Integration of Nanomaterials and Bioluminescence Resonance Energy Transfer Techniques for Sensing Biomolecules


Abstract
:1. Introduction
2. Bioluminescent Proteins
2.1. Aequorin
2.2. Bacterial Luciferase
2.3. Firefly Luciferase
2.4. Renilla Luciferase
2.5. Gaussia Luciferase
2.6. Vargula Luciferase
2.7. Metridia Luciferase
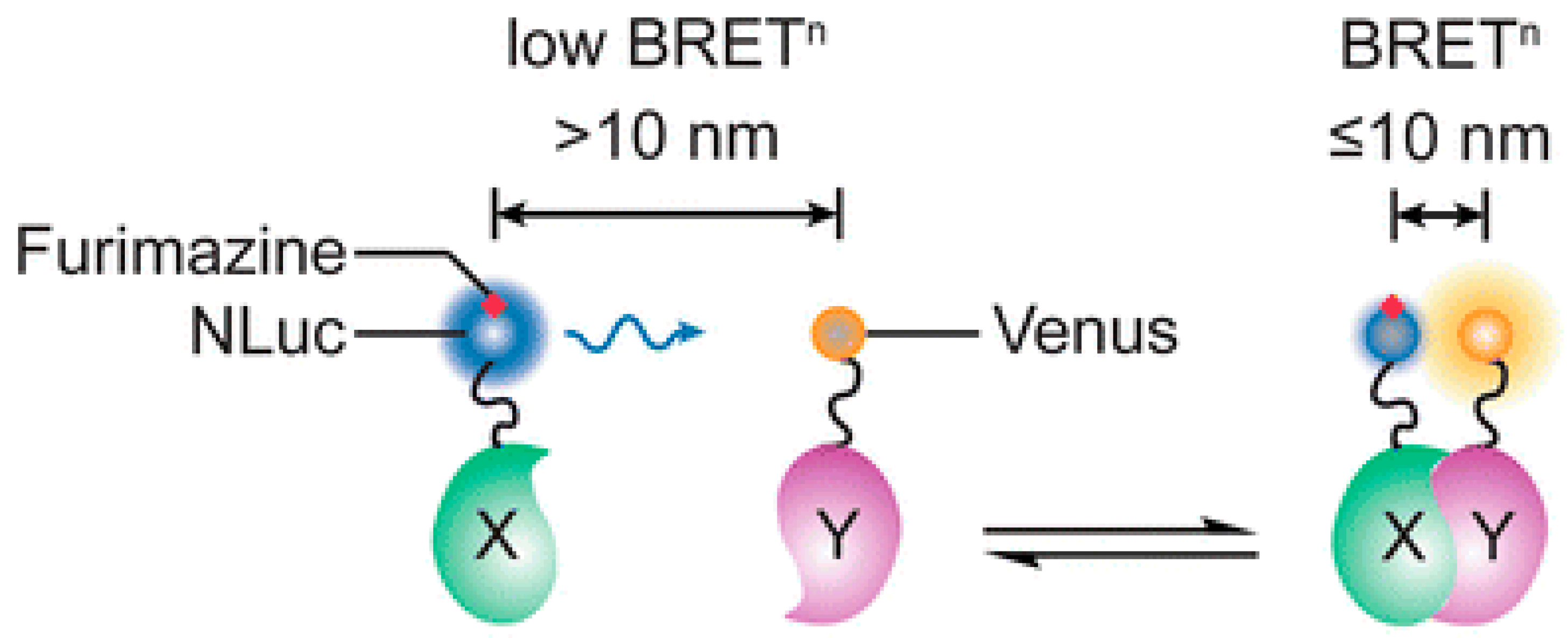
2.8. Nano Luciferase
3. QD-Based Biosensors by Applying Bioluminescent Resonance Energy Transfer Techniques
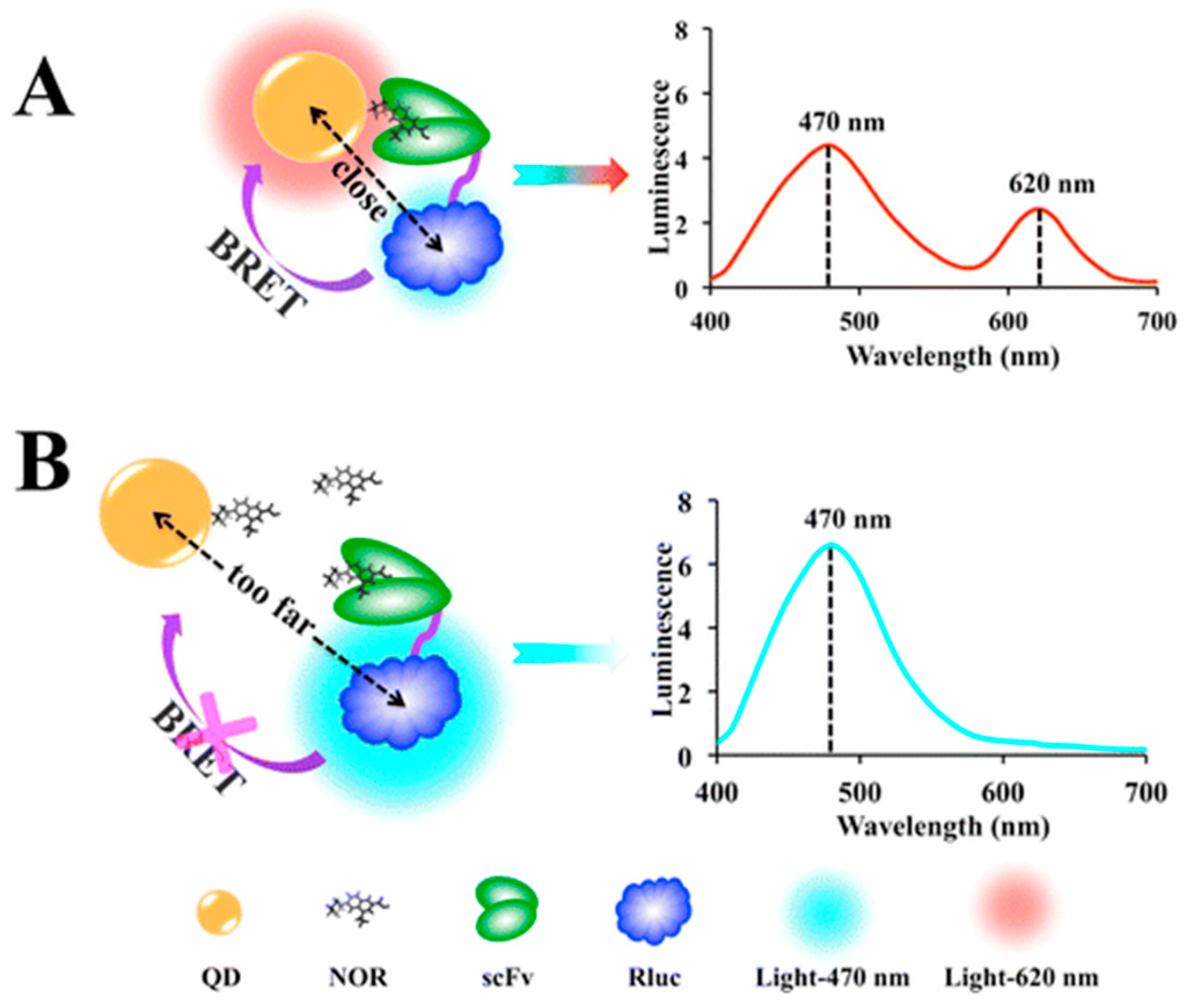
3.1. QD-Based BRET Sensor for Detecting Biomolecules
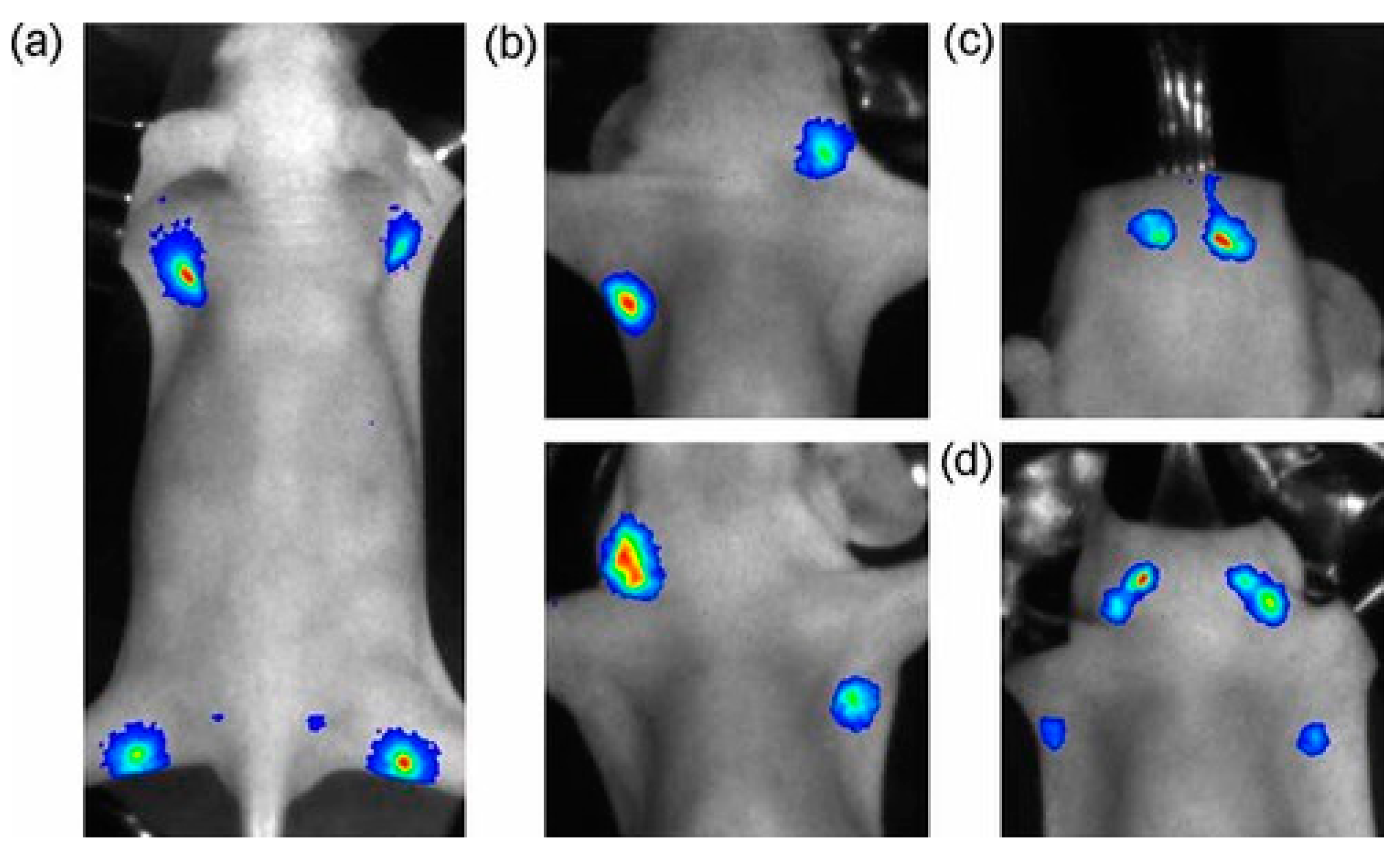
3.2. QD-Based BRET Sensor Used in Bioimaging In Vivo
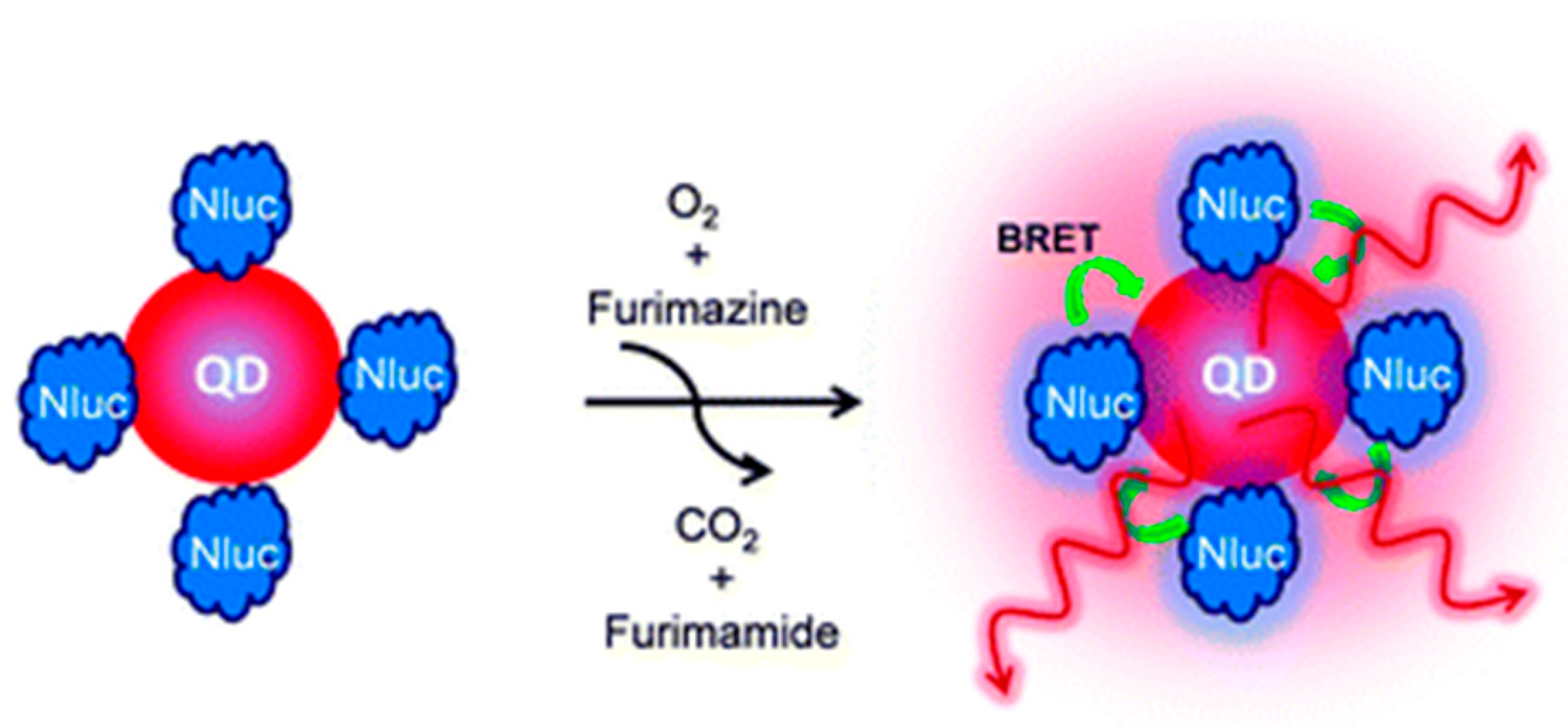
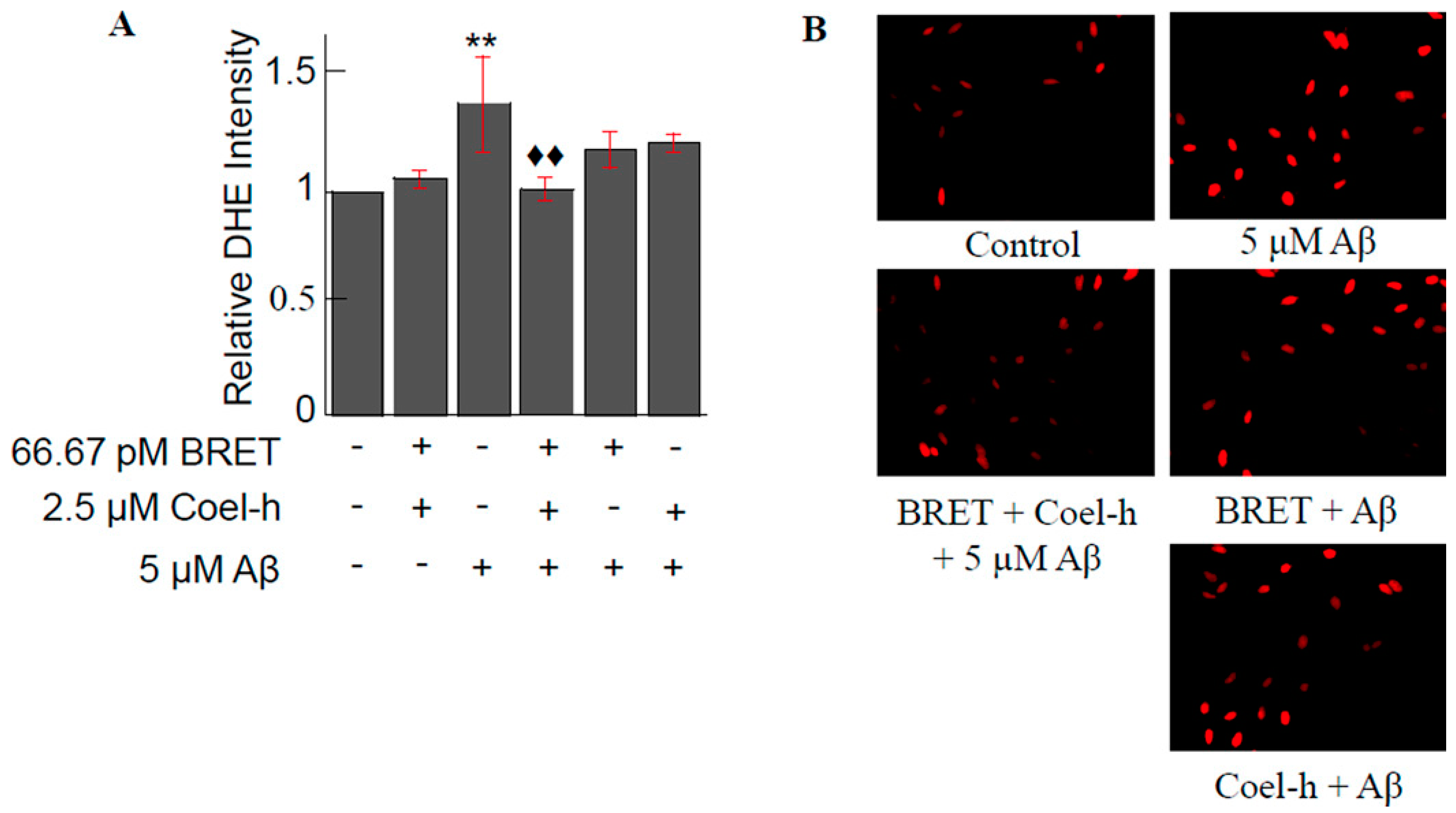
3.3. Other QDs-Based Applications
4. Biocompatibility
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Morin, J.G.; Hastings, J.W. Energy transfer in a bioluminescent system. J. Cell. Physiol. 1971, 77, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Ward, W.W.; Cormier, M.J. An energy transfer protein in coelenterate bioluminescence. Characterization of the Renilla green-fluorescent protein. J. Biol. Chem. 1979, 254, 781–788. [Google Scholar] [PubMed]
- Lorenz, W.W.; McCann, R.O.; Longiaru, M.; Cormier, M.J. Isolation and expression of a cDNA encoding Renilla reniformis luciferase. Proc. Natl. Acad. Sci. USA 1991, 88, 4438–4442. [Google Scholar] [CrossRef] [PubMed]
- Förster, T. Zwischenmolekulare Energiewanderung und Fluoreszenz. Ann. Phys. 1948, 437, 55–75. [Google Scholar] [CrossRef]
- Li, H.; Luo, Y.; Sun, X. Fluorescence resonance energy transfer dye-labeled probe for fluorescence-enhanced DNA detection: An effective strategy to greatly improve discrimination ability toward single-base mismatch. Biosens. Bioelectron. 2011, 27, 167–171. [Google Scholar] [CrossRef]
- Prada, M.P.; Syed, A.U.; Buonarati, O.R.; Reddy, G.R.; Nystoriak, M.A.; Ghosh, D.; Simo, S.; Sato, D.; Sasse, K.C.; Ward, S.M.; et al. A Gs-coupled purinergic receptor boosts Ca(2+) influx and vascular contractility during diabetic hyperglycemia. Elife 2019, 8, e42214. [Google Scholar] [CrossRef]
- Greotti, E.; Capitanio, P.; Wong, A.; Pozzan, T.; Pizzo, P.; Pendin, D. Familial Alzheimer’s disease-linked presenilin mutants and intracellular Ca(2+) handling: A single-organelle, FRET-based analysis. Cell Calcium 2019, 79, 44–56. [Google Scholar] [CrossRef]
- Solis, C.; Russell, B. CapZ integrates several signaling pathways in response to mechanical stiffness. J. Gen. Physiol. 2019, 151. [Google Scholar] [CrossRef]
- Zhang, L.; Bouadjel, K.; Manoury, B.; Vandecasteele, G.; Fischmeister, R.; Leblais, V. Cyclic nucleotide signalling compartmentation by phosphodiesterases in cultured vascular smooth muscle cells. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Manoharan, G.B.; Kopra, K.; Eskonen, V.; Härmä, H.; Abankwa, D. High-throughput amenable fluorescence-assays to screen for calmodulin-inhibitors. Anal. Biochem. 2019, 18, 31185–31190. [Google Scholar] [CrossRef]
- Pfleger, K.D.G.; Eidne, K.A. Illuminating insights into protein-protein interactions using bioluminescence resonance energy transfer (BRET). Nat. Methods 2006, 3, 165–174. [Google Scholar] [CrossRef] [PubMed]
- del Rosal, B.; Benayas, A. Strategies to Overcome Autofluorescence in Nanoprobe-Driven In Vivo Fluorescence Imaging. Small Methods 2018, 2, 1800075. [Google Scholar] [CrossRef]
