Specific and Generic Immunorecognition of Glycopeptide Antibiotics Promoted by Unique and Multiple Orientations of Hapten



Abstract
:1. Introduction
- Additional facilities for group recognition of analytes by polyclonal antibodies can be provided by the affinity fractionation approach, using the principle of hapten heterology. A structurally related analyte as a heterologous immobilized hapten can bind only to the portion of antiserum antibodies directed against common epitopes. The assay developed according to the mentioned approach demonstrates improved group specificity, in comparison with homologous assay formats [16,17,18].
2. Materials and Methods
2.1. Chemicals
2.2. Synthesis of Conjugated Antigens
2.3. Immunization and Polyclonal Antibody Preparation
2.4. ELISA Procedure
3. Results and Discussion
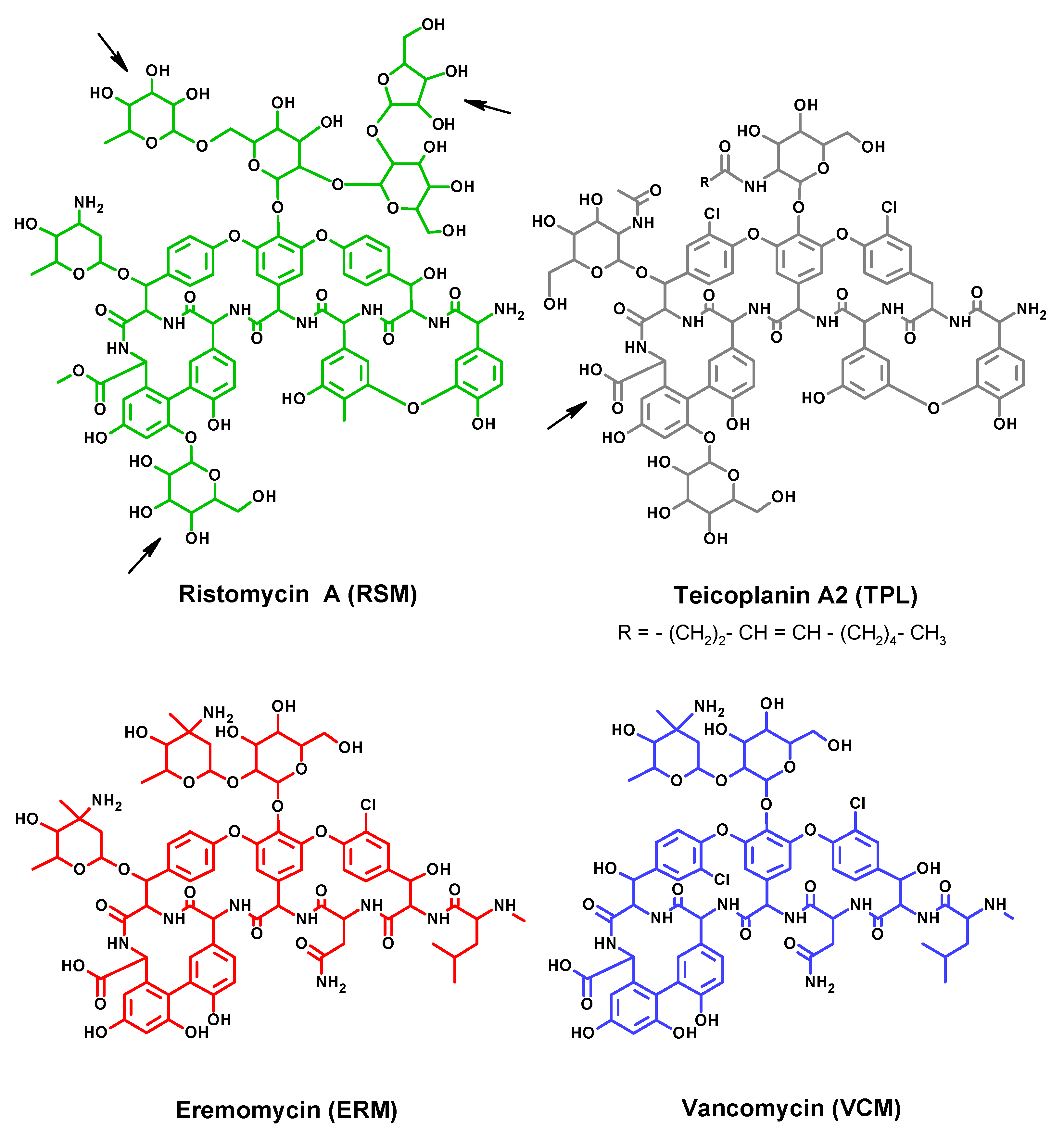
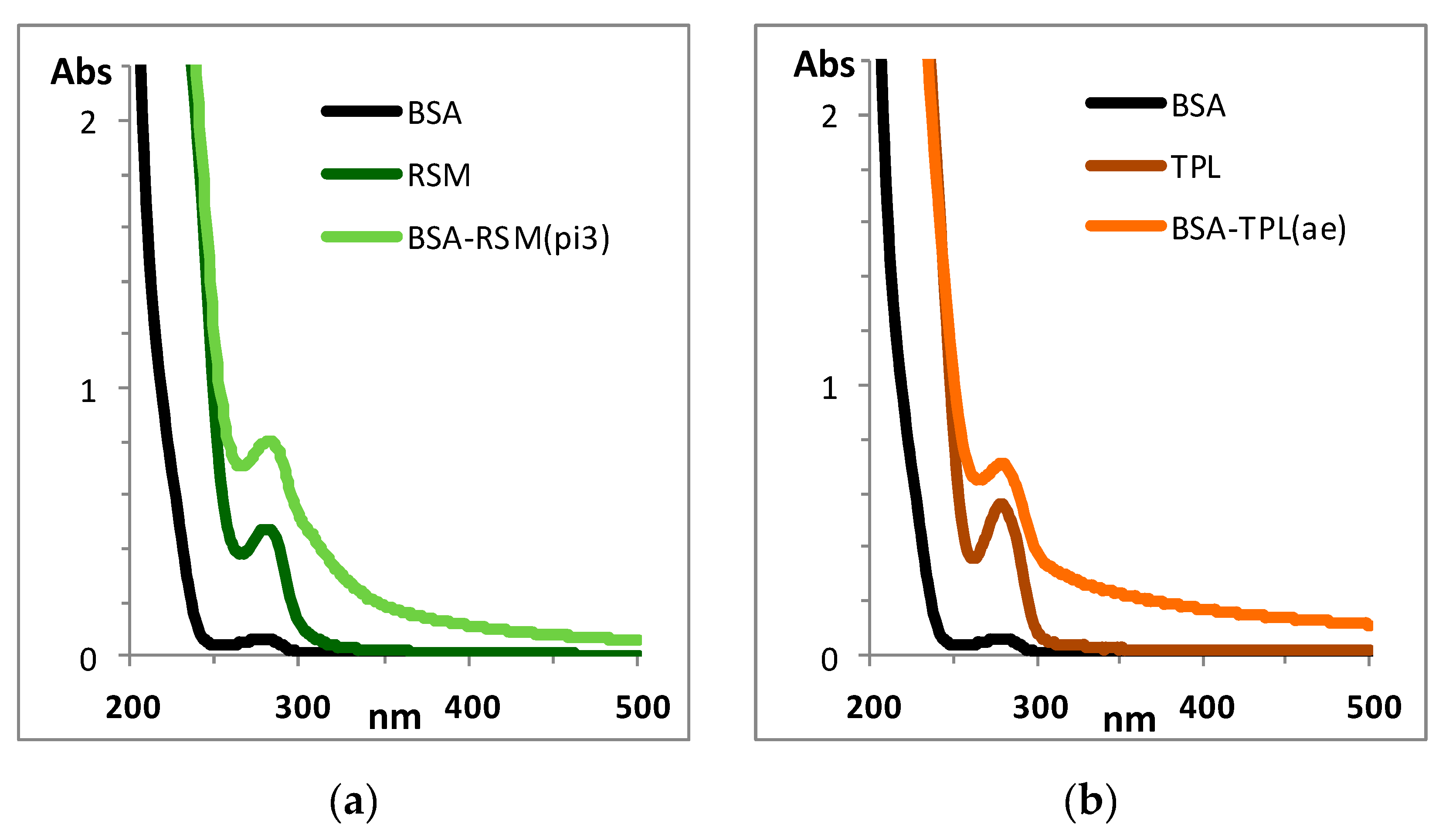
3.1. Synthesis and Characterization of Conjugated Antigens
3.2. Preparation of Polyclonal Antibodies
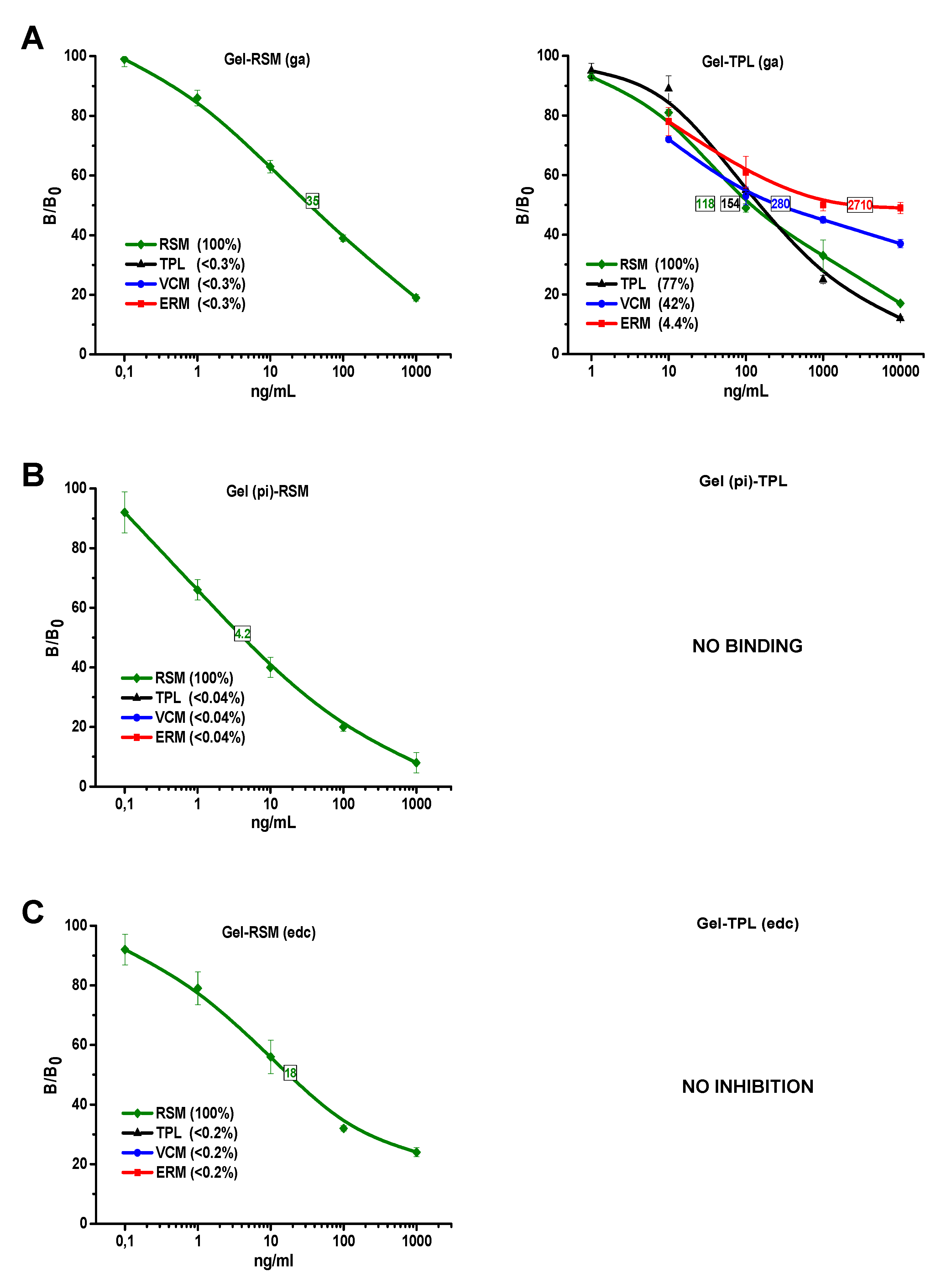
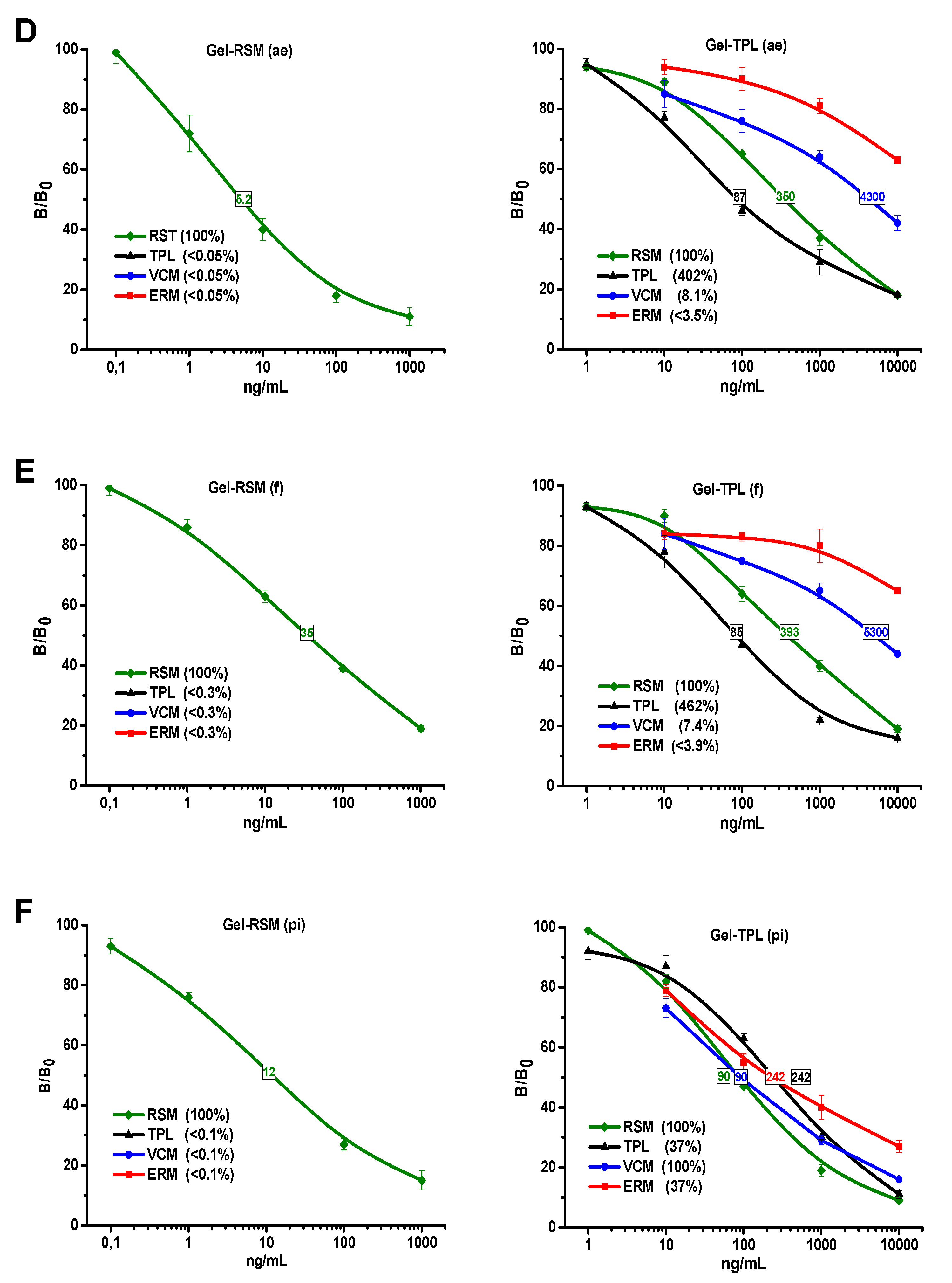
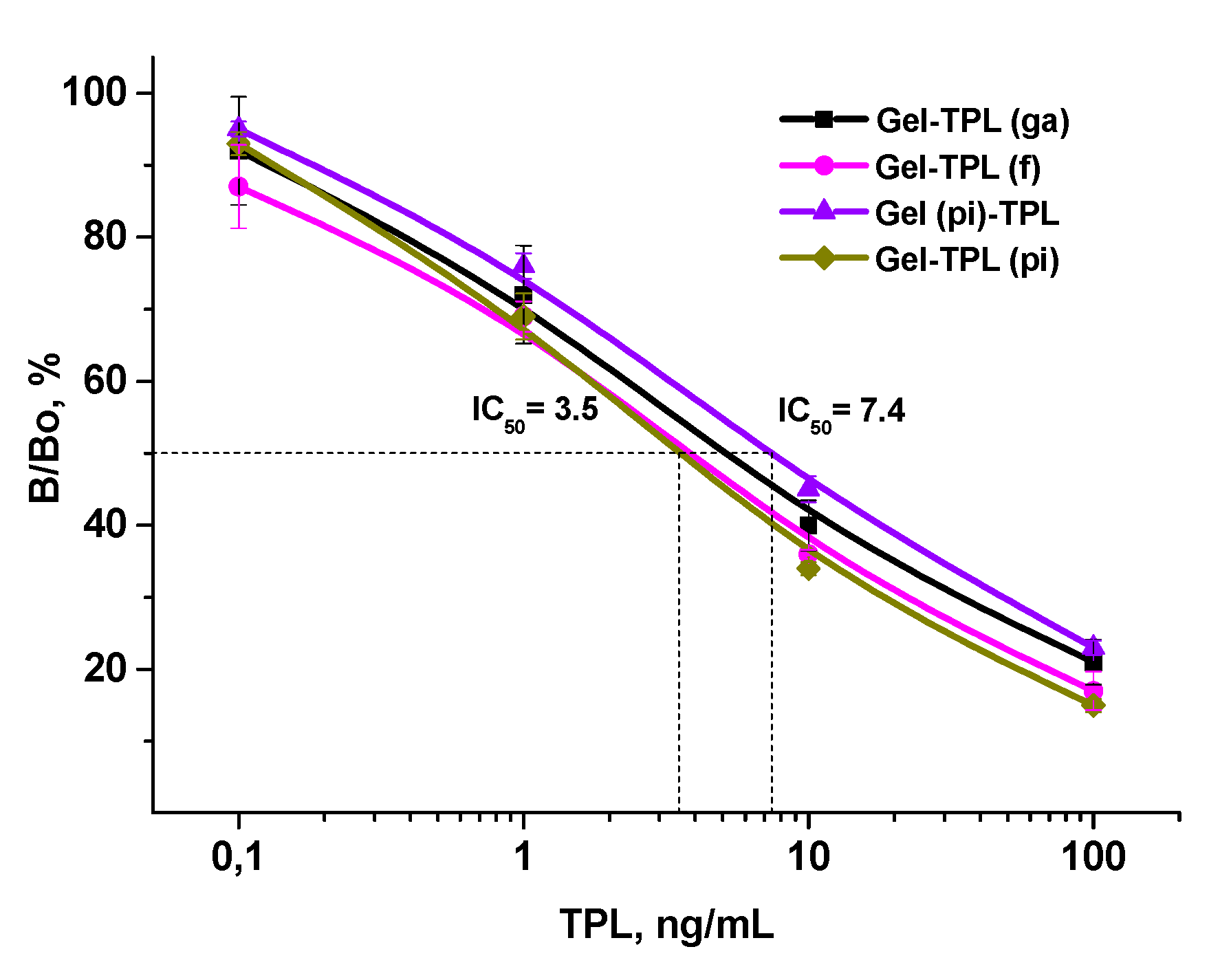
3.3. Examination of Assay Specificity Based on Prepared Immunoreagents
3.3.1. Anti-BSA-RSM(pi3)
3.3.2. Anti-BSA-TPL(ae)
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Xiao, Q.; Lin, J.-M. Advances and applications of chemiluminescence immunoassay in clinical diagnosis and foods safety. Chin. J. Anal. Chem. 2015, 43, 929–938. [Google Scholar] [CrossRef]
