Label-Free Detection of T4 Polynucleotide Kinase Activity and Inhibition via Malachite Green Aptamer Generated from Ligation-Triggered Transcription


Abstract
:1. Introduction
2. Materials and Methods
2.1. Apparatus
2.2. Materials and Reagents
2.3. Amplified Fluorescence Assay for T4 PNK-Catalyzed Phosphorylation
2.4. Selectivity Evaluation of the Assay
2.5. Kinase Inhibitor Screening
2.6. Determination of T4 PNK Activity in Diluted Cell Lysates
3. Results and Discussion
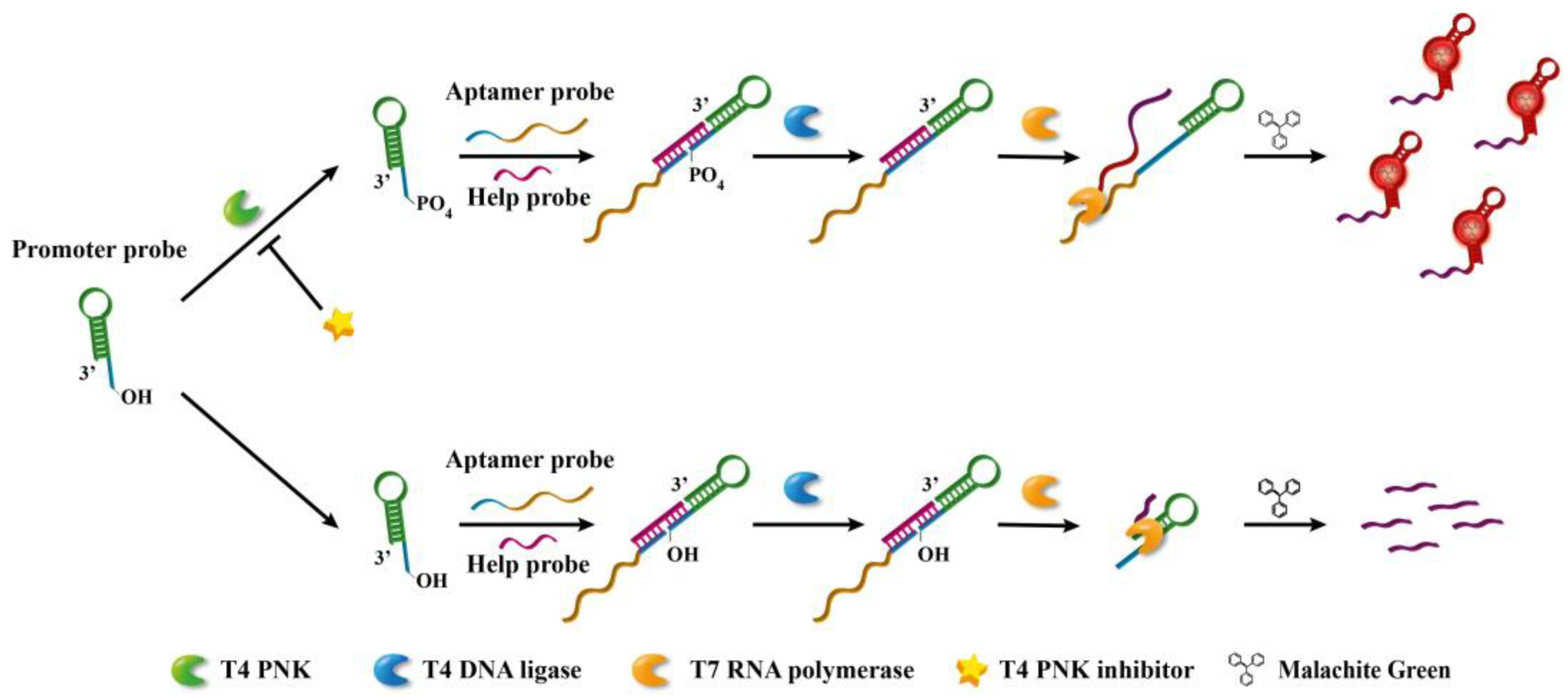
3.1. Sensing Principle of the Proposed Assay for T4 PNK Activity
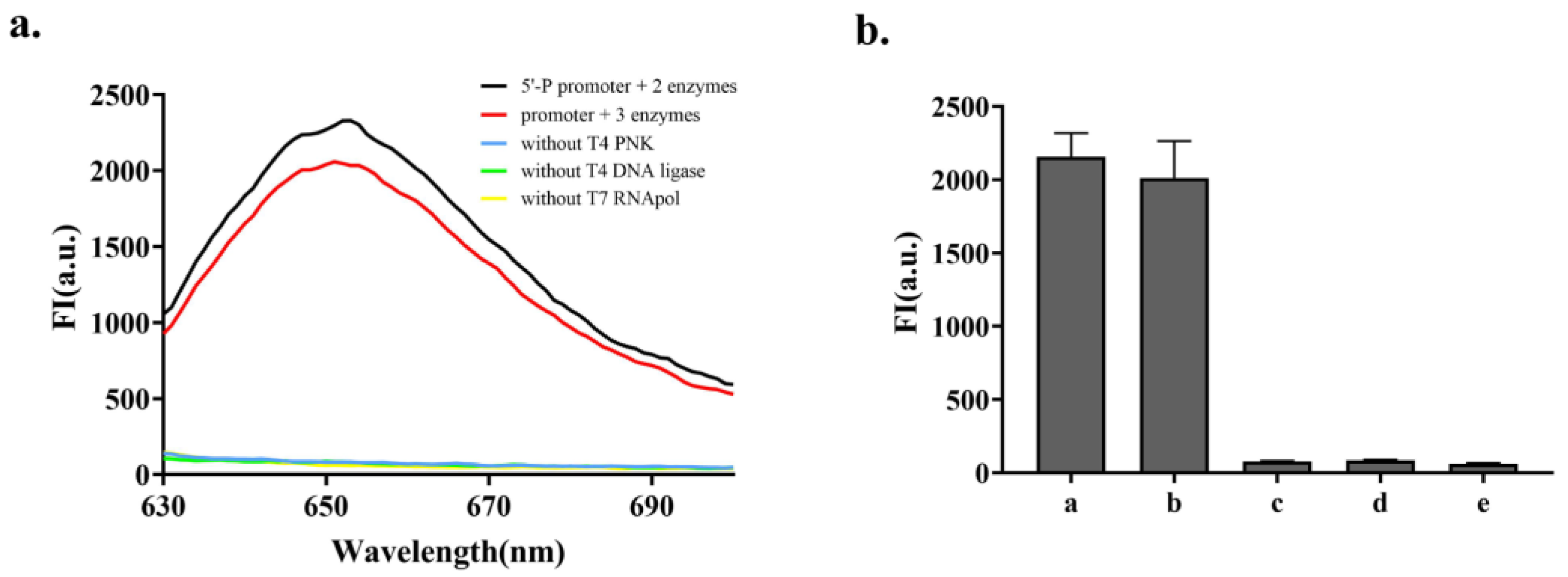
3.2. Feasibility Verification of the Strategy
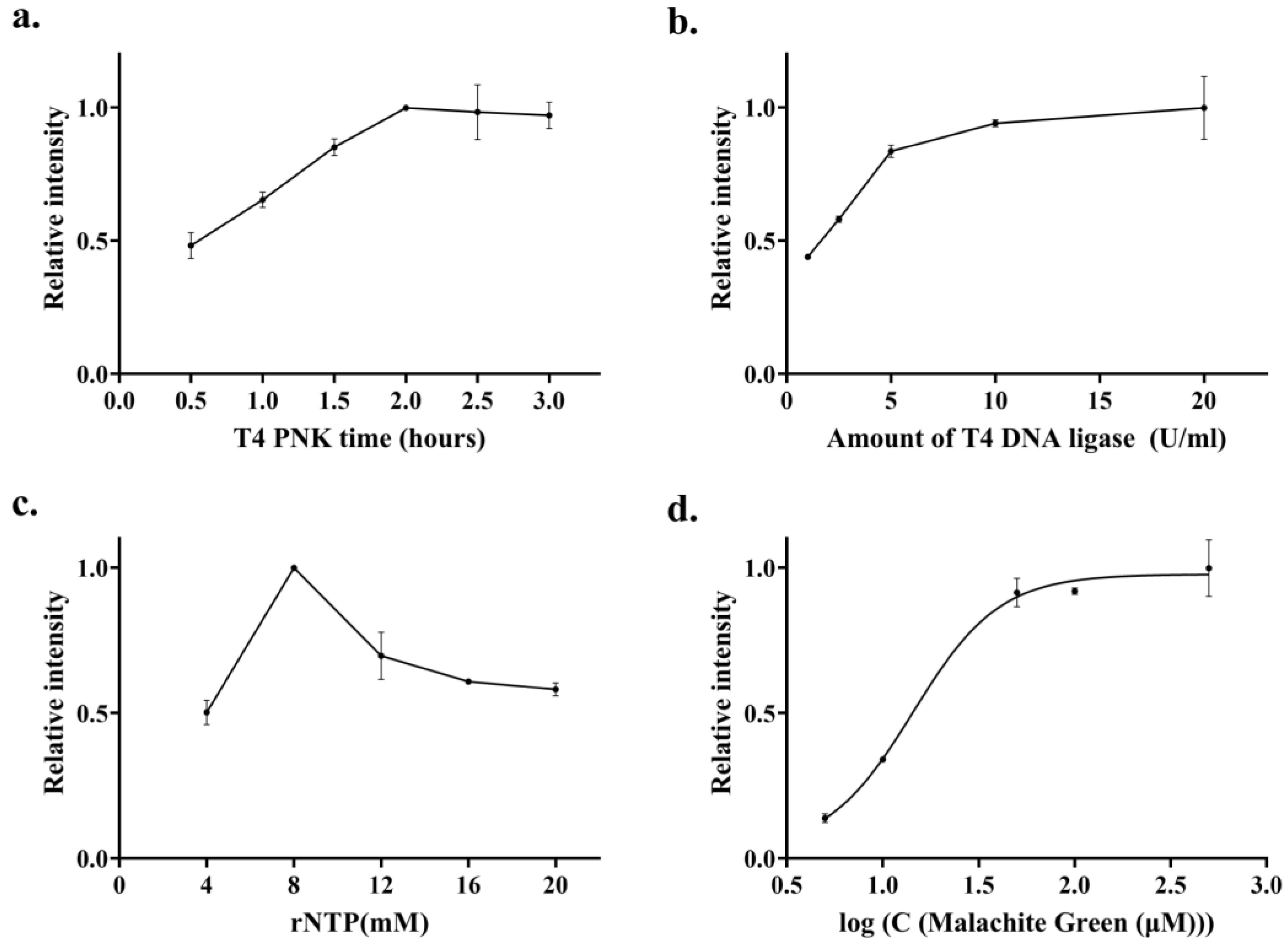
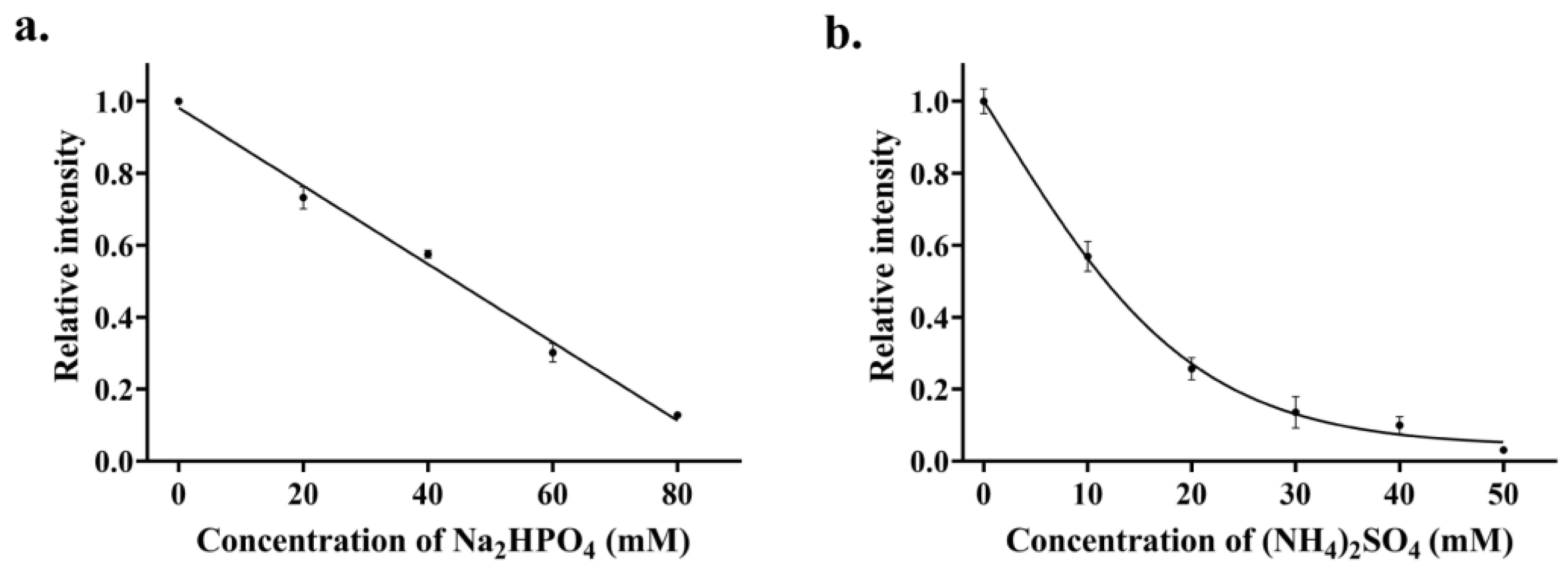
3.3. Optimization of Experimental Conditions
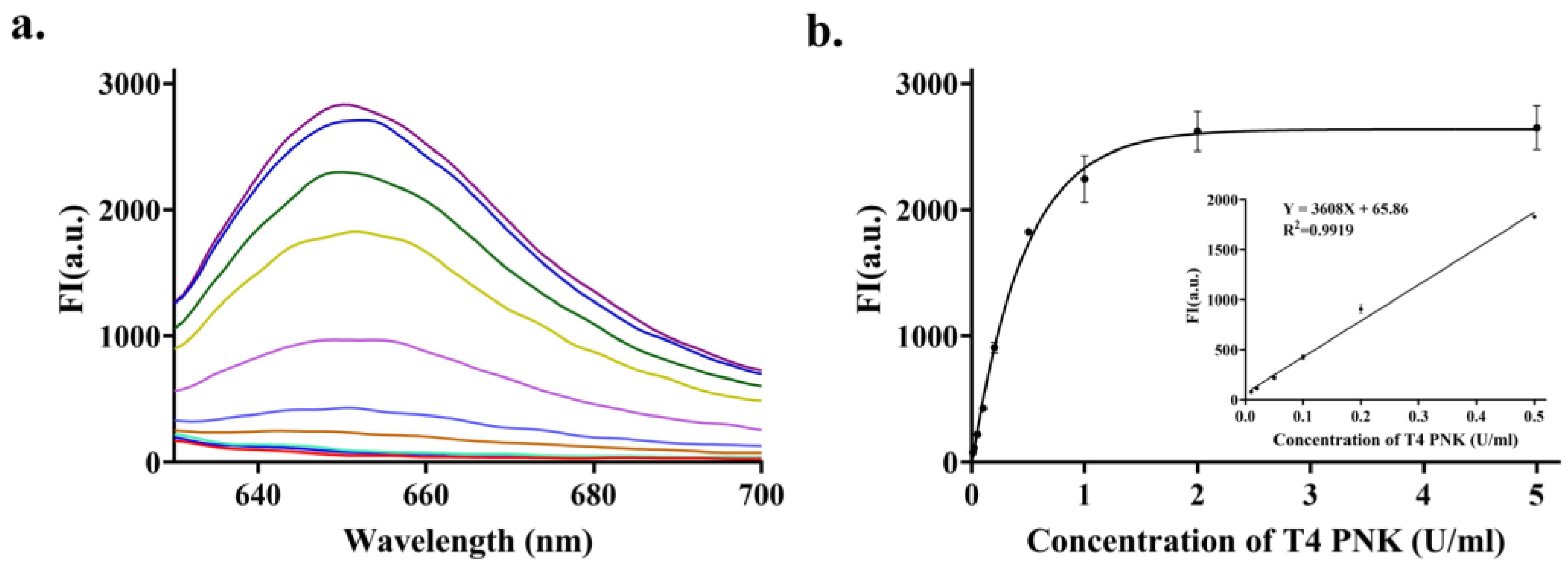
3.4. Quantitative Detection of T4 PNK Activity
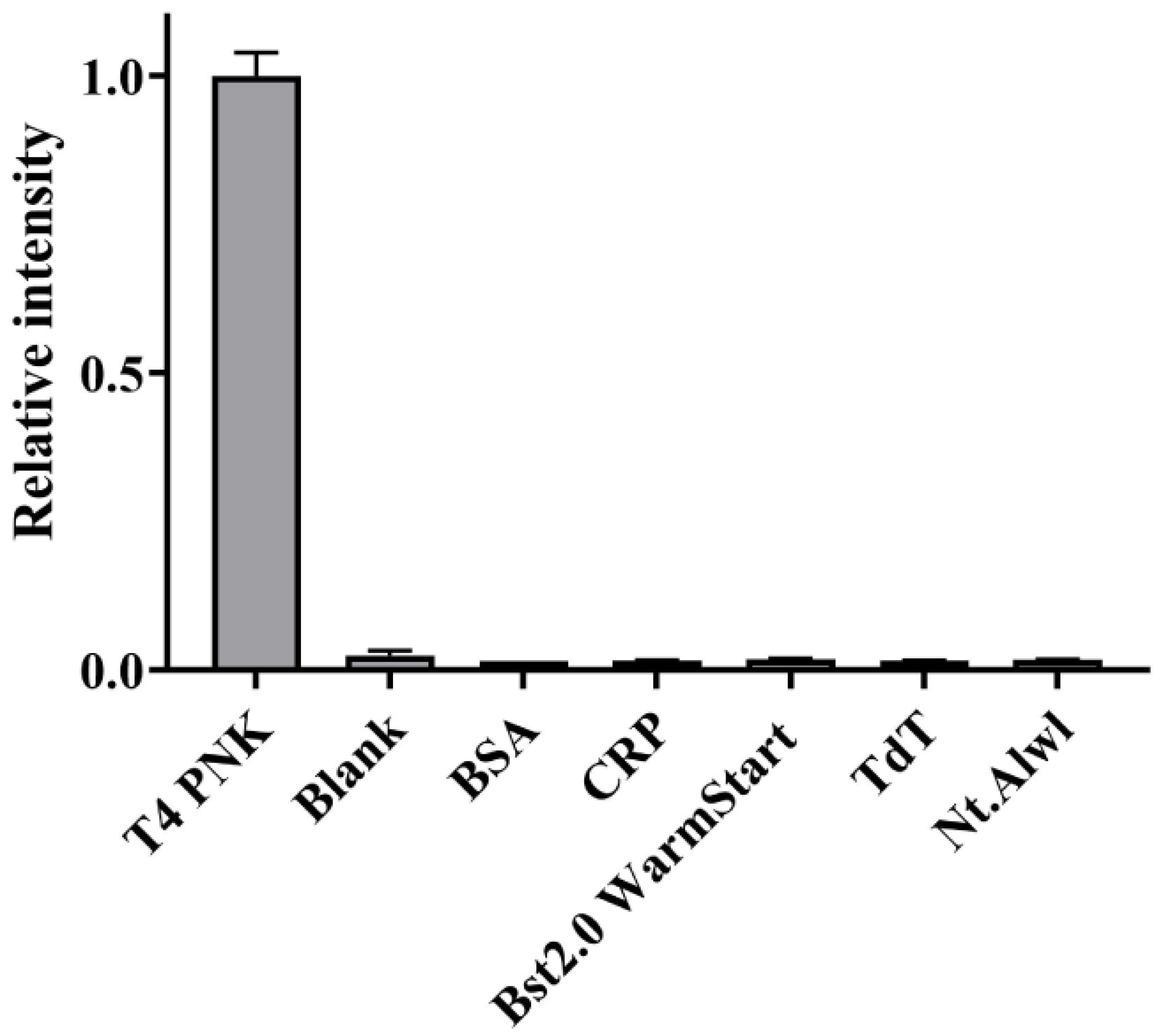
3.5. Selectivity Evaluation of the Assay
3.6. Inhibition Evaluation of T4 PNK Activity
3.7. PNK Activity Detection in Diluted Cell Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bukhari, A.B.; Chan, G.K.; Gamper, A.M. Targeting the DNA Damage Response for Cancer Therapy by Inhibiting the Kinase Wee1. Front. Oncol. 2022, 12, 828684. [Google Scholar] [CrossRef]
- Sun, C.; Fang, Y.; Labrie, M.; Li, X.; Mills, G.B. Systems approach to rational combination therapy: PARP inhibitors. Biochem. Soc. Trans. 2020, 48, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zou, Z.; Luo, X.; Mi, Y.; Chang, H.; Xing, D. LRH1 enhances cell resistance to chemotherapy by transcriptionally activating MDC1 expression and attenuating DNA damage in human breast cancer. Oncogene 2018, 37, 3243–3259. [Google Scholar] [CrossRef]
