The Application of a Nanomaterial Optical Fiber Biosensor Assay for Identification of Brucella Nomenspecies


Abstract
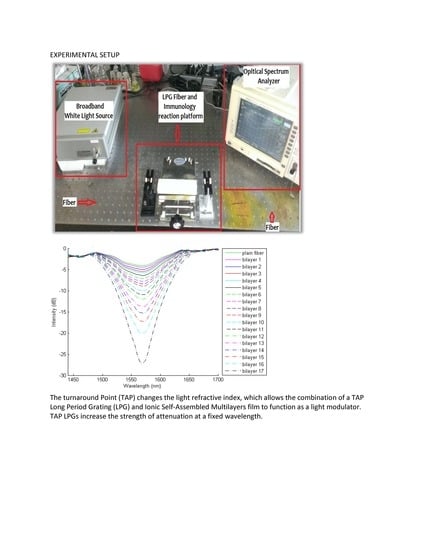
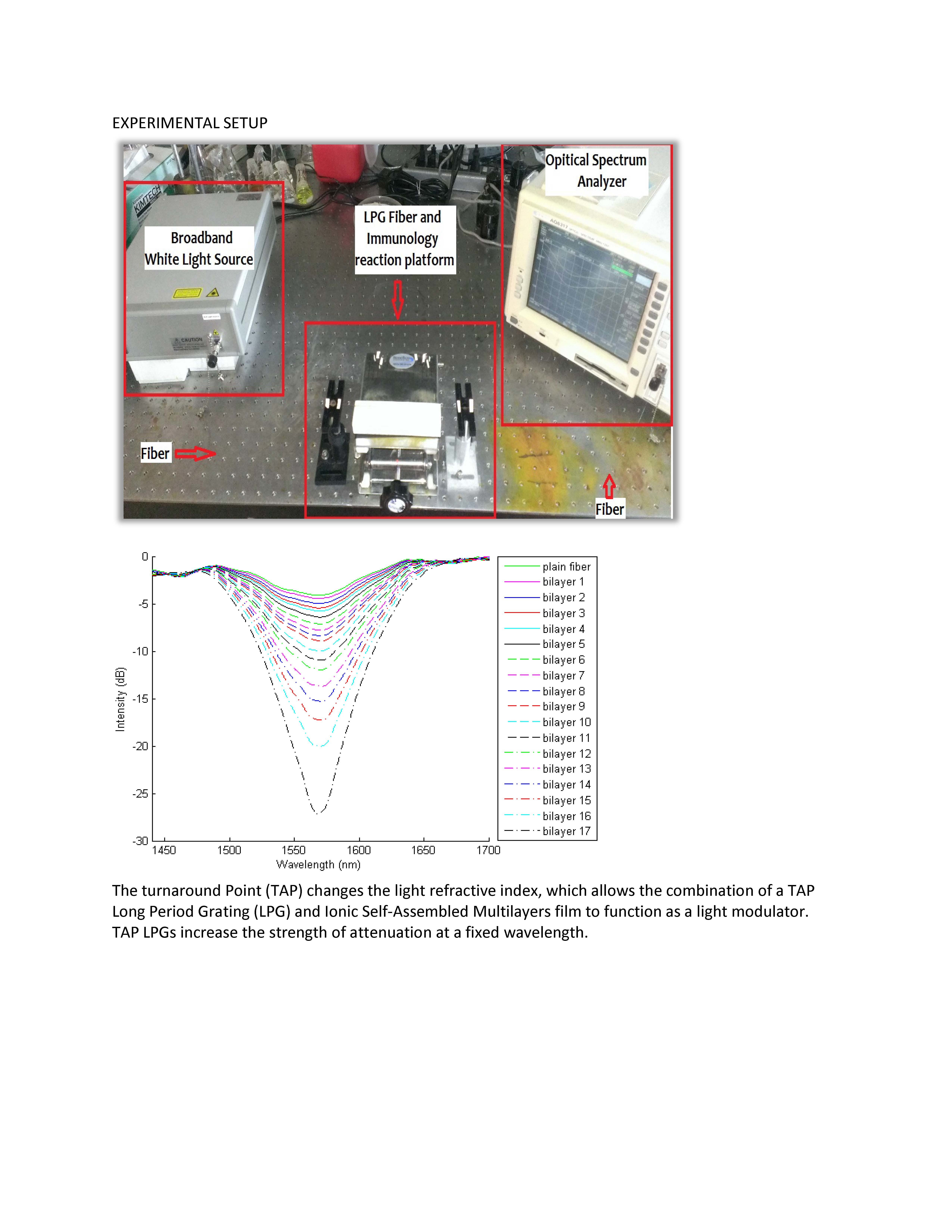
:
1. Introduction
2. Materials and Methods
2.1. Oligonucleotide Primers and Probes
2.2. Bacterial Strains and Culture Conditions Used
2.3. PCR
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
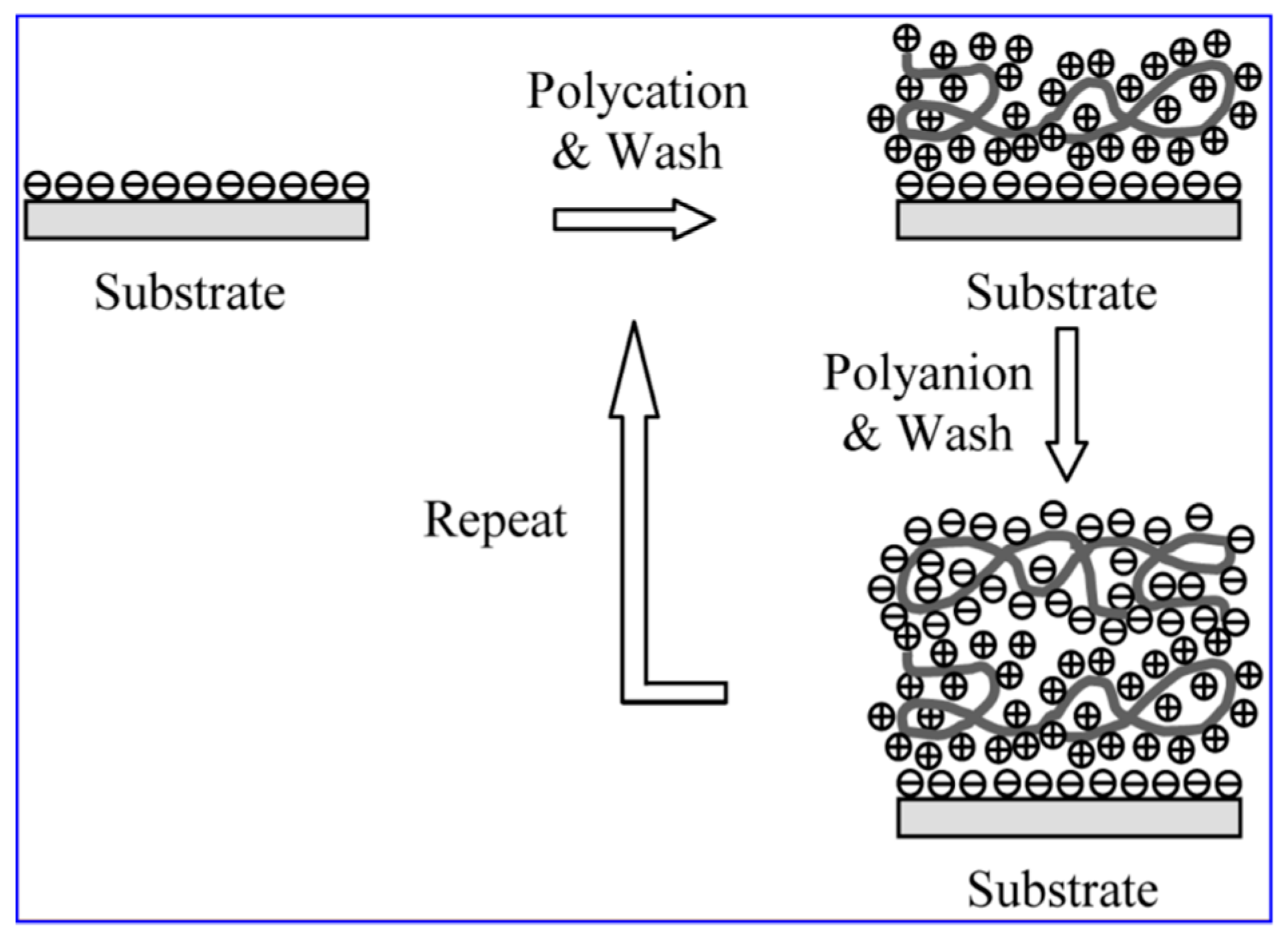
2.5. Fabrication of the ISAM Film
2.6. Coupling the Probe to the ISAM Film
2.7. Conjugation of Streptavidin to the ISAM Film
2.8. NOFS Assay
2.9. Detection of Brucella DNA in Tissues of Infected Mice
2.10. Statistical Analyses
3. Results
3.1. Specificity of the DNA Probes
3.2. Validation of Target DNA for Hybridization to the DNA Probe
3.3. Identification of Brucella Nomenspecies by NOFS Assay
3.4. NOFS Assay to Detect and Distinguish Different Brucella Nomenspecies and to Distingusih Brucella from Non-Brucella Bacterial Types
3.5. Identification of Brucella in Tissues from Infected Mice by NOFS Assay
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Corbel, M.J. International committee on systematic bacteriology subcommittee on the taxonomy of Brucella—Report of the Meeting, 5 September 1986, Manchester, England. Int. J. Syst. Bacteriol. 1988, 38, 450–452. [Google Scholar] [CrossRef]
- Mercier, E.; JumasBilak, E.; AllardetServent, A.; Ocallahan, D.; Ramuz, M. Polymorphism in Brucella strains detected by studying distribution of two short repetitive DNA elements. J. Clin. Microbiol. 1996, 34, 1299–1302. [Google Scholar]
- Jahans, K.L.; Foster, G.; Broughton, E.S. The characterisation of Brucella strains isolated from marine mammals. Vet. Microbiol. 1997, 57, 373–382. [Google Scholar] [CrossRef]
- Verger, J.M.; Grayon, M.; Cloeckaert, A.; Lefevre, M.; Ageron, E.; Grimont, F. Classification of Brucella strains isolated from marine mammals using DNA-DNA hybridization and ribotyping. Res. Microbiol. 2000, 151, 797–799. [Google Scholar] [CrossRef]