- González-Martínez, M.Á.; Puchades, R.; Maquieira, Á. Immunoanalytical technique: Enzyme-linked immunosorbent assay (ELISA). In Modern Techniques for Food Authentication, 2nd ed.; Academic Press: Cambridge, MA, USA, 2018; Chapter 15; pp. 617–657. [Google Scholar] [CrossRef]
- Raman Suri, C.; Boro, R.; Nangia., Y.; Gandhi, S.; Sharma, P.; Wangoo, N.; Rajesh, K.; Shekhawat, G.S. Immunoanalytical techniques for analyzing pesticides in the environment. Trends Anal. Chem. 2009, 28, 29–39. [Google Scholar] [CrossRef]
- Hao, X.; Huang, Y.; Qiu, M.; Yin, C.; Ren, H.; Gan, H.; Li, H.; Zhou, Y.; Xia, J.; Li, W.; et al. Immunoassay of S-adenosylmethionine and S-adenosylhomocysteine: The methylation index as a biomarker for disease and health status. BMC Res. Notes 2016, 9, 498. [Google Scholar] [CrossRef]
- Hino, Y.; Ojanperä, I.; Rasanen, I.; Vuori, E. Performance of immunoassays in screening for opiates, cannabinoids and amphetamines in post-mortem blood. Forensic Sci. Int. 2003, 131, 148–155. [Google Scholar] [CrossRef]
- Kononenko, G.P.; Burkin, A.A. Distribution of mycotoxins and usnic acid in the thalli of epigeous lichens. Biol. Bull. Russ. Acad. Sci. 2015, 42, 213–219. [Google Scholar] [CrossRef]
- Li, Y.-F.; Sun, Y.-M.; Beier, R.C.; Lei, H.-T.; Gee, S.; Hammock, B.D.; Wang, H.; Wang, Z.; Sun, X.; Shen, Y.; et al. Immunochemical techniques for multianalyte analysis of chemical residues in food and the environment: A review. TrAC Trends Anal. Chem. 2017, 88, 25–40. [Google Scholar] [CrossRef]