- Rocha, C.R.R.; Silva, M.M.; Quinet, A.; Cabral-Neto, J.B.; Menck, C.F.M. DNA repair pathways and cisplatin resistance: An intimate relationship. Clinics 2018, 73, e478s. [Google Scholar] [CrossRef]
- Chen, S.H.; Chang, J.Y. New Insights into Mechanisms of Cisplatin Resistance: From Tumor Cell to Microenvironment. Int. J. Mol. Sci. 2019, 20, 4136. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Sinha, D.; Bhattacharya, S.; Srinivasan, K.; Abdisalaam, S.; Asaithamby, A. Werner Syndrome Protein and DNA Replication. Int. J. Mol. Sci. 2018, 19, 3442. [Google Scholar] [CrossRef] [Green Version]
- Schiess, N.; Zee, D.S.; Siddiqui, K.A.; Szolics, M.; El-Hattab, A.W. Novel PNKP mutation in siblings with ataxia-oculomotor apraxia type 4. J. Neurogenet. 2017, 31, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.K.; Lima, C.D.; Shuman, S. Structure and mechanism of T4 polynucleotide kinase: An RNA repair enzyme. EMBO J. 2002, 21, 3873–3880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, C.C. Phosphorylation of nucleic acid by an enzyme from T4 bacteriophage-infected Escherichia coli. Proc. Natl. Acad. Sci. USA 1965, 54, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.K.; Shuman, S. Domain structure and mutational analysis of T4 polynucleotide kinase. J. Biol. Chem. 2001, 276, 26868–26874. [Google Scholar] [CrossRef] [Green Version]
- Karimi-Busheri, F.; Daly, G.; Robins, P.; Canas, B.; Pappin, D.J.; Sgouros, J.; Miller, G.G.; Fakhrai, H.; Davis, E.M.; Le Beau, M.M.; et al. Molecular characterization of a human DNA kinase. J. Biol. Chem. 1999, 274, 24187–24194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Yan, C.; Jiang, J.; Yu, R. Colorimetric assay for T4 polynucleotide kinase activity based on the horseradish peroxidase-mimicking DNAzyme