
Antifungal and Anti-Biofilm Effects of Caffeic Acid Phenethyl Ester on Different Candida Species

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Susceptibility of Candida Planktonic Cells to CAPE
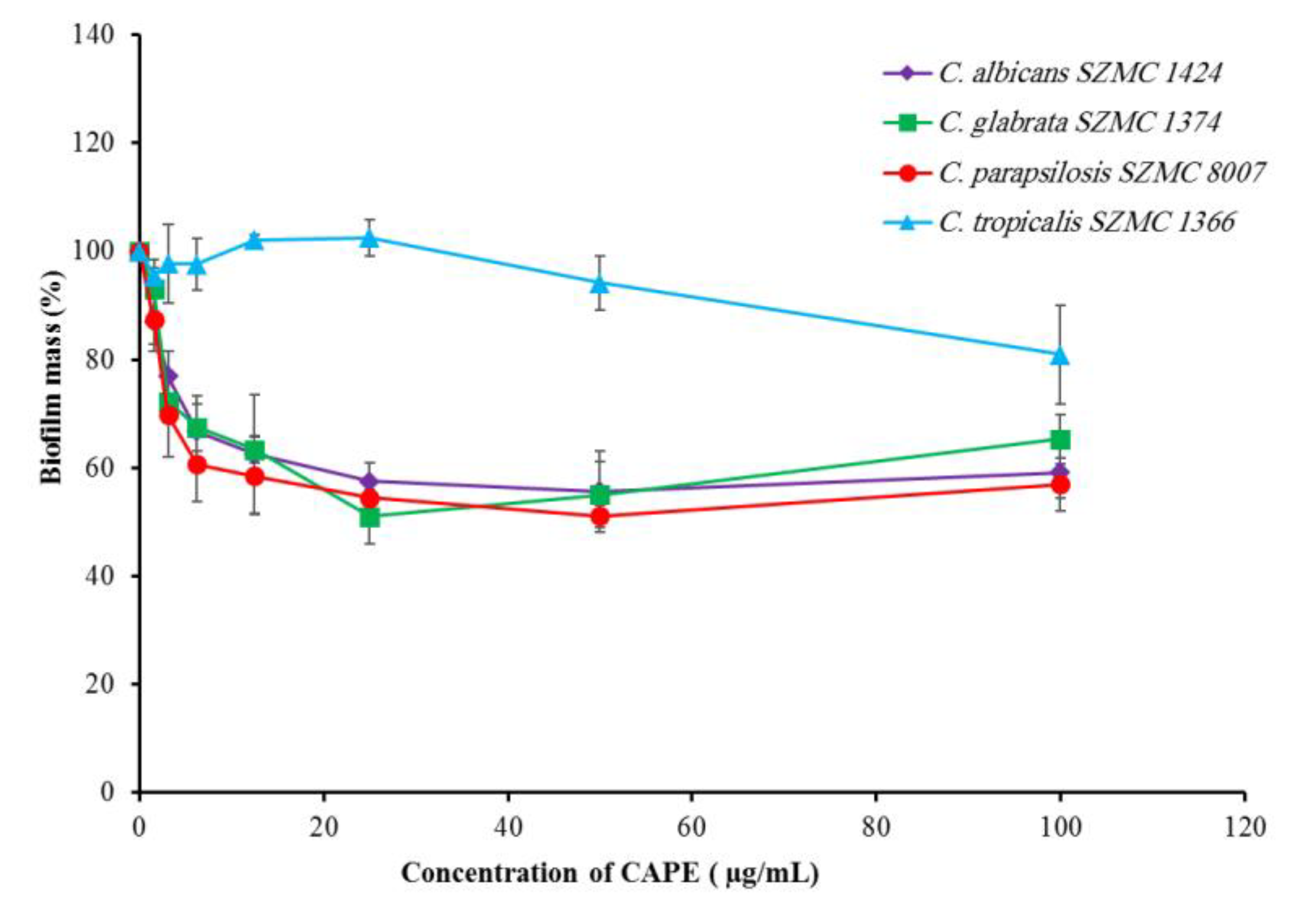
2.2. Effect of CAPE on Candida Biofilm-Forming Ability
2.3. Effect of CAPE on Candida Biofilm Eradication
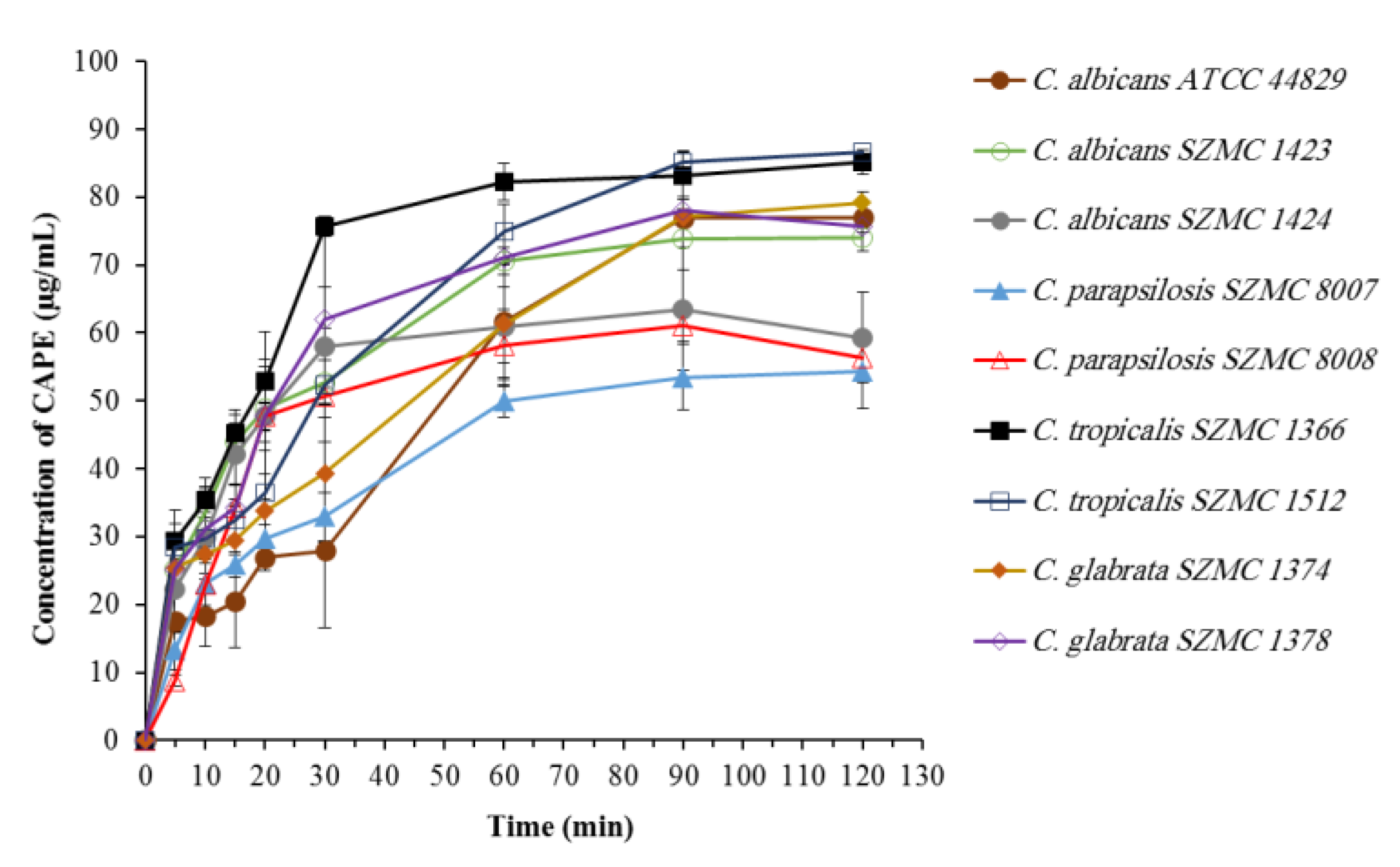
2.4. Biosorption of CAPE by Candida Cells
2.5. Induction of Apoptotic Cell Death in Candida spp. by CAPE
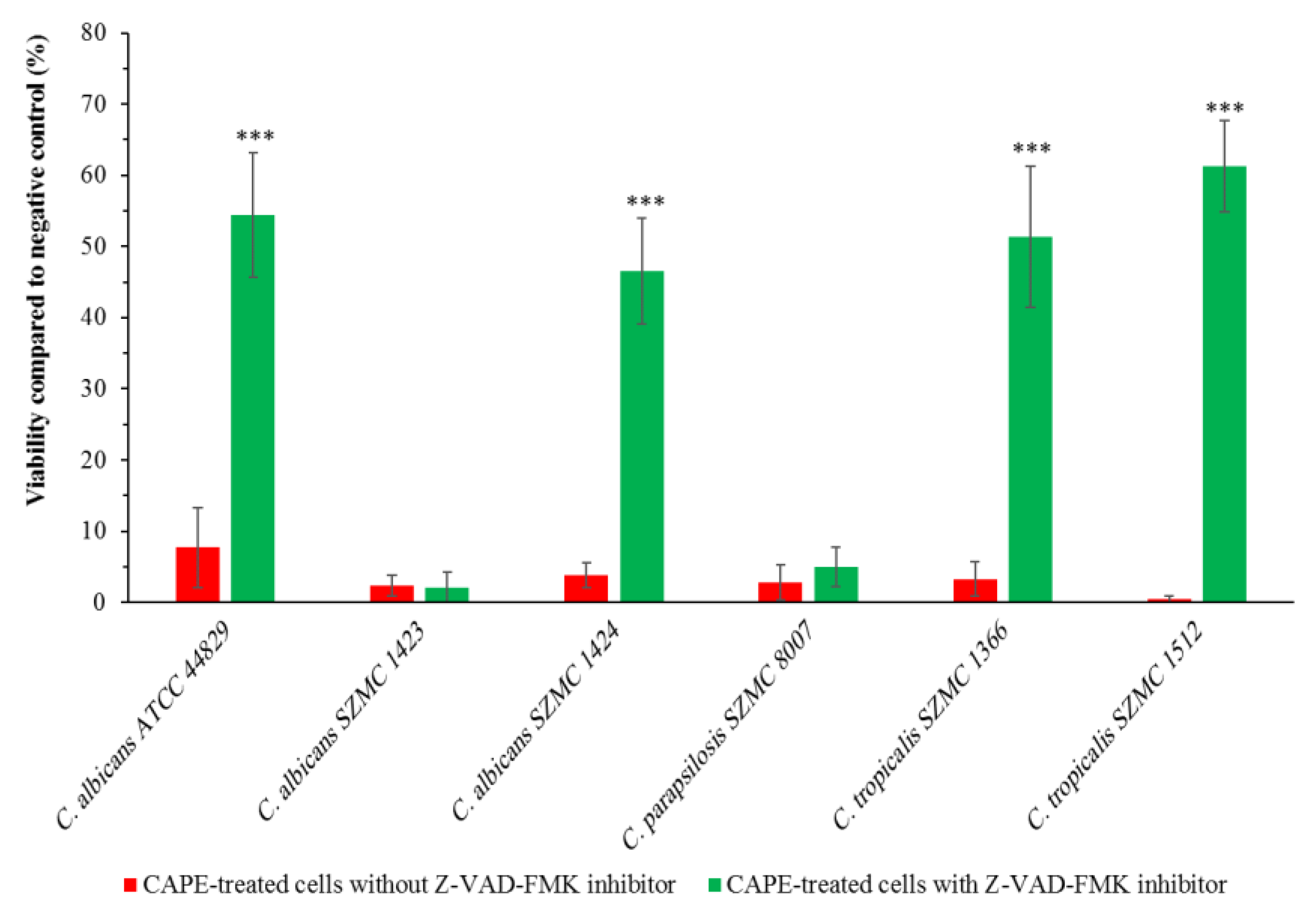
2.6. Effect of Caspase Inhibitor on the Growth of CAPE-Treated Candida Cells
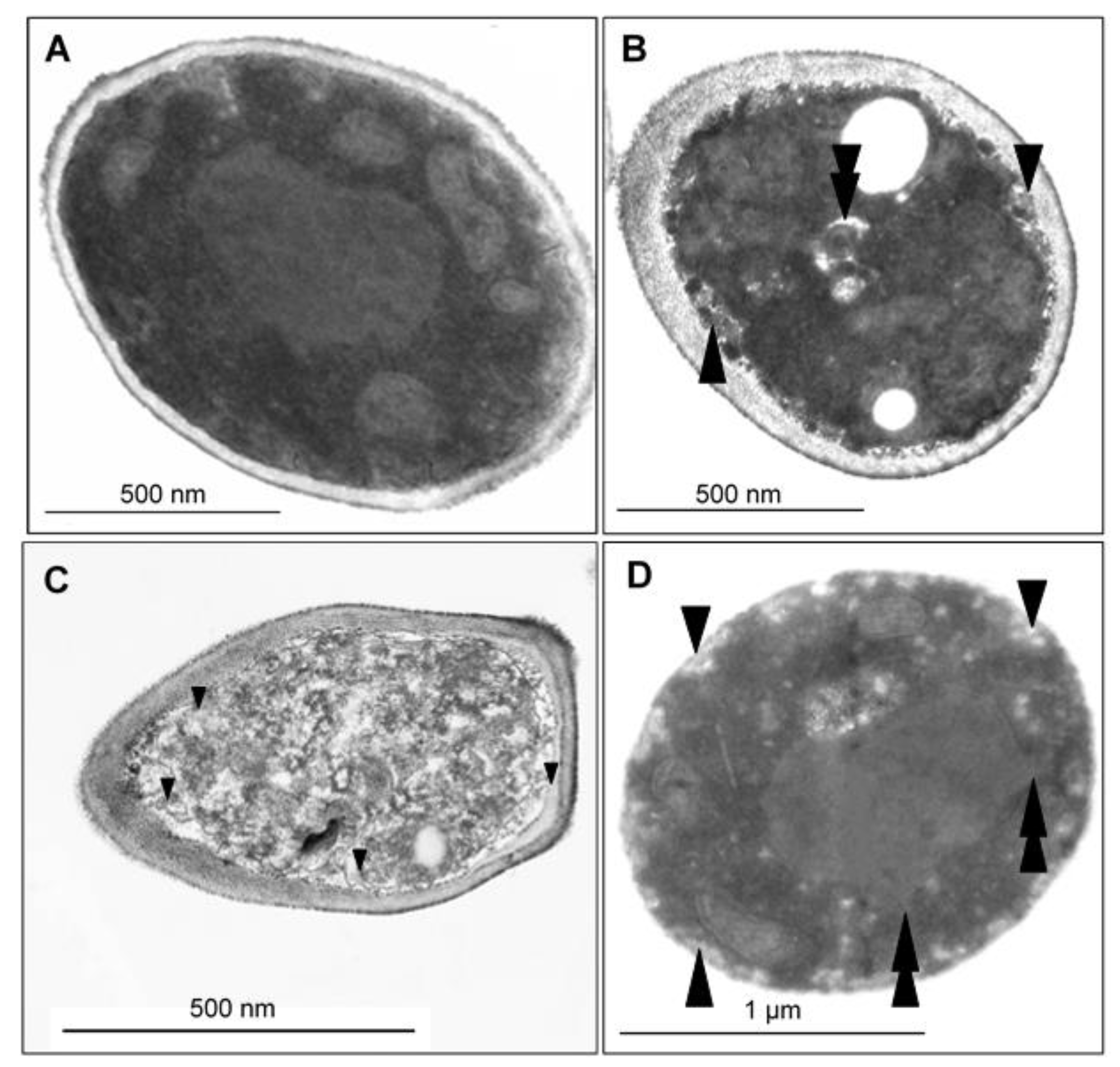
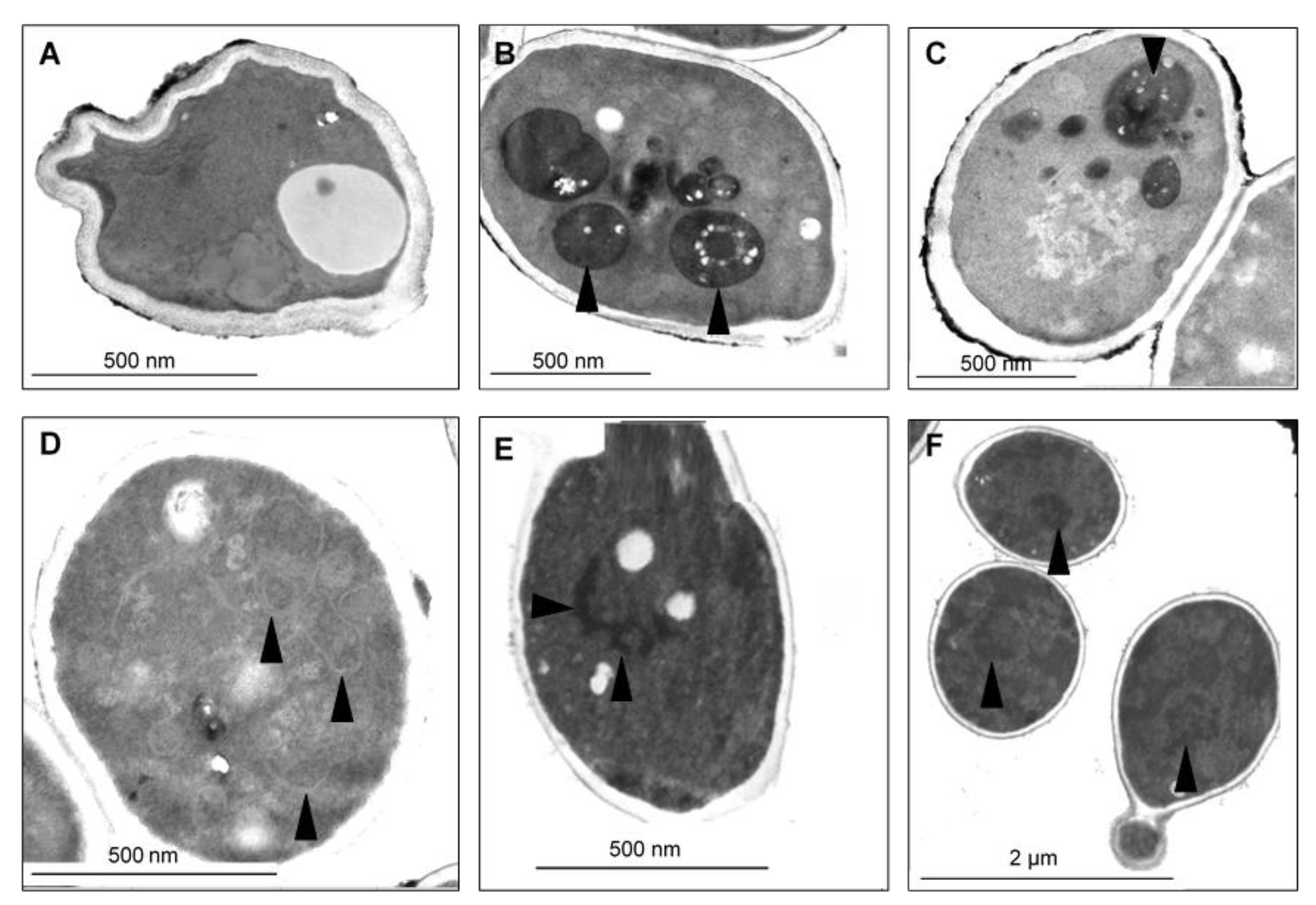
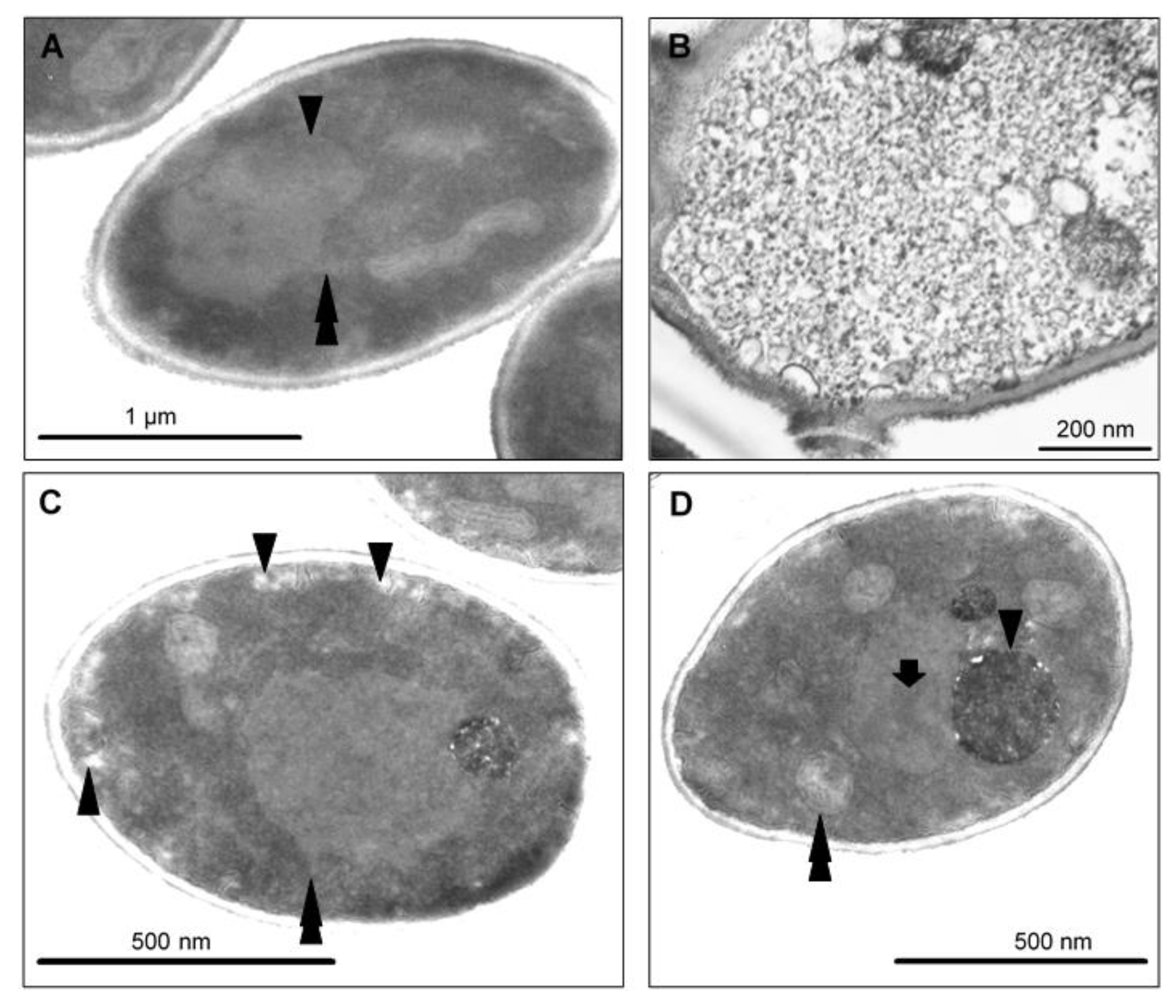
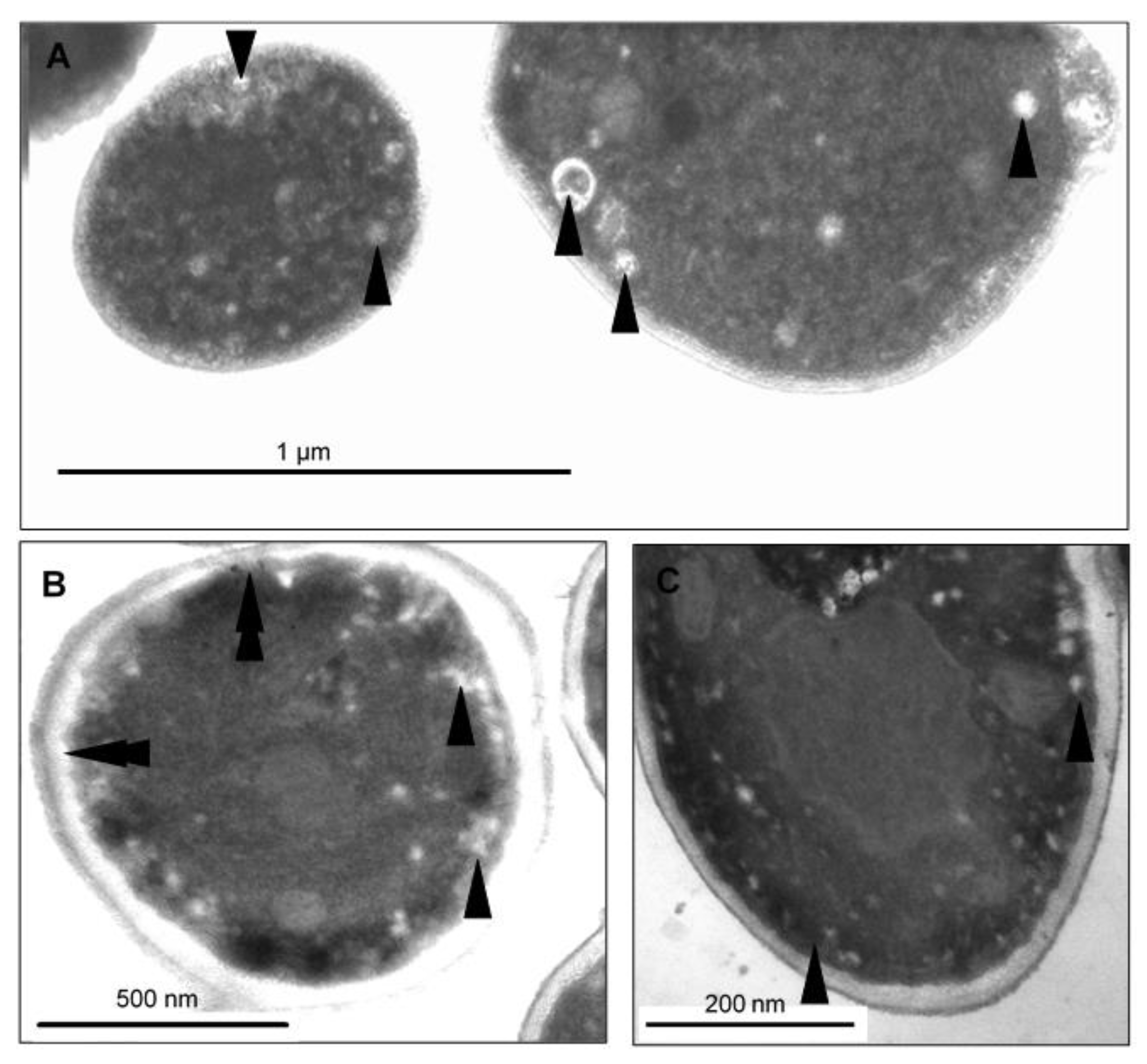
2.7. CAPE-Induced Subcellular Cell Death Markers Determined by TEM
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Instruments Used in the Experiments
4.3. Test Microorganisms, Culture Media, and Growth Conditions
4.4. Preparation of Stock Solution of CAPE
4.5. Antifungal Susceptibility Testing
4.6. Biofilm-Forming Ability Assay
4.7. Biofilm Eradication Assay
4.8. Biosorption of CAPE by Candida Cells
4.9. Cell Death Examination Assay
4.10. Caspase Inhibitor Assay
4.11. Ultrastructural Examination of Candida Species by TEM
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Muñoz, J.E.; Rossi, D.C.P.; Jabes, D.L.; Barbosa, D.A.; Cunha, F.F.M.; Nunes, L.R.; Arruda, D.C.; Pelleschi Taborda, C. In Vitro and In Vivo Inhibitory Activity of Limonene against Different Isolates of Candida spp. J. Fungi 2020, 6, 183. [Google Scholar] [CrossRef]