- Buchanan, T.M.; Hendrick, S.L.; Patton, C.M.; Feldman, R.A. Brucellosis in United States, 1960–1972—Abattoir associated disease. 3. Epidemiology and evidence for acquired immunity. Medicine 1974, 53, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Young, E.J. Human Brucellosis. Rev. Infect. Dis. 1983, 5, 821–842. [Google Scholar] [CrossRef]
- Cheville, N.F.; McCullough, D.R.; Paulson, L.R. Brucellosis in the Greater Yellowstone area. National Research Council. In Brucellosis in the Greater Yellowstone Area; The National Acadimies Press: Washington, DC, USA, 1998. [Google Scholar]
- Thorne, E.T. Brucellosis. In Infectious Diseases of Wild Mammals, 3rd ed.; Williams, E.S., Barker, I.K., Eds.; Wiley Online Library: Hoboken, NJ, USA, 2001; pp. 72–395. [Google Scholar]
- Cross, P.C.; Edwards, W.H.; Scurlock, B.M.; Maichak, E.J.; Rogerson, J.D. Effects of management and climate on elk brucellosis in the Greater Yellowstone Ecosystem. Ecol. Appl. 2007, 17, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Maichak, E.J.; Scurlock, B.M.; Rogerson, J.D.; Meadows, L.L.; Barbknecht, A.E.; Edwards, W.H.; Cross, P.C. Effects of management, behavior, and scavenging on risk of brucellosis transmission ifn Elk of Western Wyoming. J. Wildl. Dis. 2009, 45, 398–410. [Google Scholar] [CrossRef]
- Ramsden, J.J. Optical biosensors. J. Mol. Recognit. 1997, 10, 109–120. [Google Scholar] [CrossRef]
- Kersey, A.D.; Davis, M.A.; Patrick, H.J.; LeBlanc, M.; Koo, K.P.; Askins, C.G.; Putnam, M.A.; Friebele, E.J. Fiber Grating Sensors. J. Lightwave Technol. 1997, 15, 1442–1463. [Google Scholar] [CrossRef]
- Decher, G. Fuzzy nanoassemblies. Science 1997, 277, 1232–1237. [Google Scholar] [CrossRef]
- Decher, G.H.J.D.; Hong, J.D.; Schmitt, J. Buildup of ultrathin multilayer films by a self-assembly process: Consecutively alternating adsorption of anionic and cationic polyelectrolytes on charged surfaces. Thin Solid Film. 1992, 210/211, 831–835. [Google Scholar] [CrossRef]
- Wang, Z.; Heflin, J.R.; Van Cott, K.; Stolen, R.H.; Ramachandran, S.; Ghalmi, S. Biosensors employing ionic self-assembled multilayers adsorbed on long-period fiber gratings. Sens. Actuators B Chem. 2009, 139, 618–623. [Google Scholar] [CrossRef]
