Comparison of Single-Stranded DNA Probes Conjugated with Magnetic Particles for Trans-Cleavage in Cas12a-Based Biosensors

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Syntheses of dsDNA Targets for Activation of Cas12a (Cis-Targets)
2.3. Syntheses of gRNAs
2.4. Conjugation of ssDNA Reporters with MPs
2.5. Synthesis of Au@Pt Nanozyme and Its Conjugate with PolyA-80
2.6. Trans-Cleavage of ssDNA Reporters Attached to MPs Using Cas12a
2.7. Detection of dsDNA Targets Using CRISPR–Cas12 Assay in a Homogeneous Format
2.8. Detection of dsDNA Targets Using CRISPR–Cas12 Assay in a Heterogeneous Format
3. Results
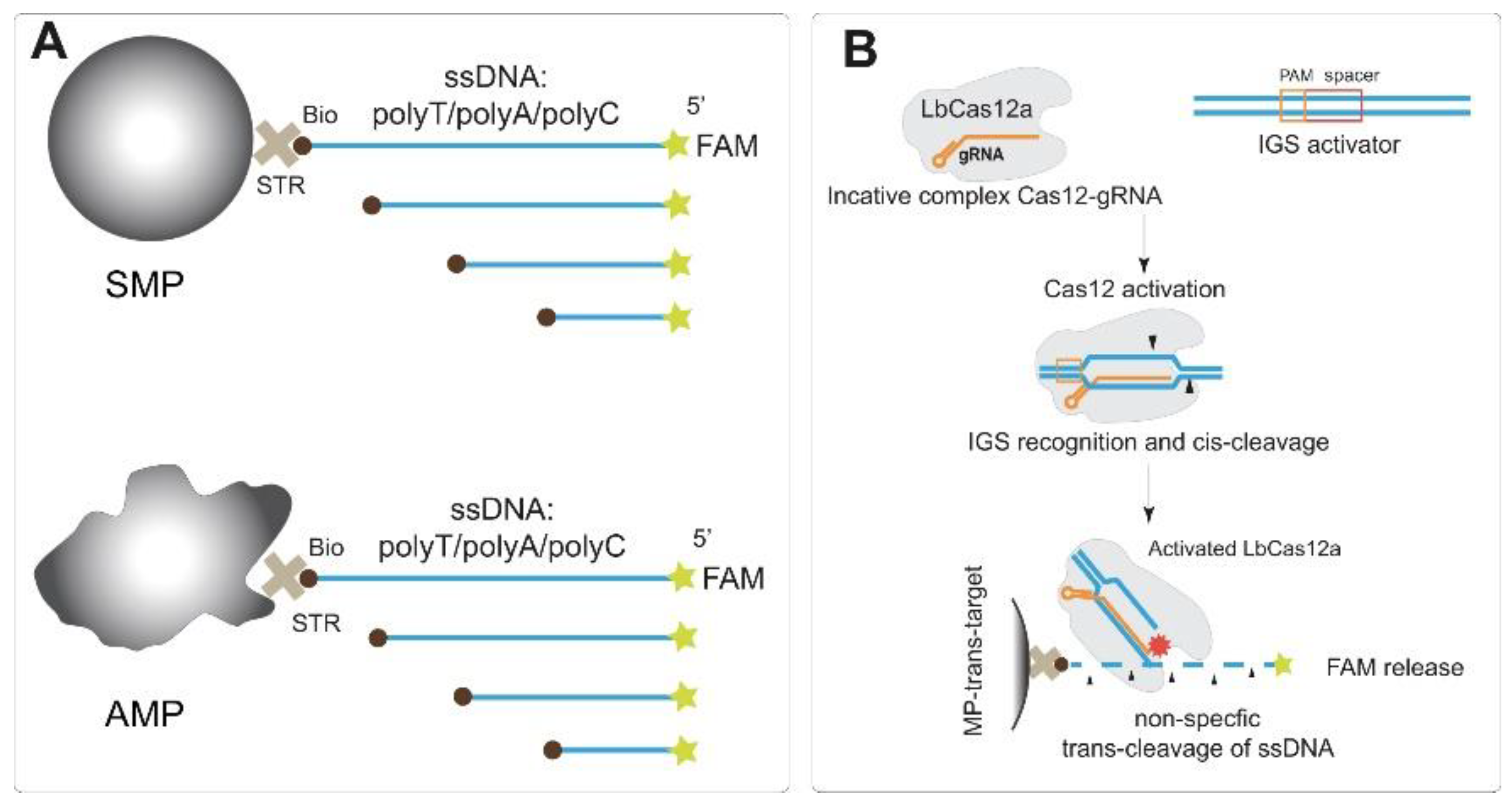
3.1. Design of Experiments
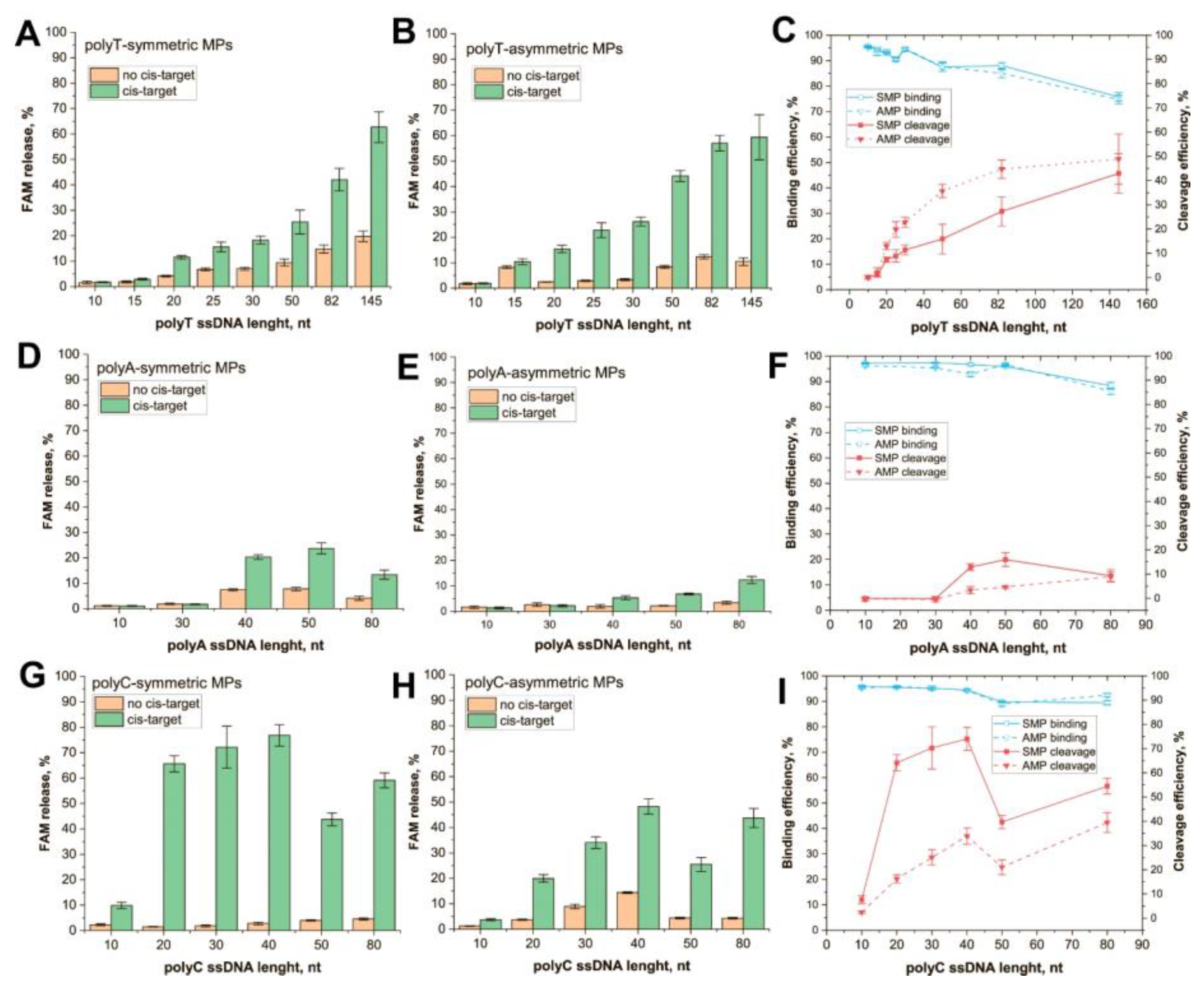
3.2. Trans-Cleavage of Different Types of ssDNA Immobilized on MPs
- PolyC ssDNA is the most effective trans-target that compares polyA or polyT of the same length (see Figure 2C,F,I). This feature is more expressed for SMP conjugates. It agrees with data from previous research that establish polyC as being the most effective trans-target in the solution. In our research, the effectivity of cleavage increases in the row polyA < polyT < polyC, wherein, the cleavage of each immobilized ssDNA was less effective than the cleavage of the corresponding free ssDNA (Figures S6 and S7). This can be caused by steric hindrance and/or restricted diffusion of Cas12s near the MP surface. In experiments in which polyT was immobilized on the gold layer, the cleavage efficiency was 20–25% of its maximum value [17]. Meanwhile, conjugate polyT, with small (13 nm) GNPs, demonstrated the same cleavage efficiency as unbound polyT [25]. As such, it can be expected that the size of carrier particles influences the extent of the decrease in cleavage efficiency.