- So, M.-K.; Xu, C.; Loening, A.M.; Gambhir, S.S.; Rao, J. Self-illuminating quantum dot conjugates for in vivo imaging. Nat. Biotechnol. 2006, 24, 339. [Google Scholar] [CrossRef] [PubMed]
- Troy, T.; Jekic-McMullen, D.; Sambucetti, L.; Rice, B. Quantitative Comparison of the Sensitivity of Detection of Fluorescent and Bioluminescent Reporters in Animal Models. Mol. Imaging 2004, 3, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, L.; Qian, H.; Dong, C.; Ren, J. A Resonance Energy Transfer between Chemiluminescent Donors and Luminescent Quantum-Dots as Acceptors (CRET). Angew. Chem. Int. Ed. 2006, 45, 5140–5143. [Google Scholar] [CrossRef]
- Zhao, S.; Huang, Y.; Shi, M.; Liu, R.; Liu, Y.-M. Chemiluminescence Resonance Energy Transfer-Based Detection for Microchip Electrophoresis. Anal. Chem. 2010, 82, 2036–2041. [Google Scholar] [CrossRef] [PubMed]
- Freeman, R.; Liu, X.Q.; Willner, I. Chemiluminescent and Chemiluminescence Resonance Energy Transfer (CRET) Detection of DNA, Metal Ions, and Aptamer-Substrate Complexes Using Hemin/G-Quadruplexes and CdSe/ZnS Quantum Dots. J. Am. Chem. Soc. 2011, 133, 11597–11604. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Q.; Freeman, R.; Golub, E.; Willner, I. Chemiluminescence and Chemiluminescence Resonance Energy Transfer (CRET) Aptamer Sensors Using Catalytic Hemin/G-Quadruplexes. ACS Nano 2011, 5, 7648–7655. [Google Scholar] [CrossRef] [PubMed]
- Ward, W.W.; Cormier, M.J. Energy Transfer via Protein-Protein Interaction In Renilla Bioluminescence. Photochem. Photobiol. 1978, 27, 389–396. [Google Scholar] [CrossRef]
- Mo, X.-L.; Fu, H. BRET: NanoLuc-Based Bioluminescence Resonance Energy Transfer Platform to Monitor Protein-Protein Interactions in Live Cells. Methods Mol. Biol. 2016, 1439, 263–271. [Google Scholar]
- Schaufele, F.; Demarco, I.; Day, R.N. FRET Imaging in the Wide-Field Microscope. In Molecular Imaging; Periasamy, A., Day, R.N.B.T.-M.I., Eds.; American Physiological Society: San Diego, CA, USA, 2005; pp. 72–94. [Google Scholar]
- Ansbacher, T.; Srivastava, H.K.; Stein, T.; Baer, R.; Merkx, M.; Shurki, A. Calculation of transition dipole moment in fluorescent proteins—Towards efficient energy transfer. Phys. Chem. Chem. Phys. 2012, 14, 4109–4117. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-Y.; Chen, C.-W.; Yu, H.-P.; Lin, Y.-F.; Lai, P.-S. Bioluminescence resonance energy transfer using luciferase-immobilized quantum dots for self-illuminated photodynamic therapy. Biomaterials 2013, 34, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
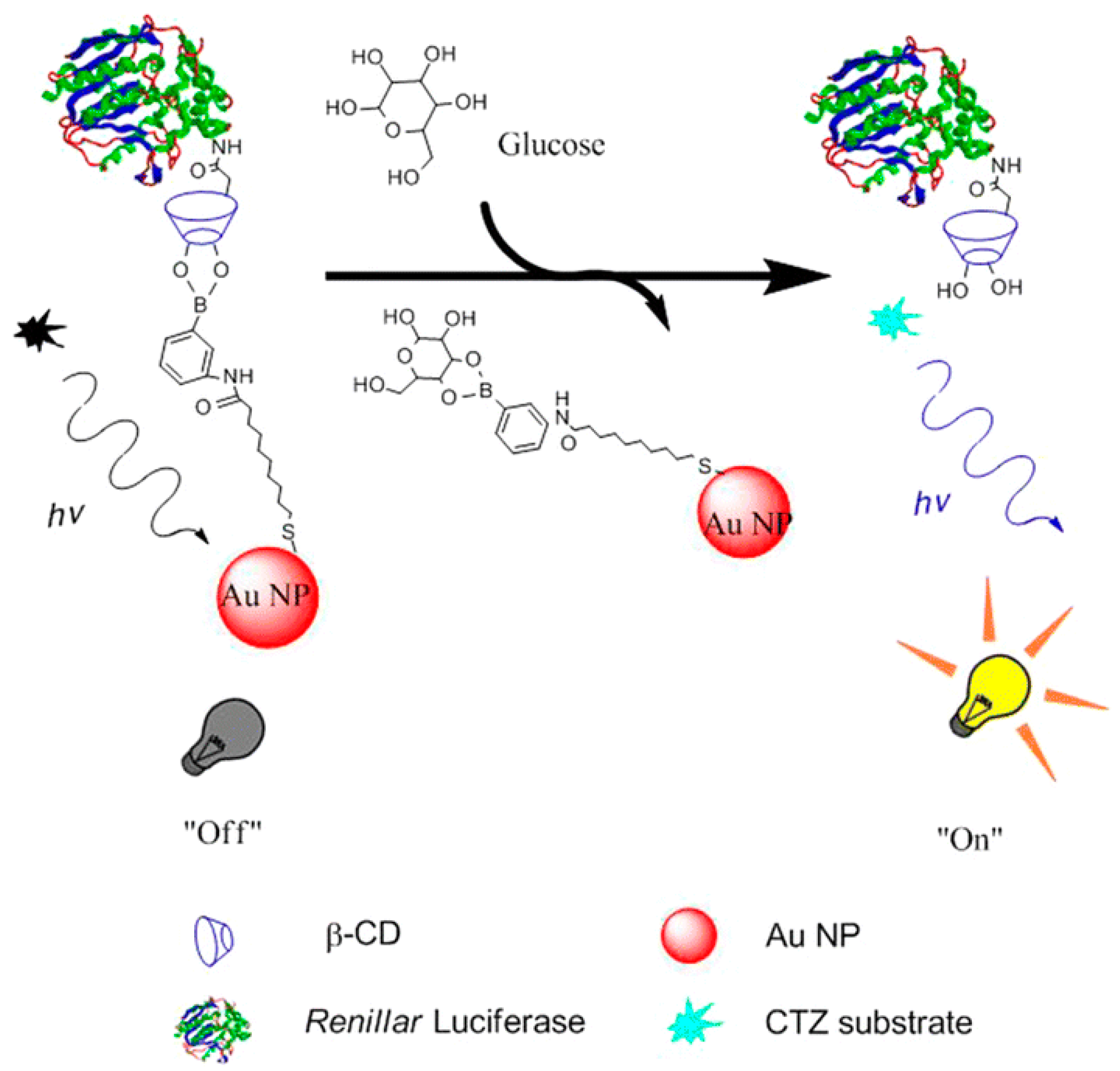
- Chen, L.; Chen, L.; Dotzert, M.; Melling, C.W.J.; Zhang, J. Nanostructured biosensor using bioluminescence quenching technique for glucose detection. J. Nanobiotechnol. 2017, 15, 59. [Google Scholar] [CrossRef]
- Hoshino, H.; Nakajima, Y.; Ohmiya, Y. Luciferase-YFP fusion tag with enhanced emission for single-cell luminescence imaging. Nat. Methods 2007, 4, 637. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, P.; Costoya, J.A. A novel BRET-based genetically encoded biosensor for functional imaging of hypoxia. Biosens. Bioelectron. 2009, 24, 3126–3130. [Google Scholar] [CrossRef]
- Rumyantsev, K.A.; Turoverov, K.K.; Verkhusha, V. V Near-infrared bioluminescent proteins for two-color multimodal imaging. Sci. Rep. 2016, 6, 36588. [Google Scholar] [CrossRef] [PubMed]
