Advances in Directly Amplifying Nucleic Acids from Complex Samples

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Literature Search
2.2. Record Screening
2.3. Data Abstraction
2.4. Visualization Process
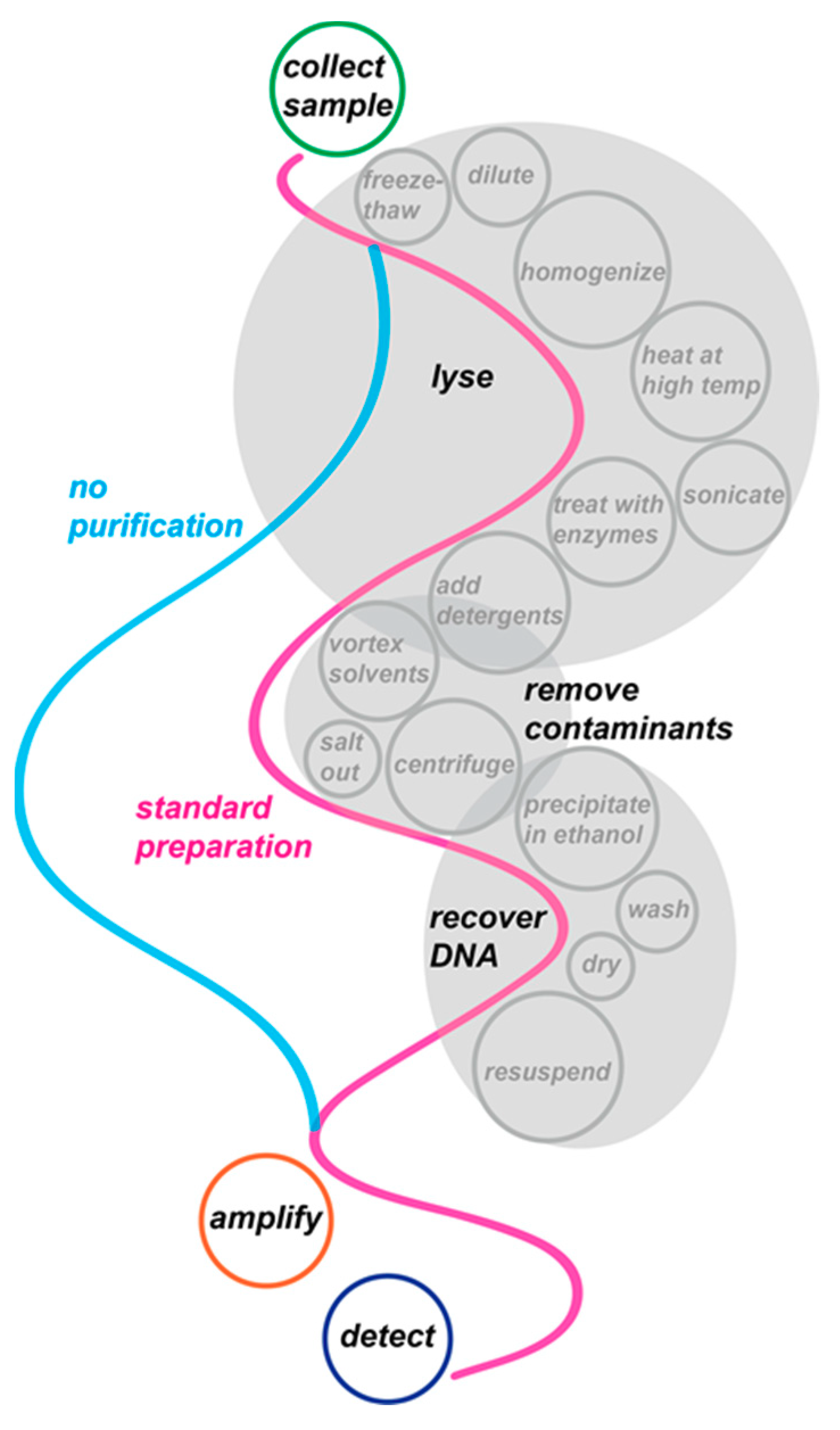
- Tolerance to the sample of interest—ideally, the assay should be able to detect its target against a high concentration of background contaminants. We note that, although sample dilution sometimes provides a convenient way of permitting amplification, doing so inevitably reduces the limit of detection (LOD). Most NAATs analyze only a fraction of the sample volume. Sample dilution therefore increases the likelihood of false negative results, especially when the samples already have low target concentrations.
- LOD—by foregoing sample preparation, one generally sacrifices the opportunity to concentrate bulk samples, reducing the limit of detection and making sensitivity an important consideration.
- Clinical evaluation—recognizing assays that have been validated with clinical samples.
3. Brief Overview of Isothermal Amplification Techniques
3.1. LAMP
3.2. NASBA
3.3. SDA
3.4. RPA
3.5. SIBA
3.6. MDA
3.7. HCR
4. Analysis of Surveyed Direct NAATs
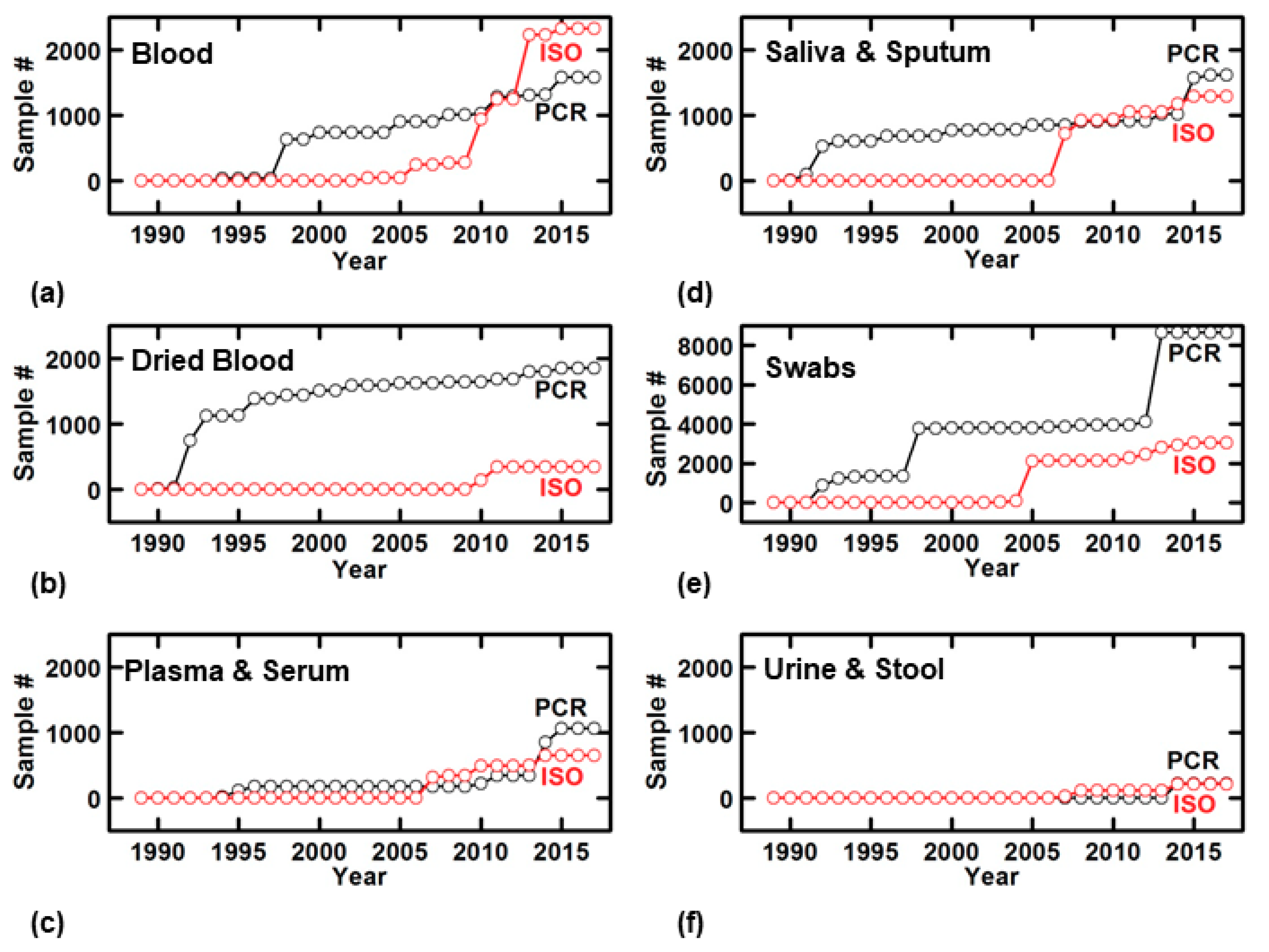
4.1. Growing Prevalence of Direct NAATs
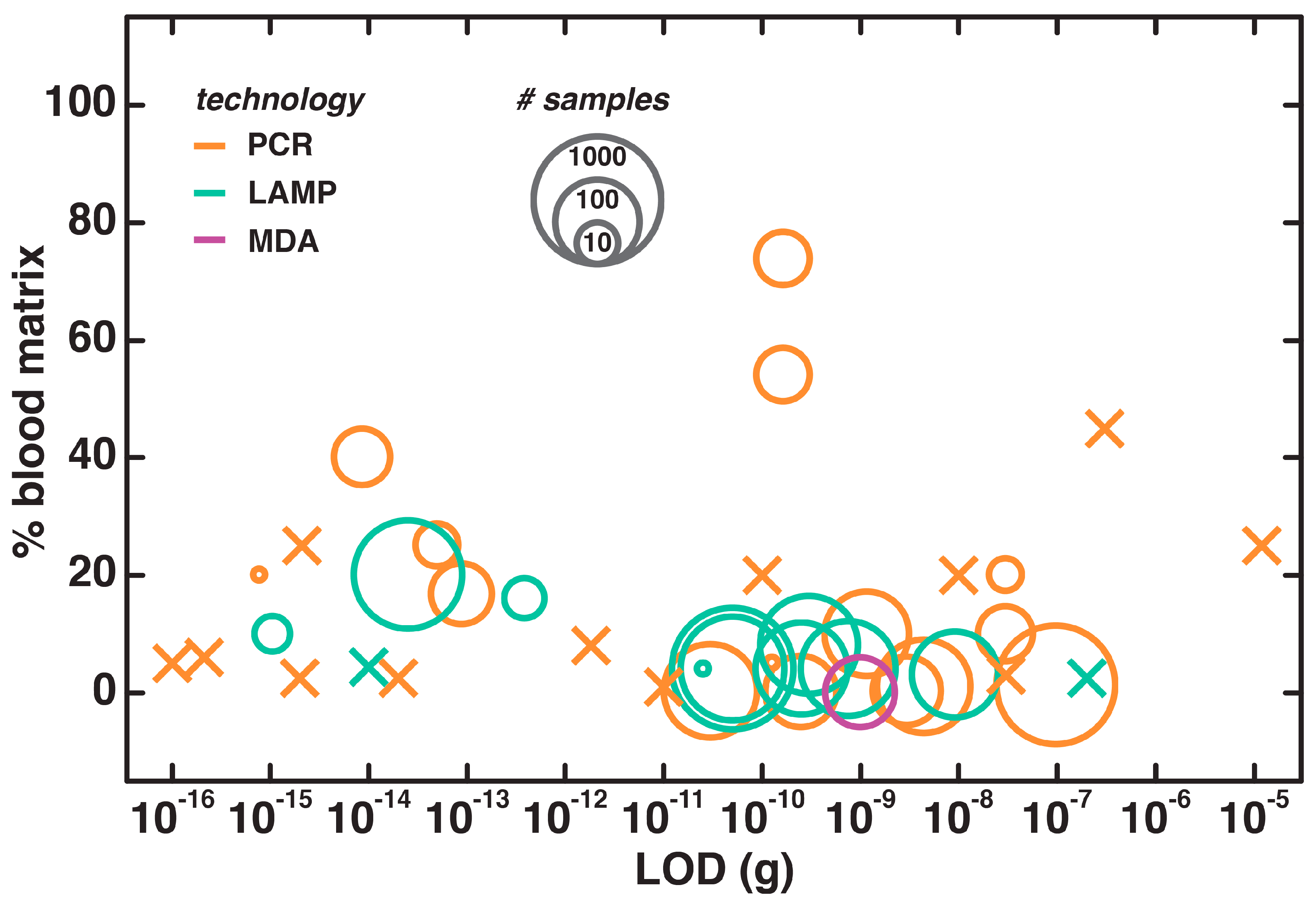
4.2. Direct NAATs for Whole Blood
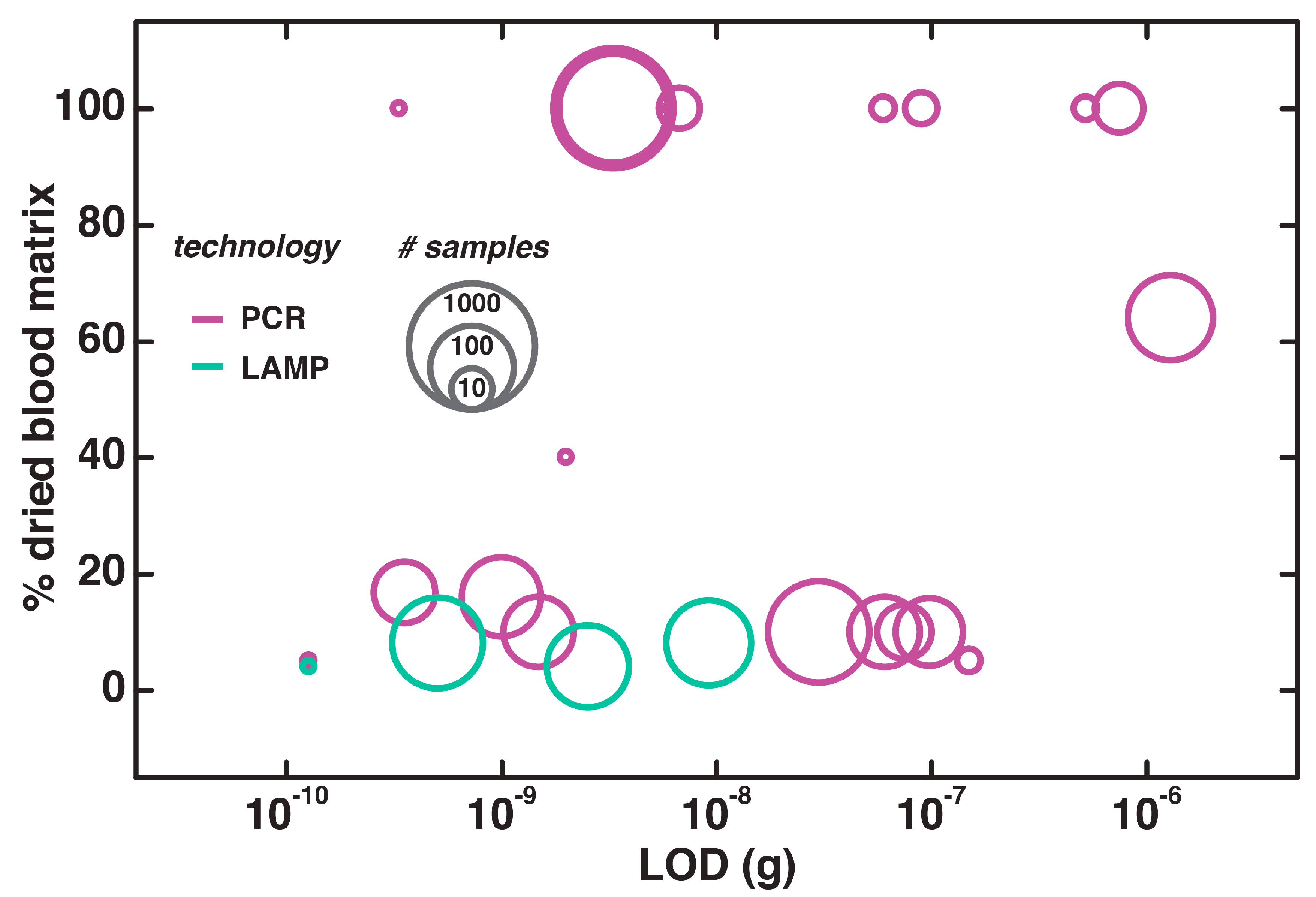
4.3. Direct NAATs for Dried Blood
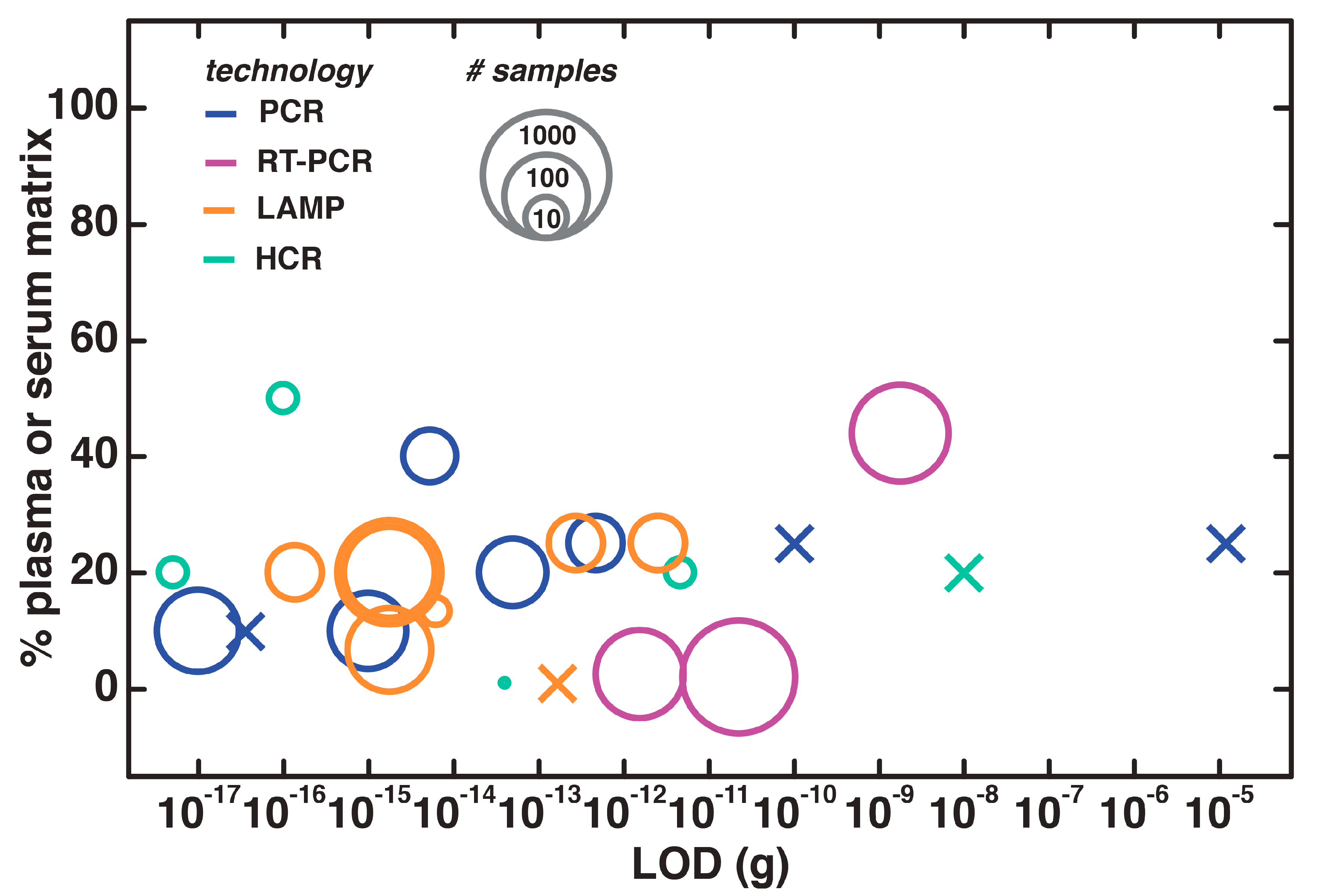
4.4. Direct NAATs for Plasma and Serum
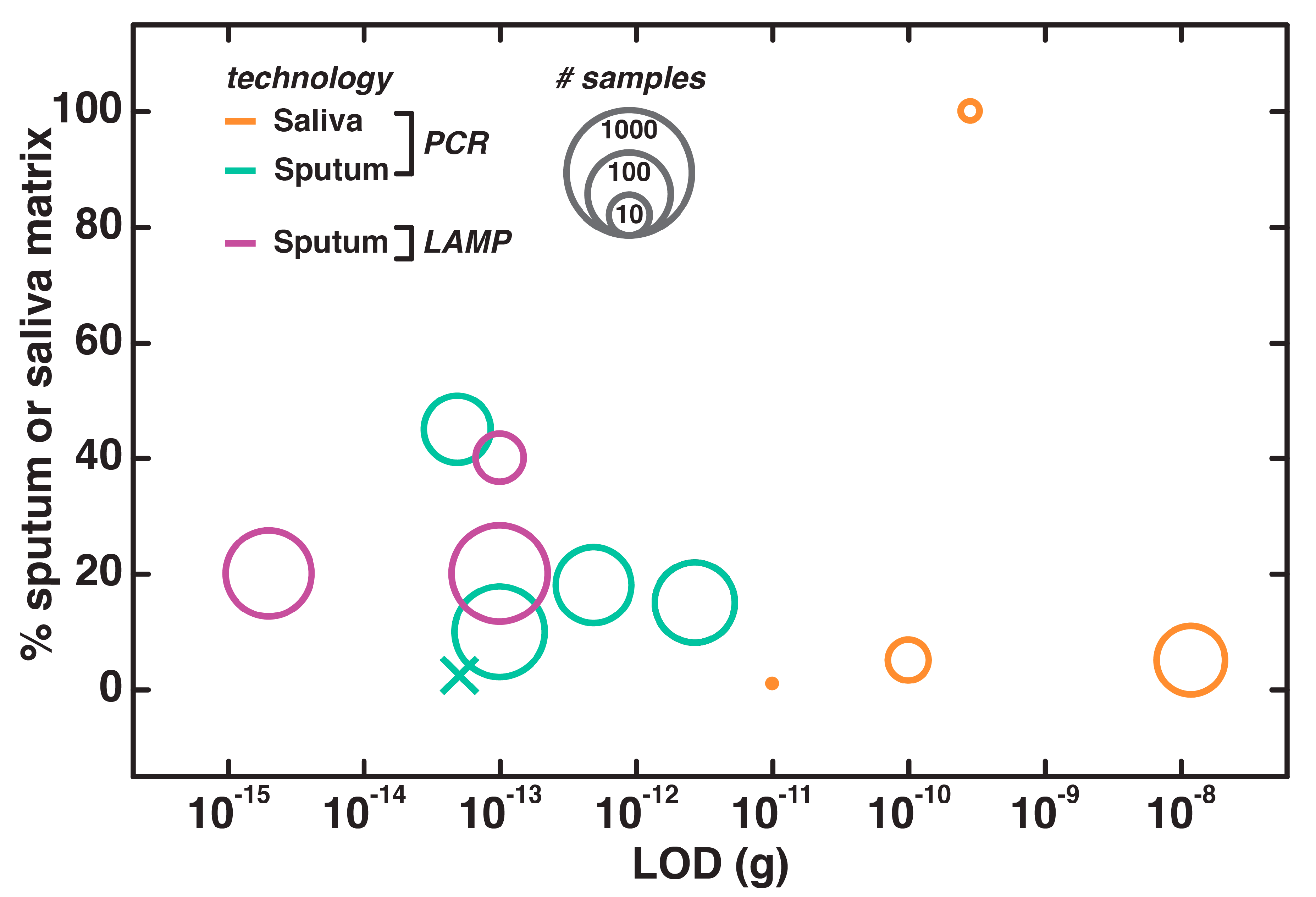
4.5. Direct NAATs for Saliva and Sputum
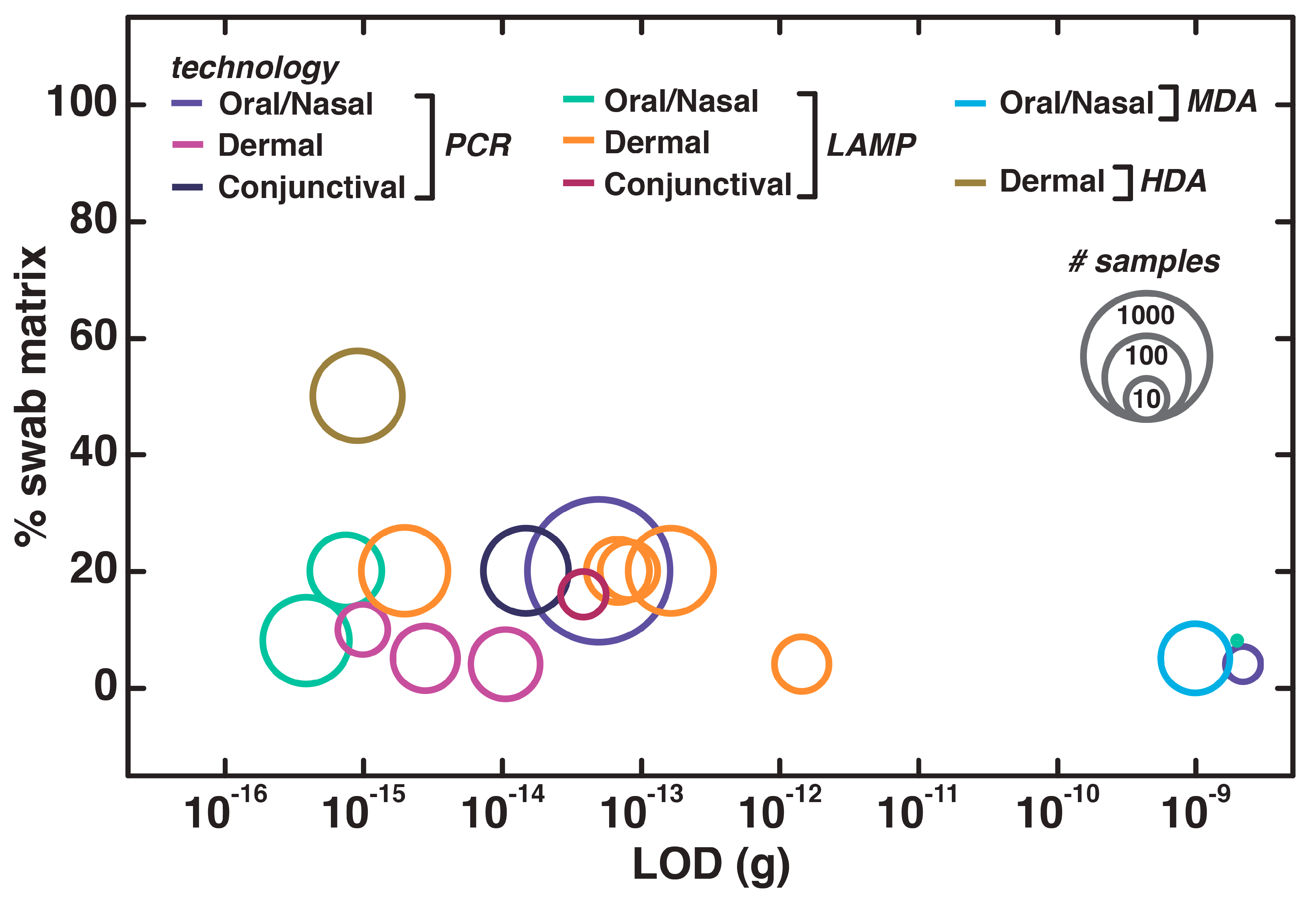
4.6. Direct NAATs for Oral, Dermal, and Conjunctival Swabs
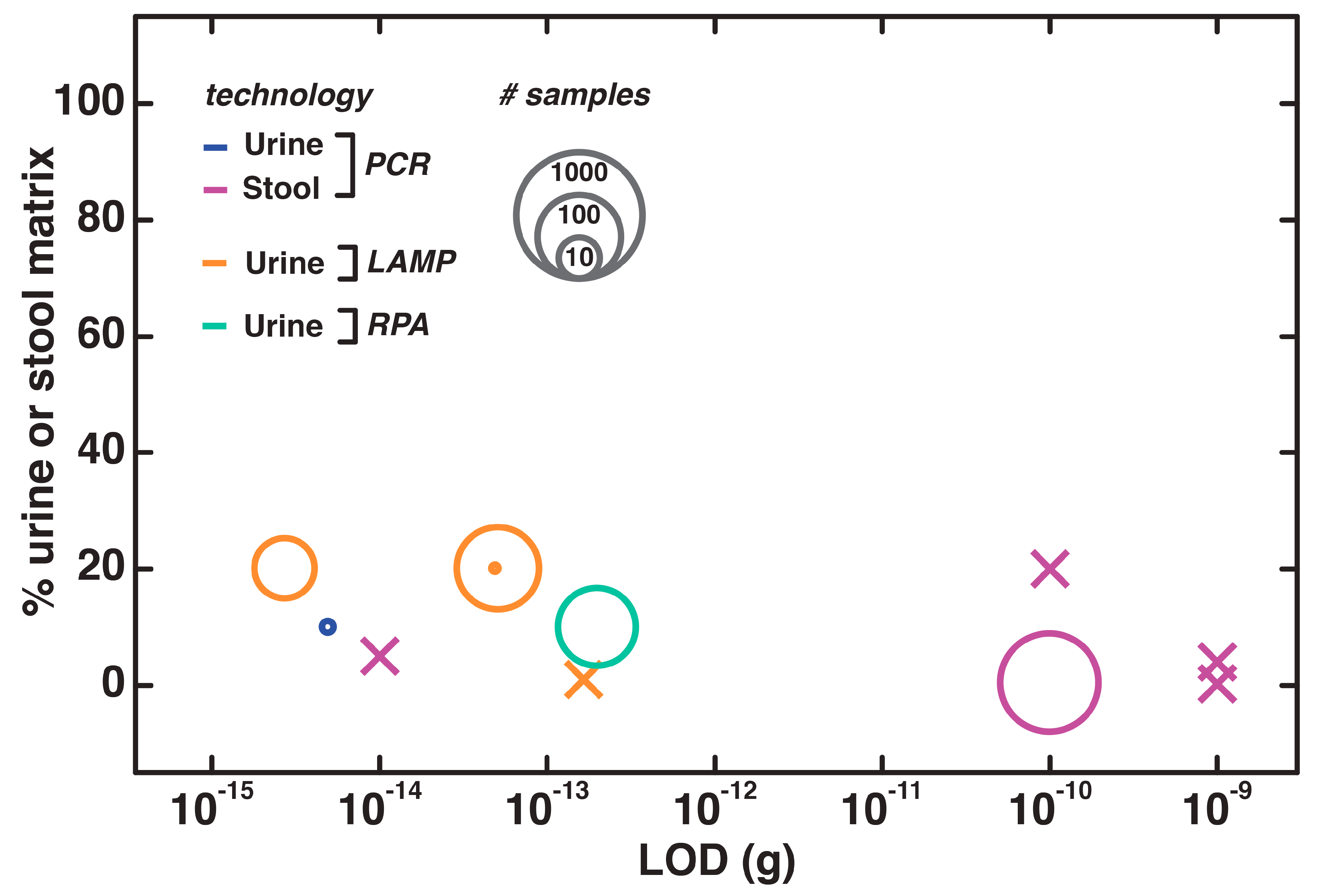
4.7. Direct NAATs for Urine and Stool
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gaydos, C.; Hardick, J. Point of care diagnostics for sexually transmitted infections: Perspectives and advances. Expert Rev. Anti. Infect. Ther. 2014, 12, 657–672. [Google Scholar] [CrossRef]
- Wang, S.; Xu, F.; Demirci, U. Advances in developing HIV-1 viral load assays for resource-limited settings. Biotechnol. Adv. 2010, 28, 770–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, C.D.; Linder, V.; Sia, S.K. Commercialization of microfluidic point-of-care diagnostic devices. Lab Chip 2012, 12, 2118–2134. [Google Scholar] [CrossRef] [PubMed]
- Kovarik, M.L.; Gach, P.C.; Ornoff, D.M.; Wang, Y.; Balowski, J.; Farrag, L.; Allbritton, N.L. Micro total analysis systems for cell biology and biochemical assays. Anal. Chem. 2012, 84, 516–540. [Google Scholar] [CrossRef]
- Zhang, Y.; Ozdemir, P. Microfluidic DNA amplification—A review. Anal. Chim. Acta 2009, 638, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.; Ghaemi, A. Nucleic acid isothermal amplification technologies: A review. Nucleosides Nucleotides Nucleic Acids 2008, 27, 224–243. [Google Scholar] [CrossRef] [PubMed]
- Niemz, A.; Ferguson, T.M.; Boyle, D.S. Point-of-care nucleic acid testing for infectious diseases. Trends Biotechnol. 2011, 29, 240–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craw, P.; Balachandran, W. Isothermal nucleic acid amplification technologies for point-of-care diagnostics: A critical review. Lab Chip 2012, 12, 2469–2486. [Google Scholar] [CrossRef]
- Dineva, M.A.; Mahilum-Tapay, L.; Lee, H. Sample preparation: A challenge in the development of point-of-care nucleic acid-based assays for resource-limited settings. Analyst 2007, 132, 1193–1199. [Google Scholar] [CrossRef]
- Schnepf, N.; Scieux, C.; Resche-Riggon, M.; Feghoul, L.; Xhaard, A.; Gallien, S.; Molina, J.M.; Socié, G.; Viglietti, D.; Simon, F.; et al. Fully automated quantification of cytomegalovirus (CMV) in whole blood with the new sensitive Abbott realTime CMV assay in the era of the CMV international standard. J. Clin. Microbiol. 2013, 51, 2096–2102. [Google Scholar] [CrossRef]
- Haliski, J. FDA Grants First CLIA Waiver for Nucleic Acid-Based Flu Diagnostic Test; Food and Drug Administration: Silver Spring, MD, USA, 2015. [Google Scholar]
- Bost, W. Focus Diagnostics Receives FDA Clearance for Moderate Complexity Simplexa HSV 1 & 2 Direct Molecular Test for Aiding the Diagnosis of Encephalitis; Quest Diagnostics: Madison, NJ, USA, 2014. [Google Scholar]
- Chapin, K.C.; Flores-Cortez, E.J. Performance of the Molecular Alere i Influenza A&B Test Compared to That of the Xpert Flu A/B Assay. J. Clin. Microbiol. 2015, 53, 706–709. [Google Scholar] [PubMed] [Green Version]
- Hazelton, B.; Gray, T.; Ho, J.; Ratnamohan, V.M.; Dwyer, D.E.; Kok, J. Detection of influenza A and B with the Alere TM i Influenza A&B: A novel isothermal nucleic acid amplification assay. Influenza Other Respi. Viruses 2015, 9, 151–154. [Google Scholar]
- Bell, J.J.; Selvarangan, R. Evaluation of the Alere i Influenza A&B Nucleic Acid Amplification Test by Use of Respiratory Specimens Collected in Viral Transport Medium. J. Clin. Microbiol. 2014, 52, 3992–3995. [Google Scholar] [PubMed] [Green Version]
- Nie, S.; Roth, R.B.; Stiles, J.; Mikhlina, A.; Lu, X.; Tang, Y.-W.; Babady, N.E. Evaluation of Alere i Influenza A&B for Rapid Detection of Influenza Viruses A and B. J. Clin. Microbiol. 2014, 52, 3339–3344. [Google Scholar] [PubMed] [Green Version]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell. Probes 2002, 16, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Thai, H.T.C.; Le, M.Q.; Vuong, C.D.; Parida, M.; Minekawa, H.; Notomi, T.; Hasebe, F.; Morita, K. Development and evaluation of a novel loop-mediated isothermal amplification method for rapid detection of severe acute respiratory syndrome coronavirus. J. Clin. Microbiol. 2004, 42, 1956–1961. [Google Scholar] [CrossRef] [PubMed]
- Compton, J. Nucleic Acid Sequence-Based Amplification. Nature 1991, 350, 91–92. [Google Scholar] [CrossRef]
- Pardee, K.; Green, A.A.; Takahashi, M.K.; Braff, D.; Lambert, G.; Lee, J.W.; Ferrante, T.; Ma, D.; Donghia, N.; Fan, M.; et al. Rapid, Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell 2016, 165, 1255–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verkooyen, R.P.; Noordhoek, G.T.; Klapper, P.E.; Schirm, J.; Cleator, G.M.; Ieven, M.; Hoddevik, G.; Reid, J. Reliability of Nucleic Acid Amplification Methods for Detection of Chlamydia trachomatis in Urine: Results of the First International Collaborative Quality Control Study among 96 Laboratories Reliability of Nucleic Acid Amplification Methods for Detectio. J. Clin. Microbiol. 2003, 41, 3013–3016. [Google Scholar] [CrossRef]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA detection using recombination proteins. PLoS Biol. 2006, 4, e204. [Google Scholar] [CrossRef] [PubMed]
- Hoser, M.J.; Mansukoski, H.K.; Morrical, S.W.; Eboigbodin, K.E. Strand Invasion Based Amplification (SIBA): A novel isothermal DNA amplification technology demonstrating high specificity and sensitivity for a single molecule of target analyte. PLoS ONE 2014, 9, e112656. [Google Scholar] [CrossRef] [PubMed]
- Eboigbodin, K.E.; Hoser, M.J. Multiplex Strand Invasion Based Amplification (mSIBA) assay for detection of Chlamydia trachomatis and Neisseria gonorrhoeae. Sci. Rep. 2016, 6, 20487. [Google Scholar] [CrossRef] [PubMed]
