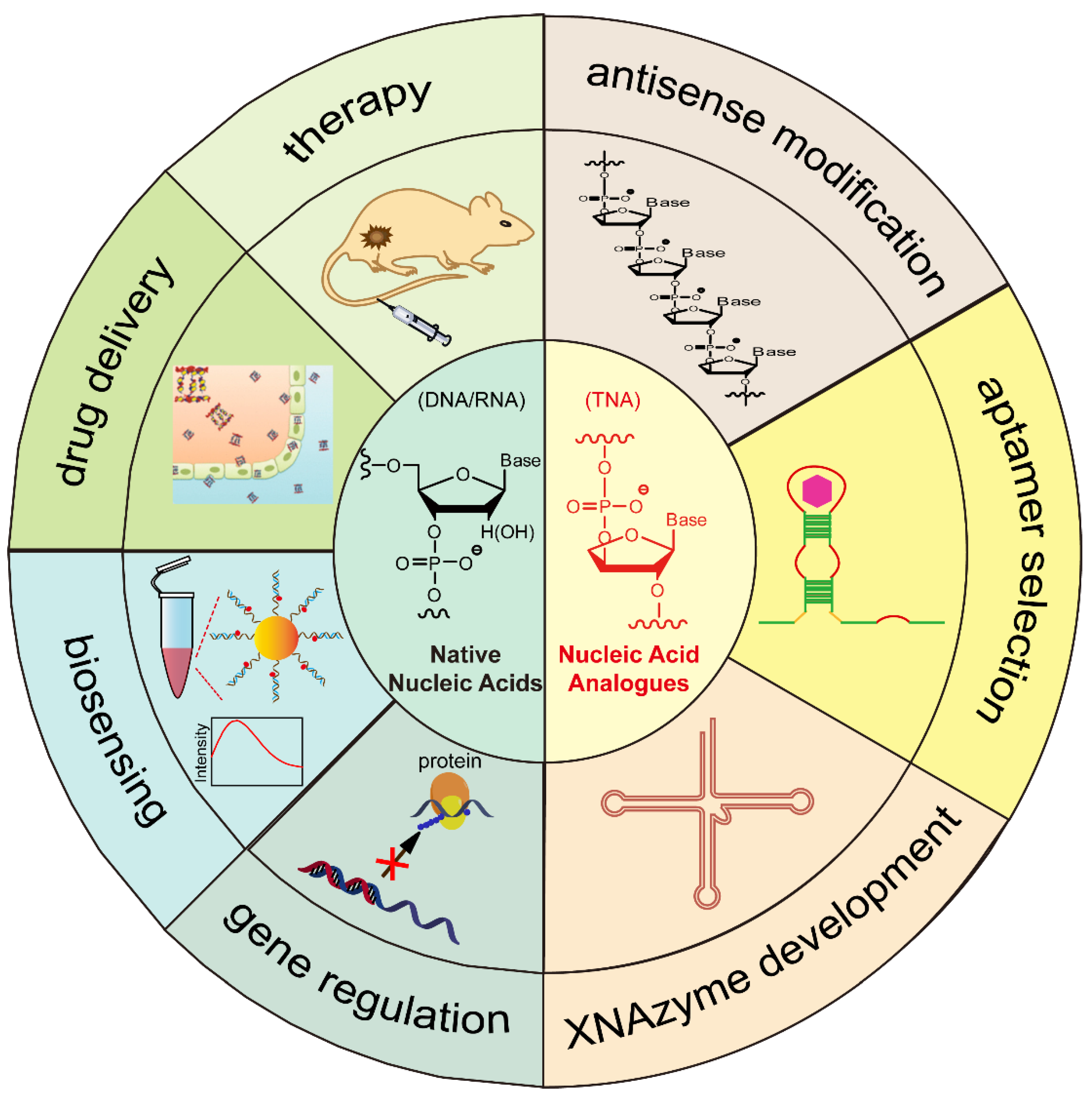
Nucleic Acids and Their Analogues for Biomedical Applications

Abstract
:1. Introduction
2. Native Nucleic Acids
2.1. Molecular Structure
2.2. Synthesis of Nucleic Acids
2.3. Biomedical Applications
2.3.1. Hybridization-Based Applications
2.3.2. Catalysis-Based Applications
2.3.3. Binding-Activity-Based Applications
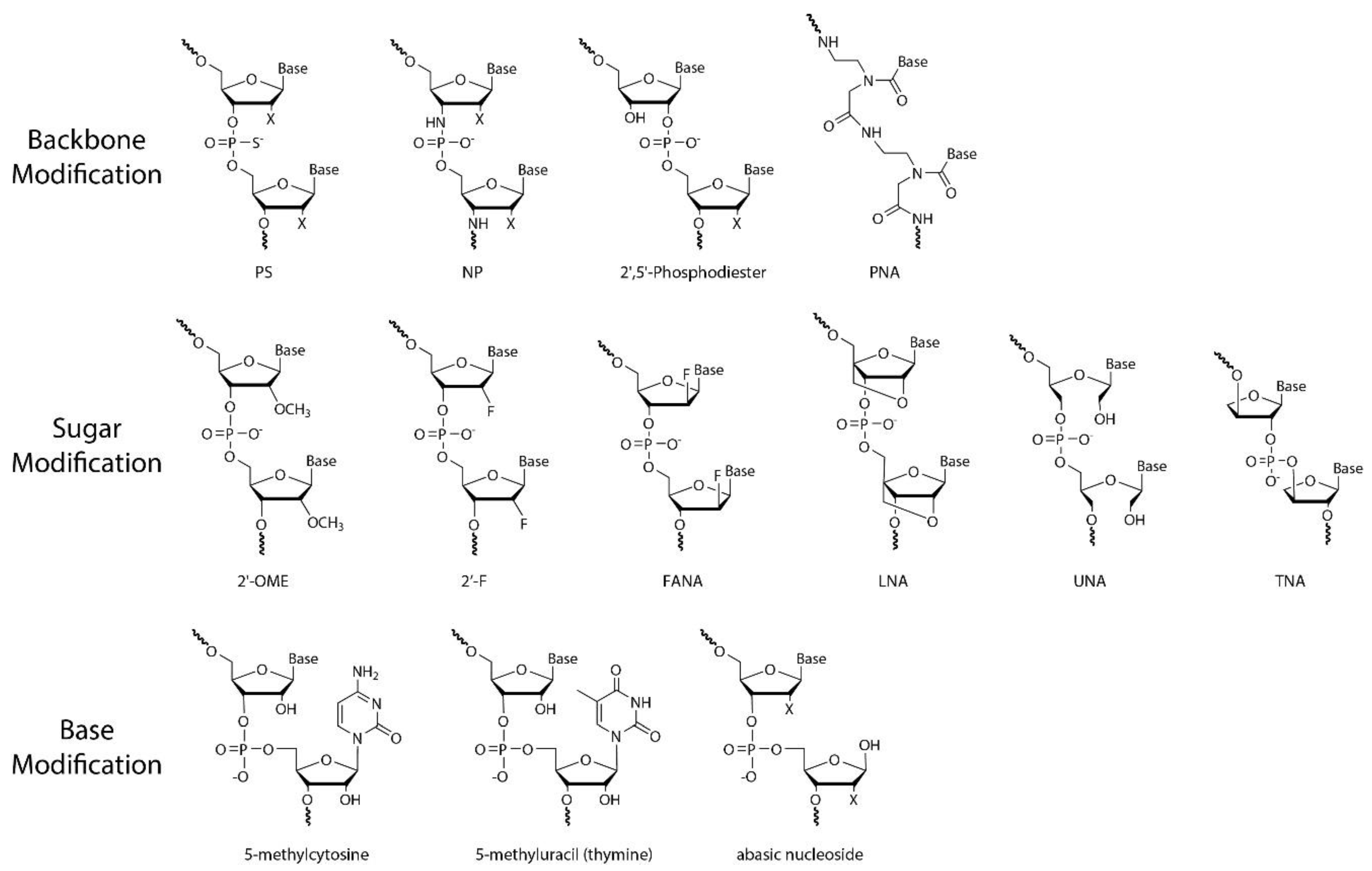
3. Chemically Modified Nucleic Acid Analogues
3.1. Phosphorothioate (PS) ONs
3.1.1. Molecular Structure of PS ONs
3.1.2. Synthesis of PS ONs
3.1.3. Biomedical Applications of PS ONs
3.2. Peptide Nucleic Acid (PNA)
3.2.1. Molecular Structure of PNA
3.2.2. Synthesis of PNA
3.2.3. Biomedical Applications of PNA
3.3. Sugar 2′-O-Methyl (2′-OMe) RNA
3.3.1. Molecular Structure of 2′-OMe RNA
3.3.2. Synthesis of 2′-OMe RNA
3.3.3. Biomedical Applications of 2′-OMe RNA
3.4. Sugar 2′-Deoxy-2′-Fluoro (2′-F) RNA
3.4.1. Molecular Structure of 2′-F RNA
3.4.2. Synthesis of 2′-F RNA
3.4.3. Biomedical Applications of 2′-F RNA
3.5. Locked Nucleic Acids (LNA)
3.5.1. Molecular Structure of LNA
3.5.2. Synthesis of LNA
3.5.3. Biomedical Applications of LNA
3.6. Threose Nucleic Acid (TNA)
3.6.1. Molecular Structure of TNA
3.6.2. Synthesis of TNA
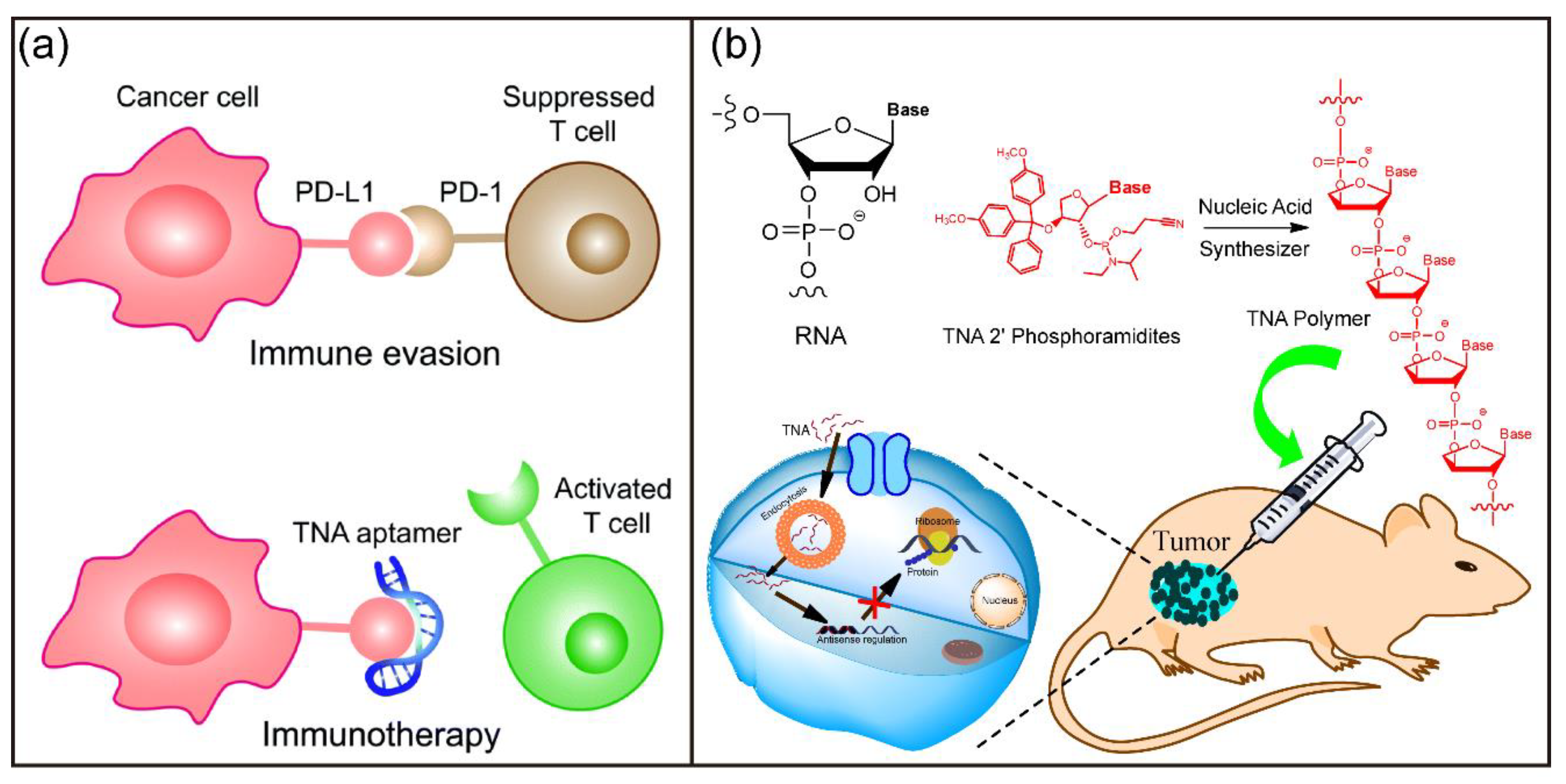
3.6.3. Biomedical Applications of TNA

4. Nucleic Acid Nanotechnology
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, Y.X.; Chen, F.; Li, Q.; Wang, L.H.; Fan, C.H. Isothermal amplification of nucleic acids. Chem. Rev. 2015, 115, 12491–12545. [Google Scholar] [CrossRef]
- Zhao, Y.; Zuo, X.; Li, Q.; Chen, F.; Chen, Y.-R.; Deng, J.; Han, D.; Hao, C.; Huang, F.; Huang, Y. Nucleic acids analysis. Sci. China Chem. 2021, 64, 171–203. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, M.; Sugimoto, N. Molecular evolution of functional nucleic acids with chemical modifications. Molecules 2010, 15, 5423–5444. [Google Scholar] [CrossRef] [PubMed]
- Serganov, A.; Patel, D.J. Ribozymes, riboswitches and beyond: Regulation of gene expression without proteins. Nat. Rev. Genet. 2007, 8, 776–790. [Google Scholar] [CrossRef] [PubMed]
- Wachowius, F.; Hobartner, C. Chemical RNA modifications for studies of RNA structure and dynamics. ChemBioChem 2010, 11, 469–480. [Google Scholar] [CrossRef]
- Chen, C.M.; Yang, Z.J.; Tang, X.J. Chemical modifications of nucleic acid drugs and their delivery systems for gene-based therapy. Med. Res. Rev. 2018, 38, 829–869. [Google Scholar] [CrossRef]
- Weng, Y.H.; Huang, Q.Q.; Li, C.H.; Yang, Y.F.; Wang, X.X.; Yu, J.; Huang, Y.Y.; Liang, X.J. Improved nucleic acid therapy with advanced nanoscale biotechnology. Mol. Ther. Nucleic Acids 2020, 19, 581–601. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, S.; Dunn, M.R.; Chaput, J.C. An efficient and faithful in vitro replication system for threose nucleic acid. J. Am. Chem. Soc. 2013, 135, 3583–3591. [Google Scholar] [CrossRef]
- Wang, F.; Chen, H.; Liu, Z.; Mi, F.; Fang, X.; Liu, J.; Wang, M.; Lo, P.K.; Li, Q. Conjugated polymer dots for biocompatible siRNA delivery. New J. Chem. 2019, 43, 14443–14449. [Google Scholar] [CrossRef]
- Mendes, L.P.; Pan, J.Y.; Torchilin, V.P. Dendrimers as nanocarriers for nucleic acid and drug delivery in cancer therapy. Molecules 2017, 22, 1401. [Google Scholar] [CrossRef] [Green Version]
- Judge, A.; Maclachlan, I. Overcoming the innate immune response to small interfering RNA. Hum. Gene. Ther. 2008, 19, 111–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.T.; He, W.C.; Du, Z.H.; Zhu, L.Y.; Huang, K.L.; Lu, Y.; Luo, Y.B. Functional nucleic acid nanomaterials: Development, properties, and applications. Angew. Chem. Int. Ed. 2021, 60, 6890–6918. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.S.; Wang, F.; Ge, Y.H.; Lo, P.K. Recent developments in aptasensors for diagnostic applications. ACS Appl. Mater. Interfaces 2021, 13, 9329–9358. [Google Scholar] [CrossRef]
- Li, C.H.; Yang, T.R.; Weng, Y.H.; Zhang, M.J.; Zhao, D.Y.; Guo, S.; Hu, B.; Shao, W.X.; Wang, X.X.; Hussain, A.; et al. Ionizable lipid-assisted efficient hepatic delivery of gene editing elements for oncotherapy. Bioact. Mater. 2022, 9, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Morihiro, K.; Kasahara, Y.; Obika, S. Biological applications of xeno nucleic acids. Mol. Biosyst. 2017, 13, 235–245. [Google Scholar] [CrossRef]
- Khvorova, A.; Watts, J.K. The chemical evolution of oligonucleotide therapies of clinical utility. Nat. Biotechnol. 2017, 35, 238–248. [Google Scholar] [CrossRef]
- Seeman, N.C.; Sleiman, H.F. DNA nanotechnology. Nat. Rev. Mater. 2018, 3, 17068. [Google Scholar] [CrossRef]
- Keller, A.; Linko, V. Challenges and perspectives of DNA nanostructures in biomedicine. Angew. Chem. Int. Ed. 2020, 59, 15818–15833. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.D.; Crick, F.H. Molecular structure of nucleic acids: A structure for deoxyribose nucleic acid. Nature 1953, 171, 737–738. [Google Scholar] [CrossRef]
- Yakovchuk, P.; Protozanova, E.; Frank-Kamenetskii, M.D. Base-stacking and base-pairing contributions into thermal stability of the DNA double helix. Nucleic Acids Res. 2006, 34, 1082. [Google Scholar] [CrossRef] [Green Version]
- Marmur, J.; Doty, P. Determination of the base composition of deoxyribonucleic acid from its thermal denaturation temperature. J. Mol. Biol. 1962, 5, 109–118. [Google Scholar] [CrossRef]
- Takahashi, S.; Sugimoto, N. Watson-Crick versus hoogsteen base pairs: Chemical strategy to encode and express genetic information in life. Acc. Chem. Res. 2021, 54, 2110–2120. [Google Scholar] [CrossRef]
- Vandivier, L.E.; Anderson, S.J.; Foley, S.W.; Gregory, B.D. The conservation and function of RNA secondary structure in plants. Annu. Rev. Plant Biol. 2016, 67, 463–488. [Google Scholar] [CrossRef] [Green Version]
- Dickerson, R.E.; Drew, H.R.; Conner, B.N.; Wing, R.M.; Fratini, A.V.; Kopka, M.L. The anatomy of A-DNA, B-DNA, and Z-DNA. Science 1982, 216, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, E.N.; Goh, G.B.; Brooks, C.L.; Al-Hashimi, H.M. Characterizing the protonation state of cytosine in transient G·C hoogsteen base pairs in duplex DNA. J. Am. Chem. Soc. 2013, 135, 6766–6769. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.Q.; Hintze, B.J.; Kimsey, I.J.; Sathyamoorthy, B.; Yang, S.; Richardson, J.S.; Al-Hashimi, H.M. New insights into Hoogsteen base pairs in DNA duplexes from a structure-based survey. Nucleic Acids Res. 2015, 43, 3420–3433. [Google Scholar] [CrossRef]
- Abou Assi, H.; Garavis, M.; Gonzalez, C.; Damha, M.J. I-motif DNA: Structural features and significance to cell biology. Nucleic Acids Res. 2018, 46, 8038–8056. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, J.; Adhikari, S.; Balasubramanian, S. The structure and function of DNA G-quadruplexes. Trends Chem. 2020, 2, 123–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; Caruthers, M. Synthesis of DNA/RNA and their analogs via phosphoramidite and H-phosphonate chemistries. Molecules 2013, 18, 14268–14284. [Google Scholar] [CrossRef] [Green Version]