combined with λ exonuclease cleavage. Anal. Chim. Acta 2013, 766, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Li, Y.; Lu, M.; Tang, B.; Zhang, C.Y. An ultrasensitive electrochemical biosensor for polynucleotide kinase assay based on gold nanoparticle-mediated lambda exonuclease cleavage-induced signal amplification. Biosens. Bioelectron. 2018, 99, 1–7. [Google Scholar] [CrossRef]
- Wang, Y.; He, X.; Wang, K.; Ni, X.; Su, J.; Chen, Z. Ferrocene-functionalized SWCNT for electrochemical detection of T4 polynucleotide kinase activity. Biosens. Bioelectron. 2012, 32, 213–218. [Google Scholar] [CrossRef]
- Hou, T.; Wang, X.; Liu, X.; Pan, C.; Li, F. Sensitive electrochemical assay for T4 polynucleotide kinase activity based on dual-signaling amplification coupled with exonuclease reaction. Sens. Actuators B Chem. 2014, 202, 588–593. [Google Scholar] [CrossRef]
- Liu, S.; Ming, J.; Lin, Y.; Wang, C.; Cheng, C.; Liu, T.; Wang, L. Highly sensitive detection of T4 polynucleotide kinase activity by coupling split DNAzyme and ligation-triggered DNAzyme cascade amplification. Biosens. Bioelectron. 2014, 55, 225–230. [Google Scholar] [CrossRef]
- Yu, L.-l.; Lan, W.; Xu, H.T.; Chen, H.; Bai, L.; Wang, W.J.S.; B-chemical, A. Label-free detection of Hg2+ based on Hg2+-triggered toehold binding, Exonuclease III assisted target recycling and hybridization chain reaction. Sens. Actuators B Chem. 2017, 248, 411–418. [Google Scholar] [CrossRef]
- Tang, Z.; Wang, K.; Tan, W.; Ma, C.; Li, J.; Liu, L.; Guo, Q.; Meng, X. Real-time investigation of nucleic acids phosphorylation process using molecular beacons. Nucleic Acids Res. 2005, 33, e97. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Xu, Y.; Dai, L.; Liu, X.; Kong, D. Ultrasensitive, label-free detection of T4 ligase and T4 polynucleotide kinase based on target-triggered hyper-branched rolling circle amplification. Sens. Actuators B Chem. 2018, 260, 70–77. [Google Scholar] [CrossRef]