- Dean, F.B.; Hosono, S.; Fang, L.; Wu, X.; Faruqi, A.F.; Bray-Ward, P.; Sun, Z.; Zong, Q.; Du, Y.; Du, J.; et al. Comprehensive human genome amplification using multiple displacement amplification. Proc. Natl. Acad. Sci. USA 2002, 99, 5261–5266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirks, R.M.; Pierce, N. a Triggered amplification by hybridization chain reaction. Proc. Natl. Acad. Sci. USA 2004, 101, 15275–15278. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ge, L.; Yu, Y.; Dong, S.; Li, F. Highly sensitive electrogenerated chemiluminescence biosensor based on hybridization chain reaction and amplification of gold nanoparticles for DNA detection. Sens. Actuators B Chem. 2015, 220, 942–948. [Google Scholar] [CrossRef]
- Mullis, K.; Faloona, F.; Scharf, S. Specific enzymatic amplification of DNA in vitro: The polymerase chain reaction. Cold Spring Harb. 1986, 51, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Pavšič, J.; Devonshire, A.S.; Parkes, H.; Schimmel, H.; Foy, C.A.; Karczmarczyk, M.; Gutiérrez-Aguirre, I.; Honeyborne, I.; Huggett, J.F.; McHugh, T.D.; et al. Standardization of nucleic acid tests for clinical measurements of bacteria and viruses. J. Clin. Microbiol. 2015, 53, 2008–2014. [Google Scholar] [CrossRef]
- Banada, P.P.; Chakravorty, S.; Shah, D.; Burday, M.; Mazzella, F.M.; Alland, D. Highly sensitive detection of staphylococcus aureus directly from patient blood. PLoS ONE 2012, 7, e31126. [Google Scholar] [CrossRef]
- De Franchis, R.; Cross, N.C.P.; Foulkes, N.S.; Cox, T.M. A potent inhibitor of Taq polymerase copurifies with human genomic DNA. Nucleic Acids Res. 1988, 16, 10355. [Google Scholar] [CrossRef]
- Akane, A.; Matsubara, K.; Nakamura, H.; Takahashi, S.; Kimura, K. Identification of the Heme Compound Copurified with Deoxyribonucleic Acid (DNA) from Bloodstains, a Major Inhibitor of Polymerase Chain Reaction (PCR) Amplification. J. Forensic Sci. 1994, 39, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Rossen, L.; Norskov, P.; Hoimstrom, K.; Rasmussen, O.F. Inhibition of PCR by components of food samples, microbial diagnostic assays and DNA-extraction solutions. Int. J. Food Microbiol. 1992, 17, 37–45. [Google Scholar] [CrossRef]
- Al-soud, W.A.; Rådström, P. Purification and Characterization of PCR-Inhibitory Components in Blood Cells. J. Clin. Microbiol. 2001, 39, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burckhardt, J. Amplification of DNA from whole blood. PCR Methods Appl. 1994, 3, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Morton, B.A.; Diniz, P.P.V.D.P. Detection of Bartonella quintana DNA in the presence of human and feline whole blood by single-tube PCR without DNA extraction. Microbiol. Discov. 2013, 1, 7. [Google Scholar] [CrossRef]
- Nishimura, N.; Nakayama, T.; Tonoike, H.; Kojima, K.; Kato, S. Direct polymerase chain reaction from whole blood without DNA isolation. Ann. Clin. Biochem. 2000, 37, 674–680. [Google Scholar] [CrossRef]
- Yang, Y.G.; Kim, J.Y.; Song, Y.H.; Kim, D.S. A novel buffer system, AnyDirect, can improve polymerase chain reaction from whole blood without DNA isolation. Clin. Chim. Acta 2007, 380, 112–117. [Google Scholar] [CrossRef]
- Walker, F.M.; Ahmad, K.M.; Eisenstein, M.; Soh, H.T. Transformation of Personal Computers and Mobile Phones into Genetic Diagnostic Systems. Anal. Chem. 2014, 86, 9236–9241. [Google Scholar] [CrossRef] [Green Version]
- Castley, A.; Higgins, M.; Ivey, J.; Mamotte, C.; Sayer, D.C.; Christiansen, F.T. Clinical applications of whole-blood PCR with real-time instrumentation. Clin. Chem. 2005, 51, 2025–2030. [Google Scholar] [CrossRef]
- Chomczynski, P.; Rymaszewski, M. Alkaline polyethylene glycol-based method for direct PCR from bacteria, eukaryotic tissue samples, and whole blood. Biotechniques 2004, 40, 454–458. [Google Scholar] [CrossRef]
- Klintschar, M.; Neuhuber, F. Evaluation of an alkaline lysis method for the extraction of DNA from whole blood and forensic stains for STR analysis. J. Forensic Sci. 2000, 45, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Rudbeck, L.; Dissing, J. Rapid, simple alkaline extraction of human genomic DNA from whole blood, buccal epithelial cells, semen and forensic stains for PCR. Biotechniques 1998, 25, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Manage, D.P.; Lauzon, J.; Atrazhev, A.; Pang, X.; Pilarski, L.M. A novel method for sample delivery and testing of whole blood: Gel strip PCR for point of care (POC) molecular diagnostics. Lab Chip 2013, 13, 4011–4014. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Virdi, A.S.; Singh, P. A novel method for whole blood PCR without pretreatment. Gene 2012, 501, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.J.; Martin, K.A.; Arango, E.; Agudelo, O.M.; Maestre, A.; Yanow, S.K. Real-time PCR detection of Plasmodium directly from whole blood and filter paper samples. Malar. J. 2011, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.; Leathart, J.B.S.; French, D.J.; Krishan, E.; Kohnke, H.; Wadelius, M.; Van Schie, R.; Verhoef, T.; Maitland-van der Zee, A.H.; Daly, A.K.; et al. Genotyping for CYP2C9 and VKORC1 alleles by a novel point of care assay with HyBeacon® probes. Clin. Chim. Acta 2011, 412, 2063–2069. [Google Scholar] [CrossRef]
- Zhang, Z.; Kermekchiev, M.B.; Barnes, W.M. Direct DNA Amplification from Crude Clinical Samples Using a PCR Enhancer Cocktail and Novel Mutants of Taq. J. Mol. Diagn. 2010, 12, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Kermekchiev, M.B.; Kirilova, L.I.; Vail, E.E.; Barnes, W.M. Mutants of Taq DNA polymerase resistant to PCR inhibitors allow DNA amplification from whole blood and crude soil samples. Nucleic Acids Res. 2009, 37, e40. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, J.Y.; Yang, Y.G.; Lee, S.H. Direct STR amplification from whole blood and blood- or saliva-spotted FTA without DNA purification. J. Forensic Sci. 2008, 53, 335–341. [Google Scholar] [CrossRef]
- Bu, Y.; Huang, H.; Zhou, G. Direct polymerase chain reaction (PCR) from human whole blood and filter-paper-dried blood by using a PCR buffer with a higher pH. Anal. Biochem. 2008, 375, 370–372. [Google Scholar] [CrossRef]
- Silvy, M.; Bres, J.C.; Grimaldi, A.; Movia, C.; Muriel, V.; Roubinet, F.; Chiaroni, J.; Bailly, P. A simple genotyping procedure without DNA extraction to identify rare blood donors. Vox Sang. 2015, 109, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.-P.; Wang, Y.; He, X.-H.; Guan, J.-J.; Wang, J.; Qin, Z.-H.; Sun, W.-P. Whole Blood PCR Amplification with Pfu DNA Polymerase and Its Application in Single-Nucleotide Polymorphism Analysis. Genet. Test. Mol. Biomark. 2015, 19, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Minogue, T.D.; Rachwal, P.A.; Trombley Hall, A.; Koehler, J.W.; Weller, S.A. Cross-institute evaluations of inhibitor-resistant PCR reagents for direct testing of aerosol and blood samples containing biological warfare agent DNA. Appl. Environ. Microbiol. 2014, 80, 1322–1329. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.T.; Zovanyi, A.M.; Christensen, D.R.; Koehler, J.W.; Minogue, T.D. Evaluation of inhibitor-resistant real-time PCR methods for diagnostics in clinical and environmental samples. PLoS ONE 2013, 8, e73845. [Google Scholar]
- Al-soud, W.A.; Rådström, P. Capacity of Nine Thermostable DNA Polymerases To Mediate DNA Amplification in the Presence of PCR-Inhibiting Samples. Appl. Environ. Microbiol. 1998, 64, 3748–3753. [Google Scholar] [PubMed]
- Panaccio, M.; Lew, A. PCR based diagnosis in the presence of 8% (v/v) blood. Nucleic Acids Res. 1991, 19, 1151. [Google Scholar] [CrossRef]
- Port, J.R.; Nguetse, C.; Adukpo, S.; Velavan, T.P. A reliable and rapid method for molecular detection of malarial parasites using microwave irradiation and loop mediated isothermal amplification. Malar. J. 2014, 13, 454. [Google Scholar] [CrossRef] [PubMed]
