerda, S.H.; Pease, L.F.; Dobrovolskaia, M.A.; Kang, H.; Read, T.D.; Sozhamannan, S.; Hwang, J. Quantitative characterization of quantum dot-labeled lambda phage for Escherichia coli detection. Biotechnol. Bioeng. 2009, 104, 1059–1067. [Google Scholar] [CrossRef]
- Goldman, E.R.; Pazirandeh, M.P.; Mauro, J.M.; King, K.D.; Frey, J.C.; Anderson, G.P. Phage-displayed peptides as biosensor reagents. J. Mol. Recognit. 2000, 13, 382–387. [Google Scholar] [CrossRef]
- Hoang, H.A.; Dien, L.T. Rapid and simple colorimetric detection of Escherichia coli O157: H7 in apple juice using a novel recombinant bacteriophage-based method. Biocontrol Sci. 2015, 20, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Zhang, Y.; Yang, H.; Yu, J.; Wei, H. Combining phagomagnetic separation with immunoassay for specific, fast and sensitive detection of Staphylococcus aureus. Talanta 2017, 170, 291–297. [Google Scholar] [CrossRef]
- Laube, T.; Cortés, P.; Llagostera, M.; Alegret, S.; Pividori, M.I. Phagomagnetic immunoassay for the rapid detection of Salmonella. Appl. Microbiol. Biotechnol. 2014, 98, 1795–1805. [Google Scholar] [CrossRef]
- Sharp, N.J.; Molineux, I.J.; Page, M.A.; Schofield, D.A. Rapid detection of viable Bacillus anthracis spores in environmental samples by using engineered reporter phages. Appl. Environ. Microbiol. 2016, 82, 2380–2387. [Google Scholar] [CrossRef] [Green Version]
- Olsen, E.V.; Sorokulova, I.B.; Petrenko, V.A.; Chen, I.-H.; Barbaree, J.M.; Vodyanoy, V.J. Affinity-selected filamentous bacteriophage as a probe for acoustic wave biodetectors of Salmonella Typhimurium. Biosens. Bioelectron. 2006, 21, 1434–1442. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Xiang, M.; Mi, X. A new bacteriophage-modified piezoelectric sensor for rapid and specific detection of mycobacterium. Anal. Lett. 2012, 45, 1242–1253. [Google Scholar] [CrossRef]
- Guntupalli, R.; Sorokulova, I.; Olsen, E.; Globa, L.; Pustovyy, O.; Moore, T.; Chin, B.; Barbaree, J.; Vodyanoy, V. Detection and identification of methicillin resistant and sensitive strains of Staphylococcus aureus using tandem measurements. J. Microbiol. Methods 2012, 90, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Shabani, A.; Marquette, C.A.; Mandeville, R.; Lawrence, M.F. Magnetically-assisted impedimetric detection of bacteria using phage-modified carbon microarrays. Talanta 2013, 116, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Richter, Ł.; Matuła, K.; Leśniewski, A.; Kwaśnicka, K.; Łoś, J.; Łoś, M.; Paczesny, J.; Hołyst, R. Ordering of bacteriophages in the electric field: Application for bacteria detection. Sens. Actuators B Chem. 2016, 224, 233–240. [Google Scholar] [CrossRef]
- Richter, Ł.; Bielec, K.; Leśniewski, A.; Łoś, M.; Paczesny, J.; Hołyst, R. Dense layer of bacteriophages ordered in alternating electric field and immobilized by surface chemical modification as sensing element for bacteria detection. ACS Appl. Mater. Interfaces 2017, 9, 19622–19629. [Google Scholar] [CrossRef]
- Ronkainen, N.J.; Halsall, H.B.; Heineman, W.R. Electrochemical biosensors. Chem. Soc. Rev. 2010, 39, 1747–1763. [Google Scholar] [CrossRef]
