A Careful Insight into DDI-Type Receptor Layers on the Way to Improvement of Click-Biology-Based Immunosensors



{kind=link}
{kind=link}
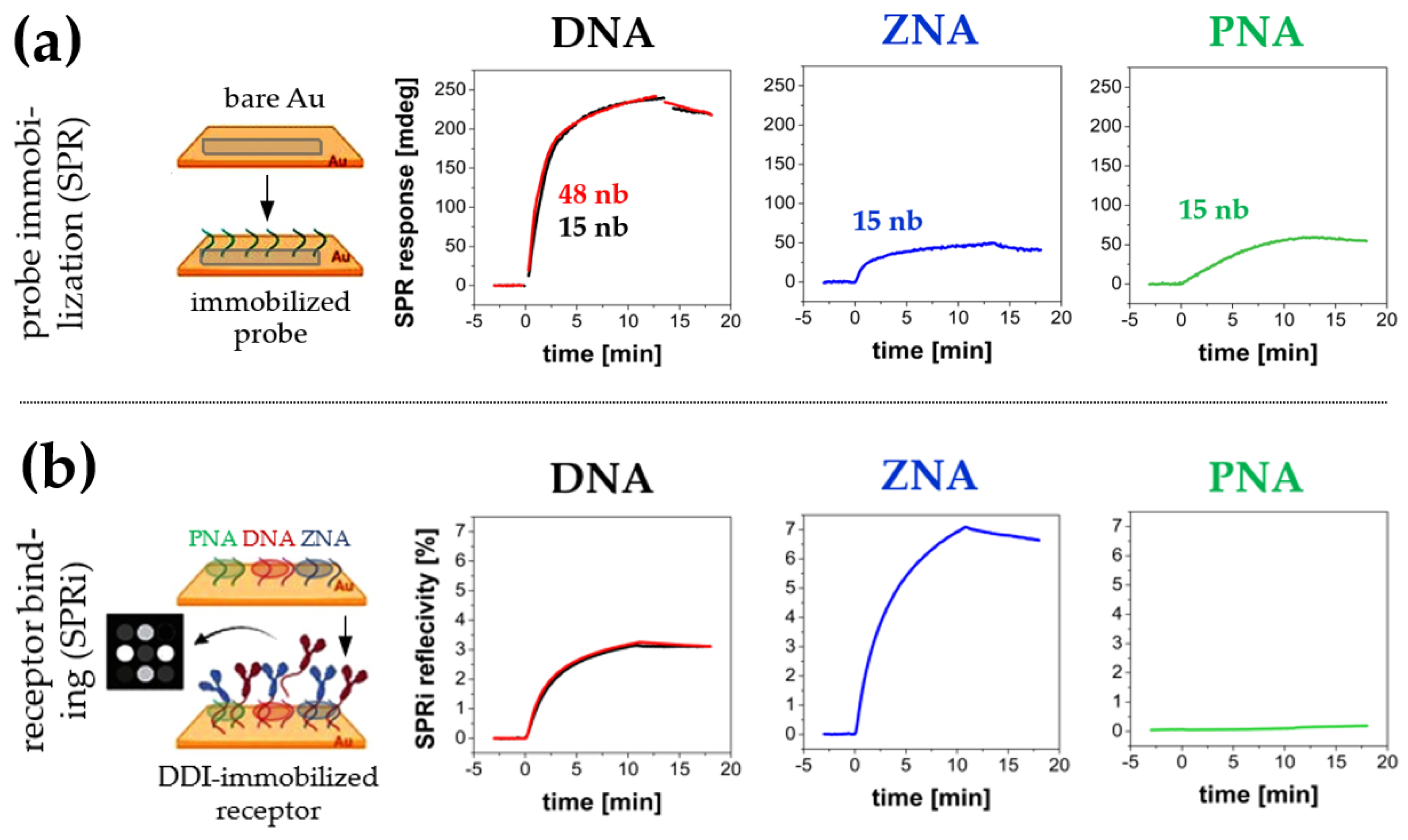{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
- -
- ligand strand (48 nt.) 5′-NH2-C6-ATC AGT ACT TGT CAA CAC GAG CAG CCC GTA TAT TCT CCT ACA GCA CTA-3′
- -
- DNA probe (long) (48 nt.) 5′-SH-C6-TAG TGC TGT AGG AGA ATA TAC GGG CTG CTC GTG TTG ACA AGT ACT GAT-3′
- -
- DNA probe (15 nt.) 5′-SH-C6-TAG TGC TGT AGG AGA-3′
- -
- PNA probe (15 nt.) 5′-SH-C6-TAG TGC TGT AGG AGA-3′
- -
- ZNA® probe (15 nt.) 5′-SH-C6-TAG TGC TGT AGG AGA-(spermine)3-3′
2.2. Surface Preparation of SPR Gold Slides and Ex Situ Immobilization of Oligonucleotide Probes
2.3. Conjugation of Receptor Proteins with ssDNA Anchors and Conjugates Purification
2.4. Ex Situ Immobilization of Protein Conjugates by DNA-Directed Immobilization (DDI)
2.5. Multi-Parametric SPR Measurements and Kinetic Analysis
2.6. Surface Plasmon Resonance Imaging Measurements
2.7. Surface ζ-Potential Measurements of Oligonucleotide Receptor Layers
3. Results and Discussion
3.1. Purification of DNA-Protein Conjugates and SPR-Based Quality Control for DDI
3.2. Development of the Oligonucleotide Probe Layer Composition for DDI
3.2.1. Selection of the Oligonucleotide Anchor Type
3.2.2. Selection of Non-Receptor Components
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aparna, G.M.; Tetala, K.K.R. Recent Progress in Development and Application of DNA, Protein, Peptide, Glycan, Antibody, and Aptamer Microarrays. Biomolecules 2023, 13, 602. [Google Scholar] [CrossRef]
- Li, S.; Song, G.; Bai, Y.; Song, N.; Zhao, J.; Liu, J.; Hu, C. Applications of Protein Microarrays in Biomarker Discovery for Autoimmune Diseases. Front. Immunol. 2021, 12, 645632. [Google Scholar] [CrossRef]
- Walter, J.; Eludin, Z.; Drabovich, A.P. Redefining Serological Diagnostics with Immunoaffinity Proteomics. Clin. Proteom. 2023, 20, 42. [Google Scholar] [CrossRef]