- Wang, Z.; Heflin, J.; Stolen, R.; Ramachandran, S. Analysis of optical response of long period fiber gratings to nm-thick thin-film coating. Opt. Express 2005, 13, 2808–2813. [Google Scholar] [CrossRef]
- Bandara, A.B.; Zuo, Z.; Ramachandran, S.; Ritter, A.; Heflin, J.R.; Inzana, T.J. Detection of methicillin-resistant staphylococci by biosensor assay consisting of nanoscale films on optical fiber long-period gratings. Biosens. Bioelectron. 2015, 70, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Chiavaioli, F.; Baldini, F.; Tombelli, S.; Trono, C.; Giannetti, A. Biosensing with optical fiber gratings. Nanophotonics 2017, 6, 663–679. [Google Scholar] [CrossRef]
- Esposito, F.; Sansone, L.; Taddei, C.; Campopiano, S.; Giordano, M.; Iadicicco, A. Ultrasensitive biosensor based on long period grating coated with polycarbonate-graphene oxide multilayer. Sens. Actuators B Chem. 2018, 274, 517–526. [Google Scholar] [CrossRef]
- Piestrzyńska, M.; Dominik, M.; Kosiel, K.; Janczuk-Richter, M.; Szot-Karpińska, K.; Brzozowska, E.; Shao, L.; Niedziółka-Jonsson, J.; Bock, W.J.; Śmietana, M. Ultrasensitive tantalum oxide nano-coated long-period gratings for detection of various biological targets. Biosens. Bioelectron. 2019, 133, 8–15. [Google Scholar] [CrossRef]
- Bandara, A.B.; Zuo, Z.; McCutcheon, K.; Ramachandran, S.; Heflin, J.R.; Inzana, T.J. Identification of Histophilus somni by a nanomaterial optical fiber biosensor assay. J. Vet. Diagn. Investig. 2018, 30, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, D.K.; Schulze, H.; Henihan, G.; Ciani, I.; Giraud, G.; Terry, J.G.; Walton, A.J.; Pethig, R.; Ghazal, P.; Crain, J.; et al. Impedimetric detection of single-stranded PCR products derived from methicillin resistant Staphylococcus aureus (MRSA) isolates. Biosens. Bioelectron. 2012, 34, 178–184. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Chen, P.; Zhou, X.; Yang, Y.; Wu, S.; Niu, L.; Han, Y.; Wang, L.; Chen, P.; et al. Label-free, electrochemical detection of methicillin-resistant Staphylococcus aureus DNA with reduced graphene oxide-modified electrodes. Biosens. Bioelectron. 2011, 26, 3883–3886. [Google Scholar] [CrossRef]
