
Research Progress on Antibacterial Activities and Mechanisms of Natural Alkaloids: A Review


Abstract
:1. Introduction
2. Antibacterial Activity of Different Types of Natural Alkaloids
2.1. Isoquinoline Alkaloids
2.2. Pyridine Alkaloids
2.3. Indole Alkaloids
2.4. Steroidal Alkaloids
2.5. Others

{kind=link}
| Alkaloids (Compound Name) | Sources | Structure of the Alkaloids | Strains Inhibited | MIC * | Positive Control | MIC * | References |
|---|---|---|---|---|---|---|---|
| 2-methoxy-4-(2-(2-pyridine)-ethyl) phenol | Zingiberis rhizoma |  | C. albicans | 1.0 mg/mL | - | - | [29] |
| Thalicfoetine | T. foetidum |  | B. subtilis | 3.12 µg/mL | Cefotaxime | 1.56 µg/mL | [24] |
| Spathullin A | P. spathulatum Em19 |  | E. coli, A. baumannii, E. cloacae, S. aureus | 4–15 µg/mL | Meropenem | 0.5–2 µg/mL | [25] |
| Spathullin B | P. spathulatum Em19 |  | E. coli, A. baumannii, E. cloacae, S. aureus | 1–15 µg/mL | [25] | ||
| 16,17,19,20-tetrahydro-2,16-dehydro-18-deoxyisostrychnine | P. pilifera |  | E. coli | 0.781 μg/mL | - | - | [30] |
| tTris(1H-indol-3-yl) methylium | Pseudomonas aeruginosa UWI-1 |  | Gram-positive bacteria, Gram-negative bacteria | 1–128 µg/mL | Kanamycin | 4–128 µg/mL | [43] |
| bis(indol-3-yl) phenylmethane | P. aeruginosa UWI-1 |  | Gram-positive bacteria | 32–128 µg/mL | [43] | ||
| indolo (2,1b) quinazoline-6,12 dione | P. aeruginosa UWI-1 |  | Gram-positive bacteria, Gram-negative bacteria | 1–32 µg/mL | [43] | ||
| Dionemycin | Streptomyces sp. SCSIO 11791 |  | M. luteus, S. aureus, MRSA | 0.5–2 μg/mL | Kanamycin | 1–128 μg/mL | [31] |
| 6-CH3O-7′,7′′-dichorochromopyrrolic acid | Streptomyces sp. SCSIO 11791 |  | M. luteus, S. aureus, MRSA | 3–128 μg/mL | [31] | ||
| Voacafricines A | V. africana |  | S. aureus, S. typhi, B. Subtilis | 3.12–25 μg/mL | Berberine, Fibraurtine | 3.12–25 μg/mL | [32] |
| Voacafricines B | V. africana |  | S. aureus, S. typhi, B. Subtilis, E. coli | 0.78–50 μg/mL | [32] | ||
| Epidihydropinidine | Picea abies (L.) Karsten |  | P. aeruginosa, E. faecalis, C. glabrata, C. albicans, S. enterica, B. cereus, S. aureus | 5.37–43 µg/mL | Amphotericin B | - | [40] |
| Myoporumine A | M. bontioides A. Gray |  | MRSA | 6.25 µg/mL | Fraction F4 | 25 µg/mL | [41] |
| Myoporumine B | M. bontioides A. Gray |  | MRSA | 6.25 µg/mL | [41] | ||
| Palmatine | Coptis chinensis |  | H. pylori | 75–200 μg/mL | Metronidazole | 0.5–2 μg/mL | [44] |
| Chelerythrine | Toddalia asiatica (Linn) Lam |  | S. aureus, MRSA, Extended-spectrum β-lactamases S. aureus | 0.156 mg/mL | - | - | [45] |
| Berberine | T. asiatica (Linn) Lam |  | S. aureus, MRSA, Extended-spectrum β-lactamases S. aureus | 0.0312 mg/mL | - | - | [45] |
| Tetrandrine | Stephania tetrandra S. Moore |  | S. aureus | 125–250 μg/mL | Ampicillin, Oxacillin | 0.9–250 μg/mL | [46] |
| Stachydrine | Ritchiea capparoides var. longipedicellata |  | S. aureus, E. coli | 5 mg/mL | Streptomycin | 0.125 mg/mL | [47] |
| Chabamide | Piper chaba |  | M. tuberculosis | 12.5 μg/mL | - | - | [48] |
| Lycorine | Pancratium Foetidum Pom |  | S. aureus, B. cereus, P. aerugin, E. cloac | 0.24 mg/mL | Streptomycin | 0.04–0.34 mg/mL | [49] |
| Lycorine | Amaryllidaceae |  | C. dubliniensis, C. albicans, L. elongisporus | 32–64 µg/mL | Tetracycline | 0.5–2 µg/mL | [27] |
| Caranine | Amaryllidaceae |  | C. dubliniensis | 128 µg/mL | [27] | ||
| 6-(pyrrolidin-2-yl)DAPG | Pseudomonas protegens UP46 |  | S. aureus, Bacillus cereus | 2–4 µg/mL | - | - | [50] |
| 6-(piperidin-2-yl)DAPG | P. protegens UP46 |  | S. aureus, B. cereus | 2 µg/mL | - | - | [50] |
| Kopsiahainanins A | Kopsia hainanensis |  | S. aureus, S. epidermidis, E. coli, E. cloacae, K. pneumoniae, P. aeruginosa, S. dysenteriae | 0.12–0.23 µg/mL | Netilmicin | 0.004–0.015 µg/mL | [51] |
| Kopsiahainanins B | K. hainanensis |  | S. aureus, S. epidermidis, E. coli, E. cloacae, K. pneumoniae, P. aeruginosa, S. dysenteriae | 0.14–0.26 µg/mL | [51] | ||
| Kopsiahainanins C | K. hainanensis |  | S. aureus, S. epidermidis, E. coli, E. cloacae, K. pneumoniae, P. aeruginosa, S. dysenteriae | 0.94–1.32 µg/mL | [51] | ||
| Kopsiahainanins D | K. hainanensis |  | S. aureus, S. epidermidis, E. coli, E. cloacae, K. pneumoniae, P. aeruginosa, S. dysenteriae | 0.92–1.24 µg/mL | [51] | ||
| Kopsiahainanins E | K. hainanensis |  | E. coli, E. cloacae, K. pneumoniae, P. aeruginosa, S. dysenteriae | 1.19–1.31 µg/mL | [51] | ||