- Kojima, R.; Takakura, H.; Ozawa, T.; Tada, Y.; Nagano, T.; Urano, Y. Rational Design and Development of Near-Infrared-Emitting Firefly Luciferins Available In Vivo. Angew. Chem. Int. Ed. 2012, 52, 1175–1179. [Google Scholar] [CrossRef]
- Wu, C.; Mino, K.; Akimoto, H.; Kawabata, M.; Nakamura, K.; Ozaki, M.; Ohmiya, Y. In vivo far-red luminescence imaging of a biomarker based on BRET from Cypridina bioluminescence to an organic dye. Proc. Natl. Acad. Sci. USA 2009, 106, 15599–15603. [Google Scholar] [CrossRef]
- Branchini, B.R.; Ablamsky, D.M.; Rosenberg, J.C. Chemically Modified Firefly Luciferase Is an Efficient Source of Near-Infrared Light. Bioconjugate Chem. 2010, 21, 2023–2030. [Google Scholar] [CrossRef]
- De, A. The New Era of Bioluminescence Resonance Energy Transfer Technology. Curr. Pharm. Biotechnol. 2011, 12, 558–568. [Google Scholar] [CrossRef]
- Xia, Z.; Rao, J. Biosensing and imaging based on bioluminescence resonance energy transfer. Curr. Opin. Biotechnol. 2009, 20, 37–44. [Google Scholar] [CrossRef]
- Mithöfer, A.; Mazars, C. Aequorin-based measurements of intracellular Ca2+-signatures in plant cells. Biol. Proc. Online 2002, 4, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.; Bertoli, A.; Sorgato, M.C.; Moccia, F. Generation and usage of aequorin lentiviral vectors for Ca2+ measurement in sub-cellular compartments of hard-to-transfect cells. Cell Calcium 2016, 59, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Cobbold, P.H.; Cuthbertson, K.S.; Goyns, M.H.; Rice, V. Aequorin measurements of free calcium in single mammalian cells. J. Cell Sci. 1983, 61, 123–136. [Google Scholar] [PubMed]
- Granatiero, V.; Patron, M.; Tosatto, A.; Merli, G.; Rizzuto, R. The use of aequorin and its variants for Ca2+ measurements. Cold Spring Harb. Protoc. 2014, 2014, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Zeinoddini, M.; Khajeh, K.; Hosseinkhani, S.; Saeedinia, A.R.; Robatjazi, S.-M. Stabilisation of Recombinant Aequorin by Polyols: Activity, Thermostability and Limited Proteolysis. Appl. Biochem. Biotechnol. 2013, 170, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Iwano, S.; Sugiyama, M.; Hama, H.; Watakabe, A.; Hasegawa, N.; Kuchimaru, T.; Tanaka, K.Z.; Takahashi, M.; Ishida, Y.; Hata, J.; et al. Single-cell bioluminescence imaging of deep tissue in freely moving animals. Science 2018, 359, 935–939. [Google Scholar] [CrossRef]
- Waidmann, M.S.; Bleichrodt, F.S.; Laslo, T.; Riedel, C.U. Bacterial luciferase reporters: The Swiss army knife of molecular biology. Bioeng. Bugs 2011, 2, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Ke, D.; Tu, S.-C. Activities, Kinetics and Emission Spectra of Bacterial Luciferase-Fluorescent Protein Fusion Enzymes. Photochem. Photobiol. 2011, 87, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Zhang, L.; Song, Y.; Wei, J.; Li, C.; Wang, T.; Wang, Y.; Zhao, T.; Shen, X. Engineering an Enhanced, Thermostable, Monomeric Bacterial Luciferase Gene As a Reporter in Plant Protoplasts. PLoS ONE 2014, 9, e107885. [Google Scholar] [CrossRef]
- Hollis, R.P.; Lagido, C.; Pettitt, J.; Porter, A.J.R.; Killham, K.; Paton, G.I.; Glover, L.A. Toxicity of the bacterial luciferase substrate, n-decyl aldehyde, to Saccharomyces cerevisiae and Caenorhabditis elegans. FEBS Lett. 2001, 506, 140–142. [Google Scholar] [CrossRef]