- Tombelli, S.; Minunni, M.; Santucci, A.; Spiriti, M.M.; Mascini, M. A DNA-based piezoelectric biosensor: Strategies for coupling nucleic acids to piezoelectric devices. Talanta 2006, 68, 806–812. [Google Scholar] [CrossRef]
- Yang, K.L.A.; Wu, S.Y.; Kwok, H.C.; Ho, H.P.; Kong, S.K. Using loop-mediated isothermal DNA amplification (LAMP) and spectral surface plasmon resonance (SPR) to detect methicillin-resistance S. aureus (MRSA). In Proceedings of the 2012 International Conference on Biomedical Engineering and Biotechnology, Macao, China, 28–30 May 2012. [Google Scholar] [CrossRef]
- Gotuzzo, E.; Carrillo, C.; Guerra, J.; Llosa, L. An evaluation of diagnostic methods for brucellosis—The value of bone marrow culture. J. Infect. Dis. 1986, 153, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Bricker, B.J.; Halling, S.M. Differentiation of Brucella abortus BV-1, Brucella abortus BV-2, and Brucella abortus BV-4, Brucella mellitensis, Brucella ovis, and Brucella suis BV-1 by PCR. J. Clin. Microbiol. 1994, 32, 2660–2666. [Google Scholar]
- Matar, G.M.; Khneisser, I.A.; Abdelnoor, A.M. Rapid laboratory confirmation of human brucellosis by PCR analysis of a target sequence on the 31-kilodalton Brucella antigen DNA. J. Clin. Microbiol. 1996, 34, 477–478. [Google Scholar] [PubMed]
- Wellinghausen, N.; Nockler, K.; Sigge, A.; Bartel, M.; Essig, A.; Poppert, S. Rapid detection of Brucella spp. in blood cultures by fluorescence in situ hybridization. J. Clin. Microbiol. 2006, 44, 1828–1830. [Google Scholar] [CrossRef]
- Rijpens, N.P.; Jannes, G.; VanAsbroeck, M.; Rossau, R.; Herman, L.M.F. Direct detection of Brucella spp. in raw milk by PCR and reverse hybridization with 16S-23S rRNA spacer probes. Appl. Environ. Microbiol. 1996, 62, 1683–1688. [Google Scholar]