| Kopsiahainanins F | K. hainanensis |  | E. coli, E. cloacae, K. pneumoniae, P. aeruginosa, S. dysenteriae | 0.99–1.32 µg/mL | [51] | ||
| Kuanoniamine D | Ascidian Cystodytes dellechiajei |  | E. coli, M. luteus | 2.2–17.4 µM | Gentamicin | 0.02–0.08 µM | [52] |
| Shermilamine B | Ascidian C. dellechiajei |  | E. coli, M. luteus | 2.0–8.0 µM | [52] | ||
| N-deacetylkuanoniamine D | Ascidian C. dellechiajei |  | E. coli, M. luteus | 2.5 µM | [52] | ||
| N-deacetylshermilamine B | Ascidian C. dellechiajei |  | E. coli, M. luteus | 1.1–4.5 µM | [52] | ||
| 11-hydroxyascididemin | Ascidian C. dellechiajei |  | E. coli, M. luteus | 2.6–10.5 µM | [52] | ||
| Cystodimine A | Ascidian C. dellechiajei |  | E. coli, M. luteus | 1.2–2.4 µM | [52] | ||
| Cystodimine B | Ascidian C. dellechiajei |  | E. coli, M. luteus | 2.6–10.5 µM | [52] | ||
| Ascididemin | Ascidian C. dellechiajei |  | E. coli, M. luteus | 0.2–0.3 µM | [52] | ||
| Sophoridine | Thermopsis lanceolata R.Brown |  | E. coli, E. aerogenes, P. vulgaris, B. subtilis, S. epidermidis | 2 × 10−2–4 × 10−2 M | - | - | [53] |
| Sophoramine | T. lanceolata R.Brown |  | E. coli, E. aerogenes, P. vulgaris, B. subtilis, S. epidermidis | 4 × 10−2–5 × 10−2 M | - | - | [53] |
| Matrine | T. lanceolata R.Brown |  | E. coli, E. aerogenes, P. vulgaris, B. subtilis, S. epidermidis | 2 × 10−2–5 × 10−2 M | - | - | [53] |
| Cytisine | T. lanceolata R.Brown |  | E. coli, E. aerogenes, P. vulgaris, B. subtilis, S. epidermidis | 3 × 10−2–5 × 10−2 M | - | - | [53] |
| Oxymatrine | T. lanceolata R.Brown |  | E. coli, E. aerogenes, P. vulgaris, B. subtilis, S. epidermidis | 5 × 10−2 M | - | - | [53] |
| Berberine | Berberis vulgaris |  | T. mentagrophytes, T. rubrum, M. canis, M. gypseum | 0.062–0.250 mg/mL | Ketoconazole | 0.125–0.250 mg/mL | [54] |
| Chelerythrine | C. majus |  | P. aeruginosa | 1.9 mg/L | - | - | [26] |
| Sanguinarine | C. majus |  | S. aureus | 1.9 mg/L | - | - | [26] |
| Chelidonine | C. majus |  | C. albicans | 62.5 mg/L | - | - | [26] |
| Berberine | C. majus |  | S. aureus | 125 mg/L | - | - | [26] |
| Allocryptopine | C. majus |  | S. aureus | 125 mg/L | - | - | [26] |
| Mokluangins B | Holarrhena pubescens |  | B. subtilis, E. coli | 16 µg/mL | Vancomycin, Gentamycin | 0.125–0.25 µg/mL | [38] |
| Mokluangins C | H. pubescens |  | E. coli | 16 µg/mL | [38] | ||
| Ageliferin | Sponge A. dilatata |  | A. baumannii, K. pneumoniae, P. aeruginosa | 64 ≥ 128 mg/L | - | - | [42] |
| Bromoageliferin | Sponge A. dilatata |  | A. baumannii, K. pneumoniae, P. aeruginosa | 8 ≥ 128 mg/L | - | - | [42] |
| Dibromoageliferin | Sponge A. dilatata |  | A. baumannii, K. pneumoniae, P. aeruginosa | 32 ≥ 128 mg/L | - | - | [42] |
| Sceptrin | Sponge A. dilatata |  | A. baumannii, K. pneumoniae, P. aeruginosa | 64 ≥ 128 mg/L | - | - | [42] |
| Nakamuric acid | Sponge A. dilatata |  | A. baumannii, K. pneumoniae, P. aeruginosa | ≥128 mg/L | - | - | [42] |
| 4-Bromo-1H-pyrrole-2-carboxylic acid | Sponge A. dilatata |  | A. baumannii, K. pneumoniae, P. aeruginosa | 64 ≥ 128 mg/L | - | - | [42] |
| 4,5-Dibromopyrrole-2-carboxylic acid | Sponge A. dilatata |  | A. baumannii, K. pneumoniae, P. aeruginosa | 64 ≥ 128 mg/L | - | - | [42] |
| 3,7-Dimethylisoguanine | Sponge A. dilatata |  | A. baumannii, K. pneumoniae, P. aeruginosa | 64 ≥ 128 mg/L | - | - | [42] |
| 9H-carbazole | Myxobacterium L. luteola |  | C. albicans, B. subtilis, E. coli, C. violaceum | 6.7–33.3 µg/mL | Methanol | - | [55] |
| 3-chloro-9H-carbazole | Myxobacterium L. luteola |  | C. albicans | 33.3 µg/mL | [55] | ||
| 4-hydroxymethyl-quinoline | Myxobacterium L. luteola |  | C. albicans | 33.3 µg/mL | [55] | ||
| Latifolianine A | Nauclea latifolia |  | H. influenzae | 25 µg/mL | Ciprofloxacin | 1.6 µg/mL | [56] |
| Latifoliaindole A | N. latifolia |  | H. influenzae | 50 µg/mL | [56] | ||
| Latifoliaindole B | N. latifolia |  | H. influenzae | 25 | [56] | ||
| Neoechinulin A | Eurotium sp. |  | B. cereus, P. vulgaris | 6.25–25 μM | Ciprofloxacin | 0.20–0.78 μM | [57] |
| l-alanyl-l-tryptophan anhydride | Eurotium sp. |  | B. cereus, P. vulgaris | 1.56–3.13 μM | [57] | ||
| Dihydroxyisoechinulin A | Eurotium sp. |  | B. cereus | 3.13 μM | [57] | ||
| Terpendole L | Tolypocladium sp. |  | M. lysodeikticus, M. luteus | 6.25–50 μg/mL | Ciprofloxacin | 0.78 μg/mL | [58] |