- Walsh, T.J.; Dixon, D.M. Spectrum of mycoses: Chapter 75. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Patil, S.; Rao, R.S.; Majumdar, B.; Anil, S. Clinical appearance of oral Candida infection and therapeutic strategies. Front. Microbiol. 2015, 6, 1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Sabouraudia 2009, 47, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Ghannoum, M.A. Candida: A causative agent of an emerging infection. J. Investig. Dermatol. Symp. Proc. 2001, 6, 188–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donadu, M.G.; Peralta-Ruiz, Y.; Usai, D.; Maggio, F.; Molina-Hernandez, J.B.; Rizzo, D.; Bussu, F.; Rubino, S.; Zanetti, S.; Paparella, A. Colombian Essential Oil of Ruta graveolens against Nosocomial Antifungal Resistant Candida Strains. J. Fungi 2021, 7, 383. [Google Scholar] [CrossRef]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. FEMS Pathog. Dis. 2016, 74, 1–13. [Google Scholar] [CrossRef] [Green Version]
- El-Houssaini, H.H.; Elnabawy, O.M.; Nasser, H.A.; Elkhatib, W.F. Correlation between antifungal resistance and virulence factors in Candida albicans recovered from vaginal specimens. Microb. Pathog. 2019, 128, 13–19. [Google Scholar] [CrossRef]
- Soliman, S.; Alnajdy, D.; El-Keblawy, A.A.; Mosa, K.A.; Khoder, G.; Noreddin, A.M. Plants’ natural products as alternative promising anti-Candida drugs. Pharmacogn. Rev. 2017, 11, 104. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, P.K.; Chandra, J. Candida biofilm resistance. Drug Resist. Updatates 2004, 7, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Seneviratne, C.J.; Jin, L.; Samaranayake, L.P. Biofilm lifestyle of Candida: A mini review. Oral Dis. 2008, 14, 582–590. [Google Scholar] [CrossRef]