- Tang, W.; Zhu, G.; Zhang, C.Y. Sensitive detection of polynucleotide kinase using rolling circle amplification-induced chemiluminescence. Chem. Commun. 2014, 50, 4733–4735. [Google Scholar] [CrossRef]
- Hou, T.; Wang, X.; Lu, T.; Liu, X.; Li, F. Sensitive detection of T4 polynucleotide kinase activity based on coupled exonuclease reaction and nicking enzyme-assisted fluorescence signal amplification. Anal. Bioanal. Chem. 2014, 406, 2943–2948. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.; Tao, M.; Shi, Z.; Zhang, X.; Jin, Y.; Li, B. Label-free and sensitive detection of T4 polynucleotide kinase activity via coupling DNA strand displacement reaction with enzymatic-aided amplification. Biosens. Bioelectron. 2015, 73, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Raso, A.; Mascelli, S.; Nozza, P.; Ugolotti, E.; Vanni, I.; Capra, V.; Biassoni, R. Troubleshooting fine-tuning procedures for qPCR system design. J. Clin. Lab. Anal. 2011, 25, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Milligan, J.F.; Groebe, D.R.; Witherell, G.W.; Uhlenbeck, O.C. Oligoribonucleotide synthesis using T7 RNA polymerase and synthetic DNA templates. Nucleic Acids Res. 1987, 15, 8783–8798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouellet, J. RNA Fluorescence with Light-Up Aptamers. Front. Chem. 2016, 4, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autour, A.; Westhof, E.; Ryckelynck, M. iSpinach: A fluorogenic RNA aptamer optimized for in vitro applications. Nucleic Acids Res. 2016, 44, 2491–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, T.; Shin, J.; Choi, H.-J.; Park, K.S. Split T7 promoter-based isothermal transcription amplification for one-step fluorescence detection of SARS-CoV-2 and emerging variants. Biosens. Bioelectron. 2022, 208, 114221. [Google Scholar] [CrossRef]