- No linear dependence of efficiency was observed from trans = target length. The dependencies had hyperbolic saturation or a bell shape. The minimum length for the cleavage was determined using the type of homopolymers, following the row polyC < polyT < polyA. Thus, the most effective homopolymer for the cleavage of the shorter oligonucleotide can be used. At the same time, the length for maximally effective cleavage (20–50 nt) is significantly higher for MP-bound targets than for soluble trans-targets (8 nt) [11]. This could be a result of steric and/or charge effects of the MP surface. An AMP with a more heterogenous surface showed less efficient trans-cleavage.
- There was no unambiguous increase in cleavage efficiency with increasing ssDNA length. Limitations for long ssDNAs can be explained due to the loss of linear structure. Curving trends appear even for very short ssDNAs, starting from 0.7–6 nm in length [34,35]. Pyrimidine polymers are more flexible; for some lengths, they tend to form compact globules [36]. Unlike them, polyA oligonucleotides tend to form rod conformations that resemble dsDNA due to adenine stacking [37,38]. This property of polyA ssDNAs would potentially require an increase in their accessibility to Cas12a, but we did not observe it. In the case of SMPs, some interactions of the nucleotides with the MP surface could lead to lower cleavage efficiency for long ssDNAs. Moreover, the binging of long ssDNA to MPs is worse than short ones.
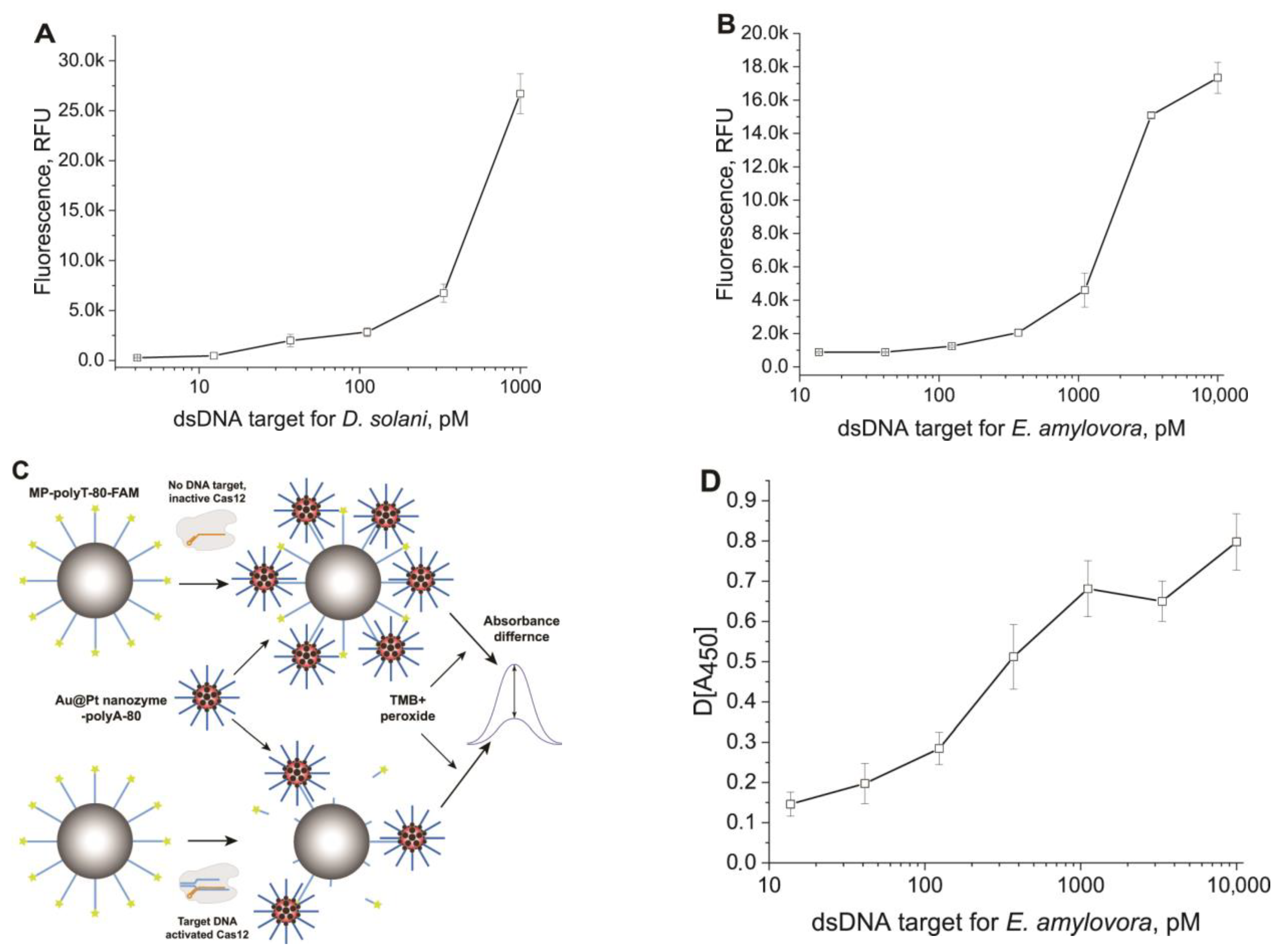
3.3. Detection of dsDNA Targets Using CRISPR–Cas12 Assay in Homogeneous and Heterogeneous Formats
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Wu, Y.; Wu, Y.; Chang, Y.; Liu, M. CRISPR-Cas systems: From gene scissors to programmable biosensors. TrAC Trends Anal. Chem. 2021, 137, 116210. [Google Scholar] [CrossRef]
- Ghouneimy, A.; Mahas, A.; Marsic, T.; Aman, R.; Mahfouz, M. CRISPR-Based Diagnostics: Challenges and Potential Solutions toward Point-of-Care Applications. ACS Synth. Biol. 2023, 12, 1–16. [Google Scholar] [CrossRef]
- Zhang, X.; Shi, Y.; Chen, G.; Wu, D.; Wu, Y.; Li, G. CRISPR/Cas Systems-Inspired Nano/Biosensors for Detecting Infectious Viruses and Pathogenic Bacteria. Small Methods 2022, 6, e2200794. [Google Scholar] [CrossRef]
- Stella, S.; Mesa, P.; Thomsen, J.; Paul, B.; Alcon, P.; Jensen, S.B.; Saligram, B.; Moses, M.E.; Hatzakis, N.S.; Montoya, G. Conformational Activation Promotes CRISPR-Cas12a Catalysis and Resetting of the Endonuclease Activity. Cell 2018, 175, 1856–1871.e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Zhang, H.; Le, X.C. Signal Amplification by the trans-Cleavage Activity of CRISPR-Cas Systems: Kinetics and Performance. Anal. Chem. 2023, 95, 206–217. [Google Scholar] [CrossRef]