- Wang, J. Glucose biosensors: 40 years of advances and challenges. Electroanalysis 2001, 13, 983–988. [Google Scholar] [CrossRef]
- Nikkhoo, N.; Gulak, P.G.; Maxwell, K. Rapid detection of E. coli bacteria using potassium-sensitive FETs in CMOS. IEEE Trans. Biomed. Circuits Syst. 2013, 7, 621–630. [Google Scholar] [CrossRef]
- Ivnitski, D.; Abdel-Hamid, I.; Atanasov, P.; Wilkins, E.; Stricker, S. Application of electrochemical biosensors for detection of food pathogenic bacteria. Electroanalysis 2000, 12, 317–325. [Google Scholar] [CrossRef]
- Cinti, S.; Volpe, G.; Piermarini, S.; Delibato, E.; Palleschi, G. Electrochemical biosensors for rapid detection of foodborne Salmonella: A critical overview. Sensors 2017, 17, 1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, S.; Alocilja, E.C.; Downes, F.P. Nanowire labeled direct-charge transfer biosensor for detecting Bacillus species. Biosens. Bioelectron. 2007, 22, 2329–2336. [Google Scholar] [CrossRef] [PubMed]
- Webster, M.; Timoshkin, I.V.; MacGregor, S.J.; Mattey, M. Computer aided modelling of an interdigitated microelectrode array impedance biosensor for the detection of bacteria. IEEE Trans. Dielectr. Electr. Insul. 2009, 16, 1356–1363. [Google Scholar] [CrossRef]
- Tlili, C.; Sokullu, E.; Safavieh, M.; Tolba, M.; Ahmed, M.U.; Zourob, M. Bacteria screening, viability, and confirmation assays using bacteriophage-impedimetric/loop-mediated isothermal amplification dual-response biosensors. Anal. Chem. 2013, 85, 4893–4901. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Bhardwaj, S.K.; Mehta, J.; Mohanta, G.C.; Deep, A. Bacteriophage immobilized graphene electrodes for impedimetric sensing of bacteria (Staphylococcus arlettae). Anal. Biochem. 2016, 505, 18–25. [Google Scholar] [CrossRef]
- Shabani, A.; Zourob, M.; Allain, B.; Marquette, C.A.; Lawrence, M.F.; Mandeville, R. Bacteriophage-modified microarrays for the direct impedimetric detection of bacteria. Anal. Chem. 2008, 80, 9475–9482. [Google Scholar] [CrossRef]
- Mejri, M.; Baccar, H.; Baldrich, E.; Del Campo, F.; Helali, S.; Ktari, T.; Simonian, A.; Aouni, M.; Abdelghani, A. Impedance biosensing using phages for bacteria detection: Generation of dual signals as the clue for in-chip assay confirmation. Biosens. Bioelectron. 2010, 26, 1261–1267. [Google Scholar] [CrossRef] [Green Version]
- Shabani, A.; Marquette, C.A.; Mandeville, R.; Lawrence, M.F. Carbon microarrays for the direct impedimetric detection of Bacillus anthracis using Gamma phages as probes. Analyst 2013, 138, 1434–1440. [Google Scholar] [CrossRef]
- Moghtader, F.; Congur, G.; Zareie, H.M.; Erdem, A.; Piskin, E. Impedimetric detection of pathogenic bacteria with bacteriophages using gold nanorod deposited graphite electrodes. RSC Adv. 2016, 6, 97832–97839. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Marar, A.; Kner, P.; Ramasamy, R.P. Charge-directed immobilization of bacteriophage on nanostructured electrode for whole-cell electrochemical biosensors. Anal. Chem. 2017, 89, 5734–5741. [Google Scholar] [CrossRef]
- Tolba, M.; Ahmed, M.U.; Tlili, C.; Eichenseher, F.; Loessner, M.J.; Zourob, M. A bacteriophage endolysin-based electrochemical impedance biosensor for the rapid detection of Listeria cells. Analyst 2012, 137, 5749–5756. [Google Scholar] [CrossRef] [PubMed]