- Esteve-Turrillas, F.A.; Mercader, J.V.; Agulló, C.; Abad-Somovilla, A.; Abad-Fuentes, A. A class-selective immunoassay for simultaneous analysis of anilinopyrimidine fungicides using a rationally designed hapten. Analyst 2017, 142, 3975–3985. [Google Scholar] [CrossRef] [PubMed]
- Burkin, A.A.; Kononenko, G.P.; Soboleva, N.A. Group-specific antibodies against zearalenone and its metabolites and synthetic analogs. Appl. Biochem. Microbiol. 2002, 38, 169–176. [Google Scholar] [CrossRef]
- Burkin, M.A.; Galvidis, I.A. Simultaneous and differential determination of drugs and metabolites using the same antibody: Difloxacin and sarafloxacin case. Anal. Methods 2016, 8, 5843–5850. [Google Scholar] [CrossRef]
- Galvidis, I.A.; Lapa, G.B.; Burkin, M.A. Group determination of 14-membered macrolide antibiotics and azithromycin using antibodies against common epitopes. Anal. Biochem. 2015, 468, 75–82. [Google Scholar] [CrossRef]
- Rabia, L.A.; Desai, A.A.; Jhajj, H.S.; Tessier, P.M. Understanding and overcoming trade-offs between antibody affinity, specificity, stability and solubility. Biochem. Engineer. J. 2018, 137, 365–374. [Google Scholar] [CrossRef]
- Li, T.; Pantazes, R.J.; Maranas, C.D. OptMAVEn: A new framework for the de novo design of antibody variable region models targeting specific antigen epitopes. PLoS ONE 2014, 9, e105954. [Google Scholar] [CrossRef] [PubMed]
- Galvidis, I.A.; Wang, Z.; Nuriev, R.I.; Burkin, M.A. Broadening the detection spectrum of small analytes using a two-antibody-designed hybrid immunoassay. Anal. Chem. 2018, 90, 4901–4908. [Google Scholar] [CrossRef]