- Kumaran, A.; Jude Serpes, N.; Gupta, T.; James, A.; Sharma, A.; Kumar, D.; Nagraik, R.; Kumar, V.; Pandey, S. Advancements in CRISPR-Based Biosensing for Next-Gen Point of Care Diagnostic Application. Biosensors 2023, 13, 202. [Google Scholar] [CrossRef]
- Xie, S.; Ji, Z.; Suo, T.; Li, B.; Zhang, X. Advancing sensing technology with CRISPR: From the detection of nucleic acids to a broad range of analytes—A review. Anal. Chim. Acta 2021, 1185, 338848. [Google Scholar] [CrossRef]
- Li, S.Y.; Cheng, Q.X.; Wang, J.M.; Li, X.Y.; Zhang, Z.L.; Gao, S.; Cao, R.B.; Zhao, G.P.; Wang, J. CRISPR-Cas12a-assisted nucleic acid detection. Cell Discov. 2018, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Lv, H.; Wang, J.; Zhang, J.; Chen, Y.; Yin, L.; Jin, D.; Gu, D.; Zhao, H.; Xu, Y.; Wang, J. Definition of CRISPR Cas12a Trans-Cleavage Units to Facilitate CRISPR Diagnostics. Front. Microbiol. 2021, 12, 766464. [Google Scholar] [CrossRef]
- Yue, H.; Shu, B.; Tian, T.; Xiong, E.; Huang, M.; Zhu, D.; Sun, J.; Liu, Q.; Wang, S.; Li, Y.; et al. Droplet Cas12a Assay Enables DNA Quantification from Unamplified Samples at the Single-Molecule Level. Nano Lett. 2021, 21, 4643–4653. [Google Scholar] [CrossRef]
- Wu, L.; Wang, X.; Wu, X.; Xu, S.; Liu, M.; Cao, X.; Tang, T.; Huang, X.; Huang, H. MnO2 Nanozyme-Mediated CRISPR-Cas12a System for the Detection of SARS-CoV-2. ACS Appl. Mater. Interfaces 2022, 14, 50534–50542. [Google Scholar] [CrossRef]
- Samanta, D.; Ebrahimi, S.B.; Ramani, N.; Mirkin, C.A. Enhancing CRISPR-Cas-Mediated Detection of Nucleic Acid and Non-nucleic Acid Targets Using Enzyme-Labeled Reporters. J. Am. Chem. Soc. 2022, 144, 16310–16315. [Google Scholar] [CrossRef]
- Yin, B.; Zhang, Q.; Xia, X.; Li, C.; Ho, W.K.H.; Yan, J.; Huang, Y.; Wu, H.; Wang, P.; Yi, C.; et al. A CRISPR-Cas12a integrated SERS nanoplatform with chimeric DNA/RNA hairpin guide for ultrasensitive nucleic acid detection. Theranostics 2022, 12, 5914–5930. [Google Scholar] [CrossRef]
- Yang, X.; Li, J.; Zhang, S.; Li, C.; Ma, J. Amplification-Free, Single-Microbead-Based Cas12a Assay for One-Step DNA Detection at the Single-Molecule Level. Anal. Chem. 2022, 94, 13076–13083. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Somoza, R.A.; Wang, L.; Welter, J.F.; Li, Y.; Caplan, A.I.; Liu, C.C. Exploring the Trans-Cleavage Activity of CRISPR-Cas12a (cpf1) for the Development of a Universal Electrochemical Biosensor. Angew. Chem. Int. Ed. Engl. 2019, 58, 17399–17405. [Google Scholar] [CrossRef]