- Caruthers, M.H. Chemical synthesis of DNA and DNA analogs. Acc. Chem. Res. 1991, 24, 278–284. [Google Scholar] [CrossRef]
- Palluk, S.; Arlow, D.H.; de Rond, T.; Barthel, S.; Kang, J.S.; Bector, R.; Baghdassarian, H.M.; Truong, A.N.; Kim, P.W.; Singh, A.K.; et al. De novo DNA synthesis using polymerase-nucleotide conjugates. Nat. Biotechnol. 2018, 36, 645–650. [Google Scholar] [CrossRef]
- Jensen, M.A.; Davis, R.W. Template-independent enzymatic oligonucleotide synthesis (TiEOS): Its history, prospects, and challenges. Biochemistry 2018, 57, 1821–1832. [Google Scholar] [CrossRef] [PubMed]
- Mackey, J.K.; Gilham, P.T. New approach to synthesis of polyribonucleotides of defined sequence. Nature 1971, 233, 551–553. [Google Scholar] [CrossRef] [PubMed]
- England, T.E.; Uhlenbeck, O.C. Enzymatic oligoribonucleotide synthesis with T4 RNA ligase. Biochemistry 1978, 17, 2069–2076. [Google Scholar] [CrossRef] [PubMed]
- Wochner, A.; Attwater, J.; Coulson, A.; Holliger, P. Ribozyme-catalyzed transcription of an active ribozyme. Science 2011, 332, 209–212. [Google Scholar] [CrossRef]
- Gheibi-Hayat, S.M.; Jamialahmadi, K. Antisense oligonucleotide (AS-ODN) technology: Principle, mechanism and challenges. Biotechnol. Appl. Biochem. 2020, 68, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Chang, H.; Nam, G.; Ko, Y.; Kim, S.H.; Roberts, T.M.; Ryu, J.H. RNAi-based approaches for pancreatic cancer therapy. Pharmaceutics 2021, 13, 1638. [Google Scholar] [CrossRef]
- Wang, H.X.; Li, M.; Lee, C.M.; Chakraborty, S.; Kim, H.W.; Bao, G.; Leong, K.W. CRISPR/Cas9-based genome editing for disease modeling and therapy: Challenges and opportunities for nonviral delivery. Chem. Rev. 2017, 117, 9874–9906. [Google Scholar] [CrossRef]
- Gupta, D.; Bhattacharjee, O.; Mandal, D.; Sen, M.K.; Dey, D.; Dasgupta, A.; Kazi, T.A.; Gupta, R.; Sinharoy, S.; Acharya, K.; et al. CRISPR-Cas9 system: A new-fangled dawn in gene editing. Life Sci. 2019, 232, 116636. [Google Scholar] [CrossRef]
- Cioca, D.P.; Aoki, Y.; Kiyosawa, K. RNA interference is a functional pathway with therapeutic potential in human myeloid leukemia cell lines. Cancer Gene Ther. 2003, 10, 125–133. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Trang, P.; Wiggins, J.F.; Patrawala, L.; Cheng, A.G.; Ford, L.; Weidhaas, J.B.; Brown, D.; Bader, A.G.; Slack, F.J. The let-7 microRNA reduces tumor growth in mouse models of lung cancer. Cell Cycle 2008, 7, 759–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juliano, R.L. The delivery of therapeutic oligonucleotides. Nucleic Acids Res. 2016, 44, 6518–6548. [Google Scholar] [CrossRef] [PubMed]
- Arshad, R.; Barani, M.; Rahdar, A.; Sargazi, S.; Cucchiarini, M.; Pandey, S.; Kang, M. Multi-functionalized nanomaterials and nanoparticles for diagnosis and treatment of retinoblastoma. Biosensors 2021, 11, 97. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Shi, B.; Wang, L.; Liu, Y.; Zou, Y.; Zhou, Y.; Chen, Y.; Zheng, M.; Zhu, Y.; Duan, J.; et al. From mouse to mouse-ear cress: Nanomaterials as vehicles in plant biotechnology. Exploration 2021, 1, 9–20. [Google Scholar] [CrossRef]
- Rosi, N.L.; Giljohann, D.A.; Thaxton, C.S.; Lytton-Jean, A.K.R.; Han, M.S.; Mirkin, C.A. Oligonucleotide-modified gold nanoparticles for intracellular gene regulation. Science 2006, 312, 1027–1030. [Google Scholar] [CrossRef]
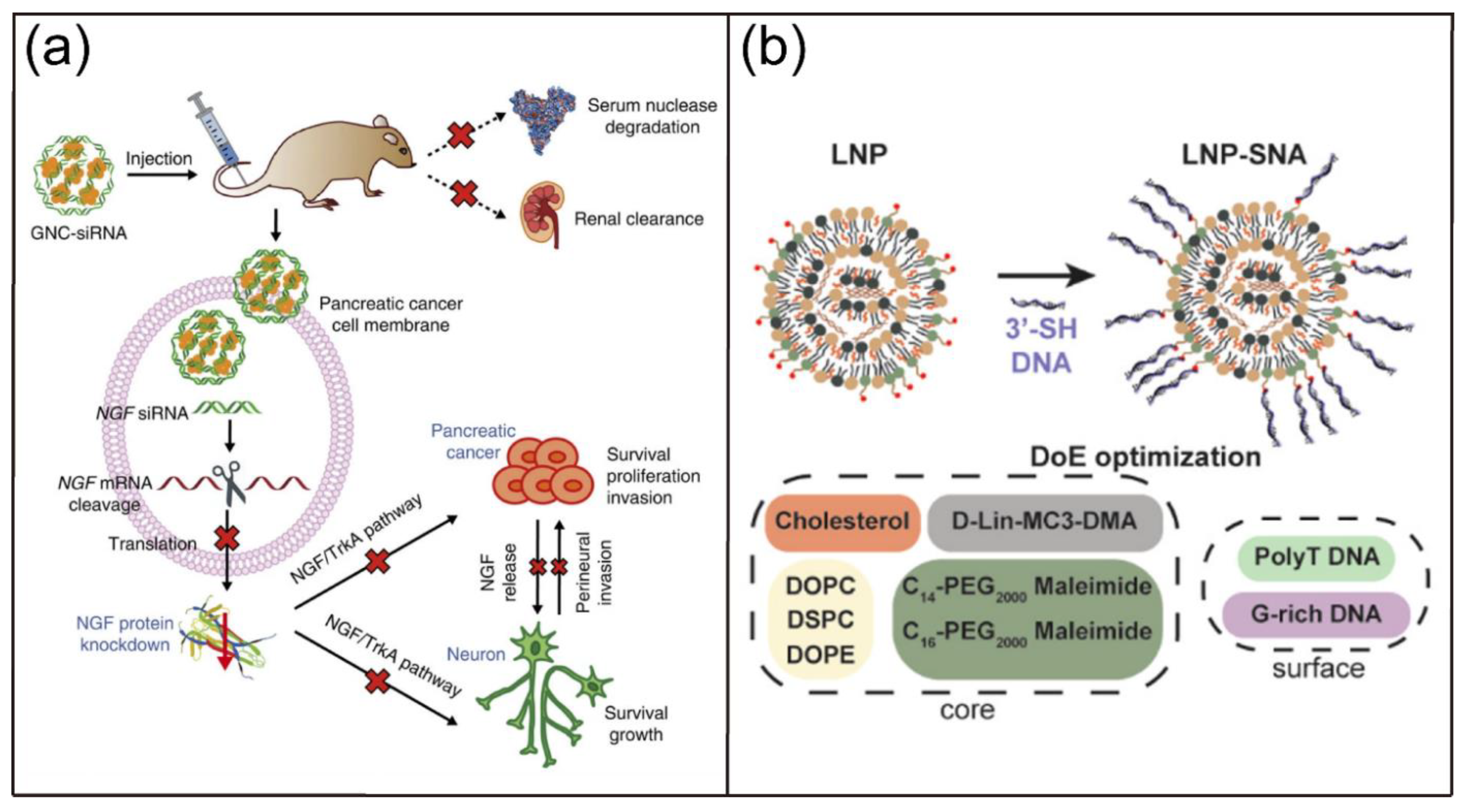
- Lei, Y.F.; Tang, L.X.; Xie, Y.Z.Y.; Xianyu, Y.L.; Zhang, L.M.; Wang, P.; Hamada, Y.; Jiang, K.; Zheng, W.F.; Jiang, X.Y. Gold nanoclusters-assisted delivery of NGF siRNA for effective treatment of pancreatic cancer. Nat. Commun. 2017, 8, 15130. [Google Scholar] [CrossRef]
- Sinegra, A.J.; Evangelopoulos, M.; Park, J.; Huang, Z.Y.; Mirkin, C.A. Lipid nanoparticle spherical nucleic acids for intracellular DNA and RNA delivery. Nano Lett. 2021, 21, 6584–6591. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Xie, Y.; Kilchrist, K.V.; Li, J.; Duvall, C.L.; Oupicky, D. Endosomolytic and tumor-penetrating mesoporous silica nanoparticles for siRNA/miRNA combination cancer therapy. ACS Appl. Mater. Interfaces 2020, 12, 4308–4322. [Google Scholar] [CrossRef]
- El Aamri, M.; Yammouri, G.; Mohammadi, H.; Amine, A.; Korri-Youssoufi, H. Electrochemical biosensors for detection of microRNA as a cancer biomarker: Pros and cons. Biosensors 2020, 10, 186. [Google Scholar] [CrossRef]
- Samanta, D.; Ebrahimi, S.B.; Mirkin, C.A. Nucleic-acid structures as intracellular probes for live cells. Adv. Mater. 2020, 32, 1901743. [Google Scholar] [CrossRef]
- Seferos, D.S.; Giljohann, D.A.; Hill, H.D.; Prigodich, A.E.; Mirkin, C.A. Nano-flares: Probes for transfection and mRNA detection in living cells. J. Am. Chem. Soc. 2007, 129, 15477–15479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.V.; Yang, J.S.; Jo, H.; Kang, B.; Oh, S.S.; Jung, G.Y. Catalytic RNA, ribozyme, and its applications in synthetic biology. Biotechnol. Adv. 2019, 37, 107452. [Google Scholar] [CrossRef] [PubMed]
- Schlosser, K.; Li, Y.F. Biologically inspired synthetic enzymes made from DNA. Chem. Biol. 2009, 16, 311–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Zhao, H.M.; Chen, S.; Yu, H.T.; Zhang, Y.B.; Quan, X. A “turn-on” fluorescent copper biosensor based on DNA cleavage-dependent graphene-quenched DNAzyme. Biosens. Bioelectron. 2011, 26, 4111–4116. [Google Scholar] [CrossRef]
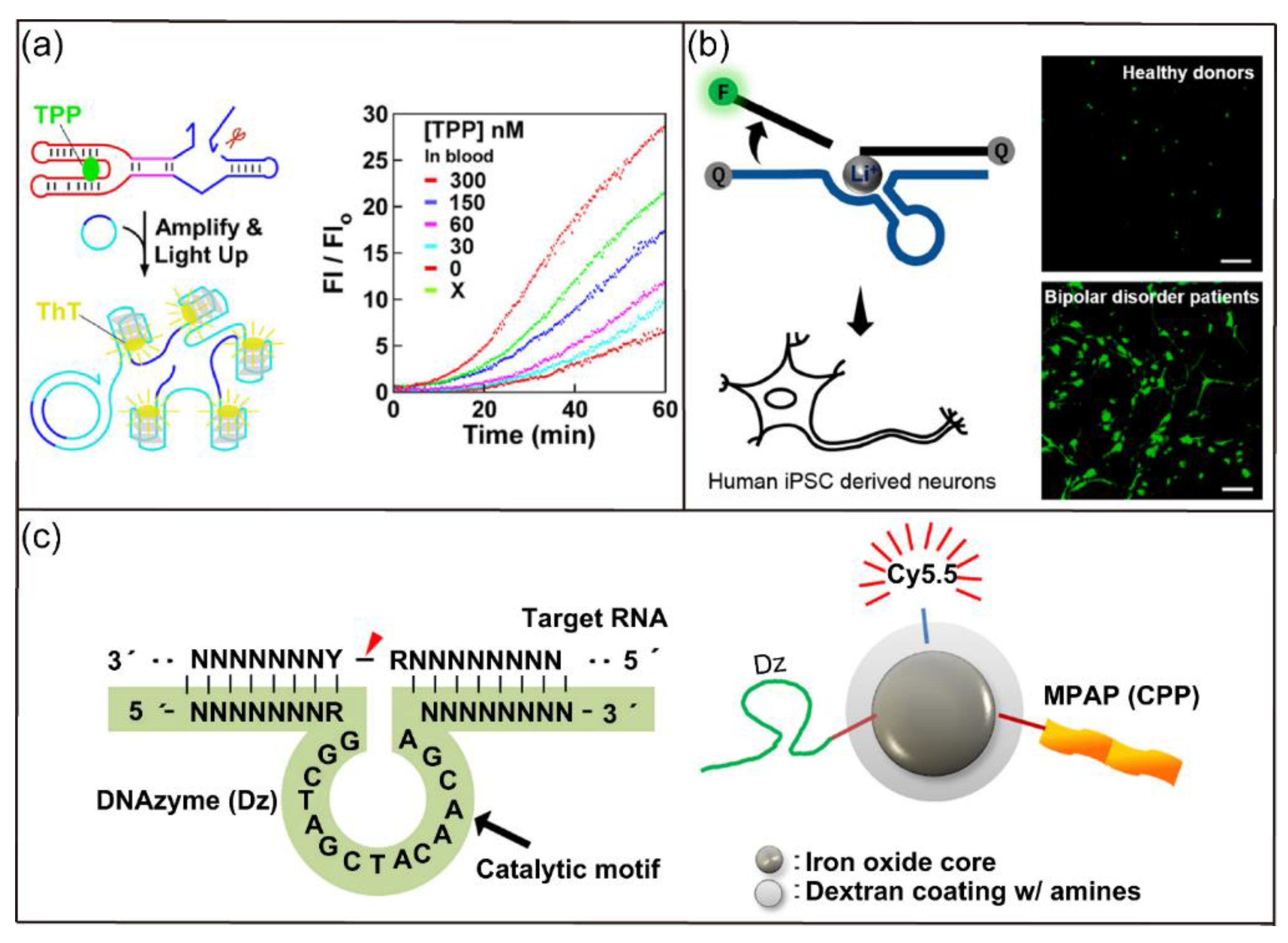
- Du, X.Y.; Cheng, X.Q.; Li, W.; Ge, Z.L.; Zhong, C.J.; Fan, C.H.; Gu, H.Z. Engineering allosteric ribozymes to detect thiamine pyrophosphate in whole blood. Anal. Chem. 2021, 93, 4277–4284. [Google Scholar] [CrossRef] [PubMed]
- McGhee, C.E.; Yang, Z.; Guo, W.; Wu, Y.; Lyu, M.; DeLong, C.J.; Hong, S.; Ma, Y.; McInnis, M.G.; O′Shea, K.S. DNAzyme-based lithium-selective imaging reveals higher lithium accumulation in bipolar disorder patient-derived neurons. Acs Cent. Sci. 2021, 7, 1809–1820. [Google Scholar] [CrossRef]
- Zhou, W.H.; Ding, J.S.; Liu, J.W. Theranostic DNAzymes. Theranostics 2017, 7, 1010–1025. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, S.R.; Jang, H.; Kim, K.S.; Lee, B.; Kim, K.B.; Kim, Y.K.; Yeo, W.S.; Lee, Y.; Kim, D.E.; Min, D.H. Functional delivery of DNAzyme with iron oxide nanoparticles for hepatitis C virus gene knockdown. Biomaterials 2012, 33, 2754–2761. [Google Scholar] [CrossRef]
- Rouge, J.L.; Sita, T.L.; Hao, L.L.; Kouri, F.M.; Briley, W.E.; Stegh, A.H.; Mirkin, C.A. Ribozyme-spherical nucleic acids. J. Am. Chem. Soc. 2015, 137, 10528–10531. [Google Scholar] [CrossRef] [Green Version]
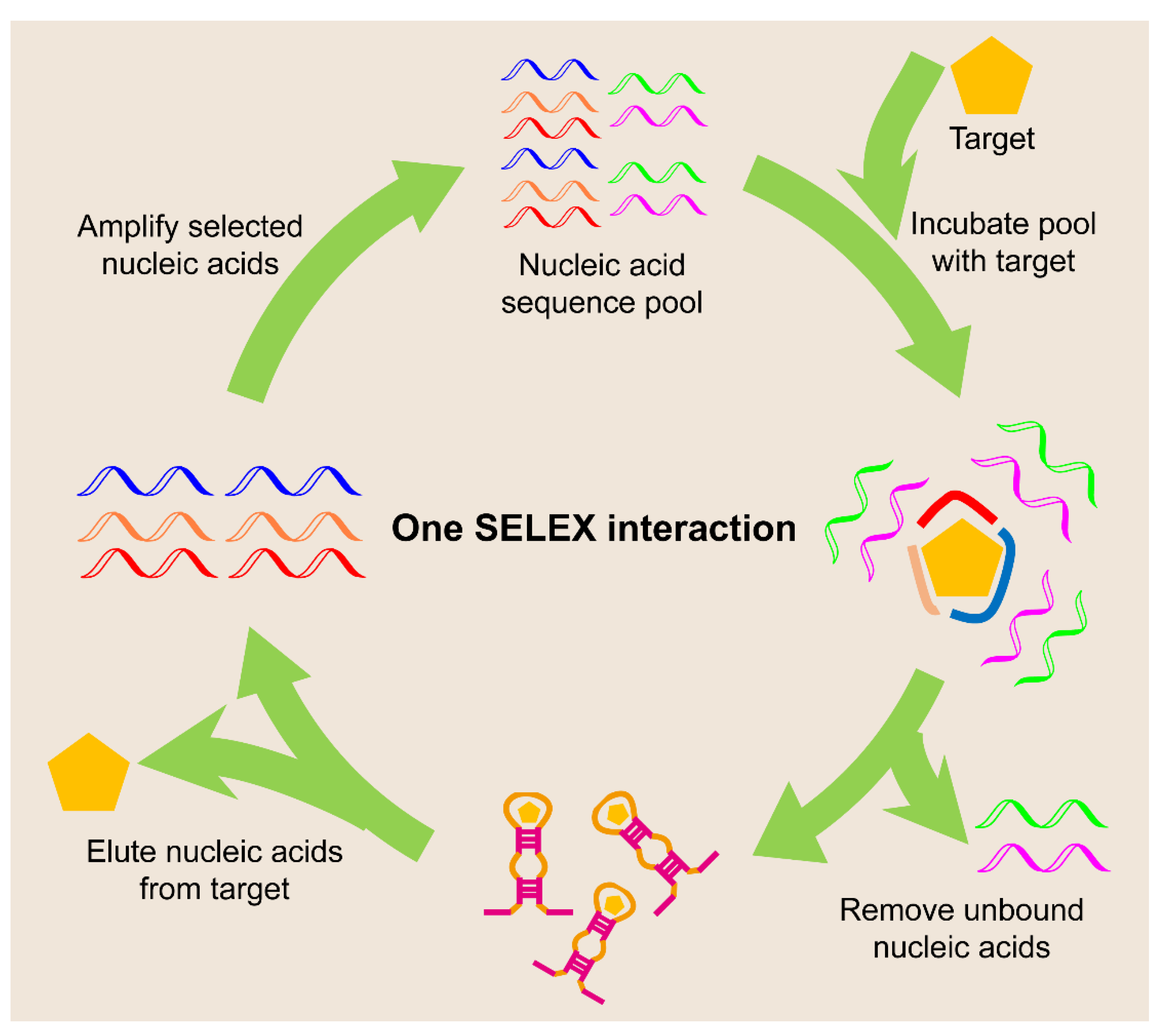