- Wang, F.; Wang, H.; Shen, Y.; Li, Y.; Dong, J.; Xu, Z.; Yang, J.; Sun, Y.; Xiao, Z. Bispecific monoclonal antibody-based multianalyte ELISA for furaltadone metabolite, malachite green, and leucomalachite green in aquatic products. J. Agric. Food Chem. 2016, 64, 8054–8061. [Google Scholar] [CrossRef] [PubMed]
- Burkin, M.A.; Galvidis, I.A. Hapten modification approach for switching immunoassay specificity from selective to generic. J. Immunol. Methods 2013, 388, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Huet, A.; Charlier, C.; Tittlemier, S.; Singh, G.; Benrejeb, S.; Delahaut, P. Simultaneous determination of (fluoro)quinolone antibiotics in kidney, marine products, eggs, and muscle by enzyme-linked immunosorbent assay (ELISA). J. Agric. Food Chem. 2006, 54, 2822–2827. [Google Scholar] [CrossRef] [PubMed]
- Fodey, T.; Murilla, G.; Cannavan, A.; Elliott, C. Characterisation of antibodies to chloramphenicol, produced in different species by enzyme-linked immunosorbent assay and biosensor technologies. Anal. Chim. Acta 2007, 592, 51–57. [Google Scholar] [CrossRef]
- Blaskovich, M.A.T.; Hansford, K.A.; Butler, M.S.; Jia, Z.; Mark, A.E.; Cooper, M.A. Developments in glycopeptide antibiotics. ACS Infect. Dis. 2018, 4, 715–735. [Google Scholar] [CrossRef] [PubMed]
- Gause, G.F.; Brazhnikova, M.G.; Lomakina, N.N.; Berdnikova, T.F.; Fedorova, G.B.; Tokareva, N.L.; Borisova, V.N.; Batta, G.Y. Eremomycin—new glycopeptide antibiotic: Chemical properties and structure. J. Antibiot. 1989, 42, 1790–1797. [Google Scholar] [CrossRef]