- Meganathan, P.; Singh, S.; Ling, L.Y.; Singh, J.; Subrayan, V.; Nissapatorn, V. Detection of Toxoplasma gondii DNA by PCR following microwave treatment of serum and whole blood. Southeast Asian J. Trop. Med. Public Health 2010, 41, 265–273. [Google Scholar]
- El-Awady, M.K.; El-Hosseiny, L.A.; Ismail, S.M.; Abdel-Aziz, M.T.; El-Demellawy, M.A. Comparison between Toxoplasma gondii DNA and specific immunoglobulins during pregnancy. La Rev. Sante la Mediterr. Orient. 2000, 6, 888–897. [Google Scholar]
- Jadaon, M.M.; Dashti, A.A.; Lewis, H.L.; Habeeb, F.M. Whole-blood polymerase chain reaction and restriction fragment length polymorphism: A simplified method by microwave irradiation. Med. Princ. Pract. 2009, 18, 280–283. [Google Scholar] [CrossRef]
- McCusker, J.; Dawson, M.T.; Noone, D.; Gannon, F.; Smith, T. Improved method for direct PCR amplification from whole blood. Nucleic Acids Res. 1992, 20, 6747. [Google Scholar] [CrossRef] [PubMed]
- Mercier, B.; Gaucher, C.; Feugeas, O.; Mazurier, C. Direct PCR from whole blood, without DNA extraction. Nucleic Acids Res. 1990, 18, 5908. [Google Scholar] [CrossRef] [PubMed]
- Modak, S.S.; Barber, C.A.; Geva, E.; Abrams, W.R.; Malamud, D.; Ongagna, Y.S.Y. Rapid Point-of-Care Isothermal Amplification Assay for the Detection of Malaria without Nucleic Acid Purification. Infect. Dis. 2016, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Paris, D.H.; Blacksell, S.D.; Newton, P.N.; Day, N.P.J. Simple, rapid and sensitive detection of Orientia tsutsugamushi by loop-isothermal DNA amplification. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 1239–1246. [Google Scholar] [CrossRef]
- Njiru, Z.K.; Mikosza, A.S.J.; Matovu, E.; Enyaru, J.C.K.; Ouma, J.O.; Kibona, S.N.; Thompson, R.C.A.; Ndung’u, J.M. African trypanosomiasis: Sensitive and rapid detection of the sub-genus Trypanozoon by loop-mediated isothermal amplification (LAMP) of parasite DNA. Int. J. Parasitol. 2008, 38, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Poon, L.L.M.; Wong, B.W.Y.; Ma, E.H.T.; Chan, K.H.; Chow, L.M.C.; Abeyewickreme, W.; Tangpukdee, N.; Yuen, K.Y.; Guan, Y.; Looareesuwan, S.; et al. Sensitive and inexpensive molecular test for falciparum malaria: Detecting Plasmodium falciparum DNA directly from heat-treated blood by loop-mediated isothermal amplification. Clin. Chem. 2006, 52, 300–303. [Google Scholar] [CrossRef]
- Britton, S.; Cheng, Q.; Sutherland, C.J.; McCarthy, J.S. A simple, high-throughput, colourimetric, field applicable loop-mediated isothermal amplification (HtLAMP) assay for malaria elimination. Malar. J. 2015, 14, 335. [Google Scholar] [CrossRef]
- Hopkins, H.; González, I.J.; Polley, S.D.; Angutoko, P.; Ategeka, J.; Asiimwe, C.; Agaba, B.; Kyabayinze, D.J.; Sutherland, C.J.; Perkins, M.D.; et al. Highly sensitive detection of malaria parasitemia in a malaria-endemic setting: Performance of a new loop-mediated isothermal amplification kit in a remote clinic in Uganda. J. Infect. Dis. 2013, 208, 645–652. [Google Scholar] [CrossRef]
- Polley, S.D.; González, I.J.; Mohamed, D.; Daly, R.; Bowers, K.; Watson, J.; Mewse, E.; Armstrong, M.; Gray, C.; Perkins, M.D.; et al. Clinical evaluation of a loop-mediated amplification kit for diagnosis of imported malaria. J. Infect. Dis. 2013, 208, 637–644. [Google Scholar] [CrossRef]
- Sirichaisinthop, J.; Buates, S.; Watanabe, R.; Han, E.T.; Suktawonjaroenpon, W.; Krasaesub, S.; Takeo, S.; Tsuboi, T.; Sattabongkot, J. Short report: Evaluation of loop-mediated isothermal amplification (LAMP) for malaria diagnosis in a field setting. Am. J. Trop. Med. Hyg. 2011, 85, 594–596. [Google Scholar] [CrossRef]
- Soejima, M.; Egashira, K.; Kawano, H.; Kawaguchi, A.; Sagawa, K.; Koda, Y. Rapid detection of haptoglobin gene deletion in alkaline-denatured blood by loop-mediated isothermal amplification reaction. J. Mol. Diagn. 2011, 13, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Polley, S.D.; Mori, Y.; Watson, J.; Perkins, M.D.; González, I.J.; Notomi, T.; Chiodini, P.L.; Sutherland, C.J. Mitochondrial DNA targets increase sensitivity of malaria detection using loop-mediated isothermal amplification. J. Clin. Microbiol. 2010, 48, 2866–2871. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Lu, F.; Lim, C.S.; Kim, J.Y.; Ahn, H.J.; Suh, I.B.; Takeo, S.; Tsuboi, T.; Sattabongkot, J.; Han, E.T. Detection of Plasmodium vivax infection in the Republic of Korea by loop-mediated isothermal amplification (LAMP). Acta Trop. 2010, 113, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Ihira, M.; Enomoto, Y.; Yano, H.; Maruyama, F.; Emi, N.; Asano, Y.; Yoshikawa, T. Heat denaturation increases the sensitivity of the cytomegalovirus loop-mediated isothermal amplification method. Microbiol. Immunol. 2010, 54, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Benzine, J.W.; Brown, K.M.; Agans, K.N.; Godiska, R.; Mire, C.E.; Gowda, K.; Converse, B.; Geisbert, T.W.; Mead, D.A.; Chander, Y. Molecular Diagnostic Field Test for Point-of-Care Detection of Ebola Virus Directly from Blood. J. Infect. Dis. 2016, 214, S234–S242. [Google Scholar] [CrossRef] [PubMed]
- Damhorst, G.L.; Duarte-Guevara, C.; Chen, W.; Ghonge, T.; Cunningham, B.T.; Bashir, R. Smartphone-Imaged HIV-1 Reverse-Transcription Loop-Mediated Isothermal Amplification (RT-LAMP) on a Chip from Whole Blood. Engineering 2015, 1, 324–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashida, K.; Kajino, K.; Hachaambwa, L.; Namangala, B.; Sugimoto, C. Direct Blood Dry LAMP: A Rapid, Stable, and Easy Diagnostic Tool for Human African Trypanosomiasis. PLoS Negl. Trop. Dis. 2015, 9, e0003578. [Google Scholar] [CrossRef] [PubMed]
- Ebbinghaus, P.; von Samson-Himmelstjerna, G.; Krücken, J. Direct loop-mediated isothermal amplification from Plasmodium chabaudi infected blood samples: Inability to discriminate genomic and cDNA sequences. Exp. Parasitol. 2012, 131, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Curtis, K.A.; Rudolph, D.L.; Owen, S.M. Sequence-specific detection method for reverse transcription, loop-mediated isothermal amplification of HIV-1. J. Med. Virol. 2009, 81, 966–972. [Google Scholar] [CrossRef]
- Priye, A.; Bird, S.W.; Light, Y.K.; Ball, C.S.; Negrete, O.A.; Meagher, R.J. A smartphone-based diagnostic platform for rapid detection of Zika, chikungunya, and dengue viruses. Sci. Rep. 2017, 7, 44778. [Google Scholar] [CrossRef] [PubMed]
- Ball, C.S.; Light, Y.K.; Koh, C.Y.; Wheeler, S.S.; Coffey, L.L.; Meagher, R.J. Quenching of Unincorporated Amplification Signal Reporters in Reverse-Transcription Loop-Mediated Isothermal Amplification Enabling Bright, Single-Step, Closed-Tube, and Multiplexed Detection of RNA Viruses. Anal. Chem. 2016, 88, 3562–3568. [Google Scholar] [CrossRef] [PubMed]
- Hosono, S.; Faruqi, A.F.; Dean, F.B.; Du, Y.; Sun, Z.; Wu, X.; Du, J.; Kingsmore, S.F.; Egholm, M.; Lasken, R.S. Unbiased whole-genome amplification directly from clinical samples. Genome Res. 2003, 13, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Johannessen, A. Dried blood spots in HIV monitoring: Applications in resource-limited settings. Bioanalysis 2010, 2, 1893–1908. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, L.; Jiang, Y.; Fang, L.; Pan, P.; Gong, S.; Yao, J.; Tang, Y.W.; Vermund, S.H.; Jia, Y. Early infant human immunodeficiency virus type 1 detection suitable for resource-limited settings with multiple circulating subtypes by use of nested three-monoplex DNA PCR and dried blood spots. J. Clin. Microbiol. 2008, 46, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Smit, P.W.; Elliott, I.; Peeling, R.W.; Mabey, D.; Newton, P.N. Review article: An overview of the clinical use of filter paper in the diagnosis of tropical diseases. Am. J. Trop. Med. Hyg. 2014, 90, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Chediak, A. The diagnosis of syphilis on a dessicated and defibrinated blood drop. Rev. Med. Cubana 1932, 953–956. [Google Scholar]