- Bricker, B.J.; Ewalt, D.R.; Halling, S.M. Brucella ‘HOOF-Prints’: Strain typing by multi-locus analysis of variable number tandem repeats (VNTRs). BMC Microbiol. 2003, 3, 15. [Google Scholar] [CrossRef]
- Ewalt, D.R.; Bricker, B.J. Validation of the abbreviated Brucella AMOS PCR as a rapid screening method for differentiation of Brucella abortus field strain isolates and the vaccine strains, 19 and RB51. J. Clin. Microbiol. 2000, 38, 3085–3086. [Google Scholar]
- Probert, W.S.; Schrader, M.N.; Khuong, N.Y.; Bystrom, S.L.; Graves, M.H. Real-time multiplex PCR assay for detection of Brucella spp., B. abortus, and B. melitensis. J. Clin. Microbiol. 2004, 42, 1290–1293. [Google Scholar] [CrossRef] [PubMed]
- Schmalz, G.; Tsigaras, S.; Rinke, S.; Kottmann, T.; Haak, R.; Ziebolz, D. Detection of five potentially periodontal pathogenic bacteria in peri-implant disease: A comparison of PCR and real-time PCR. Diagn. Microbiol. Infect. Dis. 2016, 85, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Thorn, M.; Rorsman, F.; Ronnblom, A.; Sangfelt, P.; Wanders, A.; Eriksson, B.M.; Bondeson, K. Active cytomegalovirus infection diagnosed by real-time PCR in patients with inflammatory bowel disease: A prospective, controlled observational study. Scand. J. Gastroenterol. 2016, 51, 1075–1080. [Google Scholar] [CrossRef] [PubMed]
- Fellahi, S.; Harrak, M.E.; Kuhn, J.H.; Sebbar, G.; Bouaiti, E.A.; Khataby, K.; Fihri, O.F.; Houadfi, M.E.; Ennaji, M.M. Comparison of SYBR green I real-time RT-PCR with conventional agarose gel-based RT-PCR for the diagnosis of infectious bronchitis virus infection in chickens in Morocco. BMC Res. Notes 2016, 9, 231. [Google Scholar] [CrossRef]
- Cooper, K.L.; Bandara, A.B.; Wang, Y.; Wang, A.; Inzana, T.J. Photonic biosensor assays to detect and distinguish subspecies of Francisella tularensis. Sensors 2011, 11, 3004–3019. [Google Scholar] [CrossRef]
- eXtension. Brucellosis in Feral Hogs. Cooperative Extension Service. 2012. Available online: https://articles.extension.org/pages/65499/brucellosis-in-feral-hogs (accessed on 18 March 2019).