- Dadarwal, R.; Namvar, A.; Thomas, D.; Hall, J.; Warriner, K. Organic conducting polymer electrode based sensors for detection of Salmonella infecting bacteriophages. Mater. Sci. Eng. C 2009, 29, 761–765. [Google Scholar] [CrossRef]
- Li, Y.; Xie, G.; Qiu, J.; Zhou, D.; Gou, D.; Tao, Y.; Li, Y.; Chen, H. A new biosensor based on the recognition of phages and the signal amplification of organic-inorganic hybrid nanoflowers for discriminating and quantitating live pathogenic bacteria in urine. Sens. Actuators B Chem. 2018, 258, 803–812. [Google Scholar] [CrossRef]
- Neufeld, T.; Mittelman, A.S.; Buchner, V.; Rishpon, J. Electrochemical Phagemid Assay for the Specific Detection of Bacteria Using Escherichia coli TG-1 and the M13KO7 Phagemid in a Model System. Anal. Chem. 2005, 77, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, J.; Nugen, S.R. Electrochemical detection of Escherichia coli from aqueous samples using engineered phages. Anal. Chem. 2017, 89, 1650–1657. [Google Scholar] [CrossRef]
- Olsson, A.L.J.; Wargenau, A.; Tufenkji, N. Optimizing bacteriophage surface densities for bacterial capture and sensing in quartz crystal microbalance with dissipation monitoring. ACS Appl. Mater. Interfaces 2016, 8, 13698–13706. [Google Scholar] [CrossRef]
- Burlage, R.S.; Tillmann, J. Biosensors of bacterial cells. J. Microbiol. Methods 2017, 138, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Yang, H.; Lakshmanan, R.S.; Johnson, M.; Wan, J.; Chen, I.-H.; Wikle, H.; Petrenko, V.; Barbaree, J.; Chin, B. Sequential detection of Salmonella Typhimurium and Bacillus anthracis spores using magnetoelastic biosensors. Biosens. Bioelectron. 2009, 24, 1730–1736. [Google Scholar] [CrossRef]
- Park, M.-K.; Weerakoon, K.A.; Oh, J.-H.; Chin, B.A. The analytical comparison of phage-based magnetoelastic biosensor with TaqMan-based quantitative PCR method to detect Salmonella Typhimurium on cantaloupes. Food Control 2013, 33, 330–336. [Google Scholar] [CrossRef]
- Wang, F.; Horikawa, S.; Hu, J.; Wikle, H.C.; Chen, I.-H.; Du, S.; Liu, Y.; Chin, B.A. Detection of Salmonella Typhimurium on spinach using phage-based magnetoelastic biosensors. Sensors 2017, 17, 386. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.-H.; Horikawa, S.; Bryant, K.; Riggs, R.; Chin, B.A.; Barbaree, J.M. Bacterial assessment of phage magnetoelastic sensors for Salmonella enterica Typhimurium detection in chicken meat. Food Control 2017, 71, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Li, Y.; Chen, H.; Horikawa, S.; Shen, W.; Simonian, A.; Chin, B.A. Direct detection of Salmonella Typhimurium on fresh produce using phage-based magnetoelastic biosensors. Biosens. Bioelectron. 2010, 26, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Mack, J.D.; Yehualaeshet, T.; Park, M.-K.; Tameru, B.; Samuel, T.; Chin, B.A. Phage-based biosensor and optimization of surface blocking agents to detect Salmonella Typhimurium on romaine lettuce. J. Food Saf. 2017, 37, e12299. [Google Scholar] [CrossRef]