- Watson, E.E.; Winssinger, N. Synthesis of Protein-Oligonucleotide Conjugates. Biomolecules 2022, 12, 1523. [Google Scholar] [CrossRef]
- Brambilla, D.; Sola, L.; Chiari, M. Advantageous Antibody Microarray Fabrication through DNA-Directed Immobilization: A Step toward Use of Extracellular Vesicles in Diagnostics. Talanta 2021, 222, 121542. [Google Scholar] [CrossRef]
- Langer, A.; Hampel, P.A.; Kaiser, W.; Knezevic, J.; Welte, T.; Villa, V.; Maruyama, M.; Svejda, M.; Jähner, S.; Fischer, F.; et al. Protein Analysis by Time-Resolved Measurements with an Electro-Switchable DNA Chip. Nat. Commun. 2013, 4, 2099. [Google Scholar] [CrossRef]
- Cléry, A.; Sohier, T.J.M.; Welte, T.; Langer, A.; Allain, F.H.T. SwitchSENSE: A New Technology to Study Protein-RNA Interactions. Methods 2017, 118–119, 137–145. [Google Scholar] [CrossRef]
- Washburn, A.L.; Gomez, J.; Bailey, R.C. DNA-Encoding to Improve Performance and Allow Parallel Evaluation of the Binding Characteristics of Multiple Antibodies in a Surface-Bound Immunoassay Format. Anal. Chem. 2011, 83, 3572–3580. [Google Scholar] [CrossRef]
- Huang, S.; Wang, W.; Li, J.; Zhang, T.; Liang, Y.; Wang, Q.; Jiang, Z. Multifunctional DNA Mediated Spatially Confined Assembly for Antibody Orientation: Surpassing Sensitivity and Accuracy for Rituximab Detection. Chem. Eng. J. 2021, 419, 129613. [Google Scholar] [CrossRef]
- Ünlü, N.L.; Kanik, F.E.; Seymour, E.; Connor, J.H.; Ünlü, M.S. DNA-Directed Antibody Immobilization. In Biosensors and Biodetection; Humana Press: New York, NY, USA, 2017; Volume 1571. [Google Scholar]
- Seymour, E.; Daaboul, G.G.; Zhang, X.; Scherr, S.M.; Ünlü, N.L.; Connor, J.H.; Ünlü, M.S. DNA-Directed Antibody Immobilization for Enhanced Detection of Single Viral Pathogens. Anal. Chem. 2015, 87, 10505–10512. [Google Scholar] [CrossRef]
- Bernardinelli, G.; Oloketuyi, S.; Werner, S.F.; Mazzega, E.; Högberg, B.; de Marco, A. A Compact Nanobody-DNAzyme Conjugate Enables Antigen Detection and Signal Amplification. New Biotechnol. 2020, 56, 1–8. [Google Scholar] [CrossRef]