- Li, L.; Xu, S.J.; Yan, H.; Li, X.W.; Yazd, H.S.; Li, X.; Huang, T.; Cui, C.; Jiang, J.H.; Tan, W.H. Nucleic acid aptamers for molecular diagnostics and therapeutics: Advances and perspectives. Angew. Chem. Int. Ed. 2021, 60, 2221–2231. [Google Scholar] [CrossRef]
- Dunn, M.R.; Jimenez, R.M.; Chaput, J.C. Analysis of aptamer discovery and technology. Nat. Rev. Chem. 2017, 1, 0076. [Google Scholar] [CrossRef]
- Hianik, T. Advances in electrochemical and acoustic aptamer-based biosensors and immunosensors in diagnostics of leukemia. Biosensors 2021, 11, 177. [Google Scholar] [CrossRef] [PubMed]
- Pinyou, P.; Noguer, T.; Blay, V. Enzymes, aptamers, and their use in sensors. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Zuo, X.L.; Song, S.P.; Zhang, J.; Pan, D.; Wang, L.H.; Fan, C.H. A target-responsive electrochemical aptamer switch (TREAS) for reagentless detection of nanomolar ATP. J. Am. Chem. Soc. 2007, 129, 1042–1043. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.F.; Dai, Z.W.; Tang, X.; Lin, Z.H.; Lo, P.K.; Meyyappan, M.; Lai, K.W.C. Graphene field-effect transistors for the sensitive and selective detection of Escherichia coli using pyrene-tagged DNA aptamer. Adv. Healthc. Mater. 2017, 6, 1700736. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Gao, Q.; Cheung, M.C.; Leung, H.M.; Lau, T.C.K.; Sleiman, H.F.; Lai, K.W.C.; Lo, P.K. A highly versatile platform based on geometrically well-defined 3D DNA nanostructures for selective recognition and positioning of multiplex targets. Nanoscale 2016, 8, 18291–18295. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, G.; Rengan, A.K. Aptamer-mediated nanotheranostics for cancer treatment: A review. ACS Appl. Nano Mater. 2020, 3, 9542–9559. [Google Scholar] [CrossRef]
- Ding, C.P.; Zhang, C.L.; Yin, X.Y.; Cao, X.Y.; Cai, M.F.; Xian, Y.Z. Near-infrared fluorescent Ag2S nanodot-based signal amplification for efficient detection of circulating tumor cells. Anal. Chem. 2018, 90, 6702–6709. [Google Scholar] [CrossRef]
- Li, S.P.; Jiang, Q.; Liu, S.L.; Zhang, Y.L.; Tian, Y.H.; Song, C.; Wang, J.; Zou, Y.G.; Anderson, G.J.; Han, J.Y.; et al. A DNA nanorobot functions as a cancer therapeutic in response to a molecular trigger in vivo. Nat. Biotechnol. 2018, 36, 258–264. [Google Scholar] [CrossRef]
- Li, X.W.; Yang, Y.; Zhao, H.Z.; Zhu, T.; Yang, Z.H.; Xu, H.Y.; Fu, Y.Q.; Lin, F.; Pan, X.S.; Li, L.; et al. Enhanced in vivo blood-brain barrier penetration by circular tau-transferrin receptor bifunctional aptamer for tauopathy therapy. J. Am. Chem. Soc. 2020, 142, 3862–3872. [Google Scholar] [CrossRef]
- McKenzie, L.K.; El-Khoury, R.; Thorpe, J.D.; Damha, M.J.; Hollenstein, M. Recent progress in non-native nucleic acid modifications. Chem. Soc. Rev. 2021, 50, 5126–5164. [Google Scholar] [CrossRef]
- Liu, C.X.; Zou, G.R.; Peng, S.; Wang, Y.F.; Yang, W.; Wu, F.; Jiang, Z.R.; Zhang, X.; Zhou, X. 5-formyluracil as a multifunctional building block in biosensor designs. Angew. Chem. Int. Ed. 2018, 57, 9689–9693. [Google Scholar] [CrossRef] [PubMed]
- Chaput, J.C.; Herdewijn, P. What is XNA? Angew. Chem. Int. Ed. 2019, 58, 11570–11572. [Google Scholar] [CrossRef] [PubMed]
- Saran, R.; Huang, Z.C.; Liu, J.W. Phosphorothioate nucleic acids for probing metal binding, biosensing and nanotechnology. Coord. Chem. Rev. 2021, 428, 213624. [Google Scholar] [CrossRef]
- Crooke, S.T.; Wang, S.Y.; Vickers, T.A.; Shen, W.; Liang, X.H. Cellular uptake and trafficking of antisense oligonucleotides. Nat. Biotechnol. 2017, 35, 230–237. [Google Scholar] [CrossRef]
- Eckstein, F. Phosphorothioates, essential components of therapeutic oligonucleotides. Nucleic Acid Ther. 2014, 24, 374–387. [Google Scholar] [CrossRef] [PubMed]
- Volk, D.E.; Lokesh, G.L.R. Development of phosphorothioate DNA and DNA thioaptamers. Biomedicines 2017, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Braasch, D.A.; Jensen, S.; Liu, Y.H.; Kaur, K.; Arar, K.; White, M.A.; Corey, D.R. RNA interference in mammalian cells by chemically-modified RNA. Biochemistry 2003, 42, 7967–7975. [Google Scholar] [CrossRef]
- Leung, H.M.; Chan, M.S.; Liu, L.S.; Wong, S.W.; Lo, T.W.; Lau, C.H.; Tin, C.; Lo, P.K. Dual-function, cationic, peptide-coated nanodiamond systems: Facilitating nuclear-targeting delivery for enhanced gene therapy applications. ACS Sustain. Chem. Eng. 2018, 6, 9671–9681. [Google Scholar] [CrossRef]
- King, D.J.; Ventura, D.A.; Brasier, A.R.; Gorenstein, D.G. Novel combinatorial selection of phosphorothioate oligonucleotide aptamers. Biochemistry 1998, 37, 16489–16493. [Google Scholar] [CrossRef]
- Abeydeera, N.D.; Egli, M.; Cox, N.; Mercier, K.; Conde, J.N.; Pallan, P.S.; Mizurini, D.M.; Sierant, M.; Hibti, F.E.; Hassell, T.; et al. Evoking picomolar binding in RNA by a single phosphorodithioate linkage. Nucleic Acids Res. 2016, 44, 8052–8064. [Google Scholar] [CrossRef]
- Nielsen, P.E.; Egholm, M.; Berg, R.H.; Buchardt, O. Sequence-selective recognition of DNA by strand displacement with a thymine-substituted polyamide. Science 1991, 254, 1497–1500. [Google Scholar] [CrossRef]
- Nielsen, P.E. Peptide nucleic acids (PNA) in chemical biology and drug discovery. Chem. Biodivers. 2010, 7, 786–804. [Google Scholar] [CrossRef]
- Singh, K.R.B.; Sridevi, P.; Singh, R.P. Potential applications of peptide nucleic acid in biomedical domain. Eng. Rep. 2020, 2, e12238. [Google Scholar] [CrossRef]
- Ray, A.; Norden, B. Peptide nucleic acid (PNA): Its medical and biotechnical applications and promise for the future. FASEB J. 2000, 14, 1041–1060. [Google Scholar] [CrossRef]
- Saarbach, J.; Sabale, P.M.; Winssinger, N. Peptide nucleic acid (PNA) and its applications in chemical biology, diagnostics, and therapeutics. Curr. Opin. Chem. Biol. 2019, 52, 112–124. [Google Scholar] [CrossRef]
- Gupta, A.; Mishra, A.; Puri, N. Peptide nucleic acids: Advanced tools for biomedical applications. J. Biotechnol. 2017, 259, 148–159. [Google Scholar] [CrossRef]
- Shakeel, S.; Karim, S.; Ali, A. Peptide nucleic acid (PNA)—A review. J. Chem. Technol. Biotechnol. 2006, 81, 892–899. [Google Scholar] [CrossRef]
- Lundin, K.E.; Good, L.; Stromberg, R.; Graslund, A.; Smith, C.I.E. Biological activity and biotechnological aspects of peptide nucleic acid. Adv. Genet. 2006, 56, 1–51. [Google Scholar]
- Dragulescu-Andrasi, A.; Rapireddy, S.; He, G.F.; Bhattacharya, B.; Hyldig-Nielsen, J.J.; Zon, G.; Ly, D.H. Cell-permeable peptide nucleic acid designed to bind to the 5′-untranslated region of E-cadherin transcript induces potent and sequence-specific antisense effects. J. Am. Chem. Soc. 2006, 128, 16104–16112. [Google Scholar] [CrossRef]
- Canady, T.D.; Telmer, C.A.; Oyaghire, S.N.; Armitage, B.A.; Bruchez, M.P. In vitro reversible translation control using gamma PNA probes. J. Am. Chem. Soc. 2015, 137, 10268–10275. [Google Scholar] [CrossRef]