- Coleman, S.M.; McGregor, A. A bright future for bioluminescent imaging in viral research. Future Virol. 2015, 10, 169–183. [Google Scholar] [CrossRef]
- de Wet, J.R.; Wood, K.V.; DeLuca, M.; Helinski, D.R.; Subramani, S. Firefly luciferase gene: Structure and expression in mammalian cells. Mol. Cell. Biol. 1987, 7, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Lundin, A. Optimization of the Firefly Luciferase Reaction for Analytical Purposes. In Bioluminescence: Fundamentals and Applications in Biotechnology–Volume 2; Thouand, G., Marks, R., Eds.; Advances in Biochemical Engineering/Biotechnology; Springer: Berlin/Heidelberg, Germany, 2014; pp. 31–62. [Google Scholar]
- Gibbons, A.E.; Luker, K.E.; Luker, G.D. Dual Reporter Bioluminescence Imaging with NanoLuc and Firefly Luciferase BT. In Reporter Gene Imaging: Methods and Protocols; Dubey, P., Ed.; Springer: New York, NY, USA, 2018; pp. 41–50. [Google Scholar]
- Branchini, B.R.; Southworth, T.L.; Fontaine, D.M.; Kohrt, D.; Welcome, F.S.; Florentine, C.M.; Henricks, E.R.; DeBartolo, D.B.; Michelini, E.; Cevenini, L.; et al. Red-emitting chimeric firefly luciferase for in vivo imaging in low ATP cellular environments. Anal. Biochem. 2017, 534, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, Q.; Luo, M.; Li, M.; Wang, D.; Wang, Y.; Liu, Q. Immobilization of Firefly Luciferase on PVA-co-PE Nanofibers Membrane as Biosensor for Bioluminescent Detection of ATP. ACS Appl. Mater. Interfaces 2015, 7, 20046–20052. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, D.V.; Ugarova, N.N. Firefly Luciferase-based Fusion Proteins and their Applications in Bioanalysis. Photochem. Photobiol. 2017, 93, 436–447. [Google Scholar] [CrossRef]
- Adams, S.T.; Miller, S.C. Beyond D-luciferin: Expanding the Scope of Bioluminescence Imaging in vivo. Curr. Opin. Chem. Biol. 2014, 21, 112–120. [Google Scholar] [CrossRef]
- Simonyan, H.; Hurr, C.; Young, C.N. A synthetic luciferin improves in vivo bioluminescence imaging of gene expression in cardiovascular brain regions. Physiol. Genom. 2016, 48, 762–770. [Google Scholar] [CrossRef]
- Paulmurugan, R.; Gambhir, S.S. Monitoring Protein−Protein Interactions Using Split Synthetic Renilla Luciferase Protein-Fragment-Assisted Complementation. Anal. Chem. 2003, 75, 1584–1589. [Google Scholar] [CrossRef]
- Bhaumik, S.; Gambhir, S.S. Optical imaging of Renilla luciferase reporter gene expression in living mice. Proc. Natl. Acad. Sci. USA 2002, 99, 377–382. [Google Scholar] [CrossRef]
- Eremeeva, E.V.; Markova, S.V.; Vysotski, E.S. Highly active BRET-reporter based on yellow mutant of Renilla muelleri luciferase. Dokl. Biochem. Biophys. 2013, 450, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, W.W.; Cormier, M.J.; O’Kane, D.J.; Hua, D.; Escher, A.A.; Szalay, A.A. Expression of the Renilla reniformis luciferase gene in mammalian cells. J. Biolumin. Chemilumin. 1996, 11, 31–37. [Google Scholar] [CrossRef]
- Tannous, B.A.; Kim, D.-E.; Fernandez, J.L.; Weissleder, R.; Breakefield, X.O. Codon-Optimized Gaussia Luciferase cDNA for Mammalian Gene Expression in Culture and in Vivo. Mol. Ther. 2005, 11, 435–443. [Google Scholar] [CrossRef]