| Tolypocladin A | Tolypocladium sp. |  | B. cereus, MRSA | 12.5–25 μg/mL | Ketoconazole | 0.78 μg/mL | [59] |
| Tolypocladin B | Tolypocladium sp. |  | MRSA | 50 μg/mL | [59] | ||
| Tolypocladin H | Tolypocladium sp. |  | B. cereus, M. lysodeikticus, B. paratyphosum, B. subtilis, E. aerogenes, S. typhi, P. vulgaris | 0.78–1.56 μg/mL | [59] | ||
| Alstoniascholarine A | Alstonia scholaris |  | P. aeruginosa, K. pneumoniae, E. coli, E. faecalis | 25–50 μg/mL | Gentamycin, Griseofulvin | 0.20–7.81 μg/mL | [60] |
| Alstoniascholarine C | A. scholaris |  | P. aeruginosa, K. pneumoniae, E. coli, E. faecalis | 12.5–50 μg/mL | [60] | ||
| Alstoniascholarine E | A. scholaris |  | P. aeruginosa, K. pneumoniae, E. coli, E. faecalis | 25–50 μg/mL | [60] | ||
| Alstoniascholarine F | A. scholaris |  | P. aeruginosa, K. pneumoniae, E. coli, E. faecalis | 3.13–50 μg/mL | [60] | ||
| Alstoniascholarine H | A. scholaris |  | P. aeruginosa, K. pneumoniae, E. coli | 25–50 μg/mL | [60] | ||
| Alstoniascholarine I | A. scholaris |  | P. aeruginosa, K. pneumoniae, E. coli | 12.5–25 μg/mL | [60] | ||
| Alstoniascholarine J | A. scholaris |  | S. aureus, P. aeruginosa, E. faecalis, K. pneumoniae, E. coli | 3.13–25 μg/mL | [60] | ||
| Scholarisine T | A. scholaris |  | E. coli, B. subtilis, S. typhi | 0.78–12.5 μg/mL | Cefotaxime | 0.39–3.12 μg/mL | [61] |
| Scholarisine U | A. scholaris |  | E. coli, B. subtilis | 0.78–3.12 μg/mL | [61] | ||
| Scholarisine V | A. scholaris |  | E. coli, S. aureus, S. typhi | 0.78–12.5 μg/mL | [61] | ||
| 2, 5, 6-tribromo-3-[(3′-bromo-4′-hydroxyl-phenyl)-methyl]-1H-indole | Laurencia similis |  | B. subtilis, B. thuringensis, S. aureus, A. tumefaciens, P. lachrymans, R. solanacearum, X. vesicatoria | 2–8 μg/mL | Penicillin | 0.125–250 μg/mL | [62] |
| 5, 6-dibromo-1-hydroxy-3-isopropenyl-indole-2-one | L. similis |  | S. aureus, A. tumefaciens | 12.5 μg/mL | [62] | ||
| Neofiscalin A | Neosartorya siamensis KUFA 0017 |  | MRSA, VRE | 8 μg/mL | Oxacillin | 128 μg/mL | [63] |
| Dragmacidin G | Genus Spongosorites |  | S. aureus, MRSA | 0.62 µg/mL | Chloramphenicol | 3.1–6.2 µg/mL | [64] |
| Chaetoglobinol A | Chaetomium globosum |  | B. subtilis | 50 µg/mL | - | - | [65] |
| Phutdonginin | Kopsia arborea Blume |  | E. coli | 32 g/mL | - | - | [66] |
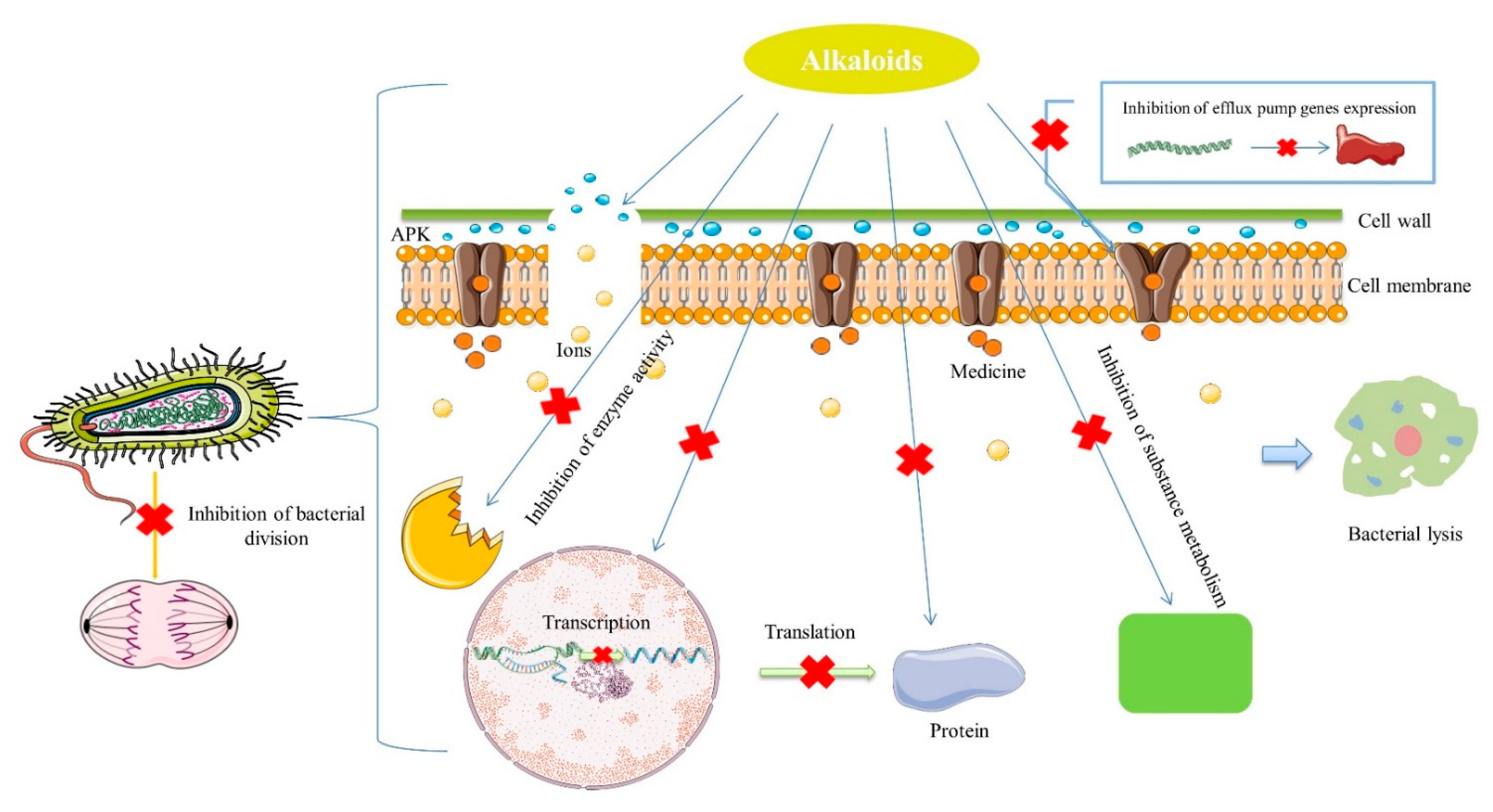
3. Antibacterial Mechanisms of Natural Alkaloids
3.1. Inhibition of Bacterial Nucleic Acid and Protein Synthesis
3.2. Effect on the Bacterial Cell Membrane Permeability—Damage of Cell Membrane and Cell Wall
3.3. Inhibition of Efflux Pumps
3.4. Inhibition of Bacterial Metabolism
3.5. Other Mechanisms
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shen, P.; Chen, X.D. Microbiology; Higher Education Press: Beijing, China, 2016. [Google Scholar]
- Li, F.; Xu, Z.K. Medical Microbiology; People’s Health Publishing House: Beijing, China, 2018. [Google Scholar]
- Zhang, Z.P. New progress in the study of antimicrobial agents. Chin. J. Clin. Pharmacol. 2000, 139–145. [Google Scholar] [CrossRef]
- Zaman, S.B.; Hussain, M.A.; Nye, R.; Mehta, V.; Mamun, K.T.; Hossain, N. A Review on Antibiotic Resistance: Alarm Bells are Ringing. Cureus 2017, 9, e1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.; Mumtaz, S.; Li, C.H.; Hussain, I.; Rotello, V.M. Combatting antibiotic-resistant bacteria using nanomaterials. Chem. Soc. Rev. 2019, 48, 415–427. [Google Scholar] [CrossRef]
- Bate, P.N.N.; Orock, A.E.; Nyongbela, K.D.; Babiaka, S.B.; Kukwah, A.; Ngemenya, M.N. In vitro activity against multi-drug resistant bacteria and cytotoxicity of lichens collected from Mount Cameroon. J. King Saud Univ. Sci. 2020, 32, 614–619. [Google Scholar] [CrossRef]
- Moloney, M.G. Natural Products as a Source for Novel Antibiotics. Trends Pharmacol. Sci. 2016, 37, 689–701. [Google Scholar] [CrossRef]
- Luo, Y.M. Natural Medicinal Chemistry; Huazhong University of Science and Technology: Wuhan, China, 2011. [Google Scholar]
- Qing, Z.-X.; Huang, J.L.; Yang, X.Y.; Liu, J.H.; Cao, H.L.; Xiang, F.; Cheng, P.; Zeng, J.G. Anticancer and Reversing Multidrug Resistance Activities of Natural Isoquinoline Alkaloids and their Structure-activity Relationship. Curr. Med. Chem. 2018, 25, 5088–5114. [Google Scholar] [CrossRef] [PubMed]