- Giurgiutiu, D.; Banis, C.; Hunt, E.; Mincer, J.; Nicolardi, C.; Weltman, A.; Stanek, D.; Matthews, S.; Siegenthaler, C.; Blackmore, C.; Tiller, R.; et al. Centers for Disease Control. Brucella suis infection associated with feral swine hunting—Three States, 2007–2008. Morb. Mortal. Wkly. Rep. 2009, 58, 618–621. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe/Primer Name | Sequence (5′ to 3′) | Comments |
|---|---|---|
| Probe-IS711-BIO | AAGCCAACACCCGGCCATTATGGT | The probe was biotinylated at the 3′ end |
| Probe-IS711-DIG | GGCCTACCGCTGCGAATA | The probe was labeled with digoxigenin at the 5′ end |
| Primer-IS711-For | TTGGCCTTGATCTGAGCCGT | |
| Primer-IS711-Rev | ATCGAAAGTCCACGCAGATG | |
| Probe-BruAb2_0168 | TGGAACGACCTTTGCAGGCGAGATC | Biotin label at the 3′ end; specific for B. abortus |
| Probe_Melitensis_0466 | CCAGCTTTTGGCCTTTTCCAGATTG | Biotin label at the 3′ end; specific for B. melitensis |
| Probe_Suis-TraJ | CCATGAGCGCCCGCATGTCCTCTTG | Biotin label at the 3′ end; specific for B. suis |
| Bacterial Species | Strain Name | Source |
|---|---|---|
| Brucella abortus | 2308 | Virginia Tech Biosafety 3 laboratory |
| Brucella melitensis | 16 M | Virginia Tech Biosafety 3 laboratory |
| Brucella suis | 1330 | Virginia Tech Biosafety 3 laboratory |
| Klebsiella pneumoniae | 2237 | Virginia Tech Veterinary Teaching Hospital |
| Klebsiella pneumoniae | 2237-13 | Virginia Tech Veterinary Teaching Hospital |
| Proteus mirabilis | 2172B | Virginia Tech Veterinary Teaching Hospital |
| Proteus mirabilis | 13-2319 | Virginia Tech Veterinary Teaching Hospital |
| Proteus mirabilis | 13-2401 | Virginia Tech Veterinary Teaching Hospital |
| Staphylococcus aures | 29213 | Virginia Tech Veterinary Teaching Hospital |
| Enterococcus faecalis | 2174 | Virginia Tech Veterinary Teaching Hospital |
| Enterococcus faecalis | 13-2321-2 | Virginia Tech Veterinary Teaching Hospital |
| Escherichia coli | 2174 | Virginia Tech Veterinary Teaching Hospital |
| Escherichia coli | Top10 | ThermoFisher Scientific |
| Escherichia coli | 13-2438 | Virginia Tech Veterinary Teaching Hospital |
| Salmonella arizonae | 13-2453 | Virginia Tech Veterinary Teaching Hospital |
| Enterobacter aerogenes | 13-2329-2 | Virginia Tech Veterinary Teaching Hospital |
| Enterococcus faecium | 13-2174-C | Virginia Tech Veterinary Teaching Hospital |
| Enterococcus faecium | 13-2248-2 | Virginia Tech Veterinary Teaching Hospital |
| Bacterial Species | Strain Name | Transmission Attenuation (Mean ± Standard Deviation) |
|---|---|---|
| Klebsiella pneumoniae | 2237 | −1.67% ± 0.15% |
| Klebsiella pneumoniae | 2237-13 | 2.09% ± 2.63% |