- Shao, N.; Han, X.; Song, Y.; Zhang, P.; Qin, L. CRISPR-Cas12a Coupled with Platinum Nanoreporter for Visual Quantification of SNVs on a Volumetric Bar-Chart Chip. Anal. Chem. 2019, 91, 12384–12391. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.S.R.; Erdogmus, E.; Shokr, A.; Kandula, H.; Thirumalaraju, P.; Kanakasabapathy, M.K.; Hardie, J.M.; Pacheco, L.G.C.; Li, J.Z.; Kuritzkes, D.R.; et al. SARS-CoV-2 RNA Detection by a Cellphone-Based Amplification-Free System with CRISPR/CAS-Dependent Enzymatic (CASCADE) Assay. Adv. Mater. Technol. 2021, 6, 2100602. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, Z.; Li, C.; Hao, Y.; Tang, Y.; Yuan, Y.; Chai, L.; Fan, T.; Yu, J.; Ma, X.; et al. CRISPR-Cas12a-Empowered Electrochemical Biosensor for Rapid and Ultrasensitive Detection of SARS-CoV-2 Delta Variant. Nanomicro Lett. 2022, 14, 159. [Google Scholar] [CrossRef]
- Xu, W.; Jin, T.; Dai, Y.; Liu, C.C. Surpassing the detection limit and accuracy of the electrochemical DNA sensor through the application of CRISPR Cas systems. Biosens. Bioelectron. 2020, 155, 112100. [Google Scholar] [CrossRef]
- Choi, J.H.; Lim, J.; Shin, M.; Paek, S.H.; Choi, J.W. CRISPR-Cas12a-Based Nucleic Acid Amplification-Free DNA Biosensor via Au Nanoparticle-Assisted Metal-Enhanced Fluorescence and Colorimetric Analysis. Nano Lett. 2021, 21, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.V.; Safenkova, I.V.; Zherdev, A.V.; Dzantiev, B.B. DIRECT2: A novel platform for a CRISPR-Cas12-based assay comprising universal DNA-IgG probe and a direct lateral flow test. Biosens. Bioelectron. 2022, 208, 114227. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Ke, Y.; Maboyi, N.; Zhi, X.; Yan, S.; Li, F.; Zhao, B.; Jia, X.; Song, S.; Ding, X. CRISPR/Cas12a Powered DNA Framework-Supported Electrochemical Biosensing Platform for Ultrasensitive Nucleic Acid Analysis. Small Methods 2021, 5, e2100935. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Shi, Y.; Peng, F.; Zhou, M.; Yin, Y.; Tan, Y.; Chen, M.; Yin, X.; Ke, G.; Zhang, X.B. Exploring the Trans-Cleavage Activity of CRISPR/Cas12a on Gold Nanoparticles for Stable and Sensitive Biosensing. Anal. Chem. 2021, 93, 4967–4974. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Safenkova, I.V.; Biketov, S.F.; Zherdev, A.V.; Dzantiev, B.B. Engineering of DNA Structures Attached to Magnetic Particles for Effective Trans- and Cis-Cleavage in Cas12-Based Biosensors. Int. J. Mol. Sci. 2023, 24, 4484. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.V.; Safenkova, I.V.; Drenova, N.V.; Zherdev, A.V.; Dzantiev, B.B. Development of lateral flow assay combined with recombinase polymerase amplification for highly sensitive detection of Dickeya solani. Mol. Cell. Probes 2020, 53, 101622. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.V.; Safenkova, I.V.; Drenova, N.V.; Zherdev, A.V.; Dzantiev, B.B. Comparison of Biosensing Methods Based on Different Isothermal Amplification Strategies: A Case Study with Erwinia amylovora. Biosensors 2022, 12, 1174. [Google Scholar] [CrossRef]