- Philip, J.E.; Schenck, J.R.; Hargie, M.P. Ristocetins A and B, two new antibiotics; isolation and properties. Antibiot. Annu. 1956, 3, 699–705. [Google Scholar]
- Weiss, H.J. Abnormalities of factor VIII and platelet aggregation: Use of ristocetin in diagnosing the von Willebrand syndrome. Blood 1975, 45, 403–412. [Google Scholar] [PubMed]
- Reynolds, P.E. Structure, biochemistry and mechanism of action of glycopeptide antibiotics. Eur. J. Clin. Microbiol. Infect. Dis. 1989, 8, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.C.; Peter, G. General immune practices. In Vaccines, 3rd ed.; Plotkin, S.A., Orenstein, W.A., Eds.; WB Saunders: Philadelphia, PA, USA, 1999; pp. 47–73. [Google Scholar]
- Ackerman, B.H.; Berg, H.G.; Strate, R.G.; Rotschafer, J.C. Comparison of radioimmunoassay and fluorescent polarization immunoassay for quantitative determination of vancomycin concentrations in serum. J. Clin. Microbiol. 1983, 18, 994–995. [Google Scholar] [PubMed]
- Mastin, S.H.; Buck, R.L.; Mueggler, P.A. Performance of a fluorescence polarization immunoassay for teicoplanin in serum. Diagn. Microbiol. Infect. Dis. 1993, 16, 17–24. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Krogstad, D.J.; Granich, G.G.; Murray, P.R. Laboratory evaluation of five assay methods for vancomycin: Bioassay, high-pressure liquid chromatography, fluorescence polarization immunoassay, radioimmunoassay, and fluorescence immunoassay. J. Clin. Microbiol. 1984, 20, 311–316. [Google Scholar]