- Alfarrayeh, I.I.S. Bioactivities and Potential Beneficial Properties of Propolis Ethanolic Extract, Caffeic acid phenethyl Ester, and Arabic Coffee Beans Extract. Ph.D. Thesis, University of Pécs, Pécs, Hungary, 2021. [Google Scholar]
- Murtaza, G.; Karim, S.; Akram, M.R.; Khan, S.A.; Azhar, S.; Mumtaz, A.; Bin Asad, M.H.H. Caffeic acid phenethyl ester and therapeutic potentials. Biomed Res. Int. 2014, 2014, 145342. [Google Scholar] [CrossRef] [Green Version]
- Koru, Ö.; Avcu, F.; Tanyüksel, M.; Ural, A.U.; Araz, R.E.; Şener, K. Cytotoxic effects of caffeic acid phenethyl ester (CAPE) on the human multiple myeloma cell line. Turkish J. Med. Sci. 2009, 39, 863–870. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, C.P.; Wang, K.; Li, G.Q.; Hu, F.L. Recent advances in the chemical composition of propolis. Molecules 2014, 19, 19610–19632. [Google Scholar] [CrossRef] [Green Version]
- Yordanov, Y. Caffeic acid phenethyl ester (CAPE): Pharmacodynamics and potential for therapeutic application. Pharmacia 2019, 66, 107. [Google Scholar] [CrossRef] [Green Version]
- Alfarrayeh, I.; Fekete, C.; Gazdag, Z.; Papp, G. Propolis ethanolic extract has double-face in vitro effect on the planktonic growth and biofilm formation of some commercial probiotics. Saudi J. Biol. Sci. 2021, 28, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Lu, W.; Zhu, L.; Shen, X.; Huang, J. Caffeic acid phenethyl ester (CAPE), an active component of propolis, inhibits Helicobacter pylori peptide deformylase activity. Biochem. Biophys. Res. Commun. 2013, 435, 289–294. [Google Scholar] [CrossRef]