- Gorpenchenko, T.Y.; Grigorchuk, V.P.; Bulgakov, D.V.; Tchernoded, G.K.; Bulgakov, V.P. Tempo-Spatial Pattern of Stepharine Accumulation in Stephania Glabra Morphogenic Tissues. Int. J. Mol. Sci. 2019, 20, 808. [Google Scholar] [CrossRef] [Green Version]
- Souza, C.R.M.; Bezerra, W.P.; Souto, J.T. Marine Alkaloids with Anti-Inflammatory Activity: Current Knowledge and Future Perspectives. Mar Drugs. 2020, 18, 147. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Cui, Y.; Lu, L.; Gong, Y.; Han, W.; Piao, G. Natural indole-containing alkaloids and their antibacterial activities. Arch. Pharm. 2020, 353, e2000120. [Google Scholar] [CrossRef]
- Rodrigues, S.; Shin, D.; Conway, M.; Smulski, S.; Trenker, E.; Shanthanna, H.; Vanniyasingam, T.; Thabane, L.; Paul, J. Hydromorphone versus morphine: A historical cohort study to evaluate the quality of postoperative analgesia. Can. J. Anesth. 2020, 68, 226–234. [Google Scholar] [CrossRef]
- Wang, S.; Dong, Y.; Su, K.; Zhang, J.; Wang, L.; Han, A.; Wen, C.; Wang, X.; He, Y. Effect of codeine on CYP450 isoform activity of rats. Pharm. Biol. 2017, 55, 1223–1227. [Google Scholar] [CrossRef] [Green Version]
- Rehman, S.; Khan, H. Advances in Antioxidant Potential of Natural Alkaloids. Curr. Bioact. Compd. 2017, 13, 101–108. [Google Scholar] [CrossRef]
- Kuang, Z.; Zhang, X.; Ma, J.; Li, J.; Zhou, L. Effects of Coptis chinensis Franch alkaloids on plants and microorganisms: A Review. Allelopath. J. 2018, 43, 139–157. [Google Scholar] [CrossRef]
- Li, N.; Tan, S.-N.; Cui, J.; Guo, N.; Wang, W.; Zu, Y.-G.; Jin, S.; Xu, X.-X.; Liu, Q.; Fu, Y.-J. PA-1, a novel synthesized pyrrolizidine alkaloid, inhibits the growth of Escherichia coli and Staphylococcus aureus by damaging the cell membrane. J. Antibiot. 2014, 67, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Larghi, E.; Bracca, A.; Aguilar, A.A.; Heredia, D.; Pergomet, J.; Simonetti, S.; Kaufman, T. Neocryptolepine: A Promising Indoloisoquinoline Alkaloid with Interesting Biological Activity. Evaluation of the Drug and its Most Relevant Analogs. Curr. Top. Med. Chem. 2015, 15, 1683–1707. [Google Scholar] [CrossRef]
- Kelley, C.; Lu, S.; Parhi, A.; Kaul, M.; Pilch, D.S.; Lavoie, E.J. Antimicrobial activity of various 4- and 5-substituted 1-phenylnaphthalenes. Eur. J. Med. Chem. 2013, 60, 395–409. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Ha, L.N.; Wang, X.Q.; Yang, F.Q.; Zhang, A.J.; Zhao, Q.G. In vitro antibacterial effect of Matrine on methicillin resistant Staphylococcus aureus. J. Changzhi Med. Coll. 2012, 26, 161–163. [Google Scholar] [CrossRef]
- Qiu, S.; Sun, H.; Zhang, A.H.; Xu, H.Y.; Yan, G.L.; Han, Y.; Wang, X.J. Natural alkaloids: Basic aspects, biological roles, and future perspectives. Chin. J. Nat. Med. 2014, 12, 401–406. [Google Scholar] [CrossRef]
- Bai, Y.J.; Yu, M.; Zhao, S.W.; Chen, Y.X.; Chang, K.L. Pharmacological action and mechanism of alkaloids. J. Harbin Univ. Com. 2013, 29, 8–11. [Google Scholar] [CrossRef]
- Hagel, J.M.; Facchini, P.J. Benzylisoquinoline Alkaloid Metabolism: A Century of Discovery and a Brave New World. Plant Cell Physiol. 2013, 54, 647–672. [Google Scholar] [CrossRef] [Green Version]
- Ding, C.F.; Qin, X.J.; Yu, H.F.; Liu, Y.P.; Wang, X.H.; Luo, X.D. Thalicfoetine, a novel isoquinoline alkaloid with antibacterial activity from Thalictrum foetidum. Tetrahedron Lett. 2019, 60. [Google Scholar] [CrossRef]
- Nord, C.; Levenfors, J.J.; Bjerketorp, J.; Sahlberg, C.; Guss, B.; Öberg, B.; Broberg, A. Antibacterial Isoquinoline Alkaloids from the Fungus Penicillium Spathulatum Em19. Molecules 2019, 24, 4616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielińska, S.; Wójciak-Kosior, M.; Dziągwa-Becker, M.; Gleńsk, M.; Sowa, I.; Fijałkowski, K.; Rurańska-Smutnicka, D.; Matkowski, A.; Junka, A. The Activity of Isoquinoline Alkaloids and Extracts from Chelidonium majus against Pathogenic Bacteria and Candida sp. Toxins 2019, 11, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ločárek, M.; Nováková, J.; Klouček, P.; Hošt’álková, A.; Kokoška, L.; Gábrlová, L.; Šafratová, M.; Opletal, L.; Cahlíková, L. Antifungal and Antibacterial Activity of Extracts and Alkaloids of Selected Amaryllidaceae Species. Nat. Prod. Commun. 2015, 10, 1537–1540. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hu, W.Z.; Chen, X.Z.; Peng, Y.B.; Song, L.Y.; Shi, Y.S. Bioactive quinolizidine alkaloids from the root of Sophora tonkinensis. J. Tradit. Chin. Med. 2016, 41, 2261–2266. [Google Scholar] [CrossRef]