- Wang, L.-j.; Liang, L.; Liu, B.-j.; Jiang, B.; Zhang, C.-y. A controlled T7 transcription-driven symmetric amplification cascade machinery for single-molecule detection of multiple repair glycosylases. Chem. Sci. 2021, 12, 5544–5554. [Google Scholar] [CrossRef]
- Yin, B.-C.; Wu, S.; Ma, J.-L.; Ye, B.-C. A novel molecular beacon-based method for isothermal detection of sequence-specific DNA via T7 RNA polymerase-aided target regeneration. Biosens. Bioelectron. 2015, 68, 365–370. [Google Scholar] [CrossRef]
- Kolpashchikov, D.M. Binary malachite green aptamer for fluorescent detection of nucleic acids. J. Am. Chem. Soc. 2005, 127, 12442–12443. [Google Scholar] [CrossRef]
- Yerramilli, V.S.; Kim, K.H. Labeling RNAs in live cells using malachite green aptamer scaffolds as fluorescent probes. ACS Synth. Biol. 2018, 7, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Rong, M.; Durbin, R.K.; McAllister, W.T. Template Strand Switching by T7 RNA Polymerase *. J. Biol. Chem. 1998, 273, 10253–10260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartje, Z.J.; Janis, H.I.; Mukhopadhyay, S.; Gagnon, K.T. Revisiting T7 RNA polymerase transcription in vitro with the Broccoli RNA aptamer as a simplified real-time fluorescent reporter. J. Biol. Chem. 2021, 296, 100175. [Google Scholar] [CrossRef] [PubMed]
- Woo, C.H.; Jang, S.; Shin, G.; Jung, G.Y.; Lee, J.W. Sensitive fluorescence detection of SARS-CoV-2 RNA in clinical samples via one-pot isothermal ligation and transcription. Nat. Biomed. Eng. 2020, 4, 1168–1179. [Google Scholar] [CrossRef] [PubMed]