- Al-Amin, R.A.; Muthelo, P.M.; Abdurakhmanov, E.; Vincke, C.; Amin, S.P.; Muyldermans, S.; Danielson, U.H.; Landegren, U. Sensitive Protein Detection Using Site-Specifically Oligonucleotide-Conjugated Nanobodies. Anal. Chem. 2022, 94, 10054–10061. [Google Scholar] [CrossRef]
- Ladd, J.; Taylor, A.D.; Piliarik, M.; Homola, J.; Jiang, S. Hybrid Surface Platform for the Simultaneous Detection of Proteins and DNAs Using a Surface Plasmon Resonance Imaging Sensor. Anal. Chem. 2008, 80, 4231–4236. [Google Scholar] [CrossRef]
- Ambrosetti, E.; Paoletti, P.; Bosco, A.; Parisse, P.; Scaini, D.; Tagliabue, E.; De Marco, A.; Casalis, L. Quantification of Circulating Cancer Biomarkers via Sensitive Topographic Measurements on Single Binder Nanoarrays. ACS Omega 2017, 2, 2618–2629. [Google Scholar] [CrossRef] [PubMed]
- Leroy, L.; Bombera, R.; Engel, E.; Calemczuk, R.; Laplatine, L.; Baganizi, D.-D.R.; Marche, P.N.; Roupioz, Y.; Livache, T. Photothermal Effect for Localized Desorption of Primary Lymphocytes Arrayed on an Antibody/DNA-Based Biochip. Lab Chip 2014, 14, 1987–1990. [Google Scholar] [CrossRef] [PubMed]
- Dovgan, I.; Koniev, O.; Kolodych, S.; Wagner, A. Antibody-Oligonucleotide Conjugates as Therapeutic, Imaging, and Detection Agents. Bioconjug. Chem. 2019, 30, 2483–2501. [Google Scholar] [CrossRef]
- Yan, X.; Zhang, H.; Wang, Z.; Peng, H.; Tao, J.; Li, X.-F.; Chris Le, X. Quantitative Synthesis of Protein-DNA Conjugates with 1:1 Stoichiometry. Chem. Commun. 2018, 54, 7491–7494. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.; Lautner, G.; Gyurcsányi, R.E. Reliable Microspotting Methodology for Peptide-Nucleic Acid Layers with High Hybridization Efficiency on Gold SPR Imaging Chips. Anal. Methods 2015, 7, 6077–6082. [Google Scholar] [CrossRef]
- Nobbmann, U. Measure Surface Zeta Potential. Available online: https://www.materials-talks.com/surface-zeta-potential-what-it-is-and-how-to-measure-it/ (accessed on 6 January 2015).