- Shao, F.; Wang, D.Q.; Xiong, W.; Zhang, P.Z.; Ma, G.Q.; Liu, R.H.; Yao, X.L. A new pyridine alkaloid from Zingiberis rhizoma. Nat. Prod. Res. 2017, 31, 1486–1489. [Google Scholar] [CrossRef]
- Liu, L.; Song, C.W.; Khan, A.; Li, X.N.; Yang, X.W.; Cheng, G.G.; Liu, Y.P.; Luo, X.D. A potent antibacterial indole alkaloid from Psychotria pilifera. J. Asian Nat. Prod. Res. 2016, 18, 798–803. [Google Scholar] [CrossRef]
- Song, Y.; Yang, J.; Yu, J.; Li, J.; Yuan, J.; Wong, N.-K.; Ju, J. Chlorinated bis-indole alkaloids from deep-sea derived Streptomyces sp. SCSIO 11791 with antibacterial and cytotoxic activities. J. Antibiot. 2020, 73, 542–547. [Google Scholar] [CrossRef]
- Ding, C.F.; Ma, H.X.; Yang, J.; Qin, X.J.; Njateng, G.S.S.; Yu, H.F.; Wei, X.; Liu, Y.P.; Huang, W.Y.; Yang, Z.F.; et al. Antibacterial Indole Alkaloids with Complex Heterocycles from Voacanga africana. Org. Lett. 2018, 20, 2702–2706. [Google Scholar] [CrossRef] [PubMed]
- Li, H.J.; Jiang, Y.; Li, P. Chemistry, bioactivity and geographical diversity of steroidal alkaloids from the Liliaceae family. Nat. Prod. Rep. 2006, 23, 735–752. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.U.; Choudhary, M.I. Diterpenoid and steroidal alkaloids. Nat. Prod. Rep. 1995, 12, 361. [Google Scholar] [CrossRef]
- Rahman, A.; Choudhary, M.I. Chemistry and biology of steroidal alkaloids. Alkaloids Chem. Biol. 1998, 50, 61–108. [Google Scholar]
- Zhou, L.N.; Ge, X.L.; Dong, T.T.; Gao, H.Y.; Sun, B.H. Antibacterial steroidal alkaloids from Holarrhena antidysenteriaca. Chin. J. Nat. Med. 2017, 15, 540–545. [Google Scholar] [CrossRef]
- Nantachit, K.; Roongjang, S. Anti-mycobacterium and Anti-cancer Activities of Combretin, an Isolated Steroidal Alkaloid from the Seeds of Combretum quadrangulare Kurz. J. Pharm. Pharmacol. 2016, 4, 261–267. [Google Scholar] [CrossRef]
- Cheenpracha, S.; Jitonnom, J.; Komek, M.; Ritthiwigrom, T.; Laphookhieo, S. Acetylcholinesterase inhibitory activity and molecular docking study of steroidal alkaloids from Holarrhena pubescens barks. Steroids 2016, 108, 92–98. [Google Scholar] [CrossRef]
- Siriyong, T.; Voravuthikunchai, S.P.; Coote, P.J. Steroidal alkaloids and conessine from the medicinal plant Holarrhena antidysenterica restore antibiotic efficacy in a Galleria mellonella model of multidrug-resistant Pseudomonas aeruginosa infection. BMC Complement. Altern. Med. 2018, 18, 285. [Google Scholar] [CrossRef] [Green Version]
- Fyhrquist, P.; Virjamo, V.; Hiltunen, E.; Julkunen-Tiitto, R. Epidihydropinidine, the main piperidine alkaloid compound of Norway spruce (Picea abies) shows promising antibacterial and anti-Candida activity. Fitoterapia 2017, 117, 138–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.M.; Huang, L.L.; Dai, H.; Xu, Q.L.; Ouyang, J.K.; Jia, X.C.; Gu, W.X.; Tan, J.W. Anti-MRSA Sesquiterpenes from the Semi-Mangrove Plant Myoporum bontioides A. Gray. Mar. Drugs 2018, 16, 438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pech-Puch, D.; Pérez-Povedano, M.; Martinez-Guitian, M.; Lasarte-Monterrubio, C.; Vázquez-Ucha, J.C.; Bou, G.; Rodríguez, J.; Beceiro, A.; Jimenez, C. In Vitro and In Vivo Assessment of the Efficacy of Bromoageliferin, an Alkaloid Isolated from the Sponge Agelas dilatata, against Pseudomonas aeruginosa. Mar. Drugs 2020, 18, 326. [Google Scholar] [CrossRef] [PubMed]
- Ramkissoon, A.; Seepersaud, M.; Maxwell, A.; Jayaraman, J.; Ramsubhag, A. Isolation and Antibacterial Activity of Indole Alkaloids from Pseudomonas aeruginosa UWI-1. Molecules 2020, 25, 3744. [Google Scholar] [CrossRef]
- Zhou, J.T.; Li, C.L.; Tan, L.H.; Xu, Y.F.; Liu, Y.H.; Mo, Z.Z.; Dou, Y.X.; Su, R.; Su, Z.R.; Huang, P.; et al. Inhibition of Helicobacter pylori and Its Associated Urease by Palmatine: Investigation on the Potential Mechanism. PLoS ONE 2017, 12, e0168944. [Google Scholar] [CrossRef] [Green Version]
- He, N.; Wang, P.; Wang, P.; Ma, C.; Kang, W. Antibacterial mechanism of chelerythrine isolated from root of Toddalia asiatica (Linn) Lam. BMC Complement. Altern. Med. 2018, 18, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Han, S.H.; Lee, S.H.; Kim, Y.G.; Park, C.B.; Kang, O.H.; Keum, J.H.; Kim, S.B.; Mun, S.H.; Seo, Y.S.; et al. The Mechanism of Antibacterial Activity of Tetrandrine AgainstStaphylococcus aureus. Foodborne Pathog. Dis. 2012, 9, 686–691. [Google Scholar] [CrossRef]