- Babar, I.A.; Cheng, C.J.; Booth, C.J.; Liang, X.P.; Weidhaas, J.B.; Saltzman, W.M.; Slack, F.J. Nanoparticle-based therapy in an in vivo microRNA-155 (miR-155)-dependent mouse model of lymphoma. Proc. Natl. Acad. Sci. USA 2012, 109, E1695–E1704. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.J.; Bahal, R.; Babar, I.A.; Pincus, Z.; Barrera, F.; Liu, C.; Svoronos, A.; Braddock, D.T.; Glazer, P.M.; Engelman, D.M.; et al. MicroRNA silencing for cancer therapy targeted to the tumour microenvironment. Nature 2015, 518, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.F.; Wu, B.L.; Wei, T.; Egholm, M.; Strauss, W.M. Unique chromosome identification and sequence-specific structural analysis with short PNA oligomers. Mamm. Genome 2000, 11, 384–391. [Google Scholar] [CrossRef]
- Machado, A.; Almeida, C.; Carvalho, A.; Boyen, F.; Haesebrouck, F.; Rodrigues, L.; Cerca, N.; Azevedo, N.F. Fluorescence in situ hybridization method using a peptide nucleic acid probe for identification of Lactobacillus spp. in milk samples. Int. J. Food Microbiol. 2013, 162, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Palecek, E.; Nielsen, P.E.; Rivas, G.; Cai, X.H.; Shiraishi, H.; Dontha, N.; Luo, D.B.; Farias, P.A.M. Peptide nucleic acid probes for sequence-specific DNA biosensors. J. Am. Chem. Soc. 1996, 118, 7667–7670. [Google Scholar] [CrossRef]
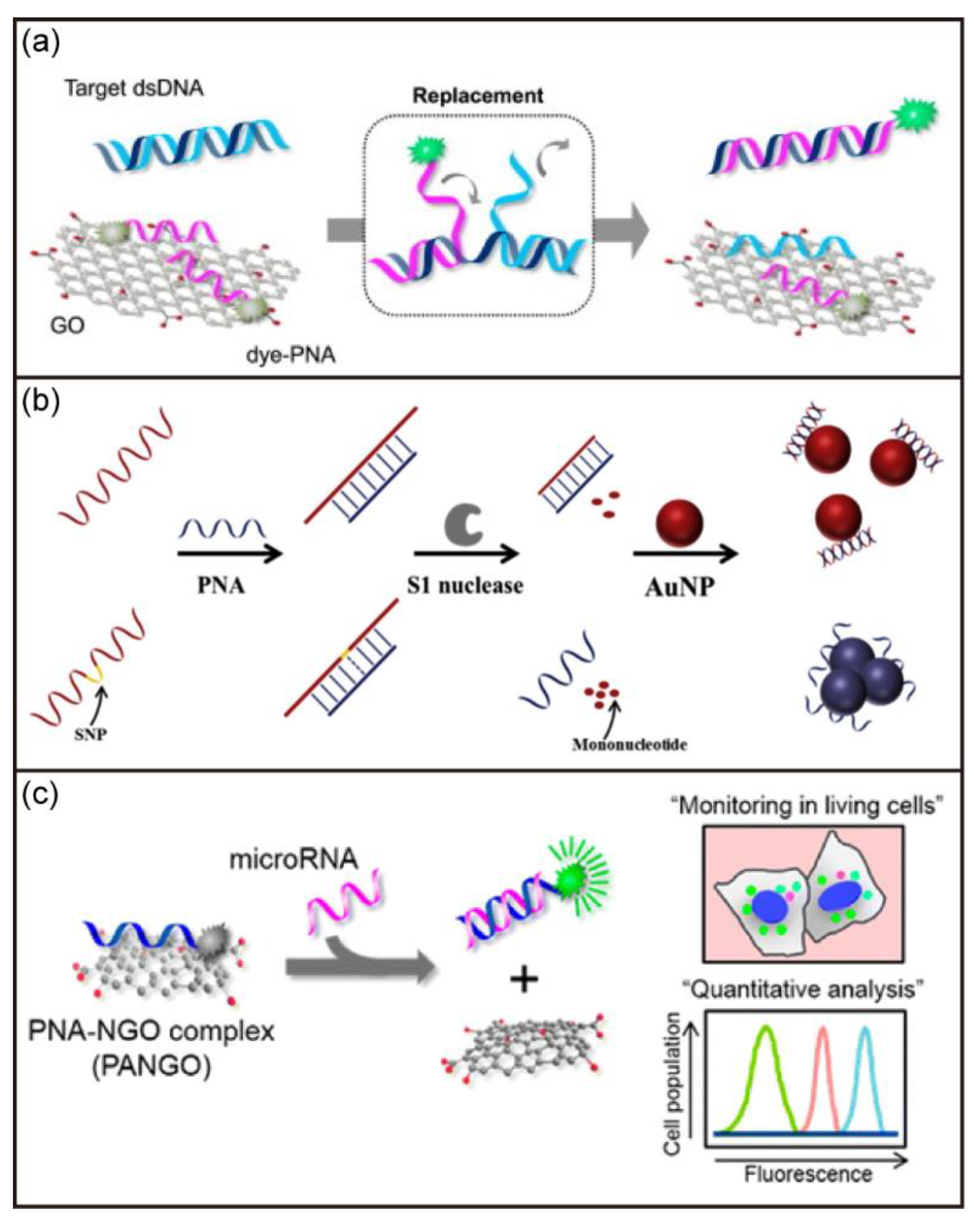
- Lee, J.; Park, I.S.; Jung, E.; Lee, Y.; Min, D.H. Direct, sequence-specific detection of dsDNA based on peptide nucleic acid and graphene oxide without requiring denaturation. Biosens. Bioelectron. 2014, 62, 140–144. [Google Scholar] [CrossRef]
- Xing, S.; Xu, X.J.; Fu, P.; Xu, M.J.; Gao, T.T.; Zhang, X.K.; Zhao, C. Colorimetric detection of single base-pair mismatches based on the interactions of PNA and PNA/DNA complexes with unmodified gold nanoparticles. Colloids Surf. B 2019, 181, 333–340. [Google Scholar] [CrossRef]
- Ryoo, S.R.; Lee, J.; Yeo, J.; Na, H.K.; Kim, Y.K.; Jang, H.; Lee, J.H.; Han, S.W.; Lee, Y.; Kim, V.N.; et al. Quantitative and multiplexed microRNA sensing in living cells based on peptide nucleic acid and nano gaphene oxide (PANGO). ACS Nano 2013, 7, 5882–5891. [Google Scholar] [CrossRef]
- Prakash, T.P. An overview of sugar-modified oligonucleotides for antisense therapeutics. Chem. Biodivers. 2011, 8, 1616–1641. [Google Scholar] [CrossRef] [PubMed]
- Nishizaki, T.; Iwai, S.; Ohtsuka, E.; Nakamura, H. Solution structure of an RNA·2′-O-methylated RNA hybrid duplex containing an RNA·DNA hybrid segment at the center. Biochemistry 1997, 36, 2577–2585. [Google Scholar] [CrossRef]
- Burmeister, P.E.; Lewis, S.D.; Silva, R.F.; Preiss, J.R.; Horwitz, L.R.; Pendergrast, P.S.; McCauley, T.G.; Kurz, J.C.; Epstein, D.M.; Wilson, C.; et al. Direct in vitro selection of a 2′-O-methyl aptamer to VEGF. Chem. Biol. 2005, 12, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Deleavey, G.F.; Damha, M.J. Designing chemically modified oligonucleotides for targeted gene silencing. Chem. Biol. 2012, 19, 937–954. [Google Scholar] [CrossRef] [Green Version]
- Inoue, H.; Hayase, Y.; Imura, A.; Iwai, S.; Miura, K.; Ohtsuka, E. Synthesis and hybridization studies on two complementary nona (2′-O-methyl) ribonucleotides. Nucleic Acids Res. 1987, 15, 6131–6148. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.; Oberhauser, B.; Holzner, A.; Brunar, H.; Issakides, G.; Schaffner, G.; Cotten, M.; Knollmuller, M.; Noe, C.R. A simple procedure for the preparation of protected 2′-O-methyl or 2′-O-ethyl ribonucleoside-3′-O-phsophoramidites. Nucleic Acids Res. 1991, 19, 5965–5971. [Google Scholar] [CrossRef] [Green Version]
- Kurreck, J. Antisense technologies-Improvement through novel chemical modifications. Eur. J. Biochem. 2003, 270, 1628–1644. [Google Scholar] [CrossRef] [PubMed]
- Fa, M.; Radeghieri, A.; Henry, A.A.; Romesberg, F.E. Expanding the substrate repertoire of a DNA polymerase by directed evolution. J. Am. Chem. Soc. 2004, 126, 1748–1754. [Google Scholar] [CrossRef]
- Chen, T.J.; Hongdilokkul, N.; Liu, Z.X.; Adhikary, R.; Tsuen, S.S.; Romesberg, F.E. Evolution of thermophilic DNA polymerases for the recognition and amplification of C2′-modified DNA. Nat. Chem. 2016, 8, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Kestemont, D.; Renders, M.; Leonczak, P.; Abramov, M.; Schepers, G.; Pinheiro, V.B.; Rozenski, J.; Herdewijn, P. XNA ligation using T4 DNA ligase in crowding conditions. Chem. Commun. 2018, 54, 6408–6411. [Google Scholar] [CrossRef] [Green Version]
- Gragoudas, E.S.; Adamis, A.P.; Cunningham, E.T.; Feinsod, M.; Guyer, D.R.; Neova, V.I.S.O. Pegaptanib for neovascular age-related macular degeneration. N. Engl. J. Med. 2004, 351, 2805–2816. [Google Scholar] [CrossRef] [Green Version]
- Jackson, A.L.; Burchard, J.; Leake, D.; Reynolds, A.; Schelter, J.; Guo, J.; Johnson, J.M.; Lim, L.; Karpilow, J.; Nichols, K.; et al. Position-specific chemical modification of siRNAs reduces “off-target” transcript silencing. RNA 2006, 12, 1197–1205. [Google Scholar] [CrossRef] [Green Version]
- Kraynack, B.A.; Baker, B.F. Small interfering RNAs containing full 2′-O-methylribonucleotide-modified sense strands display argonaute2/eIF2C2-dependent activity. RNA 2006, 12, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Green, L.S.; Jellinek, D.; Bell, C.; Beebe, L.A.; Feistner, B.D.; Gill, S.C.; Jucker, F.M.; Janjic, N. Nuclease-resistant nucleic-acid ligands to vascular-permeability factor vascular endothelial growth-factor. Chem. Biol. 1995, 2, 683–695. [Google Scholar] [CrossRef] [Green Version]
- Rigo, F.; Hua, Y.M.; Chun, S.J.; Prakash, T.P.; Krainer, A.R.; Bennett, C.F. Synthetic oligonucleotides recruit ILF2/3 to RNA transcripts to modulate splicing. Nat. Chem. Biol. 2012, 8, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, A.M.; Casper, M.D.; Freier, S.M.; Lesnik, E.A.; Zounes, M.C.; Cummins, L.L.; Gonzalez, C.; Cook, P.D. Uniformly modified 2′-deoxy-2′-fluoro phosphorothioate oligonucleotides as nuclease-resistant antisense compounds with high-affinity and specificity for RNA targets. J. Med. Chem. 1993, 36, 831–841. [Google Scholar] [CrossRef]
- Sinha, N.D.; Biernat, J.; Mcmanus, J.; Koster, H. Polymer support oligonucleotide synthesis XVIII: Use of beta-cyanoethyl-N,N-dialkylamino-/N-morpholino phosphoramidite of deoxynucleosides for the synthesis of DNA fragments simplifying deprotection and isolation of the final product. Nucleic Acids Res. 1984, 12, 4539–4557. [Google Scholar] [CrossRef]
- Pieken, W.A.; Olsen, D.B.; Benseler, F.; Aurup, H.; Eckstein, F. Kinetic characterization of ribonuclease-resistant 2′-modified hammerhead ribozymes. Science 1991, 253, 314–317. [Google Scholar] [CrossRef]
- Sousa, R.; Padilla, R. Mutant T7 RNA-polymerase as a DNA-Polymerase. Embo J. 1995, 14, 4609–4621. [Google Scholar] [CrossRef]
- Ono, T.; Scalf, M.; Smith, L.M. 2′-fluoro modified nucleic acids: Polymerase-directed synthesis, properties and stability to analysis by matrix-assisted laser desorption/ionization mass spectrometry. Nucleic Acids Res. 1997, 25, 4581–4588. [Google Scholar] [CrossRef] [Green Version]