- Panferov, V.G.; Safenkova, I.V.; Zherdev, A.V.; Dzantiev, B.B. The steadfast Au@Pt soldier: Peroxide-tolerant nanozyme for signal enhancement in lateral flow immunoassay of peroxidase-containing samples. Talanta 2021, 225, 121961. [Google Scholar] [CrossRef]
- Panferov, V.G.; Safenkova, I.V.; Zherdev, A.V.; Dzantiev, B.B. Urchin peroxidase-mimicking Au@Pt nanoparticles as a label in lateral flow immunoassay: Impact of nanoparticle composition on detection limit of Clavibacter michiganensis. Microchim. Acta 2020, 187, 268. [Google Scholar] [CrossRef]
- Li, I.; Kwon, S.-J.; Sorci, M.; Heeger, P.S.; Dordick, J.S. CRISPR-Cas12a has both cis- and trans-cleavage activities on single-stranded DNA. Cell Res. 2018, 28, 491–493. [Google Scholar] [CrossRef] [PubMed]
- Le, Q.; Nguyen, V.; Park, S. Recent advances in the engineering and application of streptavidin-like molecules. Appl. Microbiol. Biotechnol. 2019, 103, 7355–7365. [Google Scholar] [CrossRef]
- Zhao, X.; Li, S.; Liu, G.; Wang, Z.; Yang, Z.; Zhang, Q.; Liang, M.; Liu, J.; Li, Z.; Tong, Y. A versatile biosensing platform coupling CRISPR–Cas12a and aptamers for detection of diverse analytes. Sci. Bull. 2021, 66, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Chi, Q.; Wang, G.; Jiang, J. The persistence length and length per base of single-stranded DNA obtained from fluorescence correlation spectroscopy measurements using mean field theory. Phys. A Stat. Mech. Its Appl. 2013, 392, 1072–1079. [Google Scholar] [CrossRef]
- Roth, E.; Glick Azaria, A.; Girshevitz, O.; Bitler, A.; Garini, Y. Measuring the Conformation and Persistence Length of Single-Stranded DNA Using a DNA Origami Structure. Nano Lett. 2018, 18, 6703–6709. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.C.; Rasnik, I.; Cheng, W.; Lohman, T.M.; Ha, T. Probing single-stranded DNA conformational flexibility using fluorescence spectroscopy. Biophys. J. 2004, 86, 2530–2537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntosh, D.B.; Duggan, G.; Gouil, Q.; Saleh, O.A. Sequence-dependent elasticity and electrostatics of single-stranded DNA: Signatures of base-stacking. Biophys. J. 2014, 106, 659–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, G.; Giri, D.; Kumar, S. Stretching of a single-stranded DNA: Evidence for structural transition. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 2009, 79 Pt 1, 031930. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Choi, J.; Han, H.K.; Park, S.; Park, S.Y.; Park, C.; Bark, C.; Lee, T.; Min, J. Fabrication of ultrasensitive electrochemical biosensor for dengue fever viral RNA Based on CRISPR/Cpf1 reaction. Sens. Actuators B Chem. 2021, 326, 128677. [Google Scholar] [CrossRef]