- Lam, M.T.; Le, C.X. Competitive immunoassay for vancomycin using capillary electrophoresis with laser-induced fluorescence detection. Analyst 2002, 127, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Burkin, M.A.; Burkin, A.A. Enzyme immunoassay for the determinationof the glycopeptide antibiotic eremomycin. Appl. Biochem. Microbiol. 2009, 45, 210–214. [Google Scholar] [CrossRef]
- Corti, A.; Cavenaghi, L.; Giani, E.; Cassani, G. A receptor-antibody sandwich assay for teicoplanin. Clin. Chem. 1987, 33, 1615–1618. [Google Scholar]
- Karkishchenko, N.N.; Grachev, S.V. The Guide to Laboratory Animals and Alternative Models in Biomedical Researches; Profile-2C: Moscow, Russia, 2010; p. 358. [Google Scholar]
- Burkin, M.A.; Lapa, G.B.; Galvidis, I.A.; Burkin, K.M.; Zubkov, A.V.; Eremin, S.A. Three steps improving the sensitivity of sulfonamide immunodetection in milk. Anal. Meth. 2018, 10, 5773–5782. [Google Scholar] [CrossRef]
- Campbell, K.; Haughey, S.A.; van den Top, H.; van Egmond, H.; Vilariño, N.; Botana, L.M.; Elliott, C.T. Single laboratory validation of a surface plasmon resonance biosensor screening method for paralytic shellfish poisoning toxins. Anal. Chem. 2010, 82, 2977–2988. [Google Scholar] [CrossRef]
- Hermanson, G.T. Bioconjugate Techniques, 3nd ed.; Elsevier: Amsterdam, The Netherlands, 2013; p. 1140. [Google Scholar]
- Carraway, K.L.; Koshland, D.E., Jr. Reaction of tyrosine residues in proteins with carbodiimide reagents. Biochim. Biophys. Acta 1968, 160, 272–274. [Google Scholar] [CrossRef]