- Corbett, J.C.W.; McNeil-Watson, F.; Jack, R.O.; Howarth, M. Measuring Surface Zeta Potential Using Phase Analysis Light Scattering in a Simple Dip Cell Arrangement. Colloids Surf. A Physicochem. Eng. Asp. 2012, 396, 169–176. [Google Scholar] [CrossRef]
- Rodríguez, D.F.; Moglie, Y.; Ramírez-Sarmiento, C.A.; Singh, S.K.; Dua, K.; Zacconi, F.C. Bio-Click Chemistry: A Bridge between Biocatalysis and Click Chemistry. RSC Adv. 2022, 12, 1932–1949. [Google Scholar] [CrossRef] [PubMed]
- El-Sagheer, A.H.; Brown, T. Click Nucleic Acid Ligation: Applications in Biology and Nanotechnology. Acc. Chem. Res. 2012, 45, 1258–1267. [Google Scholar] [CrossRef]
- Wiener, J.; Kokotek, D.; Rosowski, S.; Lickert, H.; Meier, M. Preparation of Single- and Double-Oligonucleotide Antibody Conjugates and Their Application for Protein Analytics. Sci. Rep. 2020, 10, 1457. [Google Scholar] [CrossRef] [PubMed]
- Heerwig, A.; Kick, A.; Sommerfeld, P.; Eimermacher, S.; Hartung, F.; Laube, M.; Fischer, D.; Pietzsch, H.-J.; Pietzsch, J.; Löser, R.; et al. The Impact of Nε-Acryloyllysine Piperazides on the Conformational Dynamics of Transglutaminase 2. Int. J. Mol. Sci. 2023, 24, 1650. [Google Scholar] [CrossRef]
- Frato, K.E.; Schleif, R.F. A DNA-Assisted Binding Assay for Weak Protein-Protein Interactions. J. Mol. Biol. 2009, 394, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Cain, P.; Anguiano, V.; Shih, J.J.; Chai, Q.; Feng, Y. Impact of IgG Subclass on Molecular Properties of Monoclonal Antibodies. MAbs 2021, 13, 1993768. [Google Scholar] [CrossRef]
- Marquart, A.; Kuncova-Kallio, J.; Albers, M.; Bombera, R.; Stahlberg, R. Handbook of Multi-Parametric Surface Plasmon Resonance for Molecular Interaction Analysis—Theory and Practice, 1st ed.; The Royal Society of Chemistry: Tampere, Finland, 2019. [Google Scholar]
- Gong, P.; Levicky, R. DNA Surface Hybridization Regimes. Proc. Natl. Acad. Sci. USA 2008, 105, 5301–5306. [Google Scholar] [CrossRef] [PubMed]
- Bielec, K.; Kowalski, A.; Bubak, G.; Witkowska Nery, E.; Hołyst, R. Ion Complexation Explains Orders of Magnitude Changes in the Equilibrium Constant of Biochemical Reactions in Buffers Crowded by Nonionic Compounds. J. Phys. Chem. Lett. 2022, 13, 112–117. [Google Scholar] [CrossRef]
- Nothisen, M.; Perche-Létuvée, P.; Behr, J.-P.; Remy, J.-S.; Kotera, M. Cationic Oligospermine-Oligonucleotide Conjugates Provide Carrier-Free Splice Switching in Monolayer Cells and Spheroids. Mol. Ther. Nucleic Acids 2018, 13, 483–492. [Google Scholar] [CrossRef]
- Zhu, B.; Travas-Sejdic, J. PNA versus DNA in Electrochemical Gene Sensing Based on Conducting Polymers: Study of Charge and Surface Blocking Effects on the Sensor Signal. Analyst 2018, 143, 687–694. [Google Scholar] [CrossRef]
- Briones, C.; Mateo-Marti, E.; Gómez-Navarro, C.; Parro, V.; Román, E.; Martín-Gago, J.A. Ordered Self-Assembled Monolayers of Peptide Nucleic Acids with DNA Recognition Capability. Phys. Rev. Lett. 2004, 93, 208103. [Google Scholar] [CrossRef]
- Del Bene, A.; D’Aniello, A.; Tomassi, S.; Merlino, F.; Mazzarella, V.; Russo, R.; Chambery, A.; Cosconati, S.; Di Maro, S.; Messere, A. Ultrasound-Assisted Peptide Nucleic Acids Synthesis (US-PNAS). Ultrason. Sonochem. 2023, 95, 106360. [Google Scholar] [CrossRef] [PubMed]
- Drozd, M.; Pietrzak, M.D.; Malinowska, E. SPRi-Based Biosensing Platforms for Detection of Specific DNA Sequences Using Thiolate and Dithiocarbamate Assemblies. Front. Chem. 2018, 6, 173. [Google Scholar] [CrossRef]