- Guthrie, R.; Susi, A. A simple phenylalanine method for detecting phenylketonuria in large populations of newborn infants. Pediatrics 1963, 32, 338–343. [Google Scholar]
- Bertagnolio, S.; Parkin, N.T.; Jordan, M.; Brooks, J.; G, G.-L.J. Dried blood spots for HIV-1 drug resistance and viral load testing: A review of current knowledge and WHO efforts for global HIV drug resistance surveillance. AIDS Rev. 2010, 12, 195–208. [Google Scholar]
- Solomon, A.W.; Engels, D.; Bailey, R.L.; Blake, I.M.; Brooker, S.; Chen, J.X.; Chen, J.H.; Churcher, T.S.; Drakeley, C.J.; Edwards, T.; et al. A diagnostics platform for the integrated mapping, monitoring, and surveillance of neglected tropical diseases: Rationale and target product profiles. PLoS Negl. Trop. Dis. 2012, 6, e1746. [Google Scholar] [CrossRef]
- Raskin, S.; Iii, J.A.P.; Kaplan, G.; Mcclure, M.; Vnencak-Jones, C. Cystic Fibrosis Genotyping by Direct PCR Analysis of Guthrle Blood Spots. Genome Res. 1992, 2, 154–156. [Google Scholar] [CrossRef]
- Hall, D.E.; Roy, R. An evaluation of direct PCR amplification. Croat. Med. J. 2014, 55, 655–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makowski, G.S.; Davis, E.L.; Aslanzadeh, J.; Hopfer, S.M. Enhanced direct amplification of Guthrie card DNA following selective elution of PCR inhibitors. Nucleic Acids Res. 1995, 23, 3788–3789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raskin, S.; Phillips, J.A.; Krishnamani, M.R.; Vnencak-Jones, C.; Parker, R.A.; Rozov, T.; Cardieri, J.M.; Marostica, P.; Abreu, F.; Giugliani, R. DNA analysis of cystic fibrosis in Brazil by direct PCR amplification from Guthrie cards. Am. J. Med. Genet. 1993, 46, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Seltzer, W.K.; Accurso, F.; Fall, M.Z.; VanRiper, A.J.; Descartes, M.; Huang, Y.; McCabe, E.R. Screening for cystic fibrosis: Feasibility of molecular genetic analysis of dried blood specimens. Biochem. Med. Metab. Biol. 1991, 46, 105–109. [Google Scholar] [CrossRef]
- Schwartz, E.I.; Khalchitsky, S.E.; Eisensmith, R.C.; Woo, S.L.C. Polymerase chain reaction amplification from dried blood spots on Guthrie cards. Lancet 1990, 336, 639–640. [Google Scholar] [CrossRef]
- Caggana, M.; Conroy, J.M.; Pass, K.A. Rapid, efficient method for multiplex amplification from filter paper. Hum. Mutat. 1998, 11, 404–409. [Google Scholar] [CrossRef]
- Nelson, P.V.; Carey, W.F.; Morris, C.P. Gene amplification directly from Guthrie blood spots. Lancet 1990, 336, 1451–1452. [Google Scholar] [CrossRef]
- Jinks, D.C.; Minter, M.; Tarver, D.A.; Vanderford, M.; Hejtmancik, J.F.; McCabe, E.R. Molecular genetic diagnosis of sickle cell disease using dried blood specimens on blotters used for newborn screening. Hum. Genet. 1989, 81, 363–366. [Google Scholar] [CrossRef]
- Mackey, K.; Steinkamp, A.; Chomczynski, P. DNA Extraction from Small Blood Volumes and the Processing of Cellulose Blood Cards for Use in Polymerase Chain Reaction. Mol. Biotechnol. 1998, 9, 1–5. [Google Scholar] [CrossRef]
- Ramos-Díaz, R.; Gutiérrez-Nicolás, F.; Nazco-Casariego, G.J.; González-Perera, I.; Pérez-Pérez, J.A. Validation of a fast and low-cost alkaline lysis method for gDNA extraction in a pharmacogenetic context. Cancer Chemother. Pharmacol. 2015, 75, 1095–1098. [Google Scholar] [CrossRef]
- Song, Y.; Fahs, A.; Feldman, C.; Shah, S.; Gu, Y.; Wang, Y.; Machado, R.F.; Wunderink, R.G.; Chen, J. A reliable and effective method of DNA isolation from old human blood paper cards. Springerplus 2013, 2, 616. [Google Scholar] [CrossRef] [PubMed]
- Miura, M.; Tanigawa, C.; Fujii, Y.; Kaneko, S. Comparison of six commercially-available DNA polymerases for direct PCR. Rev. Inst. Med. Trop. Sao Paulo 2013, 55, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Bereczky, S.; Mårtensson, A.; Gil, J.P.; Farnert, A. Short Report: Rapid DNA Extraction from Archive Blood Spots on Filter Paper for Genotyping of Plasmodium Falciparum. Am. J. Trop. Med. Hyg. 2005, 72, 249–251. [Google Scholar] [CrossRef] [PubMed]
- Kline, M.C.; Duewer, D.L.; Redman, J.W.; Butler, J.M.; Boyer, D.A. Polymerase chain reaction amplification of DNA from aged blood stains: Quantitative evaluation of the “suitability for purpose” of four filter papers as archival media. Anal. Chem. 2002, 74, 1863–1869. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Hendrie, H.C.; Hall, K.; Oluwole, O.S.A.; Hodes, M.E.; Sabota, A. Improved Procedure for Eluting DNA from Dried Blood Spots. Clin. Chem. 1996, 42, 1115–1116. [Google Scholar] [PubMed]
- Tao, Z.; Zhou, H.; Xia, H.; Xu, S.; Zhu, H.; Culleton, R.L.; Han, E.; Lu, F. Adaptation of a visualized loop-mediated isothermal amplification technique for field detection of Plasmodium vivax infection. Parasit. Vectors 2011, 4, 115. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Huang, Y.Y.; Liu, X.; Zhang, X.; Ferrari, M.; Qin, L. Point-of-care technologies for molecular diagnostics using a drop of blood. Trends Biotechnol. 2014, 32, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maton, A. Human Biology and Health; Prentice Hall Science; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 1993; ISBN 9780139811760. [Google Scholar]
- Caliendo, A.M.; Valsamakis, A.; Bremer, J.W.; Ferreira-Gonzalez, A.; Granger, S.; Sabatini, L.; Tsongalis, G.J.; Wang, Y.F.W.; Yen-Lieberman, B.; Young, S.; et al. Multilaboratory evaluation of real-time PCR tests for hepatitis B virus DNA quantification. J. Clin. Microbiol. 2011, 49, 2854–2858. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; Hoon, D.S.B.; Pantel, K. Cell-free nucleic acids as biomarkers in cancer patients. Nat. Rev. Cancer 2011, 11, 426–437. [Google Scholar] [CrossRef]
- Suzuki, N.; Kamataki, A.; Yamaki, J.; Homma, Y. Characterization of circulating DNA in healthy human plasma. Clin. Chim. Acta 2008, 387, 55–58. [Google Scholar] [CrossRef]
- Stroun, M.; Anker, P.; Lyautey, J.; Lederrey, C.; Maurice, P.A. Isolation and Characterization of DNA from the Plasma of Cancer Patients. Eur. J. Cancer Clin. Oncol. 1987, 23, 707–712. [Google Scholar] [CrossRef]
- Songjaroen, T.; Dungchai, W.; Chailapakul, O.; Henry, C.S.; Laiwattanapaisal, W. Blood separation on microfluidic paper-based analytical devices. Lab Chip 2012, 12, 3392–3398. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Woenker, T.; Adamec, J.; Regnier, F.E. Simple, miniaturized blood plasma extraction method. Anal. Chem. 2013, 85, 11501–11508. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Mauk, M.; Gross, R.; Bushman, F.D.; Edelstein, P.H.; Collman, R.G.; Bau, H.H. Membrane-based, sedimentation-assisted plasma separator for point-of-care applications. Anal. Chem. 2013, 85, 10463–10470. [Google Scholar] [CrossRef] [PubMed]