- Burkin, M.A.; Galvidis, I.A. Development of competitive indirect ELISA for determination of lincomycin in milk, eggs, and honey. J. Agric. Food Chem. 2010, 58, 9893–9898. [Google Scholar] [CrossRef] [PubMed]
- Burkin, M.A.; Galvidis, I.A. Simultaneous separate and group determination of tylosin and tilmicosin in foodstuffs using single antibody-based immunoassay. Food Chem. 2012, 132, 1080–1086. [Google Scholar] [CrossRef]
- Lamont, E.; Seaton, R.A.; Macpherson, M.; Semple, L.; Bell, E.; Thomson, A.H. Development of teicoplanin dosage guidelines for patients treated within an outpatient parenteral antibiotic therapy (OPAT) programme. J. Antimicrob. Chemother. 2009, 64, 181–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Zhong, M.; Wei, Y. Direct fluorescence polarization assay for the detection of glycopeptide antibiotics. Anal. Chem. 2010, 82, 7044–7048. [Google Scholar] [CrossRef]
- Corti, A.; Rurali, C.; Borghi, A.; Cassani, G. Solid-phase enzyme-receptor assay (SPERA): A competitive-binding assay for glycopeptide antibiotics of the vancomycin class. Clin. Chem. 1985, 31, 1606–1610. [Google Scholar] [PubMed]
- Van Weemen, B.K.; Schuurs, A.H.W.M. The influence of heterologous combinations of antiserum and enzyme-labeled estrogen on the characteristics of estrogen enzyme-immunoassays. Immunochem. 1975, 12, 667–670. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hapten Functional Groups | Coating Conjugates Prepared Using Various Coupling Methods on The Basis of Glycopeptides | Type of Bond Formed Between Protein (pr) and Glycopeptide (gp) | |