- Taiwo, B.J.; Akinkunmi, E.O.; Omisore, N. Antimicrobial and antiplasmodial activities of a quaternary compound from Ritchiea capparoides var. Longipedicellata. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 528–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rukachaisirikul, T.; Prabpai, S.; Champung, P.; Suksamrarn, A. Chabamide, a Novel Piperine Dimer from Stems of Piper chaba. Planta Med. 2002, 68, 853–855. [Google Scholar] [CrossRef] [PubMed]
- Bendaif, H.; Melhaoui, A.; Ramdani, M.; Elmsellem, H.; Douez, C.; El Ouadi, Y. Antibacterial activity and virtual screening by molecular docking of lycorine from Pancratium foetidum Pom (Moroccan endemic Amaryllidaceae). Microb. Pathog. 2018, 115, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Levenfors, J.J.; Nord, C.; Bjerketorp, J.; Ståhlberg, J.; Larsson, R.; Guss, B.; Öberg, B.; Broberg, A. Antibacterial pyrrolidinyl and piperidinyl substituted 2,4-diacetylphloroglucinols from Pseudomonas protegens UP46. J. Antibiot. 2020, 73, 739–747. [Google Scholar] [CrossRef]
- Chi, W.Q.; Jiang, Y.H.; Hu, J.; Pan, J. Cytotoxic and antibacterial aspidofractinine alkaloids from Kopsia hainanensis. Fitoterapia 2018, 130, 259–264. [Google Scholar] [CrossRef]
- Bontemps, N.; Bry, D.; López-Legentil, S.; Simon-Levert, A.; Long, C.; Banaigs, B. Structures and Antimicrobial Activities of Pyridoacridine Alkaloids Isolated from Different Chromotypes of the AscidianCystodytes dellechiajei. J. Nat. Prod. 2010, 73, 1044–1048. [Google Scholar] [CrossRef]
- Xia, L.Q.; Zhao, B.G.; Ju, Y.W.; Dai, L. Inhibitory effect of bis (piperidine) alkaloids on five environmental bacterial strains. J. Nanjing For. Univ. 2001, 81–84. [Google Scholar] [CrossRef]
- Mahmoudvand, H.; Mousavi, S.A.A.; Sepahvand, A.; Sharififar, F.; Ezatpour, B.; Gorohi, F.; Dezaki, E.S.; Jahanbakhsh, S. Antifungal, Antileishmanial, and Cytotoxicity Activities of Various Extracts of Berberis vulgaris (Berberidaceae) and Its Active Principle Berberine. ISRN Pharmacol. 2014, 2014, 602436. [Google Scholar] [CrossRef]
- Mulwa, L.S.; Jansen, R.; Praditya, D.F.; Mohr, K.I.; Wink, J.; Steinmann, E.; Stadler, M. Six Heterocyclic Metabolites from the Myxobacterium Labilithrix luteola. Molecules 2018, 23, 542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankeu, J.J.K.; Kagho, D.U.K.; Fotsing, Y.S.F.; Toghueo, R.M.K.; Mba’Ning, B.M.; Feuya, G.R.T.; Fekam, F.B.; Tchouankeu, J.C.; Ngouela, S.A.; Sewald, N.; et al. Constituents from Nauclea latifolia with Anti-Haemophilus influenzae Type b Inhibitory Activities. J. Nat. Prod. 2019, 82, 2580–2585. [Google Scholar] [CrossRef]
- Zhao, D.; Cao, F.; Guo, X.J.; Zhang, Y.R.; Kang, Z.; Zhu, H.J. Antibacterial Indole Alkaloids and Anthraquinones from a Sewage-Derived Fungus Eurotium sp. Chem. Nat. Compd. 2018, 54, 399–401. [Google Scholar] [CrossRef]
- Xu, L.L.; Pang, X.J.; Shi, Q.; Xian, P.J.; Tao, Y.D.; Yang, X.L. Two New Prenylated Indole Diterpenoids from Tolypocladium sp. and Their Antimicrobial Activities. Chem. Biodivers. 2019, 16, e1900116. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.L.; Hai, P.; Zhang, S.B.; Xiao, J.F.; Gao, Y.; Ma, B.J.; Fu, H.Y.; Chen, Y.M.; Yang, X.L. Prenylated Indole Diterpene Alkaloids from a Mine-Soil-Derived Tolypocladium sp. J. Nat. Prod. 2019, 82, 221–231. [Google Scholar] [CrossRef]
- Qin, X.J.; Zhao, Y.L.; Lunga, P.K.; Yang, X.W.; Song, C.W.; Cheng, G.G.; Liu, L.; Chen, Y.Y.; Liu, Y.P.; Luo, X.D. Indole alkaloids with antibacterial activity from aqueous fraction of Alstonia scholaris. Tetrahedron 2015, 71, 4372–4378. [Google Scholar] [CrossRef]
- Yu, H.F.; Huang, W.Y.; Ding, C.F.; Wei, X.; Zhang, L.C.; Qin, X.J.; Ma, H.X.; Yang, Z.F.; Liu, Y.P.; Zhang, R.P.; et al. Cage-like monoterpenoid indole alkaloids with antimicrobial activity from Alstonia scholaris. Tetrahedron Lett. 2018, 59, 2975–2978. [Google Scholar] [CrossRef]
- Li, M.C.; Sun, W.S.; Cheng, W.; Liu, N.; Liang, H.; Zhang, Q.Y.; Lin, W.H. Four new minor brominated indole related alkaloids with antibacterial activities from Laurencia similis. Bioorg. Med. Chem. Lett. 2016, 26, 3590–3593. [Google Scholar] [CrossRef]