- Stepanyuk, G.A.; Xu, H.; Wu, C.-K.; Markova, S.V.; Lee, J.; Vysotski, E.S.; Wang, B.-C. Expression, purification and characterization of the secreted luciferase of the copepod Metridia longa from Sf9 insect cells. Protein Expr. Purif. 2008, 61, 142–148. [Google Scholar] [CrossRef]
- Inoue, Y.; Sheng, F.; Kiryu, S.; Watanabe, M.; Ratanakanit, H.; Izawa, K.; Tojo, A.; Ohtomo, K. Gaussia Luciferase for Bioluminescence Tumor Monitoring in Comparison with Firefly Luciferase. Mol. Imaging 2011, 10, 377–385. [Google Scholar] [CrossRef]
- Degeling, M.H.; Bovenberg, M.S.S.; Lewandrowski, G.K.; de Gooijer, M.C.; Vleggeert-Lankamp, C.L.A.; Tannous, M.; Maguire, C.A.; Tannous, B.A. Directed molecular evolution reveals Gaussia luciferase variants with enhanced light output stability. Anal. Chem. 2013, 85, 3006–3012. [Google Scholar] [CrossRef]
- Thompson, E.M.; Nagata, S.; Tsuji, F.I. Cloning and expression of cDNA for the luciferase from the marine ostracod Vargula hilgendorfii. Proc. Natl. Acad. Sci. USA 1989, 86, 6567–6571. [Google Scholar] [CrossRef]
- Thompson, E.M.; Nagata, S.; Tsuji, F.I. Vargula hilgendorfii luciferase: A secreted reporter enzyme for monitoring gene expression in mammalian cells. Gene 1990, 96, 257–262. [Google Scholar] [CrossRef]
- Hunt, E.A.; Moutsiopoulou, A.; Broyles, D.; Head, T.; Dikici, E.; Daunert, S.; Deo, S.K. Expression of a soluble truncated Vargula luciferase in Escherichia coli. Protein Expr. Purif. 2017, 132, 68–74. [Google Scholar] [CrossRef]
- Tanahashi, Y.; Ohmiya, Y.; Honma, S.; Katsuno, Y.; Ohta, H.; Nakamura, H.; Honma, K.I. Continuous measurement of targeted promoter activity by a secreted bioluminescence reporter, Vargula hilgendorfii luciferase. Anal. Biochem. 2001, 289, 260–266. [Google Scholar] [CrossRef]
- Markova, S.V.; Golz, S.; Frank, L.A.; Kalthof, B.; Vysotski, E.S. Cloning and Expression of cDNA for a Luciferase from the Marine Copepod Metridia longa A NOVEL SECRETED BIOLUMINESCENT REPORTER ENZYME. J. Biol. Chem. 2004, 279, 3212–3217. [Google Scholar] [CrossRef]
- Song, G.; Wu, Q.-P.; Xu, T.; Liu, Y.-L.; Xu, Z.-G.; Zhang, S.-F.; Guo, Z.-Y. Quick preparation of nanoluciferase-based tracers for novel bioluminescent receptor-binding assays of protein hormones: Using erythropoietin as a model. J. Photochem. Photobiol. B Biol. 2015, 153, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Machleidt, T.; et al. Engineered Luciferase Reporter from a Deep Sea Shrimp Utilizing a Novel Imidazopyrazinone Substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- He, S.-X.; Song, G.; Shi, J.-P.; Guo, Y.-Q.; Guo, Z.-Y. Nanoluciferase as a novel quantitative protein fusion tag: Application for overexpression and bioluminescent receptor-binding assays of human leukemia inhibitory factor. Biochimie 2014, 106, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Guo, Z.; Sato, T.; Yuan, B.; Ma, Y.; Qian, D.; Zhong, J.; Jin, M.; Huang, P.; Che, L.; et al. Optimized application of the secreted Nano-luciferase reporter system using an affinity purification strategy. PLoS ONE 2018, 13, e0196617. [Google Scholar] [CrossRef]
- Wu, Y.; Chakrabortty, S.; Gropeanu, R.A.; Wilhelmi, J.; Xu, Y.; Er, K.S.; Kuan, S.L.; Koynov, K.; Chan, Y.; Weil, T. pH-Responsive Quantum Dots via an Albumin Polymer Surface Coating. J. Am. Chem. Soc. 2010, 132, 5012–5014. [Google Scholar] [CrossRef]