- He, H.-Z.; Leung, K.-H.; Wang, W.; Chan, D.S.-H.; Leung, C.-H.; Ma, D.-L. Label-free luminescence switch-on detection of T4 polynucleotide kinase activity using a G-quadruplex-selective probe. Chem. Commun. 2014, 50, 5313–5315. [Google Scholar] [CrossRef]
- Lin, L.; Liu, Y.; Zhao, X.; Li, J. Sensitive and Rapid Screening of T4 Polynucleotide Kinase Activity and Inhibition Based on Coupled Exonuclease Reaction and Graphene Oxide Platform. Anal. Chem. 2011, 83, 8396–8402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, J.; Zhang, H.; Jiang, J.; Yu, R. Double strand DNA-templated copper nanoparticle as a novel fluorescence indicator for label-free detection of polynucleotide kinase activity. Biosens. Bioelectron. 2013, 44, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xiong, Y.; Lin, L.; Zhang, X.; Wang, G. A simple label-free electrochemical method for the detection of polynucleotide kinase activity by a peroxidase mimic: TiO2 nanotube array. Anal. Methods 2015, 7, 10345–10349. [Google Scholar] [CrossRef]
- Song, C.; Zhao, M. Real-Time Monitoring of the Activity and Kinetics of T4 Polynucleotide Kinase by a Singly Labeled DNA-Hairpin Smart Probe Coupled with λ Exonuclease Cleavage. Anal. Chem. 2009, 81, 1383–1388. [Google Scholar] [CrossRef]
- Song, C.; Yang, X.; Wang, K.; Wang, Q.; Liu, J.; Huang, J.; He, L.; Liu, P.; Qing, Z.; Liu, W. A sensitive detection of T4 polynucleotide kinase activity based on β-cyclodextrin polymer enhanced fluorescence combined with an exonuclease reaction. Chem. Commun. 2015, 51, 1815–1818. [Google Scholar] [CrossRef]