- Bessa, L.J.; Buttachon, S.; Dethoup, T.; Martins, R.; Vasconcelos, V.; Kijjoa, A.; Da Costa, P.M. Neofiscalin A and fiscalin C are potential novel indole alkaloid alternatives for the treatment of multidrug-resistant Gram-positive bacterial infections. FEMS Microbiol. Lett. 2016, 363, fnw150. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.E.; Killday, K.B.; Chakrabarti, D.; Guzmán, E.A.; Harmody, D.; McCarthy, P.J.; Pitts, T.; Pomponi, S.A.; Reed, J.K.; Roberts, B.F.; et al. Dragmacidin G, a Bioactive Bis-Indole Alkaloid from a Deep-Water Sponge of the Genus Spongosorites. Mar. Drugs 2017, 15, 16. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.B.; He, G.; Bai, H.H.; Yang, T.; Zhang, G.L.; Wu, L.W.; Li, G.Y. Indole Alkaloids fromChaetomium globosum. J. Nat. Prod. 2015, 78, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Cheenpracha, S.; Raksat, A.; Ritthiwigrom, T.; Laphookhieo, S. Monoterpene indole alkaloids from the twigs of Kopsia arborea. Nat. Prod. Commun. 2014, 9, 1441. [Google Scholar] [CrossRef] [Green Version]
- Elisabetsky, E.; Costa-Campos, L. The Alkaloid Alstonine: A Review of Its Pharmacological Properties. Evid.-Based Complement. Altern. Med. 2006, 3, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Lyu, Y.; Huang, J.; Zhang, X.; Yu, N.; Wen, Z.; Chen, S. Antibacterial activity and mechanism of sanguinarine against Providencia rettgeri in vitro. PeerJ 2020, 8, e9543. [Google Scholar] [CrossRef] [PubMed]
- Salton, M.R. Structure and function of bacterial cell membranes. Annu. Rev. Microbiol. 1967, 21, 417–442. [Google Scholar] [CrossRef] [PubMed]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial Activity of Polyphenols and Alkaloids in Middle Eastern Plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef] [PubMed]
- Barrows, J.M.; Goley, E.D. FtsZ dynamics in bacterial division: What, how, and why?—ScienceDirect. Curr. Opin. Cell Biol. 2021, 68, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Petronio, G.P.; Cutuli, M.A.; Magnifico, I.; Venditti, N.; Pietrangelo, L.; Vergalito, F.; Pane, A.; Scapagnini, G.; Di Marco, R. In Vitro and In Vivo Biological Activity of Berberine Chloride against Uropathogenic E. coli Strains Using Galleria mellonella as a Host Model. Molecules 2020, 25, 5010. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, K.; Moriyasu, M.; Yamori, T.; Turuo, T.; Lee, D.U.; Wiegrebe, W. In Vitro Cytotoxicity of the Protoberberine-Type Alkaloids. J. Nat. Prod. 2001, 64, 896–898. [Google Scholar] [CrossRef]
- Jin, J.L.; Hua, G.Q.; Meng, Z.; Gao, P.J. Antibacterial Mechanisms of Berberine and Reasons for Little Resistance of Bacteria. Chin. Herb. Med. 2010, 3, 27–35. [Google Scholar] [CrossRef]
- Imenshahidi, M.; Hosseinzadeh, H. Berberine and barberry (Berberis vulgaris): A clinical review. Phytother. Res. 2019, 33, 504–523. [Google Scholar] [CrossRef]
- Pasqual, M.S.; Lauer, C.P.; Moyna, P.; Henriques, J.A.P. Genotoxicity of the isoquinoline alkaloid berberine in prokaryotic and eukaryotic organisms. Mutat. Res. Mol. Mech. Mutagen. 1993, 286, 243–252. [Google Scholar] [CrossRef]
- Ortiz, L.M.G.; Lombardi, P.; Tillhon, M.; Scovassi, A.I. Berberine, an Epiphany against Cancer. Molecules 2014, 19, 12349–12367. [Google Scholar] [CrossRef]
- Rahman, M.U.; Wang, P.; Wang, N.; Chen, Y. The key bacterial cell division protein FtsZ as a novel antibacterial drug target. Bosn. J. Basic Med. Sci. 2020, 20. [Google Scholar] [CrossRef]
- Beuria, T.K.; Santra, A.M.K.; Panda, D. Sanguinarine Blocks Cytokinesis in Bacteria by Inhibiting FtsZ Assembly and Bundling. Biochemistry 2005, 44, 16584–16593. [Google Scholar] [CrossRef]
- Chen, D.H. In vitro antibacterial activity of alkaloids from Sophora flavescens. Chin. Anim. Health 2010, 12, 28–30. [Google Scholar] [CrossRef]
- Boberek, J.M.; Stach, J.; Good, L. Genetic Evidence for Inhibition of Bacterial Division Protein FtsZ by Berberine. PLoS ONE 2010, 5, e13745. [Google Scholar] [CrossRef] [PubMed]
- Mabhiza, D.; Chitemerere, T.; Mukanganyama, S. Antibacterial Properties of Alkaloid Extracts from Callistemon citrinus and Vernonia adoensis against Staphylococcus aureus and Pseudomonas aeruginosa. Int. J. Med. Chem. 2016, 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Qian, L.H.; Tao, Y.; Xie, J. Antibacterial mechanism of tea polyphenols against Staphylococcus aureus and Pseudomonas aeruginosa. Microbiol. China 2010, 37, 1628–1633. [Google Scholar]