- Liu, H.-B.; Yan, Q.; Wang, C.; Liu, X.; Wang, C.; Zhou, X.-H.; Xiao, S.-J. Saccharide- and temperature-responsive polymer brushes grown on gold nanoshells for controlled release of diols. Colloids Surfaces A Physicochem. Eng. Asp. 2011, 386, 131–134. [Google Scholar] [CrossRef]
- Song, F.; Tang, P.S.; Durst, H.; Cramb, D.T.; Chan, W.C.W. Nonblinking Plasmonic Quantum Dot Assemblies for Multiplex Biological Detection. Angew. Chem. Int. Ed. 2012, 51, 8773–8777. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.; Zayats, M.; Willner, I. Semiconductor quantum dots for bioanalysis. Angew. Chem. Int. Ed. Engl. 2008, 47, 7602–7625. [Google Scholar] [CrossRef] [PubMed]
- Alivisatos, A.P.; Gu, W.; Larabell, C. Quantum dots as cellular probes. Annu. Rev. Biomed. Eng. 2005, 7, 55–76. [Google Scholar] [CrossRef]
- Gerion, D.; Pinaud, F.; Williams, S.C.; Parak, W.J.; Zanchet, D.; Weiss, S.; Alivisatos, A.P. Synthesis and Properties of Biocompatible Water-Soluble Silica-Coated CdSe/ZnS Semiconductor Quantum Dots. J. Phys. Chem. B 2001, 105, 8861–8871. [Google Scholar] [CrossRef]
- Aubert, T.; Soenen, S.J.; Wassmuth, D.; Cirillo, M.; Van Deun, R.; Braeckmans, K.; Hens, Z. Bright and Stable CdSe/CdS@SiO2 Nanoparticles Suitable for Long-Term Cell Labeling. ACS Appl. Mater. Interfaces 2014, 6, 11714–11723. [Google Scholar] [CrossRef]
- Aubert, T.; Grasset, F.; Mornet, S.; Duguet, E.; Cador, O.; Cordier, S.; Molard, Y.; Demange, V.; Mortier, M.; Haneda, H. Functional silica nanoparticles synthesized by water-in-oil microemulsion processes. J. Colloid Interface Sci. 2010, 341, 201–208. [Google Scholar] [CrossRef]
- Selvan, S.T.; Tan, T.T.; Ying, J.Y. Robust, Non-Cytotoxic, Silica-Coated CdSe Quantum Dots with Efficient Photoluminescence. Adv. Mater. 2005, 17, 1620–1625. [Google Scholar] [CrossRef]
- De, A.; Loening, A.M.; Gambhir, S.S. An improved bioluminescence resonance energy transfer strategy for imaging intracellular events in single cells and living subjects. Cancer Res. 2007, 67, 7175–7183. [Google Scholar] [CrossRef] [PubMed]
- Venisnik, K.M.; Olafsen, T.; Loening, A.M.; Iyer, M.; Gambhir, S.S.; Wu, A.M. Bifunctional antibody-Renilla luciferase fusion protein for in vivo optical detection of tumors. Protein Eng. Des. Sel. 2006, 19, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Dacres, H.; Michie, M.; Wang, J.; Pfleger, K.D.G.; Trowell, S.C. Effect of enhanced Renilla luciferase and fluorescent protein variants on the Forster distance of Bioluminescence resonance energy transfer (BRET). Biochem. Biophys. Res. Commun. 2012, 425, 625–629. [Google Scholar] [CrossRef]
- Alam, R.; Karam, L.M.; Doane, T.L.; Coopersmith, K.; Fontaine, D.M.; Branchini, B.R.; Maye, M.M. Probing Bioluminescence Resonance Energy Transfer in Quantum Rod-Luciferase Nanoconjugates. ACS Nano 2016, 10, 1969–1977. [Google Scholar] [CrossRef]
- Nie, S.; Emory, S.R. Probing Single Molecules and Single Nanoparticles by Surface-Enhanced Raman Scattering. Science 1997, 275, 1102–1106. [Google Scholar] [CrossRef] [PubMed]
- Bishop, K.J.M.; Wilmer, C.E.; Soh, S.; Grzybowski, B.A. Nanoscale forces and their uses in self-assembly. Small 2009, 5, 1600–1630. [Google Scholar] [CrossRef]
- Valentini, F.; Carbone, M.; Palleschi, G. Carbon nanostructured materials for applications in nano-medicine, cultural heritage, and electrochemical biosensors. Anal. Bioanal. Chem. 2013, 405, 451–465. [Google Scholar] [CrossRef]