- Meyuhas, S.; Assali, M.; Huleihil, M.; Huleihel, M. Antimicrobial activities of caffeic acid phenethyl ester. J. Mol. Biochem. 2015, 4. [Google Scholar]
- Mirzoeva, O.K.; Grishanin, R.N.; Calder, P.C. Antimicrobial action of propolis and some of its components: The effects on growth, membrane potential and motility of bacteria. Microbiol. Res. 1997, 152, 239–246. [Google Scholar] [CrossRef]
- Lv, L.; Cui, H.; Ma, Z.; Liu, X.; Yang, L. Recent progresses in the pharmacological activities of caffeic acid phenethyl ester. Naunyn Schmiedebergs Arch. Pharmacol. 2021, 394, 1327–1339. [Google Scholar] [CrossRef]
- Niu, Y.; Wang, K.; Zheng, S.; Wang, Y.; Ren, Q.; Li, H.; Ding, L.; Li, W.; Zhang, L. Antibacterial effect of caffeic acid phenethyl ester on cariogenic bacteria and Streptococcus mutans biofilms. Antimicrob. Agents Chemother. 2020, 64, e00251-20. [Google Scholar] [CrossRef] [PubMed]
- De Barros, P.P.; Rossoni, R.D.; Garcia, M.T.; de Lima Kaminski, V.; Loures, F.V.; Fuchs, B.B.; Mylonakis, E.; Junqueira, J.C. The anti-biofilm efficacy of caffeic acid phenethyl ester (CAPE) in vitro and a murine model of oral candidiasis. Front. Cell. Infect. Microbiol. 2021, 11, 700305. [Google Scholar] [CrossRef]
- Sun, L.; Liao, K.; Hang, C. Caffeic acid phenethyl ester synergistically enhances the antifungal activity of fluconazole against resistant Candida albicans. Phytomedicine 2018, 40, 55–58. [Google Scholar] [CrossRef]