- Kersaudy-Kerhoas, M.; Sollier, E. Micro-scale blood plasma separation: From acoustophoresis to egg-beaters. Lab Chip 2013, 13, 3323–3346. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, Y.; Ming, J.; Lin, Y.; Cheng, C.; Li, F. Enzyme-free and ultrasensitive electrochemical detection of nucleic acids by target catalyzed hairpin assembly followed with hybridization chain reaction. Biosens. Bioelectron. 2013, 49, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Nixon, G.; Garson, J.A.; Grant, P.; Nastouli, E.; Foy, C.A.; Huggett, J.F. Comparative Study of Sensitivity, Linearity, and Resistance to Inhibition of Digital and Nondigital Polymerase Chain Reaction and Loop Mediated Isothermal Amplification Assays for Quantification of Human Cytomegalovirus. Anal. Chem. 2014, 86, 4387–4394. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, X.; Zhang, Y.; Liu, X.; Zheng, G.; Yang, Y.; Wang, L.; Du, L.; Wang, C. Direct Serum Assay for Cell-Free Bmi-1 mRNA and Its Potential Diagnostic and Prognostic Value for Colorectal Cancer. Clin. Cancer Res. 2014, 21, 1225–1233. [Google Scholar] [CrossRef]
- Asaga, S.; Kuo, C.; Nguyen, T.; Terpenning, M.; Giuliano, A.E.; Hoon, D.S.B. Direct serum assay for microRNA-21 concentrations in early and advanced breast cancer. Clin. Chem. 2011, 57, 84–91. [Google Scholar] [CrossRef]
- Zhang, L.; Song, X.; Wang, X.; Xie, Y.; Wang, Z.; Xu, Y.; You, X.; Liang, Z.; Cao, H. Circulating DNA of HOTAIR in serum is a novel biomarker for breast cancer. Breast Cancer Res. Treat. 2015, 152, 199–208. [Google Scholar] [CrossRef]
- Jang, J.S.; Lee, K.-J. Simple and rapid identification of low level hepatitis B virus DNA by the nested polymerase chain reaction. Arch. Pharm. Res. 1996, 19, 469–474. [Google Scholar] [CrossRef]
- Costa, J.; López-Labrador, F.X.; Sánchez-Tapias, J.M.; Mas, A.; Vilella, A.; Olmedo, E.; Vidal, J.; Jimenez de Anta, M.T.; Rodés, J. Microwave treatment of serum facilitates detection of hepatitis B virus DNA by the polymerase chain reaction. Results of a study in anti-HBe positive chronic hepatitis B. J. Hepatol. 1995, 22, 35–42. [Google Scholar] [CrossRef]
- Cheyrou, A.; Guyomarc’h, C.; Jasserand, P.; Blouin, P. Improved detection of HBV DNA by PCR after microwave treatment of serum. Nucleic Acids Res. 1991, 19, 4006. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, H.; Kawana, T.; Fukushima, E.; Suzutani, T. Tolerance of loop-mediated isothermal amplification to a culture medium and biological substances. J. Biochem. Biophys. Methods 2007, 70, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Matsuo, T.; Kawamura, Y.; Ohashi, M.; Yonekawa, T.; Kanda, H.; Notomi, T.; Ihira, M. Direct detection of human herpesvirus 6B by the LAMP method using newly developed dry-reagents. J. Virol. Methods 2014, 201, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Ihira, M.; Sugiyama, H.; Enomoto, Y.; Higashimoto, Y.; Sugata, K.; Asano, Y.; Yoshikawa, T. Direct detection of human herpesvirus 6 DNA in serum by variant specific loop-mediated isothermal amplification in hematopoietic stem cell transplant recipients. J. Virol. Methods 2010, 167, 103–106. [Google Scholar] [CrossRef]
- Curtis, K.A.; Rudolph, D.L.; Owen, S.M. Rapid detection of HIV-1 by reverse-transcription, loop-mediated isothermal amplification (RT-LAMP). J. Virol. Methods 2008, 151, 264–270. [Google Scholar] [CrossRef]
- Bista, B.R.; Ishwad, C.; Wadowsky, R.M.; Manna, P.; Randhawa, P.S.; Gupta, G.; Adhikari, M.; Tyagi, R.; Gasper, G.; Vats, A. Development of a loop-mediated isothermal amplification assay for rapid detection of BK Virus. J. Clin. Microbiol. 2007, 45, 1581–1587. [Google Scholar] [CrossRef]
- Ihira, M.; Akimoto, S.; Miyake, F.; Fujita, A.; Sugata, K.; Suga, S.; Ohashi, M.; Nishimura, N.; Ozaki, T.; Asano, Y.; et al. Direct detection of human herpesvirus 6 DNA in serum by the loop-mediated isothermal amplification method. J. Clin. Virol. 2007, 39, 22–26. [Google Scholar] [CrossRef]
- Liu, S.; Lin, Y.; Liu, T.; Cheng, C.; Wei, W.; Wang, L.; Li, F. Enzyme-free and label-free ultrasensitive electrochemical detection of DNA and adenosine triphosphate by dendritic DNA concatamer-based signal amplification. Biosens. Bioelectron. 2014, 56, 12–18. [Google Scholar] [CrossRef]
- Nie, J.; Zhang, D.W.; Tie, C.; Zhou, Y.L.; Zhang, X.X. G-quadruplex based two-stage isothermal exponential amplification reaction for label-free DNA colorimetric detection. Biosens. Bioelectron. 2014, 56, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wu, Y.; Chen, Y.; Zhu, Z.; Yang, X.; Yang, C.J.; Wang, K.; Tan, W. Pyrene-excimer probes based on the hybridization chain reaction for the detection of nucleic acids in complex biological fluids. Angew. Chem. Int. Ed. Engl. 2011, 50, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lifsond, M.A.; Incid, F.; Liang, L.-G.; Sheng, Y.-F.; Demircid, U. Advances in addressing technical challenges of point-of-care diagnostics in resource-limited settings. Expert Rev. Mol. Diagn. 2016, 16, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Farnaud, S.J.C.; Kosti, O.; Getting, S.J.; Renshaw, D. Saliva: Physiology and diagnostic potential in health and disease. Sci. World J. 2010, 10, 434–456. [Google Scholar] [CrossRef] [PubMed]
- Deneer, H.G.; Knight, I. Inhibition of the Polymerase Chain Reaction by Mucolytic Agents. Clin. Chem. 1994, 40, 171–172. [Google Scholar] [PubMed]
- Nolte, F.S.; Metchock, B.; Mcgowan, J.E.; Edwards, A.; Okwumabua, O.G.I.; Thurmond, C.; Mitchell, P.S.; Plikaytis, B.; Shinnick, T. Direct Detection of Mycobacterium tuberculosis in Sputum by Polymerase Chain Reaction and DNA Hybridization. J. Clin. Microbiol. 1993, 31, 1777–1782. [Google Scholar]
- Aas, J.A.; Paster, B.J.; Stokes, L.N.; Olsen, I.; Dewhirst, F.E. Defining the Normal Bacterial Flora of the Oral Cavity Defining the Normal Bacterial Flora of the Oral Cavity. J. Clin. Microbiol. 2005, 43, 5721–5732. [Google Scholar] [CrossRef] [PubMed]
- Shenai, S.; Armstrong, D.T.; Valli, E.; Dolinger, D.L.; Nakiyingi, L.; Dietze, R.; Dalcolmo, M.P.; Nicol, M.P.; Zemanay, W.; Manabe, Y.; et al. Analytical and clinical evaluation of the epistem genedrive assay for detection of mycobacterium tuberculosis. J. Clin. Microbiol. 2016, 54, 1051–1057. [Google Scholar] [CrossRef]
- Shenai, S.; Amisano, D.; Ronacher, K.; Kriel, M.; Banada, P.P.; Song, T.; Lee, M.; Joh, J.S.; Winter, J.; Thayer, R.; et al. Exploring alternative biomaterials for diagnosis of pulmonary tuberculosis in HIV-negative patients by use of the geneXpert MTB/RIF assay. J. Clin. Microbiol. 2013, 51, 4161–4166. [Google Scholar] [CrossRef]
- Theron, G.; Peter, J.; Calligaro, G.; Meldau, R.; Hanrahan, C.; Khalfey, H.; Matinyenya, B.; Muchinga, T.; Smith, L.; Pandie, S.; et al. Determinants of PCR performance (Xpert MTB/RIF), including bacterial load and inhibition, for TB diagnosis using specimens from different body compartments. Sci. Rep. 2014, 4, 5658. [Google Scholar] [CrossRef]
- Boehme, C.C.; Pamela, N.; Hillemann, D.; Nicol, M.P.; Shenai, S.; Krapp, F.; Allen, J.; Tahirli, R.; Blakemore, R.; Rustomjee, R.; et al. Rapid Molecular Detection of Tuberculosis and Rifampin Resistance. N. Engl. J. Med. 2010, 363, 2373–2383. [Google Scholar] [CrossRef]
- Yamamoto, N.; Hamaguchi, S.; Akeda, Y.; Santanirand, P.; Kerdsin, A.; Seki, M.; Ishii, Y.; Paveenkittiporn, W.; Bonomo, R.A.; Oishi, K.; et al. Clinical Specimen-Direct LAMP: A Useful Tool for the Surveillance of blaOXA-23-Positive Carbapenem-Resistant Acinetobacter baumannii. PLoS ONE 2015, 10, e0133204. [Google Scholar] [CrossRef] [PubMed]
- Nuchprayoon, S.; Saksirisampant, W.; Jaijakul, S.; Nuchprayoon, I. Flinders technology associates (FTA) filter paper–based DNA extraction with polymerase chain reaction (PCR) for detection of Pneumocystis jirovecii from respiratory specimens of immunocompromised patients. J. Clin. Lab. Anal. 2007, 21, 382–386. [Google Scholar] [CrossRef]