- Wang, C.; Gao, X.; Su, X. In vitro and in vivo imaging with quantum dots. Anal. Bioanal. Chem. 2010, 397, 1397–1415. [Google Scholar] [CrossRef] [PubMed]
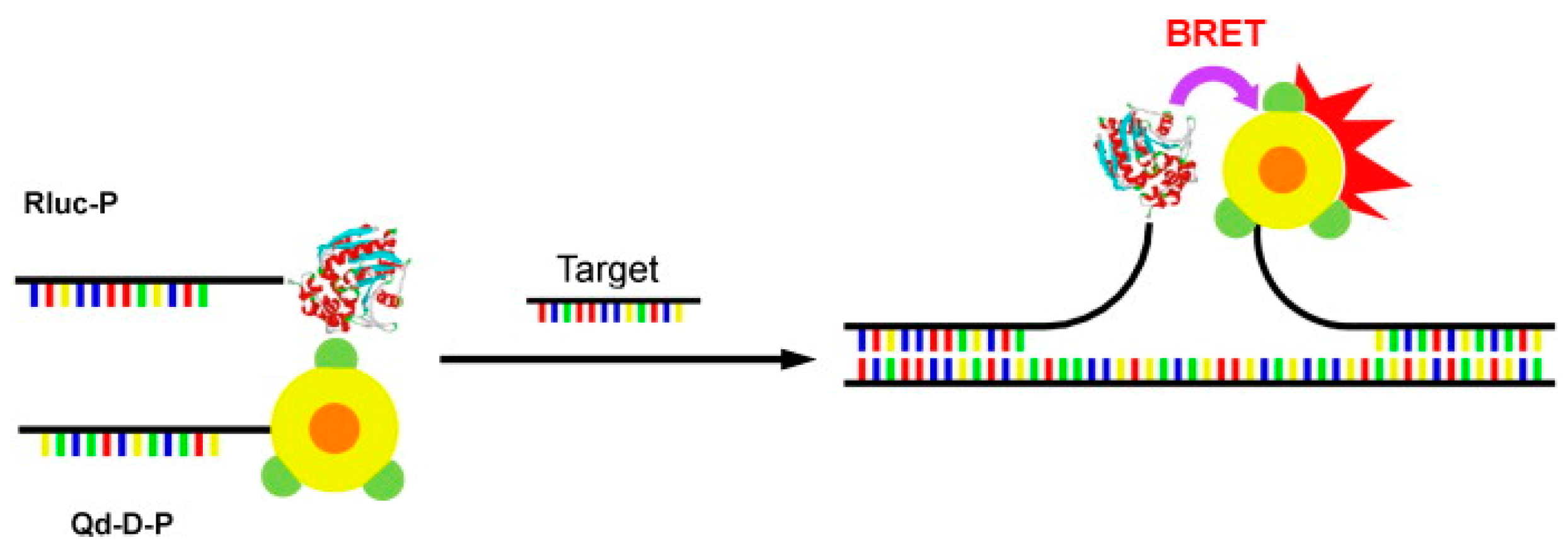
- Kumar, M.; Zhang, D.; Broyles, D.; Deo, S.K. A rapid, sensitive, and selective bioluminescence resonance energy transfer (BRET)-based nucleic acid sensing system. Biosens. Bioelectron. 2011, 30, 133–139. [Google Scholar] [CrossRef] [PubMed]
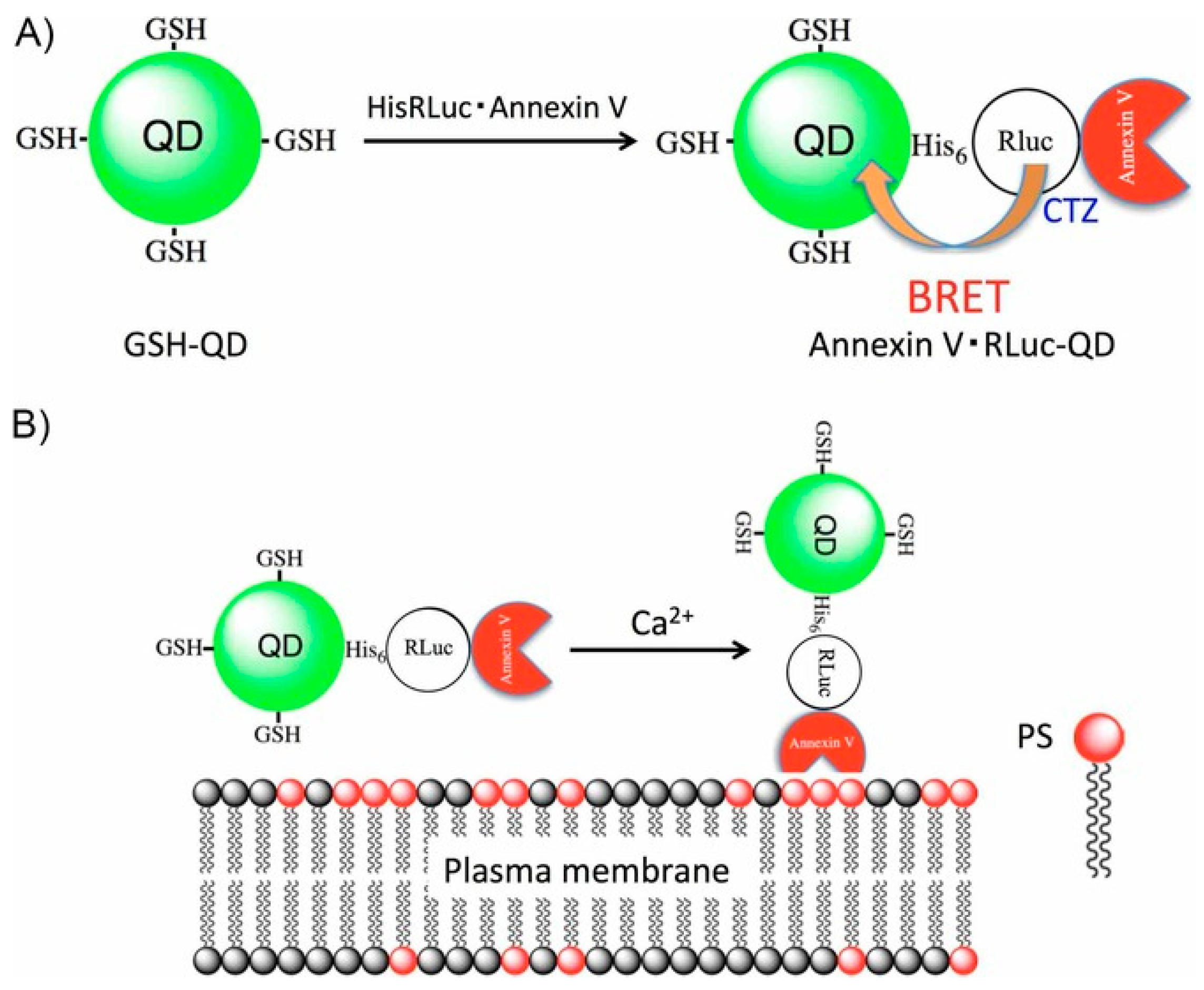
- Tsuboi, S.; Jin, T. Bioluminescence Resonance Energy Transfer (BRET)-coupled Annexin V-functionalized Quantum Dots for Near-Infrared Optical Detection of Apoptotic Cells. ChemBioChem 2017, 18, 2231–2235. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wen, K.; Wang, Z.; Zhang, X.; Li, C.; Zhang, S.; Shen, J. General Bioluminescence Resonance Energy Transfer Homogeneous Immunoassay for Small Molecules Based on Quantum Dots. Anal. Chem. 2016, 88, 3512–3520. [Google Scholar] [CrossRef] [PubMed]
- Kamkaew, A.; Sun, H.; England, C.G.; Cheng, L.; Liu, Z.; Cai, W. Quantum dot-NanoLuc bioluminescence resonance energy transfer enables tumor imaging and lymph node mapping in vivo. Chem. Commun. 2016, 52, 6997–7000. [Google Scholar] [CrossRef]
- Kosaka, N.; Mitsunaga, M.; Bhattacharyya, S.; Miller, S.C.; Choyke, P.L.; Kobayashi, H. Self-illuminating in vivo lymphatic imaging using a bioluminescence resonance energy transfer quantum dot nano-particle. Contrast Media Mol. Imaging 2011, 6, 55–59. [Google Scholar] [CrossRef]
- Feugang, J.M.; Youngblood, R.C.; Greene, J.M.; Fahad, A.S.; Monroe, W.A.; Willard, S.T.; Ryan, P.L. Application of quantum dot nanoparticles for potential non-invasive bio-imaging of mammalian spermatozoa. J. Nanobiotechnol. 2012, 10, 45. [Google Scholar] [CrossRef]
- Bungart, B.L.; Dong, L.; Sobek, D.; Sun, G.Y.; Yao, G.; Lee, J.C.M. Nanoparticle-emitted light attenuates amyloid-β-induced superoxide and inflammation in astrocytes. Nanomed. Nanotechnol. Biol. Med. 2014, 10, 15–17. [Google Scholar] [CrossRef]
- Dabbousi, B.O.; Rodriguez-Viejo, J.; Mikulec, F.V.; Heine, J.R.; Mattoussi, H.; Ober, R.; Jensen, K.F.; Bawendi, M.G. (CdSe)ZnS Core−Shell Quantum Dots: Synthesis and Characterization of a Size Series of Highly Luminescent Nanocrystallites. J. Phys. Chem. B 1997, 101, 9463–9475. [Google Scholar] [CrossRef]
- Yang, R.S.H.; Chang, L.W.; Wu, J.-P.; Tsai, M.-H.; Wang, H.-J.; Kuo, Y.-C.; Yeh, T.-K.; Yang, C.S.; Lin, P. Persistent Tissue Kinetics and Redistribution of Nanoparticles, Quantum Dot 705, in Mice: ICP-MS Quantitative Assessment. Environ. Health Perspect. 2007, 115, 1339–1343. [Google Scholar] [CrossRef]
- Widy-Tyszkiewicz, E.; Piechal, A.; Gajkowska, B.; Śmiałek, M. Tellurium-induced cognitive deficits in rats are related to neuropathological changes in the central nervous system. Toxicol. Lett. 2002, 131, 203–214. [Google Scholar] [CrossRef]
- Vinceti, M.; Wei, E.T.; Malagoli, C.; Bergomi, M.; Vivoli, G. Adverse health effects of selenium in humans. Rev. Environ. Health 2001, 16, 233–251. [Google Scholar] [CrossRef]
- UMEMURA, T. Experimental reproduction of itai-itai disease, a chronic cadmium poisoning of humans, in rats and monkeys. Jpn. J. Vet. Res. 2000, 48, 15–28. [Google Scholar]
- Klaassen, C.D.; Liu, J. Role of Metallothionein in Cadmium-Induced Hepatotoxicity and Nephrotoxicity. Drug Metab. Rev. 1997, 29, 79–102. [Google Scholar] [CrossRef] [PubMed]