- Sun, L.; Hang, C.; Liao, K. Synergistic effect of caffeic acid phenethyl ester with caspofungin against Candida albicans is mediated by disrupting iron homeostasis. Food Chem. Toxicol. 2018, 116, 51–58. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K.; Witek-Krowiak, A. State of the art for the biosorption process—A review. Appl. Biochem. Biotechnol. 2013, 170, 1389–1416. [Google Scholar] [CrossRef] [Green Version]
- Sutcu, M.; Salman, N.; Akturk, H.; Dalgıc, N.; Turel, O.; Kuzdan, C.; Kadayifci, E.K.; Sener, D.; Karbuz, A.; Erturan, Z. Epidemiologic and microbiologic evaluation of nosocomial infections associated with Candida spp in children: A multicenter study from Istanbul, Turkey. Am. J. Infect. Control 2016, 44, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Bona, E.; Cantamessa, S.; Pavan, M.; Novello, G.; Massa, N.; Rocchetti, A.; Berta, G.; Gamalero, E. Sensitivity of Candida albicans to essential oils: Are they an alternative to antifungal agents? J. Appl. Microbiol. 2016, 121, 1530–1545. [Google Scholar] [CrossRef] [PubMed]
- Mandras, N.; Roana, J.; Scalas, D.; Del Re, S.; Cavallo, L.; Ghisetti, V.; Tullio, V. The Inhibition of Non-albicans Candida Species and Uncommon Yeast Pathogens by Selected Essential Oils and Their Major Compounds. Molecules 2021, 26, 4937. [Google Scholar] [CrossRef]
- Possamai Rossatto, F.C.; Tharmalingam, N.; Escobar, I.E.; D’Azevedo, P.A.; Zimmer, K.R.; Mylonakis, E. Antifungal Activity of the Phenolic Compounds Ellagic Acid (EA) and Caffeic Acid Phenethyl Ester (CAPE) against Drug-Resistant Candida auris. J. Fungi 2021, 7, 763. [Google Scholar] [CrossRef] [PubMed]
- Cigut, T.; Polak, T.; Gasperlin, L.; Raspor, P.; Jamnik, P. Antioxidative activity of propolis extract in yeast cells. J. Agric. Food Chem. 2011, 59, 11449–11455. [Google Scholar] [CrossRef]