| Proteus mirabilis | 2172B | 0.65% ± 0.31% |
| Proteus mirabilis | 13-2319 | 0.70% ± 1.64% |
| Proteus mirabilis | 13-2401 | 0.25% ± 0.95% |
| Staphylococcus aures | 29213 | 1.30% ± 0.70% |
| Enterococcus faecalis | 2174 | 1.57% ± 1.03% |
| Enterococcus faecalis | 13-2321-2 | 1.04% ± 0.85% |
| Escherichia coli | 2174 | 0.23% ± 0.06% |
| Escherichia coli | Top10 | −0.62% ± 0.94% |
| Escherichia coli | 13-2438 | 1.54% ± 0.12% |
| Salmonella arizonae | 13-2453 | −0.59% ± 1.27% |
| Enterobacter aerogenes | 13-2329-2 | 1.64% ± 0.46% |
| Enterococcus faecium | 13-2174-C | 2.14% ± 0.10% |
| Enterococcus faecium | 13-2248-2 | 1.69% ± 0.38% |
| Challenge | Organ | Average % Attenuation ± Standard Deviation | Test Result b |
|---|---|---|---|
| With probe BruAb2_0168 (specific for B. abortus) B. abortus | Spleen (from mice 1 and 2) | 6.80 ± 0.32 | Positive |
| Liver (from mice 1 and 2) | 3.38 ± 0.78 | Positive | |
| PBS | Spleen (from mice 7 and 8) | −0.74 ± 1.12 | Negative |
| Liver (from mice 7 and 8) | −1.78 ± 0.29 | Negative | |
| With probe Melitensis_0466 (specific for B. melitensis) B. melitensis | Spleen (from mice 3 and 4) | 7.60 ± 1.86 | Positive |
| Liver (from mice 3 and 4) | 6.13 ± 2.81 | Positive | |
| PBS | Spleen (from mice 7 and 8) | −1.78 ± 2.60 | Negative |
| Liver (from mice 7 and 8) | −0.64 ± 0.41 | Negative | |
| With probe Suis_TraJ (specific for B. suis) B. suis | Spleen (from mice 5 and 6) | 6.94 ± 0.59 | Positive |
| Liver (from mice 5 and 6) | 5.10 ± 1.45 | Positive | |
| PBS | Spleen (from mice 7 and 8) | 0.72 ± 2.93 | Negative |
| Liver (from mice 7 and 8) | 0.34 ± 1.59 | Negative |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCutcheon, K.; Bandara, A.B.; Zuo, Z.; Heflin, J.R.; Inzana, T.J. The Application of a Nanomaterial Optical Fiber Biosensor Assay for Identification of Brucella Nomenspecies. Biosensors 2019, 9, 64. https://doi.org/10.3390/bios9020064
McCutcheon K, Bandara AB, Zuo Z, Heflin JR, Inzana TJ. The Application of a Nanomaterial Optical Fiber Biosensor Assay for Identification of Brucella Nomenspecies. Biosensors. 2019; 9(2):64. https://doi.org/10.3390/bios9020064
Chicago/Turabian StyleMcCutcheon, Kelly, Aloka B. Bandara, Ziwei Zuo, James R. Heflin, and Thomas J. Inzana. 2019. "The Application of a Nanomaterial Optical Fiber Biosensor Assay for Identification of Brucella Nomenspecies" Biosensors 9, no. 2: 64. https://doi.org/10.3390/bios9020064
APA StyleMcCutcheon, K., Bandara, A. B., Zuo, Z., Heflin, J. R., & Inzana, T. J. (2019). The Application of a Nanomaterial Optical Fiber Biosensor Assay for Identification of Brucella Nomenspecies. Biosensors, 9(2), 64. https://doi.org/10.3390/bios9020064