- Cozens, C.; Pinheiro, V.B.; Vaisman, A.; Woodgate, R.; Holliger, P. A short adaptive path from DNA to RNA polymerases. Proc. Natl. Acad. Sci. USA 2012, 109, 8067–8072. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Hernandez, A.; Tan, M.; Wollenhaupt, J.; Tabor, S.; Richardson, C.C. Synthesis of 2′-Fluoro RNA by Syn5 RNA polymerase. Nucleic Acids Res. 2015, 43, e94. [Google Scholar] [CrossRef] [Green Version]
- Manoharan, M.; Akinc, A.; Pandey, R.K.; Qin, J.; Hadwiger, P.; John, M.; Mills, K.; Charisse, K.; Maier, M.A.; Nechev, L.; et al. Unique gene-silencing and structural properties of 2′-fluoro-modified siRNAs. Angew. Chem. Int. Ed. 2011, 50, 2284–2288. [Google Scholar] [CrossRef] [Green Version]
- Pagratis, N.C.; Bell, C.; Chang, Y.F.; Jennings, S.; Fitzwater, T.; Jellinek, D.; Dang, C. Potent 2′-amino-, and 2′-fluoro-2′-deoxyribonucleotide RNA inhibitors of keratinocyte growth factor. Nat. Biotechnol. 1997, 15, 68–73. [Google Scholar] [CrossRef]
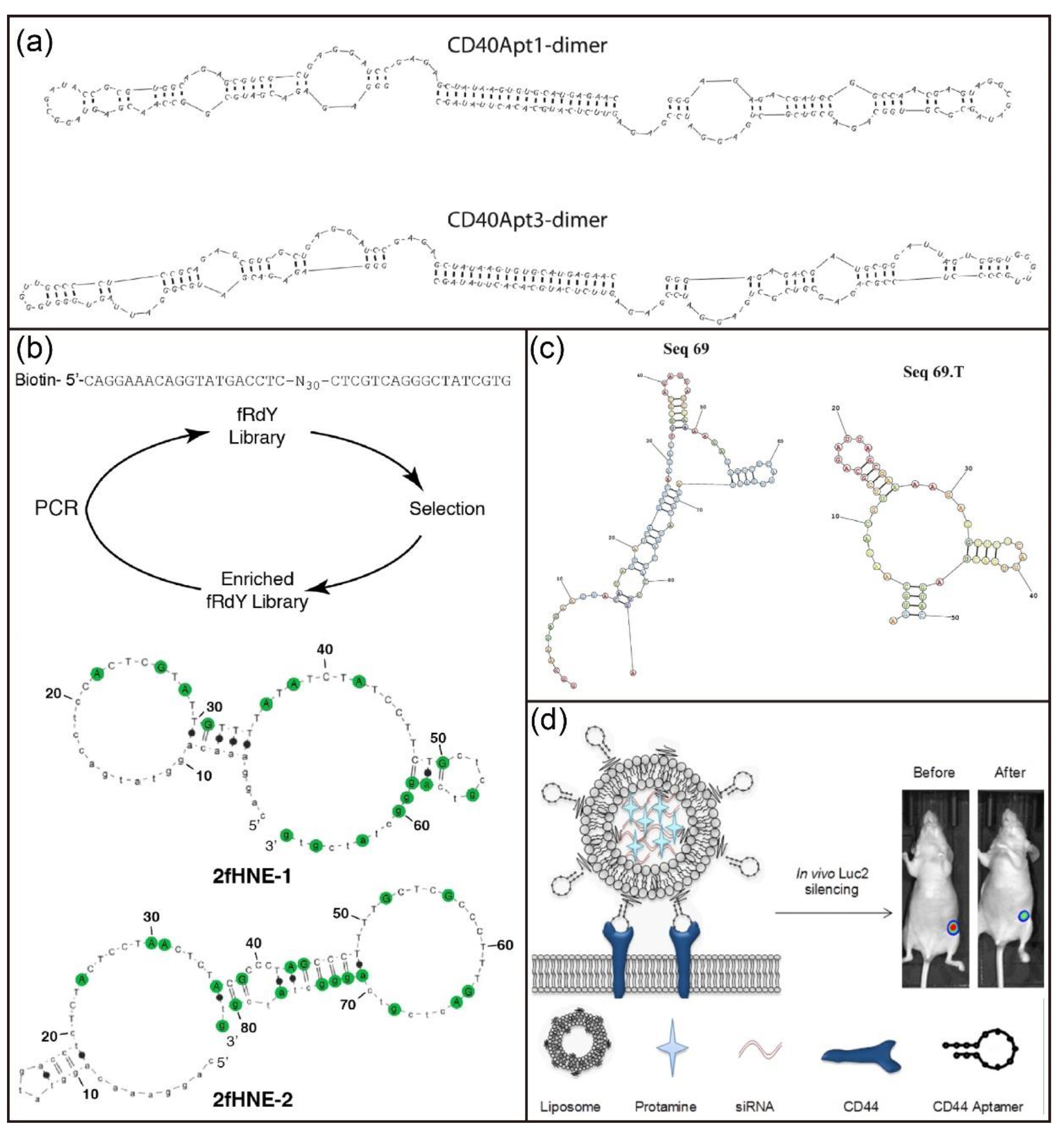
- Soldevilla, M.M.; Villanueva, H.; Bendandi, M.; Inoges, S.; de Cerio, A.L.D.; Pastor, F. 2-fluoro-RNA oligonucleotide CD40 targeted aptamers for the control of B lymphoma and bone-marrow aplasia. Biomaterials 2015, 67, 274–285. [Google Scholar] [CrossRef]
- Thirunavukarasu, D.; Chen, T.J.; Liu, Z.X.; Hongdilokkul, N.; Romesberg, F.E. Selection of 2′-fluoro-modified aptamers with optimized properties. J. Am. Chem. Soc. 2017, 139, 2892–2895. [Google Scholar] [CrossRef]
- Catuogno, S.; Di Martino, M.T.; Nuzzo, S.; Esposito, C.L.; Tassone, P.; de Franciscis, V. An anti-BCMA RNA aptamer for miRNA intracellular delivery. Mol. Ther. Nucleic Acids 2019, 18, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Ababneh, N.; Alshaer, W.; Allozi, O.; Mahafzah, A.; El-Khateeb, M.; Hillaireau, H.; Noiray, M.; Fattal, E.; Ismail, S. In vitro selection of modified RNA aptamers against CD44 cancer stem cell marker. Nucleic Acid Ther. 2013, 23, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Alshaer, W.; Hillaireau, H.; Vergnaud, J.; Ismail, S.; Fattal, E. Functionalizing liposomes with anti-CD44 aptamer for selective targeting of cancer cells. Bioconjugate Chem. 2015, 26, 1307–1313. [Google Scholar] [CrossRef]
- Alshaer, W.; Hillaireau, H.; Vergnaud, J.; Mura, S.; Delomenie, C.; Sauvage, F.; Ismail, S.; Fattal, E. Aptamer-guided siRNA-loaded nanomedicines for systemic gene silencing in CD-44 expressing murine triple-negative breast cancer model. J. Control. Release 2018, 271, 98–106. [Google Scholar] [CrossRef]
- Obika, S.; Nanbu, D.; Hari, Y.; Andoh, J.; Morio, K.; Doi, T.; Imanishi, T. Stability and structural features of the duplexes containing nucleoside analogues with a fixed N-type conformation, 2′-O,4′-C-methyleneribonucleosides. Tetrahedron Lett. 1998, 39, 5401–5404. [Google Scholar] [CrossRef]
- Koshkin, A.A.; Singh, S.K.; Nielsen, P.; Rajwanshi, V.K.; Kumar, R.; Meldgaard, M.; Olsen, C.E.; Wengel, J. LNA (Locked Nucleic Acids): Synthesis of the adenine, cytosine, guanine, 5-methylcytosine, thymine and uracil bicyclonucleoside monomers, oligomerisation, and unprecedented nucleic acid recognition. Tetrahedron 1998, 54, 3607–3630. [Google Scholar] [CrossRef]
- Kaur, H.; Babu, B.R.; Maiti, S. Perspectives on chemistry and therapeutic applications of Locked Nucleic Acid (LNA). Chem. Rev. 2007, 107, 4672–4697. [Google Scholar] [CrossRef]
- Hagedorn, P.H.; Persson, R.; Funder, E.D.; Albaek, N.; Diemer, S.L.; Hansen, D.J.; Moller, M.R.; Papargyri, N.; Christiansen, H.; Hansen, B.R.; et al. Locked nucleic acid: Modality, diversity, and drug discovery. Drug Discov. Today 2018, 23, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Campbell, M.A.; Wengel, J. Locked vs. unlocked nucleic acids (LNA vs. UNA): Contrasting structures work towards common therapeutic goals. Chem. Soc. Rev. 2011, 40, 5680–5689. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.Z.; Chattopadhyaya, J. The synthesis of therapeutic locked nucleos(t)ides. Curr. Opin. Drug Discov. 2009, 12, 876–898. [Google Scholar]
- Sharma, V.K.; Rungta, P.; Maikhuri, V.K.; Prasad, A.K. An astute synthesis of locked nucleic acid monomers. Sustain. Chem. Process. 2015, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Wahlestedt, C.; Salmi, P.; Good, L.; Kela, J.; Johnsson, T.; Hokfelt, T.; Broberger, C.; Porreca, F.; Lai, J.; Ren, K.K.; et al. Potent and nontoxic antisense oligonucleotides containing locked nucleic acids. Proc. Natl. Acad. Sci. USA 2000, 97, 5633–5638. [Google Scholar] [CrossRef] [Green Version]
- Arzumanov, A.; Walsh, A.P.; Rajwanshi, V.K.; Kumar, R.; Wengel, J.; Gait, M.J. Inhibition of HIV-1 Tat-dependent trans activation by steric block chimeric 2′-O-methyl/LNA oligoribonucleotides. Biochemistry 2001, 40, 14645–14654. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, V.M.; Scaria, V.; Maiti, S. Antagomirzymes: Oligonucleotide enzymes that specifically silence microRNA function. Angew. Chem. Int. Ed. 2009, 48, 2557–2560. [Google Scholar] [CrossRef]
- Mook, O.R.; Baas, F.; de Wissel, M.B.; Fluiter, K. Evaluation of locked nucleic acid-modified small interfering RNA in vitro and in vivo. Mol. Cancer Ther. 2007, 6, 833–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
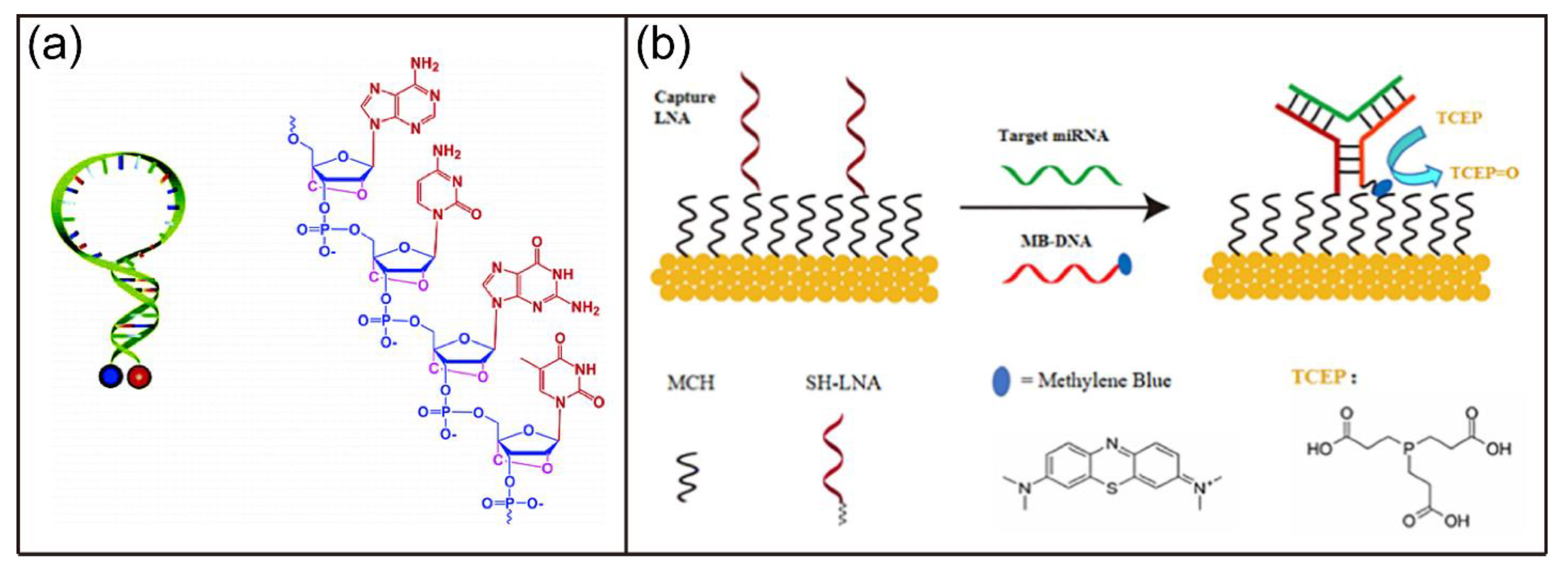
- Wang, L.; Yang, C.Y.J.; Medley, C.D.; Benner, S.A.; Tan, W.H. Locked nucleic acid molecular beacons. J. Am. Chem. Soc. 2005, 127, 15664–15665. [Google Scholar] [CrossRef]
- Hu, Z.H.; Zhao, B.Y.; Miao, P.; Hou, X.L.; Xing, F.F.; Chen, Y.Y.; Feng, L.Y. Three-way junction DNA based electrochemical biosensor for microRNAs detection with distinguishable locked nucleic acid recognition and redox cycling signal amplification. J. Electroanal. Chem. 2021, 880, 114861. [Google Scholar] [CrossRef]
- Tao, Y.Z.; Wang, W.J.; Fu, C.L.; Luo, F.; Guo, L.H.; Qiu, B.; Lin, Z.Y. Sensitive biosensor for p53 DNA sequence based on the photothermal effect of gold nanoparticles and the signal amplification of locked nucleic acid functionalized DNA walkers using a thermometer as readout. Talanta 2020, 220, 121398. [Google Scholar] [CrossRef] [PubMed]
- Darfeuille, F.; Hansen, J.B.; Orum, H.; Di Primo, C.; Toulme, J.J. LNA/DNA chimeric oligomers mimic RNA aptamers targeted to the TAR RNA element of HIV-1. Nucleic Acids Res. 2004, 32, 3512. [Google Scholar] [CrossRef] [Green Version]
- Darfeuille, F.; Reigadas, S.; Hansen, J.B.; Orum, H.; Di Primo, C.; Toulme, J.J. Aptamers targeted to an RNA hairpin show improved specificity compared to that of complementary oligonucleotides. Biochemistry 2006, 45, 12076–12082. [Google Scholar] [CrossRef]