- Breger, J.; Fuchs, B.B.; Aperis, G.; Moy, T.I.; Ausubel, F.M.; Mylonakis, E. Antifungal chemical compounds identified using a C. elegans pathogenicity assay. PLoS Pathog. 2007, 3, e18. [Google Scholar] [CrossRef]
- Su, Z.Z.; Lin, J.; Prewett, M.; Goldstein, N.I.; Fisher, P.B. Apoptosis mediates the selective toxicity of caffeic acid phenethyl ester (CAPE) toward oncogene-transformed rat embryo fibroblast cells. Anticancer Res. 1995, 15, 1841–1848. [Google Scholar]
- Marin, E.H.; Paek, H.; Li, M.; Ban, Y.; Karaga, M.K.; Shashidharamurthy, R.; Wang, X. Caffeic acid phenethyl ester exerts apoptotic and oxidative stress on human multiple myeloma cells. Investig. New Drugs 2019, 37, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Srivastava, S. Anti-Candida activity of two-peptide bacteriocins, plantaricins (Pln E/F and J/K) and their mode of action. Fungal Biol. 2014, 118, 264–275. [Google Scholar] [CrossRef] [PubMed]
- De Nollin, S.; Borgers, M. Scanning electron microscopy of Candida albicans after in vitro treatment with miconazole. Antimicrob. Agents Chemother. 1975, 7, 704–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lone, S.A.; Wani, M.Y.; Fru, P.; Ahmad, A. Cellular apoptosis and necrosis as therapeutic targets for novel Eugenol Tosylate congeners against Candida albicans. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard, 2nd ed.; Clinical and Laboratory Standards Institute: Pittsburgh, PA, USA, 2002; Volume 22, ISBN 1562384694. [Google Scholar]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Nostro, A.; Roccaro, A.S.; Bisignano, G.; Marino, A.; Cannatelli, M.A.; Pizzimenti, F.C.; Cioni, P.L.; Procopio, F.; Blanco, A.R. Effects of oregano, carvacrol and thymol on Staphylococcus aureus and Staphylococcus epidermidis biofilms. J. Med. Microbiol. 2007, 56, 519–523. [Google Scholar] [CrossRef]
- Sun, X.; Zhao, Y.; Liu, L.; Jia, B.; Zhao, F.; Huang, W.; Zhan, J. Copper tolerance and biosorption of Saccharomyces cerevisiae during alcoholic fermentation. PLoS ONE 2015, 10, e0128611. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Zhou, X.; Leng, Q.; Zhang, L.; Cheng, B.; Zhang, X. 7-ketocholesterol-induced caspase-mediated apoptosis in Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 13, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.J.; Sudbery, I.; Ramsdale, M. Apoptosis induced by environmental stresses and amphotericin B in Candida albicans. Proc. Natl. Acad. Sci. USA 2003, 100, 14327–14332. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MIC80 (µg/mL) |
|---|---|
| C. albicans ATCC 44829 | 50 |
| C. albicans SZMC 1423 | 100 |
| C. albicans SZMC 1424 | 50 |
| C. glabrata SZMC 1374 | 12.5 |
| C. glabrata SZMC 1378 | 12.5 |
| C. parapsilosis SZMC 8007 | 25 |
| C. parapsilosis SZMC 8008 | 12.5 |
| C. tropicalis SZMC 1366 | 50 |
| C. tropicalis SZMC 1512 | 50 |
| Species | Collection Code | Origin | Biofilm-Forming Ability |
|---|---|---|---|
| C. albicans | ATCC 44829 | auxotrophic mutant isolated after N-methyl N’-nitro-N-nitrosoguanidine treatment of a wild-type strain of C. albicans. | Non-biofilm forming |
| C. albicans | SZMC 1423 | clinical sample/Debrecen, Hungary | Non-biofilm forming |