- Zhang, D.; Yan, Y.; Que, H.; Yang, T.; Cheng, X.; Ding, S.; Zhang, X.; Cheng, W. CRISPR/Cas12a-Mediated Interfacial Cleaving of Hairpin DNA Reporter for Electrochemical Nucleic Acid Sensing. ACS Sens. 2020, 5, 557–562. [Google Scholar] [CrossRef]
- Wang, S.Y.; Du, Y.C.; Wang, D.X.; Ma, J.Y.; Tang, A.N.; Kong, D.M. Signal amplification and output of CRISPR/Cas-based biosensing systems: A review. Anal. Chim. Acta 2021, 15, 338882. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.X.; Wang, Y.X.; Wang, J.; Ma, J.Y.; Liu, B.; Tang, A.N.; Kong, D.M. MnO2 nanosheets as a carrier and accelerator for improved live-cell biosensing application of CRISPR/Cas12a. Chem. Sci. 2022, 13, 4364–4371. [Google Scholar] [CrossRef]
- Sha, Y.; Huang, R.; Huang, M.; Yu, H.; Shan, Y.; Hu, J.; Xing, D. Cascade CRISPR/cas enables amplification-free microRNA sensing with fM-sensitivity and single-base-specificity. Chem. Commun. 2021, 57, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Frens, G. Controlled Nucleation for the Regulation of the Particle Size in Monodisperse Gold Suspensions. Nat. Phys. Sci. 1973, 241, 20. [Google Scholar] [CrossRef]
- Cock, P.J.A.; Antao, T.; Chang, J.T.; Chapman, B.A.; Cox, C.J.; Dalke, A.; Friedberg, I.; Hamelryck, T.; Kauff, F.; Wilczynski, B.; et al. Biopython: Freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 2009, 25, 1422–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanov, A.V.; Safenkova, I.V.; Zherdev, A.V.; Wan, Y.; Dzantiev, B.B. Comparison of Single-Stranded DNA Probes Conjugated with Magnetic Particles for Trans-Cleavage in Cas12a-Based Biosensors. Biosensors 2023, 13, 700. https://doi.org/10.3390/bios13070700
Ivanov AV, Safenkova IV, Zherdev AV, Wan Y, Dzantiev BB. Comparison of Single-Stranded DNA Probes Conjugated with Magnetic Particles for Trans-Cleavage in Cas12a-Based Biosensors. Biosensors. 2023; 13(7):700. https://doi.org/10.3390/bios13070700
Chicago/Turabian StyleIvanov, Aleksandr V., Irina V. Safenkova, Anatoly V. Zherdev, Yi Wan, and Boris B. Dzantiev. 2023. "Comparison of Single-Stranded DNA Probes Conjugated with Magnetic Particles for Trans-Cleavage in Cas12a-Based Biosensors" Biosensors 13, no. 7: 700. https://doi.org/10.3390/bios13070700
APA StyleIvanov, A. V., Safenkova, I. V., Zherdev, A. V., Wan, Y., & Dzantiev, B. B. (2023). Comparison of Single-Stranded DNA Probes Conjugated with Magnetic Particles for Trans-Cleavage in Cas12a-Based Biosensors. Biosensors, 13(7), 700. https://doi.org/10.3390/bios13070700