|---|---|---|---|
| RSM | TPL | ||
| Amine | Gel-RSM(ga) | Gel-TPL(ga) | [Pr-NH]-(CH2)5-[NH-Gp] |
| Gel(pi)-RSM | Gel(pi)-TPL | [Pr=CH]-[NH-Gp] | |
| Carboxyl | Gel-TPL(edc) | [Pr-NH]-[CO-Gp] | |
| Gel-TPL(ae) | [Pr-NH]-[CO-Gp] | ||
| Phenolic, Resorcylic | Gel-RSM(f) | Gel-TPL(f) | [Pr-NH]-CH2-[CH=Gp] |
| Gel-RSM(edc) * | |||
| Gel-RSM(ae) * | |||
| Carbohydrate hydroxyls | Gel-RSM(pi-1) | Gel-TPL(pi-1) | [Pr-NH]-[CH=Gp] |
| Gel-RSM(pi-3) | Gel-TPL(pi-3) | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burkin, M.A.; Galvidis, I.A.; Eremin, S.A. Specific and Generic Immunorecognition of Glycopeptide Antibiotics Promoted by Unique and Multiple Orientations of Hapten. Biosensors 2019, 9, 52. https://doi.org/10.3390/bios9020052
Burkin MA, Galvidis IA, Eremin SA. Specific and Generic Immunorecognition of Glycopeptide Antibiotics Promoted by Unique and Multiple Orientations of Hapten. Biosensors. 2019; 9(2):52. https://doi.org/10.3390/bios9020052
Chicago/Turabian StyleBurkin, Maksim A., Inna A. Galvidis, and Sergei A. Eremin. 2019. "Specific and Generic Immunorecognition of Glycopeptide Antibiotics Promoted by Unique and Multiple Orientations of Hapten" Biosensors 9, no. 2: 52. https://doi.org/10.3390/bios9020052
APA StyleBurkin, M. A., Galvidis, I. A., & Eremin, S. A. (2019). Specific and Generic Immunorecognition of Glycopeptide Antibiotics Promoted by Unique and Multiple Orientations of Hapten. Biosensors, 9(2), 52. https://doi.org/10.3390/bios9020052