| C. albicans | SZMC 1424 | clinical sample/Debrecen, Hungary | High-biofilm forming |
| C. tropicalis | SZMC 1366 | hemoculture/Debrecen, Hungary | High-biofilm forming |
| C. tropicalis | SZMC 1512 | -/Pécs, Hungary | Non-biofilm forming |
| C. glabrata | SZMC 1374 | clinical sample/Debrecen, Hungary | High-biofilm forming |
| C. glabrata | SZMC 1378 | clinical sample/Debrecen, Hungary | Non-biofilm forming |
| C. parapsilosis | SZMC 8007 | clinical sample/Szeged, Hungary | High-biofilm forming |
| C. parapsilosis | SZMC 8008 | unknown | Non-biofilm forming |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfarrayeh, I.; Pollák, E.; Czéh, Á.; Vida, A.; Das, S.; Papp, G. Antifungal and Anti-Biofilm Effects of Caffeic Acid Phenethyl Ester on Different Candida Species. Antibiotics 2021, 10, 1359. https://doi.org/10.3390/antibiotics10111359
Alfarrayeh I, Pollák E, Czéh Á, Vida A, Das S, Papp G. Antifungal and Anti-Biofilm Effects of Caffeic Acid Phenethyl Ester on Different Candida Species. Antibiotics. 2021; 10(11):1359. https://doi.org/10.3390/antibiotics10111359
Chicago/Turabian StyleAlfarrayeh, Ibrahim, Edit Pollák, Árpád Czéh, András Vida, Sourav Das, and Gábor Papp. 2021. "Antifungal and Anti-Biofilm Effects of Caffeic Acid Phenethyl Ester on Different Candida Species" Antibiotics 10, no. 11: 1359. https://doi.org/10.3390/antibiotics10111359
APA StyleAlfarrayeh, I., Pollák, E., Czéh, Á., Vida, A., Das, S., & Papp, G. (2021). Antifungal and Anti-Biofilm Effects of Caffeic Acid Phenethyl Ester on Different Candida Species. Antibiotics, 10(11), 1359. https://doi.org/10.3390/antibiotics10111359