- Boehme, C.C.; Nabeta, P.; Henostroza, G.; Raqib, R.; Rahim, Z.; Gerhardt, M.; Sanga, E.; Hoelscher, M.; Notomi, T.; Hase, T.; et al. Operational feasibility of using loop-mediated isothermal amplification for diagnosis of pulmonary tuberculosis in microscopy centers of developing countries. J. Clin. Microbiol. 2007, 45, 1936–1940. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, H.; Imura, M.; Tateno, A.; Tsunoda, T.; Inui, A.; Sogo, T.; Fujisawa, T. Successful use of saliva without DNA extraction for detection of macrolide-resistant Mycoplasma pneumoniae DNA in children using LNA probe-based real-time PCR. J. Infect. Chemother. 2013, 19, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Gale, N.; French, D.J.; Howard, R.L.; McDowell, D.G.; Debenham, P.G.; Brown, T. Rapid typing of STRs in the human genome by HyBeacon melting. Org. Biomol. Chem. 2008, 6, 4553–4559. [Google Scholar] [CrossRef] [PubMed]
- French, D.J.; Archard, C.L.; Andersen, M.T.; McDowell, D.G. Ultra-rapid DNA analysis using HyBeacon probes and direct PCR amplification from saliva. Mol. Cell. Probes 2002, 16, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, J.; Sato, Y.; Shinozaki, N.; Ye, B.; Tsuboi, A.; Nagasaki, M.; Yamashita, R. Comparison of boiling and robotics automation method in DNA extraction for metagenomic sequencing of human oral microbes. PLoS ONE 2016, 11, e0154389. [Google Scholar] [CrossRef]
- Lazarevic, V.; Gaïa, N.; Girard, M.; François, P.; Schrenzel, J. Comparison of DNA Extraction Methods in Analysis of Salivary Bacterial Communities. PLoS ONE 2013, 8, e67699. [Google Scholar] [CrossRef]
- Lazarevic, V.; Whiteson, K.; Hernandez, D.; François, P.; Schrenzel, J. Study of inter- and intra-individual variations in the salivary microbiota. BMC Genom. 2010, 11, 523. [Google Scholar] [CrossRef]
- Faveri, M.; Mayer, M.P.A.; Feres, M.; De Figueiredo, L.C.; Dewhirst, F.E.; Paster, B.J. Microbiological diversity of generalized aggressive periodontitis by 16S rRNA clonal analysis. Oral Microbiol. Immunol. 2008, 23, 112–118. [Google Scholar] [CrossRef]
- Du, Y.; Hughes, R.A.; Bhadra, S.; Jiang, Y.S.; Ellington, A.D.; Li, B. A Sweet Spot for Molecular Diagnostics: Coupling Isothermal Amplification and Strand Exchange Circuits to Glucometers. Sci. Rep. 2015, 5, 11039. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, P.; Rosenau, J.; Springer, B.; Teschner, K.; Feldmann, K.; Bottger, E.C. Diagnosis of mycobacterial infections by nucleic acid amplification: 18-month prospective study. J. Clin. Microbiol. 1996, 34, 304–312. [Google Scholar] [PubMed]
- Ferguson, T.M.; Weigel, K.M.; Lakey Becker, A.; Ontengco, D.; Narita, M.; Tolstorukov, I.; Doebler, R.; Cangelosi, G.A.; Niemz, A. Pilot study of a rapid and minimally instrumented sputum sample preparation method for molecular diagnosis of tuberculosis. Sci. Rep. 2016, 6, 19541. [Google Scholar] [CrossRef] [PubMed]
- Geojith, G.; Dhanasekaran, S.; Chandran, S.P.; Kenneth, J. Efficacy of loop mediated isothermal amplification (LAMP) assay for the laboratory identification of Mycobacterium tuberculosis isolates in a resource limited setting. J. Microbiol. Methods 2011, 84, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Aryan, E.; Makvandi, M.; Farajzadeh, A.; Huygen, K.; Bifani, P.; Mousavi, S.L.; Fateh, A.; Jelodar, A.; Gouya, M.M.; Romano, M. A novel and more sensitive loop-mediated isothermal amplification assay targeting IS6110 for detection of Mycobacterium tuberculosis complex. Microbiol. Res. 2010, 165, 211–220. [Google Scholar] [CrossRef]
- Pandey, B.D.; Poudel, A.; Yoda, T.; Tamaru, A.; Oda, N.; Fukushima, Y.; Lekhak, B.; Risal, B.; Acharya, B.; Sapkota, B.; et al. Development of an in-house loop-mediated isothermal amplification (LAMP) assay for detection of Mycobacterium tuberculosis and evaluation in sputum samples of Nepalese patients. J. Med. Microbiol. 2008, 57, 439–443. [Google Scholar] [CrossRef]
- Boyle, D.S.; McNerney, R.; Teng Low, H.; Leader, B.T.; Pérez-Osorio, A.C.; Meyer, J.C.; O’Sullivan, D.M.; Brooks, D.G.; Piepenburg, O.; Forrest, M.S. Rapid detection of Mycobacterium tuberculosis by recombinase polymerase amplification. PLoS ONE 2014, 9, e103091. [Google Scholar] [CrossRef] [PubMed]
- Afghani, B.; Stutman, H.R. Polymerase chain reaction for diagnosis of M. tuberculosis: Comparison of simple boiling and a conventional method for DNA extraction. Biochem. Mol. Med. 1996, 57, 14–18. [Google Scholar] [CrossRef]
- Kocagöz, T.; Yilmaz, E.; Ozkara, S.; Kocagöz, S.; Hayran, M.; Sachedeva, M.; Chambers, H.F. Detection of Mycobacterium tuberculosis in sputum samples by polymerase chain reaction using a simplified procedure. J. Clin. Microbiol. 1993, 31, 1435–1438. [Google Scholar] [Green Version]
- Shawar, R.M.; el Zaatari, F.A.; Nataraj, A.; Clarridge, J.E. Detection of Mycobacterium tuberculosis in clinical samples by two-step polymerase chain reaction and nonisotopic hybridization methods. J. Clin. Microbiol. 1993, 31, 61–65. [Google Scholar] [PubMed]
- Buck, G.E.; O’Hara, L.C.; Summersgill, J.T. Rapid, simple method for treating clinical specimens containing Mycobacterium tuberculosis to remove DNA for polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 1331–1334. [Google Scholar] [PubMed]
- Thomson, L.M.; Traore, H.; Yesilkaya, H.; Doig, C.; Steingrimsdottir, H.; Garcia, L.; Forbes, K.J. An extremely rapid and simple DNA-release method for detection of M. tuberculosis from clinical specimens. J. Microbiol. Methods 2005, 63, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Mitarai, S.; Okumura, M.; Toyota, E.; Yoshiyama, T.; Aono, A.; Sejimo, A.; Azuma, Y.; Sugahara, K.; Nagasawa, T.; Nagayama, N.; et al. Evaluation of a simple loop-mediated isothermal amplification test kit for the diagnosis of tuberculosis. Int. J. Tuberc. Lung Dis. 2011, 15, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Tarhan, G.; Ceyhan, I.; Şimşek, H.; Tuncer, S. Evaluation of the efficacy of five DNA extraction methods for the detection of Mycobacterium tuberculosis DNA in direct and processed sputum by an in-house PCR method. Turk. J. Med. Sci. 2009, 39, 253–257. [Google Scholar]
- Nihonyanagi, S.; Kanoh, Y.; Okada, K.; Uozumi, T.; Kazuyama, Y.; Yamaguchi, T.; Nakazaki, N.; Sakurai, K.; Hirata, Y.; Munekata, S.; et al. Clinical usefulness of multiplex PCR lateral flow in MRSA detection: A novel, rapid genetic testing method. Inflammation 2012, 35, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Mutingwende, I.; Vermeulen, U.; Steyn, F.; Viljoen, H.; Grobler, A. Development and evaluation of a rapid multiplex-PCR based system for Mycobacterium tuberculosis diagnosis using sputum samples. J. Microbiol. Methods 2015, 116, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Sjöbring, U.; Mecklenburg, M.; Andersen, A.B.; Miörner, H. Polymerase chain reaction for detection of Mycobacterium tuberculosis. J. Clin. Microbiol. 1990, 28, 2200–2204. [Google Scholar] [Green Version]
- Sritharan, V.; Barker, R.H. A simple method for diagnosing M. tuberculosis infection in clinical samples using PCR. Mol. Cell. Probes 1991, 5, 385–395. [Google Scholar] [CrossRef]
- Lunny, C.; Taylor, D.; Hoang, L.; Wong, T.; Gilbert, M.; Lester, R.; Krajden, M.; Ogilvie, G. Self-Collected versus Clinician-Collected Sampling for Chlamydia and Gonorrhea Screening: A Systemic Review and Meta-Analysis. PLoS ONE 2015, 10, e0132776. [Google Scholar] [CrossRef]
- Thompson, P.P.; Kowalski, R.P.; Shanks, R.M.Q.; Gordon, Y.J. Validation of real-time PCR for laboratory diagnosis of Acanthamoeba keratitis. J. Clin. Microbiol. 2008, 46, 3232–3236. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, P.; Rostane, H.; Saint-Jean, C.; Batellier, L.; Alouch, C.; Zito, E.; Bourcier, T.; Laroche, L.; Chaumeil, C. Effects of topical anaesthetics and fluorescein on the real-time PCR used for the diagnosis of Herpesviruses and Acanthamoeba keratitis. Br. J. Ophthalmol. 2006, 90, 1354–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiedbrauk, D.L.; Werner, J.C.; Drevon, A.N.N.M. Inhibition of PCR by Aqueous and Vitreous Fluids. J. Clin. Microbiol. 1995, 33, 2643–2646. [Google Scholar]