- Bertin, G.; Averbeck, D. Cadmium: Cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (a review). Biochimie 2006, 88, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Sukhanova, A.; Bozrova, S.; Sokolov, P.; Berestovoy, M.; Karaulov, A.; Nabiev, I. Dependence of Nanoparticle Toxicity on Their Physical and Chemical Properties. Nanoscale Res. Lett. 2018, 13, 44. [Google Scholar] [CrossRef] [PubMed]
- Hardman, R. A Toxicologic Review of Quantum Dots: Toxicity Depends on Physicochemical and Environmental Factors. Environ. Health Perspect. 2006, 114, 165–172. [Google Scholar] [CrossRef]
- Xu, G.; Lin, G.; Lin, S.; Wu, N.; Deng, Y.; Feng, G.; Chen, Q.; Qu, J.; Chen, D.; Chen, S.; et al. The Reproductive Toxicity of CdSe/ZnS Quantum Dots on the in vivo Ovarian Function and in vitro Fertilization. Sci. Rep. 2016, 6, 37677. [Google Scholar] [CrossRef]
- Liu, J.; Yang, C.; Liu, J.; Hu, R.; Hu, Y.; Chen, H.; Law, W.-C.; Swihart, M.T.; Ye, L.; Wang, K.; et al. Effects of Cd-based Quantum Dot Exposure on the Reproduction and Offspring of Kunming Mice over Multiple Generations. Nanotheranostics 2017, 1, 23–37. [Google Scholar] [CrossRef]
- Reyes-Esparza, J.; Martínez-Mena, A.; Gutiérrez-Sancha, I.; Rodríguez-Fragoso, P.; de la Cruz, G.G.; Mondragón, R.; Rodríguez-Fragoso, L. Synthesis, characterization and biocompatibility of cadmium sulfide nanoparticles capped with dextrin for in vivo and in vitro imaging application. J. Nanobiotechnol. 2015, 13, 83. [Google Scholar] [CrossRef] [PubMed]
- Bargheer, D.; Giemsa, A.; Freund, B.; Heine, M.; Waurisch, C.; Stachowski, G.M.; Hickey, S.G.; Eychmüller, A.; Heeren, J.; Nielsen, P. The distribution and degradation of radiolabeled superparamagnetic iron oxide nanoparticles and quantum dots in mice. Beilstein J. Nanotechnol. 2015, 6, 111–123. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Size (kDa) | Emission (nm) | Substrate |
|---|---|---|---|
| Aequorin | 22 | 469 | Coelenterazine |
| Bacterial luciferase (Lux) | Alpha subunit: 40 Beta subunit: 35 | 490 | FMNH2 long-chain aliphatic aldehyde |
| Firefly luciferase (Fluc) | 61 | 562 | D-luciferin |
| Renilla luciferase (Rluc) | 36 | 480 | Coelenterazine |
| Gaussia luciferase (Gluc) | 19.9 | 480 | Coelenterazine |
| Vargula luciferase (Vluc) or Cypridina luciferase | 62 | 460 | Vargulin (Cypridina luciferin) |
| Metridia luciferase | 24 | 480 | Coelenterazine |
| Nano luciferase (Nluc) | 19 | 460 | Furimazine |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, E.; Song, J.; Zhang, J. Integration of Nanomaterials and Bioluminescence Resonance Energy Transfer Techniques for Sensing Biomolecules. Biosensors 2019, 9, 42. https://doi.org/10.3390/bios9010042
Hwang E, Song J, Zhang J. Integration of Nanomaterials and Bioluminescence Resonance Energy Transfer Techniques for Sensing Biomolecules. Biosensors. 2019; 9(1):42. https://doi.org/10.3390/bios9010042
Chicago/Turabian StyleHwang, Eugene, Jisu Song, and Jin Zhang. 2019. "Integration of Nanomaterials and Bioluminescence Resonance Energy Transfer Techniques for Sensing Biomolecules" Biosensors 9, no. 1: 42. https://doi.org/10.3390/bios9010042
APA StyleHwang, E., Song, J., & Zhang, J. (2019). Integration of Nanomaterials and Bioluminescence Resonance Energy Transfer Techniques for Sensing Biomolecules. Biosensors, 9(1), 42. https://doi.org/10.3390/bios9010042