- Chen, F.; Zhao, Y.; Qi, L.; Fan, C. One-step highly sensitive florescence detection of T4 polynucleotide kinase activity and biological small molecules by ligation-nicking coupled reaction-mediated signal amplification. Biosens. Bioelectron. 2013, 47, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Guo, J.; Song, Y.; Zhu, Z.; Yang, C.J. Detection of T4 Polynucleotide Kinase via Allosteric Aptamer Probe Platform. ACS Appl. Mater. Interfaces 2017, 9, 38356–38363. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ming, J.; Lin, Y.; Wang, C.; Liu, T.; Cheng, C.; Li, F. Amplified detection of T4 polynucleotide kinase activity based on a λ-exonuclease cleavage-induced DNAzyme releasing strategy. Sens. Actuators B Chem. 2014, 192, 157–163. [Google Scholar] [CrossRef]
- Lillehaug, J.R.; Kleppe, K. Effect of salts and polyamines on T4 polynucleotide kinase. Biochemistry 1975, 14, 1225–1229. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) |
|---|---|
| Promoter probe | AGGTTACAGATGTGGCCCTATAGTGAGTCGTATTAATTTCGCGACAACACGCGAAATTAATACGACTCACTATAGGG |
| 5′ P-promoter probe | P-AGGTTACAGATGTGGCCCTATAGTGAGTCGTATTAATTTCGCGACAACACGCGAAATTAATACGACTCACTATAGGG |
| Aptamer probe | GGATCCATTCGTTACCTGGCTCTCGCCAGTCGGGATCCTGATAGCTTGACAAC |
| Help probe | CCACATCTGTAACCTGTTGTCAAGCTATCA |
| Sensing Strategy | Detection Limit | Ref. |
|---|---|---|
| G-quadruplex-based luminescent assay coupled with λ exo cleavage reaction | 0.05 U/mL | [35] |
| Graphene oxide-based fluorescent assay assisted by λ exo | 0.05 U/mL | [36] |
| Fluorescent assay with dsDNA-templated CuNPs as the fluorescence indicator | 0.49 U/mL | [37] |
| Colorimetric assay based on horseradish peroxidase-mimicking DNAzyme combined with λ exonuclease cleavage | 0.06 U/mL | [12] |
| Electrochemical assay by TiO2 nanotube array as a peroxidase mimic | 0.15 U/mL | [38] |