- Schläpfer, G.; Senn, H.P.; Berger, R.; Just, M. Use of the polymerase chain reaction to detect Bordetella pertussis in patients with mild or atypical symptoms of infection. Eur. J. Clin. Microbiol. Infect. Dis. 1993, 12, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Harding-Esch, E.M.; Holland, M.J.; Schémann, J.F.; Molina, S.; Sarr, I.; Andreasen, A.A.; Roberts, C.H.; Sillah, A.; Sarr, B.; Harding, E.F.; et al. Diagnostic accuracy of a prototype point-of-care test for ocular chlamydia trachomatis under field conditions in the Gambia and Senegal. PLoS Negl. Trop. Dis. 2011, 5, e1234. [Google Scholar] [CrossRef]
- Sim, J.E.; Park, S.J.; Lee, H.C.; Kim, S.-Y.; Kim, J.Y.; Lee, S.H. High-throughput STR analysis for DNA database using direct PCR. J. Forensic Sci. 2013, 58, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.; Bonner, A.; Cohen, D.M.; Birkhahn, R.; Yogev, R.; Triner, W.; Cohen, J.; Palavecino, E.; Selvarangan, R. Multicenter clinical evaluation of the novel Alere i Influenza A&B isothermal nucleic acid amplification test. J. Clin. Virol. 2014, 61, 81–86. [Google Scholar]
- Harty, L.C.; Rothman, N.; Reid, Y.A.; Tucker, M.A.; Hartge, P. Collection of Buccal Cell DNA Using Treated Cards. Cancer Epidemiol. Biomark. Prev. 2000, 9, 501–506. [Google Scholar]
- Manage, D.P.; Lauzon, J.; Atrazhev, A.; Morrissey, Y.C.; Edwards, A.L.; Stickel, A.J.; Crabtree, H.J.; Pabbaraju, K.; Zahariadis, G.; Yanow, S.K.; et al. A miniaturized and integrated gel post platform for multiparameter PCR detection of herpes simplex viruses from raw genital swabs. Lab Chip 2012, 12, 1664–1671. [Google Scholar] [CrossRef]
- Manage, D.P.; Lauzon, J.; Atrazev, A.; Chavali, R.; Samuel, R.A.; Chan, B.; Morrissey, Y.C.; Gordy, W.; Edwards, A.L.; Larison, K.; et al. An enclosed in-gel PCR amplification cassette with multi-target, multi-sample detection for platform molecular diagnostics. Lab Chip 2013, 13, 2576–2584. [Google Scholar] [CrossRef] [Green Version]
- Sakai, K.; Wakasugi, S.; Muchemwa, F.C.; Ihn, H. Quick detection of herpes viruses from skin vesicles and exudates without nucleic acid extraction using multiplex PCR. Biosci. Trends 2008, 2, 164–168. [Google Scholar] [PubMed]
- Pandori, M.W.; Lei, J.; Wong, E.H.; Klausner, J.; Liska, S. Real-Time PCR for detection of herpes simplex virus without nucleic acid extraction. BMC Infect. Dis. 2006, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Loeffelholz, M.J.; Lewinski, C.A.; Silver, S.R.; Purohit, A.P.; Buonagurio, D.A.; Dragon, E.A. Detection of Chlamydia trachomatis in Endocervical Specimens by Polymerase Chain Reaction. J. Clin. Microbiol. 1992, 30, 2847–2851. [Google Scholar] [PubMed]
- Manage, D.P.; Ma, L.; Lauzon, J.; Howell, A.; Belch, A.R.; Mackey, J.R.; Pilarski, L.M. Genotyping Single Nucleotide Polymorphisms in Human Genomic DNA with an Automated and Self-Contained PCR Cassette. J. Mol. Diagn. 2014, 16, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Grujičić, N.K.; Davidović, S.; Takić, D.; Mojsin, M.; Stevanović, M. Direct PCR amplification of the HVSI region in mitochondrial DNA from buccal cell swabs. Arch. Biol. Sci. 2012, 64, 851–858. [Google Scholar] [CrossRef]
- Lind-brandberg, L.; Welinder-olsson, C.; Taranger, J.; Trollfors, B.; Lagergård, T.; Zackrisson, G. Evaluation of PCR for Diagnosis of Bordetella pertussis and Bordetella parapertussis Infections These include: Evaluation of PCR for Diagnosis of Bordetella pertussis and Bordetella parapertussis Infections. J. Clin. Microbiol. 1998, 36, 679–683. [Google Scholar] [PubMed]
- Kinchington, P.R.; Turse, S.E.; Kowalski, R.P.; Gordon, Y.J. Use of polymerase chain amplification reaction for the detection of adenoviruses in ocular swab specimens. Investig. Ophthalmol. Vis. Sci. 1994, 35, 4126–4134. [Google Scholar]
- Tabrizi, S.N.; Lees, M.I.; Garland, S.M. Comparison of polymerase chain reaction and culture techniques for detection of Chlamydia trachomatis. Mol. Cell. Probes 1993, 7, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.L.; French, D.J.; Richardson, J.A.; O’Neill, C.E.; Andreou, M.P.; Brown, T.; Clark, D.; Clarke, I.N.; Holloway, J.W.; Marsh, P.; et al. Rapid detection of diagnostic targets using isothermal amplification and HyBeacon probes—A homogenous system for sequence-specific detection. Mol. Cell. Probes 2014, 29, 92–98. [Google Scholar] [CrossRef]
- Kobayashi, T.; Yagami, A.; Suzuki, K.; Ihira, M.; Yoshikawa, T.; Matsunaga, K. Clinical utility of loop-mediated isothermal amplification assay for the diagnosis of common alpha herpesvirus skin infections. J. Dermatol. 2013, 40, 1033–1037. [Google Scholar] [CrossRef]
- Enomoto, Y.; Yoshikawa, T.; Ihira, M.; Akimoto, S.; Miyake, F.; Usui, C.; Suga, S.; Suzuki, K. Rapid Diagnosis of Herpes Simplex Virus Infection by a Loop-Mediated Isothermal Amplification Method. J. Clin. Microbiol. 2005, 43, 951–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, S.; Yoshikawa, T.; Ihira, M.; Suzuki, K.; Shimokata, K.; Nishiyama, Y.; Asano, Y. Rapid detection of varicella-zoster virus infection by a loop-mediated isothermal amplification method. J. Med. Virol. 2004, 74, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, T.; Yamashita, R.; Kakita, T.; Kakita, M.; Oshika, T. Rapid and sensitive diagnosis of adenoviral keratoconjunctivitis by loop-mediated isothermal amplification (LAMP) method. Curr. Eye Res. 2004, 28, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhao, L.; Zhao, M.; Zhu, R.; Deng, J.; Wang, F.; Li, F.; Ding, Y.; Tian, R.; Qian, Y. Four DNA extraction methods used in loop-mediated isothermal amplification for rapid adenovirus detection. J. Virol. Methods 2014, 204, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Mahony, J.; Chong, S.; Bulir, D.; Ruyter, A.; Mwawasi, K.; Waltho, D. Development of a sensitive loop-mediated isothermal amplification assay that provides specimen-to-result diagnosis of respiratory syncytial virus infection in 30 min. J. Clin. Microbiol. 2013, 51, 2696–2701. [Google Scholar] [CrossRef]
- Mahony, J.; Chong, S.; Bulir, D.; Ruyter, A.; Mwawasi, K.; Waltho, D. Multiplex loop-mediated isothermal amplification (M-LAMP) assay for the detection of influenza A/H1, A/H3 and influenza B can provide a specimen-to-result diagnosis in 40min with single genome copy sensitivity. J. Clin. Virol. 2013, 58, 127–131. [Google Scholar] [CrossRef]
- Nie, K.; Qi, S.X.; Zhang, Y.; Luo, L.; Xie, Y.; Yang, M.J.; Zhang, Y.; Li, J.; Shen, H.; Li, Q.; et al. Evaluation of a Direct Reverse Transcription Loop-Mediated Isothermal Amplification Method without RNA Extraction for the Detection of Human Enterovirus 71 Subgenotype C4 in Nasopharyngeal Swab Specimens. PLoS ONE 2012, 7, e52486. [Google Scholar] [CrossRef] [PubMed]
- Ablordey, A.; Amissah, D.A.; Aboagye, I.F.; Hatano, B.; Yamazaki, T.; Sata, T.; Ishikawa, K.; Katano, H. Detection of Mycobacterium ulcerans by the loop mediated isothermal amplification method. PLoS Negl. Trop. Dis. 2012, 6, e1590. [Google Scholar] [CrossRef]
- Iwata, S.; Shibata, Y.; Kawada, J.; Hara, S.; Nishiyama, Y.; Morishima, T.; Ihira, M.; Yoshikawa, T.; Asano, Y.; Kimura, H. Rapid detection of Epstein-Barr virus DNA by loop-mediated isothermal amplification method. J. Clin. Virol. 2006, 37, 128–133. [Google Scholar] [CrossRef]