- Virno, A.; Randazzo, A.; Giancola, C.; Bucci, M.; Cirinoc, G.; Mayol, L. A novel thrombin binding aptamer containing a G-LNA residue. Bioorg. Med. Chem. 2007, 15, 5710–5718. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, F.J.; Kalra, N.; Wengel, J.; Vester, B. Aptamers as a model for functional evaluation of LNA and 2′-amino LNA. Bioorg. Med. Chem. Lett. 2009, 19, 6585–6587. [Google Scholar] [CrossRef]
- Schoning, K.U.; Scholz, P.; Guntha, S.; Wu, X.; Krishnamurthy, R.; Eschenmoser, A. Chemical etiology of nucleic acid structure: The alpha-threofuranosyl-(3′-> 2′) oligonucleotide system. Science 2000, 290, 1347–1351. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Y.; Zhang, S.; Chaput, J.C. Darwinian evolution of an alternative genetic system provides support for TNA as an RNA progenitor. Nat. Chem. 2012, 4, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Pallan, P.S.; Wilds, C.J.; Wawrzak, Z.; Krishnamurthy, R.; Eschenmoser, A.; Egli, M. Why does TNA cross-pair more strongly with RNA than with DNA? An answer from X-ray analysis. Angew. Chem. Int. Ed. 2003, 42, 5893–5895. [Google Scholar] [CrossRef]
- Ebert, M.O.; Mang, C.; Krishnamurthy, R.; Eschenmoser, A.; Jaun, B. The structure of a TNA-TNA complex in solution: NMR study of the octamer duplex derived from alpha-(L)-threofuranosyl-(3′-2′)-CGAATTCG. J. Am. Chem. Soc. 2008, 130, 15105–15115. [Google Scholar] [CrossRef]
- Anosova, I.; Kowal, E.A.; Sisco, N.J.; Sau, S.; Liao, J.Y.; Bala, S.; Rozners, E.; Egli, M.; Chaput, J.C.; Van Horn, W.D. Structural insights into conformation differences between DNA/TNA and RNA/TNA chimeric duplexes. Chembiochem 2016, 17, 1705–1708. [Google Scholar] [CrossRef] [PubMed]
- Lackey, H.H.; Peterson, E.M.; Chen, Z.; Harris, J.M.; Heemstra, J.M. Thermostability trends of TNA:DNA duplexes reveal strong purine dependence. ACS Synth. Biol. 2019, 8, 1144–1152. [Google Scholar] [CrossRef]
- Lackey, H.H.; Chen, Z.; Harris, J.M.; Peterson, E.M.; Heemstra, J.M. Single-molecule kinetics show DNA pyrimidine content strongly affects RNA:DNA and TNA:DNA heteroduplex dissociation rates. ACS Synth. Biol. 2020, 9, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zhang, J.Z. Thermodynamic insights of base flipping in TNA duplex: Force fields, salt concentrations, and free-energy simulation methods. CCS Chem. 2021, 3, 1026–1039. [Google Scholar] [CrossRef]
- Muniz, M.I.; Lackey, H.H.; Heemstra, J.M.; Weber, G. DNA/TNA mesoscopic modeling of melting temperatures suggests weaker hydrogen bonding of CG than in DNA/RNA. Chem. Phys. Lett. 2020, 749, 137413. [Google Scholar] [CrossRef]
- Wu, X.L.; Delgado, G.; Krishnamurthy, R.; Eschenmoser, A. 2,6-diaminopurine in TNA: Effect on duplex stabilities and on the efficiency of template-controlled ligations. Org. Lett. 2002, 4, 1283–1286. [Google Scholar] [CrossRef]
- Wu, X.L.; Guntha, S.; Ferencic, M.; Krishnamurthy, R.; Eschenmoser, A. Base-pairing systems related to TNA: Alpha-threofuranosyl oligonucleotides containing phosphoramidate linkages. Org. Lett. 2002, 4, 1279–1282. [Google Scholar] [CrossRef]
- Liao, J.Y.; Anosova, I.; Bala, S.; Van Horn, W.D.; Chaput, J.C. A parallel stranded G-quadruplex composed of threose nucleic acid (TNA). Biopolymers 2017, 107, e22999. [Google Scholar] [CrossRef] [PubMed]
- Sau, S.P.; Fahmi, N.E.; Liao, J.Y.; Bala, S.; Chaput, J.C. A scalable synthesis of alpha-L-threose nucleic acid monomers. J. Org. Chem. 2016, 81, 2302–2307. [Google Scholar] [CrossRef] [PubMed]
- Sau, S.P.; Chaput, J.C. A one-pot synthesis of alpha-L-threofuranosyl nucleoside triphosphates (tNTPs). Bioorg. Med. Chem. Lett. 2016, 26, 3271–3273. [Google Scholar] [CrossRef] [Green Version]
- Mei, H.; Shi, C.H.; Jimenez, R.M.; Wang, Y.J.; Kardouh, M.; Chaput, J.C. Synthesis and polymerase activity of a fluorescent cytidine TNA triphosphate analogue. Nucleic Acids Res. 2017, 45, 5629–5638. [Google Scholar] [CrossRef] [Green Version]
- Bala, S.; Liao, J.Y.; Mei, H.; Chaput, J.C. Synthesis of alpha-L-threofuranosyl nucleoside 3′-monophosphates, 3′-phosphoro(2-methyl)imidazolides, and 3′-triphosphates. J. Org. Chem. 2017, 82, 5910–5916. [Google Scholar] [CrossRef]
- Sau, S.P.; Chaput, J.C. A gram-scale HPLC-free synthesis of TNA triphosphates using an iterative phosphorylation strategy. Org. Lett. 2017, 19, 4379–4382. [Google Scholar] [CrossRef]
- Bala, S.; Liao, J.Y.; Zhang, L.; Tran, C.N.; Chim, N.; Chaput, J.C. Synthesis of 2′-deoxy-alpha-L-threofuranosyl nucleoside triphosphates. J. Org. Chem. 2018, 83, 8840–8850. [Google Scholar] [CrossRef] [PubMed]
- Mei, H.; Wang, Y.; Yik, E.J.; Chaput, J.C. Synthesis and polymerase recognition of a pyrrolocytidine TNA triphosphate. Biopolymers 2021, 112, e23388. [Google Scholar] [CrossRef]
- Li, Q.F.; Maola, V.A.; Chim, N.; Hussain, J.; Lozoya-Colinas, A.; Chaput, J.C. Synthesis and polymerase recognition of threose nucleic acid triphosphates equipped with diverse chemical functionalities. J. Am. Chem. Soc. 2021, 143, 17761–17768. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.S.; Leung, H.M.; Tam, D.Y.; Lo, T.W.; Wong, S.W.; Lo, P.K. alpha-L-threose nucleic acids as biocompatible antisense oligonucleotides for suppressing gene expression in living cells. ACS Appl. Mater. Interfaces 2018, 10, 9736–9743. [Google Scholar] [CrossRef]
- Chaput, J.C.; Ichida, J.K.; Szostak, J.W. DNA polymerase-mediated DNA synthesis on a TNA template. J. Am. Chem. Soc. 2003, 125, 856–857. [Google Scholar] [CrossRef] [Green Version]
- Chaput, J.C.; Szostak, J.W. TNA synthesis by DNA polymerases. J. Am. Chem. Soc. 2003, 125, 9274–9275. [Google Scholar] [CrossRef] [PubMed]
- Kempeneers, V.; Vastmans, K.; Rozenski, J.; Herdewijn, P. Recognition of threosyl nucleotides by DNA and RNA polymerases. Nucleic Acids Res. 2003, 31, 6221–6226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichida, J.K.; Zou, K.; Horhota, A.; Yu, B.; McLaughlin, L.W.; Szostak, J.W. An in vitro selection system for TNA. J. Am. Chem. Soc. 2005, 127, 2802–2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horhota, A.; Zou, K.Y.; Ichida, J.K.; Yu, B.; McLaughlin, L.W.; Szostak, J.W.; Chaput, J.C. Kinetic analysis of an efficient DNA-dependent TNA polymerase. J. Am. Chem. Soc. 2005, 127, 7427–7434. [Google Scholar] [CrossRef] [Green Version]
- Dunn, M.R.; Otto, C.; Fenton, K.E.; Chaput, J.C. Improving polymerase activity with unnatural substrates by sampling mutations in homologous protein architectures. ACS Chem. Biol. 2016, 11, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Chim, N.; Shi, C.H.; Sau, S.P.; Nikoomanzar, A.; Chaput, J.C. Structural basis for TNA synthesis by an engineered TNA polymerase. Nat. Commun. 2017, 8, 1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCloskey, C.M.; Liao, J.Y.; Bala, S.; Chaput, J.C. Ligase-mediated threose nucleic acid synthesis on DNA templates. ACS Synth. Biol. 2019, 8, 282–286. [Google Scholar] [CrossRef]