- Sarcina, L.; Torsi, L.; Picca, R.A.; Manoli, K.; Macchia, E. Assessment of Gold Bio-Functionalization for Wide-Interface Biosensing Platforms. Sensors 2020, 20, 3678. [Google Scholar] [CrossRef]
- Li, Z.; Ruiz, V.G.; Kanduč, M.; Dzubiella, J. Ion-Specific Adsorption on Bare Gold (Au) Nanoparticles in Aqueous Solutions: Double-Layer Structure and Surface Potentials. Langmuir 2020, 36, 13457–13468. [Google Scholar] [CrossRef] [PubMed]
- Saito, N.; Matsuda, T. Protein Adsorption on Self-Assembled Monolayers with Water-Soluble Non-Ionic Oligomers Using Quartz-Crystal Microbalance. Mater. Sci. Eng. C 1998, 6, 261–266. [Google Scholar] [CrossRef]
- Szymczyk, A.; Soliwodzka, K.; Moskal, M.; Różanowski, K.; Ziółkowski, R. Further Insight into the Possible Influence of Electrode Blocking Agents on the Stem-Loop Based Electrochemical DNA Sensor Parameters. Sens. Actuators B Chem. 2022, 354, 131086. [Google Scholar] [CrossRef]
- Havens, A.; El-Shaer, E.; Garcia, L.; Godino, J.L.; Thompson, R.S. Protein Adsorption on Mixed Self-Assembled Monolayers: Influence of Chain Length and Terminal Group. Langmuir 2023, 39, 16712–16720. [Google Scholar] [CrossRef]
- Simon, L.; Gyurcsányi, R.E. Multiplexed Assessment of the Surface Density of DNA Probes on DNA Microarrays by Surface Plasmon Resonance Imaging. Anal. Chim. Acta 2019, 1047, 131–138. [Google Scholar] [CrossRef]
- Voigt, N.V.; Tørring, T.; Rotaru, A.; Jacobsen, M.F.; Ravnsbæk, J.B.; Subramani, R.; Mamdouh, W.; Kjems, J.; Mokhir, A.; Besenbacher, F.; et al. Single-Molecule Chemical Reactions on DNA Origami. Nat. Nanotechnol. 2010, 5, 200–203. [Google Scholar] [CrossRef]
- Gothelf, K.V. Chemical Modifications and Reactions in DNA Nanostructures. MRS Bull. 2017, 42, 897–903. [Google Scholar] [CrossRef]
- Verdolino, V.; Cammi, R.; Munk, B.H.; Schlegel, H.B. Calculation of PKa Values of Nucleobases and the Guanine Oxidation Products Guanidinohydantoin and Spiroiminodihydantoin Using Density Functional Theory and a Polarizable Continuum Model. J. Phys. Chem. B 2008, 112, 16860–16873. [Google Scholar] [CrossRef] [PubMed]
- Bryantsev, V.S.; Diallo, M.S.; Goddard III, W.A. PKa Calculations of Aliphatic Amines, Diamines, and Aminoamides via Density Functional Theory with a Poisson-Boltzmann Continuum Solvent Model. J. Phys. Chem. A 2007, 111, 4422–4430. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karoń, S.; Drozd, M.; Malinowska, E. A Careful Insight into DDI-Type Receptor Layers on the Way to Improvement of Click-Biology-Based Immunosensors. Biosensors 2024, 14, 136. https://doi.org/10.3390/bios14030136
Karoń S, Drozd M, Malinowska E. A Careful Insight into DDI-Type Receptor Layers on the Way to Improvement of Click-Biology-Based Immunosensors. Biosensors. 2024; 14(3):136. https://doi.org/10.3390/bios14030136
Chicago/Turabian StyleKaroń, Sylwia, Marcin Drozd, and Elżbieta Malinowska. 2024. "A Careful Insight into DDI-Type Receptor Layers on the Way to Improvement of Click-Biology-Based Immunosensors" Biosensors 14, no. 3: 136. https://doi.org/10.3390/bios14030136
APA StyleKaroń, S., Drozd, M., & Malinowska, E. (2024). A Careful Insight into DDI-Type Receptor Layers on the Way to Improvement of Click-Biology-Based Immunosensors. Biosensors, 14(3), 136. https://doi.org/10.3390/bios14030136