- Walker, G.T.; Little, M.C.; Nadeau, J.G.; Shank, D.D. Isothermal in vitro amplification of DNA by a restriction enzyme/DNA polymerase system. Proc. Natl. Acad. Sci. USA 1992, 89, 392–396. [Google Scholar] [CrossRef]
- Lafleur, L.; Bishop, J.D.; Heiniger, E.K.; Gallagher, R.P.; Wheeler, M.D.; Kauffman, P.C.; Zhang, X.; Kline, E.; Buser, J.; Ramachandran, S.; et al. A rapid, instrument-free, sample-to-result nucleic acid amplification test. Lab Chip 2016, 52, 3377–3383. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, N.M.; Linnes, J.C.; Fan, A.; Ellenson, C.K.; Pollock, N.R.; Klapperich, C.M. Paper-Based RNA Extraction, in Situ Isothermal Amplification, and Lateral Flow Detection for Low-Cost, Rapid Diagnosis of Influenza A (H1N1) from Clinical Specimens. Anal. Chem. 2015, 87, 7872–7879. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Chow, W.H.A.; Li, Y.; Kong, H.; Tang, Y.-W.; Lemieux, B. Nucleic acid assay system for tier II laboratories and moderately complex clinics to detect HIV in low-resource settings. J. Infect. Dis. 2010, 201, S46–S51. [Google Scholar] [CrossRef]
- Kim, H.-J.; Tong, Y.; Tang, W.; Quimson, L.; Cope, V.A.; Pan, X.; Motre, A.; Kong, R.; Hong, J.; Kohn, D.; et al. A rapid and simple isothermal nucleic acid amplification test for detection of herpes simplex virus types 1 and 2. J. Clin. Virol. 2011, 50, 26–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akduman, D.; Ehret, J.M.; Messina, K.; Ragsdale, S.; Judson, F.N. Evaluation of a strand displacement amplification assay (BD ProbeTec-SDA) for detection of Neisseria gonorrhoeae in urine specimens. J. Clin. Microbiol. 2002, 40, 281–283. [Google Scholar] [CrossRef]
- Mahony, J.; Song, X.; Chong, S. NucliSens Basic Kit for Detection of Chlamydia trachomatis and Neisseria gonorrhoeaein Genital Tract Specimens Using Nucleic Acid Sequence-Based Amplification. J. Clin. Microbiol. 2001, 39, 1429–1435. [Google Scholar] [CrossRef] [PubMed]
- Khan, G.; Kangro, H.; Coates, P.J.; Heath, R.B. Inhibitory effects of urine on the polymerase chain reaction for cytomegalovirus DNA. J. Clin. Pathol. 1991, 44, 360–365. [Google Scholar] [CrossRef]
- McElgunn, C.J.; Pereira, C.R.; Parham, N.J.; Smythe, J.E.; Wigglesworth, M.J.; Smielewska, A.; Parmar, S.A.; Gandelman, O.A.; Brown, N.M.; Tisi, L.C.; et al. A low complexity rapid molecular method for detection of Clostridium difficile in stool. PLoS ONE 2014, 9, e83808. [Google Scholar] [CrossRef]
- Al-soud, W.A.; Rådström, P. Effects of Amplification Facilitators on Diagnostic PCR in the Presence of Blood, Feces, and Meat Effects of Amplification Facilitators on Diagnostic PCR in the Presence of Blood, Feces, and Meat. J. Clin. Microbiol. 2000, 38, 4463–4470. [Google Scholar]
- Humphries, R.M.; Uslan, D.Z.; Rubin, Z. Performance of clostridium difficile toxin enzyme immunoassay and nucleic acid amplification tests stratified by patient disease severity. J. Clin. Microbiol. 2013, 51, 869–873. [Google Scholar] [CrossRef]
- Gyorke, C.E.; Wang, S.; Leslie, J.L.; Cohen, S.H.; Solnick, J.V.; Polage, C.R. Evaluation of Clostridium difficile fecal load and limit of detection during a prospective comparison of two molecular tests, the illumigene C. difficile and xpert C. difficile/epi tests. J. Clin. Microbiol. 2013, 51, 278–280. [Google Scholar] [CrossRef] [PubMed]
- Viala, C.; Le Monnier, A.; Maataoui, N.; Rousseau, C.; Collignon, A.; Poilane, I. Comparison of commercial molecular assays for toxigenic Clostridium difficile detection in stools: BD GeneOhm Cdiff, XPert C. difficile and illumigene C. difficile. J. Microbiol. Methods 2012, 90, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Pancholi, P.; Kelly, C.; Raczkowski, M.; Balada-Llasat, J.M. Detection of toxigenic Clostridium difficile: Comparison of the cell culture neutralization, Xpert C. difficile, Xpert C. difficile/Epi, and Illumigene C. difficile assays. J. Clin. Microbiol. 2012, 50, 1331–1335. [Google Scholar] [CrossRef]
- Pasternack, R.; Vuorinen, P.; Miettinen, A. Evaluation of the Gen-Probe Chlamydia trachomatis transcription-mediated amplification assay with urine specimens from women. J. Clin. Microbiol. 1997, 35, 676–678. [Google Scholar] [PubMed]
- Krõlov, K.; Frolova, J.; Tudoran, O.; Suhorutsenko, J.; Lehto, T.; Sibul, H.; Mäger, I.; Laanpere, M.; Tulp, I.; Langel, Ü. Sensitive and rapid detection of Chlamydia trachomatis by recombinase polymerase amplification directly from urine samples. J. Mol. Diagn. 2014, 16, 127–135. [Google Scholar] [CrossRef]
- Al-Talib, H.; Latif, B.; Mohd-Zain, Z. Pentaplex PCR assay for detection of hemorrhagic bacteria from stool samples. J. Clin. Microbiol. 2014, 52, 3244–3249. [Google Scholar] [CrossRef] [PubMed]
- Lantz, P.G.; Matsson, M.; Wadström, T.; Rådström, P. Removal of PCR inhibitors from human faecal samples through the use of an aqueous two-phase system for sample preparation prior to PCR. J. Microbiol. Methods 1997, 28, 159–167. [Google Scholar] [CrossRef]
- Moore, M.D.; Jaykus, L.-A. Development of a Recombinase Polymerase Amplification Assay for Detection of Epidemic Human Noroviruses. Sci. Rep. 2017, 7, 40244. [Google Scholar] [CrossRef] [Green Version]
- Nixon, G.J.; Svenstrup, H.F.; Donald, C.E.; Carder, C.; Stephenson, J.M.; Morris-Jones, S.; Huggett, J.F.; Foy, C.A. A novel approach for evaluating the performance of real time quantitative loop-mediated isothermal amplification-based methods. Biomol. Detect. Quantif. 2014, 2, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Edwards, T.; Burke, P.A.; Smalley, H.B.; Gillies, L.; Hobbs, G. Loop-mediated isothermal amplification test for detection of Neisseria gonorrhoeae in urine samples and tolerance of the assay to the presence of urea. J. Clin. Microbiol. 2014, 52, 2163–2165. [Google Scholar] [CrossRef]
- Hill, J.; Beriwal, S.; Chandra, I.; Paul, V.K.; Kapil, A.; Singh, T.; Wadowsky, R.M.; Singh, V.; Goyal, A.; Jahnukainen, T.; et al. Loop-mediated isothermal amplification assay for rapid detection of common strains of Escherichia coli. J. Clin. Microbiol. 2008, 46, 2800–2804. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.W.; Procop, G.W.; Persing, D.H. Molecular diagnostics of infectious diseases. Clin. Chem. 1997, 43, 2021–2038. [Google Scholar] [PubMed]
- Dvorin, N. The Global Molecular Diagnostics Market Report #A344; Medtech Insight: New York, NY, USA, 2015. [Google Scholar]
- Peeling, R.W.; Mabey, D. Point-of-care tests for diagnosing infections in the developing world. Clin. Microbiol. Infect. 2010, 16, 1062–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.H.; Dineva, M.A.; Chua, Y.L.; Ritchie, A.V.; Ushiro-Lumb, I.; Wisniewski, C.A. Simple amplification-based assay: A nucleic acid-based point-of-care platform for HIV-1 testing. J. Infect. Dis. 2010, 201, S65–S72. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker, F.M.; Hsieh, K. Advances in Directly Amplifying Nucleic Acids from Complex Samples. Biosensors 2019, 9, 117. https://doi.org/10.3390/bios9040117
Walker FM, Hsieh K. Advances in Directly Amplifying Nucleic Acids from Complex Samples. Biosensors. 2019; 9(4):117. https://doi.org/10.3390/bios9040117
Chicago/Turabian StyleWalker, Faye M., and Kuangwen Hsieh. 2019. "Advances in Directly Amplifying Nucleic Acids from Complex Samples" Biosensors 9, no. 4: 117. https://doi.org/10.3390/bios9040117
APA StyleWalker, F. M., & Hsieh, K. (2019). Advances in Directly Amplifying Nucleic Acids from Complex Samples. Biosensors, 9(4), 117. https://doi.org/10.3390/bios9040117