- Shen, W.; Lakshmanan, R.S.; Mathison, L.C.; Petrenko, V.A.; Chin, B.A. Phage coated magnetoelastic micro-biosensors for real-time detection of Bacillus anthracis spores. Sens. Actuators B Chem. 2009, 137, 501–506. [Google Scholar] [CrossRef]
- Chai, Y.; Horikawa, S.; Li, S.; Wikle, H.C.; Chin, B.A. A surface-scanning coil detector for real-time, in-situ detection of bacteria on fresh food surfaces. Biosens. Bioelectron. 2013, 50, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Farooq, U.; Yang, Q.; Ullah, M.W.; Wang, S. Bacterial biosensing: Recent advances in phage-based bioassays and biosensors. Biosens. Bioelectron. 2018, 118, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Dhanjal, D.S.; Sonali; Thotapalli, S.; Kumar, V.; Datta, S.; Kumar, V.; Kumar, M.; Singh, J. An insight in bacteriophage based biosensors with focus on their detection methods and recent advancements. Environ. Technol. Innov. 2020, 20, 101081. [Google Scholar] [CrossRef]
- Dunne, M.; Hupfeld, M.; Klumpp, J.; Loessner, M.J. Molecular basis of bacterial host interactions by Gram-positive targeting bacteriophages. Viruses 2018, 10, 397. [Google Scholar] [CrossRef] [PubMed]






| Transducer | Host Bacterium | Phage | Food Sample | LOD (CFU/mL) | Assay Time | Ref. |
|---|---|---|---|---|---|---|
| Magnetoelastic | S. Typhimurium | E2 | NR | 5 × 102 | NR | [133] |
| S. Typhimurium | E2 | Romaine lettuce | 5 × 102 | NR | [134] | |
| B. anthracis | JRB7 | NR | NR | NR | [135] | |
| S. Typhimurium | NR | NR | 1.5 × 103 CFU/mm2 | NR | [136] | |
| QCM-based assays | S. Typhimurium | Filamentous phage | Chicken wash | 102 | 3 min | [102] |
| M. tuberculosis and M. smegmatis | D29 | NR | 103 | NR | [103] | |
| S. aureus | 12,600 | NR | 104 | NR | [104] | |
| E. coli | T4 | Milk | NR | NR | [16] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Hindi, R.R.; Teklemariam, A.D.; Alharbi, M.G.; Alotibi, I.; Azhari, S.A.; Qadri, I.; Alamri, T.; Harakeh, S.; Applegate, B.M.; Bhunia, A.K. Bacteriophage-Based Biosensors: A Platform for Detection of Foodborne Bacterial Pathogens from Food and Environment. Biosensors 2022, 12, 905. https://doi.org/10.3390/bios12100905
Al-Hindi RR, Teklemariam AD, Alharbi MG, Alotibi I, Azhari SA, Qadri I, Alamri T, Harakeh S, Applegate BM, Bhunia AK. Bacteriophage-Based Biosensors: A Platform for Detection of Foodborne Bacterial Pathogens from Food and Environment. Biosensors. 2022; 12(10):905. https://doi.org/10.3390/bios12100905
Chicago/Turabian StyleAl-Hindi, Rashad R., Addisu D. Teklemariam, Mona G. Alharbi, Ibrahim Alotibi, Sheren A. Azhari, Ishtiaq Qadri, Turki Alamri, Steve Harakeh, Bruce M. Applegate, and Arun K. Bhunia. 2022. "Bacteriophage-Based Biosensors: A Platform for Detection of Foodborne Bacterial Pathogens from Food and Environment" Biosensors 12, no. 10: 905. https://doi.org/10.3390/bios12100905
APA StyleAl-Hindi, R. R., Teklemariam, A. D., Alharbi, M. G., Alotibi, I., Azhari, S. A., Qadri, I., Alamri, T., Harakeh, S., Applegate, B. M., & Bhunia, A. K. (2022). Bacteriophage-Based Biosensors: A Platform for Detection of Foodborne Bacterial Pathogens from Food and Environment. Biosensors, 12(10), 905. https://doi.org/10.3390/bios12100905