- Culbertson, M.C.; Temburnikar, K.W.; Sau, S.P.; Liao, J.Y.; Bala, S.; Chaput, J.C. Evaluating TNA stability under simulated physiological conditions. Bioorg. Med. Chem. Lett. 2016, 26, 2418–2421. [Google Scholar] [CrossRef] [Green Version]
- Mei, H.; Liao, J.Y.; Jimenez, R.M.; Wang, Y.J.; Bala, S.; McCloskey, C.; Switzer, C.; Chaput, J.C. Synthesis and evolution of a threose nucleic acid aptamer bearing 7-deaza-7-substituted guanosine residues. J. Am. Chem. Soc. 2018, 140, 5706–5713. [Google Scholar] [CrossRef]
- Rangel, A.E.; Chen, Z.; Ayele, T.M.; Heemstra, J.M. In vitro selection of an XNA aptamer capable of small-molecule recognition. Nucleic Acids Res. 2018, 46, 8057–8068. [Google Scholar] [CrossRef]
- Zhang, L.; Chaput, J.C. In vitro selection of an ATP-binding TNA aptamer. Molecules 2020, 25, 4194. [Google Scholar] [CrossRef]
- Li, X.T.; Li, Z.; Yu, H.Y. Selection of threose nucleic acid aptamers to block PD-1/PD-L1 interaction for cancer immunotherapy. Chem. Commun. 2020, 56, 14653–14656. [Google Scholar] [CrossRef]
- Dunn, M.R.; McCloskey, C.M.; Buckley, P.; Rhea, K.; Chaput, J.C. Generating biologically stable TNA aptamers that function with high affinity and thermal stability. J. Am. Chem. Soc. 2020, 142, 7721–7724. [Google Scholar] [CrossRef]
- McCloskey, C.M.; Li, Q.; Yik, E.J.; Chim, N.; Ngor, A.K.; Medina, E.; Grubisic, I.; Co Ting Keh, L.; Poplin, R.; Chaput, J.C. Evolution of functionally enhanced α-l-threofuranosyl nucleic acid aptamers. ACS Synth. Biol. 2021, 10, 3190–3199. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, L.S.; Lau, C.H.; Han Chang, T.J.; Tam, D.Y.; Leung, H.M.; Tin, C.; Lo, P.K. Synthetic α-l-threose nucleic acids targeting BcL-2 show gene silencing and in vivo antitumor activity for cancer therapy. ACS Appl. Mater. Interfaces 2019, 11, 38510–38518. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.J.; Burke, D.H.; Chaput, J.C. Activation of innate immune responses by a CpG oligonucleotide sequence composed entirely of threose nucleic acid. Nucleic Acid Ther. 2019, 29, 51–59. [Google Scholar] [CrossRef]
- Lu, X.H.; Wu, X.H.; Wu, T.T.; Han, L.; Liu, J.B.; Ding, B.Q. Efficient construction of a stable linear gene based on a TNA loop modified primer pair for gene delivery. Chem. Commun. 2020, 56, 9894–9897. [Google Scholar] [CrossRef]
- Wang, Y.J.; Nguyen, K.; Spitale, R.C.; Chaput, J.C. A biologically stable DNAzyme that efficiently silences gene expression in cells. Nat. Chem. 2021, 13, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.; Wang, Y.J.; England, W.E.; Chaput, J.C.; Spitale, R.C. Allele-specific RNA knockdown with a biologically stable and catalytically efficient XNAzyme. J. Am. Chem. Soc. 2021, 143, 4519–4523. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.F.; Chaput, J.C. REVEALR: A multicomponent XNAzyme-based nucleic acid detection system for SARS-CoV-2. J. Am. Chem. Soc. 2021, 143, 8957–8961. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.Y.; Song, D.F.; Sun, X.; Zhang, Z.; Li, X.T.; Li, Z.; Yu, H.Y. A threose nucleic acid enzyme with RNA ligase activity. J. Am. Chem. Soc. 2021, 143, 8154–8163. [Google Scholar] [CrossRef]
- Wang, X.; Chandrasekaran, A.R.; Shen, Z.Y.; Ohayon, Y.P.; Wang, T.; Kizer, M.E.; Sha, R.J.; Mao, C.D.; Yan, H.; Zhang, X.P.; et al. Paranemic crossover DNA: There and back again. Chem. Rev. 2019, 119, 6273–6289. [Google Scholar] [CrossRef]
- Xiao, L.; Jun, Y.W.; Kool, E.T. DNA tiling enables precise acylation-based labeling and control of mRNA. Angew. Chem. Int. Ed. 2021, 60, 26798–26805. [Google Scholar] [CrossRef]
- Chan, M.S.; Tam, D.Y.; Dai, Z.W.; Liu, L.S.; Ho, J.W.T.; Chan, M.L.; Xu, D.; Wong, M.S.; Tin, C.; Lo, P.K. Mitochondrial delivery of therapeutic agents by amphiphilic DNA nanocarriers. Small 2016, 12, 770–781. [Google Scholar] [CrossRef] [PubMed]
- Tam, D.Y.; Ho, J.W.T.; Chan, M.S.; Lau, C.H.; Chang, T.J.H.; Leung, H.M.; Liu, L.S.; Wang, F.; Chan, L.L.H.; Tin, C.; et al. Penetrating the blood-brain barrier by self-assembled 3D DNA nanocages as drug delivery vehicles for brain cancer therapy. ACS Appl. Mater. Interfaces 2020, 12, 28928–28940. [Google Scholar] [CrossRef]
- Liu, J.B.; Song, L.L.; Liu, S.L.; Jiang, Q.; Liu, Q.; Li, N.; Wang, Z.G.; Ding, B.Q. A DNA-based nanocarrier for efficient gene delivery and combined cancer therapy. Nano Lett. 2018, 18, 3328–3334. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Jiang, Q.; Beziere, N.; Song, L.L.; Zhang, Q.; Peng, D.; Chi, C.W.; Yang, X.; Guo, H.B.; Diot, G.; et al. DNA-nanostructure-gold-nanorod hybrids for enhanced in vivo optoacoustic imaging and photothermal therapy. Adv. Mater. 2016, 28, 10000–10007. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.S.; Nie, C.P.; Hu, Y.L.; Yi, J.T.; Liu, C.; Zhang, J.; He, M.M.; He, M.Y.; Chen, T.T.; Chu, X. Aptamer-functionalized DNA origami for targeted codelivery of antisense oligonucleotides and doxorubicin to enhance therapy in drug-resistant cancer cells. ACS Appl. Mater. Interfaces 2020, 12, 400–409. [Google Scholar] [CrossRef]
- Dai, Z.W.; Leung, H.M.; Lo, P.K. Stimuli-responsive self-assembled DNA nanomaterials for biomedical applications. Small 2017, 13, 1602881. [Google Scholar] [CrossRef] [PubMed]
- Tam, D.Y.; Zhuang, X.Y.; Wong, S.W.; Lo, P.K. Photoresponsive self-assembled DNA nanomaterials: Design, working principles, and applications. Small 2019, 15, 1805481. [Google Scholar] [CrossRef]
- Yu, Y.; Jin, B.; Li, Y.L.; Deng, Z.X. Stimuli-responsive DNA self-assembly: From principles to applications. Chem. Eur. J. 2019, 25, 9785–9798. [Google Scholar] [CrossRef]
- Tam, D.Y.; Leung, H.M.; Chan, M.S.; Lo, P.K. G-quadruplex-mediated molecular switching of self-assembled 3D DNA nanocages. Chemnanomat 2017, 3, 750–754. [Google Scholar] [CrossRef]
- Chan, M.S.; Leung, H.M.; Wong, S.W.; Lin, Z.H.; Gao, Q.; Chang, T.J.H.; Lai, K.W.C.; Lo, P.K. Reversible reconfiguration of high-order DNA nanostructures by employing G-quartet toeholds as adhesive units. Nanoscale 2020, 12, 2464–2471. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.J.; Jiang, T.Y.; Lu, Y.; Reiff, M.; Mo, R.; Gu, Z. Cocoon-like self-degradable DNA nanoclew for anticancer drug delivery. J. Am. Chem. Soc. 2014, 136, 14722–14725. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.L.; Zhao, Y.M.; Xu, X.M.; Xu, K.L.; Zhang, M.Z.; Huang, K.; Kang, H.Z.; Lin, H.C.; Yang, Y.; Han, D. An intelligent DNA nanorobot for autonomous anticoagulation. Angew. Chem. Int. Ed. 2020, 59, 17697–17704. [Google Scholar] [CrossRef] [PubMed]
- Tam, D.Y.; Dai, Z.W.; Chan, M.S.; Liu, L.S.; Cheung, M.C.; Bolze, F.; Tin, C.; Lo, P.K. A reversible DNA logic gate platform operated by one-and two-photon excitations. Angew. Chem. Int. Ed. 2016, 55, 164–168. [Google Scholar] [CrossRef]
- Dai, Z.W.; Lo, P.K. Photo-switchable patterning of gold nanoparticles along 3D DNA nanotubes. Nanoscale 2018, 10, 5431–5435. [Google Scholar] [CrossRef]