| Electrochemical Assay Based on Dual-Signaling Amplification Coupled with Exonuclease Reaction | 0.02 U/mL | [15] |
| Ferrocene-functionalized SWCNT for electrochemical detection | 0.01 U/mL | [14] |
| Fluorescent assay by a Singly Labeled DNA-Hairpin Smart Probe Coupled with λ Exonuclease Cleavage | 0.04 U/mL | [39] |
| Fluorescent assay based on β-cyclodextrin polymer combined with an exonuclease reaction | 0.02 U/mL | [40] |
| Fluorescent detection assay via malachite green aptamer generated from ligation-triggered transcription | 0.01 U/mL | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Si, J.; Zhou, W.; Fang, Y.; Zhou, D.; Gao, Y.; Yao, Q.; Shen, X.; Zhu, C. Label-Free Detection of T4 Polynucleotide Kinase Activity and Inhibition via Malachite Green Aptamer Generated from Ligation-Triggered Transcription. Biosensors 2023, 13, 449. https://doi.org/10.3390/bios13040449
Si J, Zhou W, Fang Y, Zhou D, Gao Y, Yao Q, Shen X, Zhu C. Label-Free Detection of T4 Polynucleotide Kinase Activity and Inhibition via Malachite Green Aptamer Generated from Ligation-Triggered Transcription. Biosensors. 2023; 13(4):449. https://doi.org/10.3390/bios13040449
Chicago/Turabian StyleSi, Jingyi, Wei Zhou, Ying Fang, Da Zhou, Yifan Gao, Qunyan Yao, Xizhong Shen, and Changfeng Zhu. 2023. "Label-Free Detection of T4 Polynucleotide Kinase Activity and Inhibition via Malachite Green Aptamer Generated from Ligation-Triggered Transcription" Biosensors 13, no. 4: 449. https://doi.org/10.3390/bios13040449
APA StyleSi, J., Zhou, W., Fang, Y., Zhou, D., Gao, Y., Yao, Q., Shen, X., & Zhu, C. (2023). Label-Free Detection of T4 Polynucleotide Kinase Activity and Inhibition via Malachite Green Aptamer Generated from Ligation-Triggered Transcription. Biosensors, 13(4), 449. https://doi.org/10.3390/bios13040449