- Deng, X.W.; Cao, M.J.; Zhang, J.K.; Hu, K.L.; Yin, Z.X.; Zhou, Z.X.; Xiao, X.Q.; Yang, Y.S.; Sheng, W.; Wu, Y.; et al. Hyaluronic acid-chitosan nanoparticles for co-delivery of MiR-34a and doxorubicin in therapy against triple negative breast cancer. Biomaterials 2014, 35, 4333–4344. [Google Scholar] [CrossRef]
- Devulapally, R.; Sekar, N.M.; Sekar, T.V.; Foygel, K.; Massoud, T.F.; Willmann, J.K.; Paulmurugan, R. Polymer nanoparticles mediated codelivery of antimiR-10b and antimiR-21 for achieving triple negative breast cancer therapy. ACS Nano 2015, 9, 2290–2302. [Google Scholar] [CrossRef] [Green Version]
- Chin, Y.R.; Yuan, X.; Balk, S.P.; Toker, A. PTEN-deficient tumors depend on AKT2 for maintenance and survival. Cancer Discov. 2014, 4, 942–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, S.H.; Choi, Y.; Choi, J. Stimuli-responsive nanomaterials for application in antitumor therapy and drug delivery. Pharmaceutics 2020, 12, 630. [Google Scholar] [CrossRef]
- Shu, D.; Li, H.; Shu, Y.; Xiong, G.F.; Carson, W.E.; Haque, F.; Xu, R.; Guo, P.X. Systemic delivery of anti-miRNA for suppression of triple negative breast cancer utilizing RNA nanotechnology. ACS Nano 2015, 9, 9731–9740. [Google Scholar] [CrossRef]
- Deng, Z.J.; Morton, S.W.; Ben-Akiva, E.; Dreaden, E.C.; Shopsowitz, K.E.; Hammond, P.T. Layer-by-layer nanoparticles for systemic codelivery of an anticancer drug and siRNA for potential triple-negative breast cancer treatment. ACS Nano 2013, 7, 9571–9584. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhu, Y.H.; Mao, C.Q.; Dou, S.; Shen, S.; Tan, Z.B.; Wang, J. Triple negative breast cancer therapy with CDK1 siRNA delivered by cationic lipid assisted PEG-PLA nanoparticles. J. Control. Release 2014, 192, 114–121. [Google Scholar] [CrossRef]
- Li, Y.F.; Xiao, Y.J.; Lin, H.P.; Reichel, D.; Bae, Y.; Lee, E.Y.; Jiang, Y.G.; Huang, X.F.; Yang, C.F.; Wang, Z.S. In vivo beta-catenin attenuation by the integrin alpha 5-targeting nano-delivery strategy suppresses triple negative breast cancer sternness and metastasis. Biomaterials 2019, 188, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.C.; Zaks, T.; Langer, R.; Dong, Y.Z. Lipid nanoparticles for mRNA delivery. Nat. Rev. Mater. 2021, 6, 1078–1094. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Li, K.; Hu, B.; Li, C.; Zhang, M.; Hussain, A.; Wang, X.; Cheng, Q.; Yang, F.; Ge, K.; et al. Membrane-destabilizing ionizable lipid empowered imaging-guided siRNA delivery and cancer treatment. Exploration 2021, 1, 35–49. [Google Scholar] [CrossRef]
- Cai, K.M.; He, X.; Song, Z.Y.; Yin, Q.; Zhang, Y.F.; Uckun, F.M.; Jiang, C.; Cheng, J.J. Dimeric drug polymeric nanoparticles with exceptionally high drug loading and quantitative loading efficiency. J. Am. Chem. Soc. 2015, 137, 3458–3461. [Google Scholar] [CrossRef]
- Wei, D.S.; Yu, Y.J.; Huang, Y.; Jiang, Y.M.; Zhao, Y.; Nie, Z.X.; Wang, F.Y.; Ma, W.; Yu, Z.Q.; Huang, Y.Y.; et al. A near-infrared-II polymer with tandem fluorophores demonstrates superior biodegradability for simultaneous drug tracking and treatment efficacy feedback. ACS Nano 2021, 15, 5428–5438. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.W.; Tam, D.Y.; Xu, H.L.; Chan, M.S.; Liu, L.S.; Bolze, F.; Sun, X.H.; Lo, P.K. Conformational change of self-assembled DNA nanotubes induced by two-photon excitation. Small 2015, 11, 4090–4096. [Google Scholar] [CrossRef] [PubMed]
- Han, J.P.; Cui, Y.C.; Gu, Z.; Yang, D.Y. Controllable assembly/disassembly of polyphenol-DNA nanocomplex for cascade-responsive drug release in cancer cells. Biomaterials 2021, 273, 120846. [Google Scholar] [CrossRef]
- Zhang, H.Q.; Li, F.; Li, X.F.; Le, X.C. Yoctomole detection of proteins using solid phase binding-induced DNA assembly. Methods 2013, 64, 322–330. [Google Scholar] [CrossRef]
- Chen, Z.B.; Li, L.D.; Tian, Y.; Mu, X.J.; Guo, L. Signal amplification architecture for electrochemical aptasensor based on network-like thiocyanuric acid/gold nanoparticle/ssDNA. Biosens. Bioelectron. 2012, 38, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Lo, P.K.; Sleiman, H.F. Nucleobase-Templated Polymerization: Copying the chain length and polydispersity of living polymers into conjugated polymers. J. Am. Chem. Soc. 2009, 131, 4182–4183. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Duplex Formation | Nuclease Stability | RNase H Recruitment | Commercial Availability | Limitations |
|---|---|---|---|---|---|
| PS ONs | yes | increased | capable | yes | relatively unstable duplex |
| PNA | enhanced binding affinity | increased | incapable | yes | low aqueous solubility, self-aggregation |
| 2′-OMe RNA | stronger binding affinity to RNA than DNA | increased | incapable | yes | reduced silencing activity of modified siRNA |
| 2′-F RNA | enhanced binding affinity to RNA | not significantly increased | incapable | yes | other modifications required to enhance nuclease stability |
| LNA | increased | increased | poor substrate | yes | severe hepatotoxicity |
| TNA | stronger binding affinity to RNA than DNA | increased | incapable | no | Limited length in chemical synthesis |
| Sensor | Target | Detection Limit/Range | Ref. |
|---|---|---|---|
| DNA-AuNPs nano-flares | Surviving mRNA | in vitro imaging | [51] |
| Graphene-DNAzyme | Cu2+ | 0.365 nM | [54] |
| Ribozyme-based biosensor | TPP | a few nM | [55] |
| DNAzyme sensor | Li+ | in vitro imaging | [56] |
| electrochemical aptasensor | ATP | 10 nM to 1 mM | [64] |
| Aptamer-modified graphene transistor | E. coli | 102 CFU/mL | [65] |
| Aptamer-modified DNA nanotube | thrombin, ATP, and insulin | ~17.6 nM, ~116 nM, and ~55 nM | [66] |
| Aptamer-modified Ag2S nanodots | CTCs | 6 tumor cells/mL | [68] |
| PNA electrochemical biosensor | DNA | 10 pmol | [96] |
| PNA-graphene oxide | dsDNA | 260 pM | [97] |
| PNA-AuNPs | single nucleotide polymorphism | 2.3 nM | [98] |
| PNA-graphene oxide | miRNAs | ~1 pM | [99] |
| LNA MB | single nucleotide polymorphism | NA | [141] |
| LNA electrochemical biosensor | miRNA | 77 aM | [142] |
| LNA-modified PT biosensor | P53 DNA sequence | 60 pM | [143] |
| TNA-based biosensor | SARS-CoV-2 RNA | ≤20 aM | [189] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Li, P.; Chu, H.C.; Lo, P.K. Nucleic Acids and Their Analogues for Biomedical Applications. Biosensors 2022, 12, 93. https://doi.org/10.3390/bios12020093
Wang F, Li P, Chu HC, Lo PK. Nucleic Acids and Their Analogues for Biomedical Applications. Biosensors. 2022; 12(2):93. https://doi.org/10.3390/bios12020093
Chicago/Turabian StyleWang, Fei, Pan Li, Hoi Ching Chu, and Pik Kwan Lo. 2022. "Nucleic Acids and Their Analogues for Biomedical Applications" Biosensors 12, no. 2: 93. https://doi.org/10.3390/bios12020093
APA StyleWang, F., Li, P., Chu, H. C., & Lo, P. K. (2022). Nucleic Acids and Their Analogues for Biomedical Applications. Biosensors, 12(2), 93. https://doi.org/10.3390/bios12020093