- Lan, W.Q.; Xie, J.; Hou, W.F.; Li, D.W. Antibacterial activity and mechanism of compound biological preservatives against Staphylococcus squirrel. Res. Dve. Nat. Prod. 2012, 24, 741–746, 753. [Google Scholar] [CrossRef]
- Hara, S.; Yamakawa, M. Moricin, a Novel Type of Antibacterial Peptide Isolated from the Silkworm, Bombyx mori. J. Biol. Chem. 1995, 270, 29923–29927. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Li, X.D.; Hong, J.; Liu, C.; Zhang, X.L.; Zheng, J.-P.; Xu, Y.J.; Ou, Z.Y.; Zheng, J.L.; Yu, D.J. Inhibitory Effect of Two Traditional Chinese Medicine Monomers, Berberine and Matrine, on the Quorum Sensing System of Antimicrobial-Resistant Escherichia coli. Front. Microbiol. 2019, 10, 2584. [Google Scholar] [CrossRef]
- Wang, T.P.; Gao, Y.X.; Zhao, Q.; Yuan, Y.J.; Ma, W.C.; Xue, L. Preliminary study on the antibacterial effect and mechanism of alkaloids from Rabdosia Rubra on Klebsiella pneumoniae. J. Tianshui Norm. Univ. 2018, 38, 24–28. [Google Scholar]
- Dvorak, Z.; Sovadinova, I.; Bláha, L.; Giesy, J.P.; Ulrichová, J. Quaternary benzo[c]phenathridine alkaloids sanguinarine and chelerythrine do not affect transcriptional activity of aryl hydrocarbon receptor: Analyses in rat hepatoma cell line H4IIE.luc. Food Chem. Toxicol. 2006, 44, 1466–1473. [Google Scholar] [CrossRef]
- Lei, Q.; Liu, H.; Peng, Y.; Xiao, P. In silico target fishing and pharmacological profiling for the isoquinoline alkaloids of Macleaya cordata (Bo Luo Hui). Chin. Med. 2015, 10, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teelucksingh, T.; Thompson, L.K.; Cox, G. The Evolutionary Conservation of Escherichia coli Drug Efflux Pumps Supports Physiological Functions. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef]
- Wei, J.T.; Qian, J.; Su, Q.L.Y.; Liu, Z.X.; Wang, X.L.; Wang, Y.P. Research Progress on the mechanism of bacterial biofilm induced drug resistance and the effect of antimicrobial peptide LL-37 on biofilm. J. Hexi Univ. 2020, 36, 38–43. [Google Scholar]
- Ito, A.; Taniuchi, A.; May, T.; Kawata, K.; Okabe, S. Increased Antibiotic Resistance of Escherichia coli in Mature Biofilms. Appl. Environ. Microbiol. 2009, 75, 4093–4100. [Google Scholar] [CrossRef] [Green Version]
- Maesaki, S.; Marichal, P.; Bossche, H.V.; Sanglard, D.; Kohno, S. Rhodamine 6G efflux for the detection of CDR1-overexpressing azole-resistant Candidaalbicans strains. J. Antimicrob. Chemother. 1999, 44, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, Y.; Wang, X.; Guo, J.; Wang, H.; Zhang, H.; Du, F. Jatrorrhizine suppresses the antimicrobial resistance of methicillin-resistant Staphylococcus aureus. Exp. Ther. Med. 2019, 18, 3715–3722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohtar, M.; Johari, S.A.; Li, A.R.; Isa, M.M.; Mustafa, S.; Ali, A.M.; Basri, D.F. Inhibitory and Resistance-Modifying Potential of Plant-Based Alkaloids Against Methicillin-Resistant Staphylococcus aureus (MRSA). Curr. Microbiol. 2009, 59, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Stavri, M.; Piddock, L.J.V.; Gibbons, S. Bacterial efflux pump inhibitors from natural sources. J. Antimicrob. Chemother. 2007, 59, 1247–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, R.A.; Hooper, D.C. Efflux Pumps and Nosocomial Antibiotic Resistance: A Primer for Hospital Epidemiologists. Clin. Infect. Dis. 2005, 40, 1811–1817. [Google Scholar] [CrossRef]
- Sobti, M.; Ishmukhametov, R.; Stewart, A.G. ATP Synthase: Expression, Purification, and Function. Methods Mol. Biol. 2020, 2073, 73–84. [Google Scholar] [CrossRef]
- Du, G.F.; Le, Y.J.; Sun, X.; Yang, X.Y.; He, Q.Y. Proteomic investigation into the action mechanism of berberine against Streptococcus pyogenes. J. Proteom. 2020, 215, 103666. [Google Scholar] [CrossRef]
- Nie, Y.; Zhang, H.L. Microcalorimetric study on the metabolism of alkaloids from Radix Aconiti and Dendrobium against Staphylococcus aureus. Acta Chin. Med. Pharm. 2004, 18–20. [Google Scholar] [CrossRef]
- Nie, Y.; Liu, X.J.; Zhang, H.L.; Zhuo, K. Microcalorimetric study on the metabolism of alkaloids from Radix Aconiti kusnezoffii against bacteria. J. Qufu Norm. Univ. 2003, 82–84. [Google Scholar] [CrossRef]
- Pommier, Y.; Leo, E.; Zhang, H.; Marchand, C. DNA Topoisomerases and Their Poisoning by Anticancer and Antibacterial Drugs. Chem. Biol. 2010, 17, 421–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Yang, Y.; Yang, G.; Yu, L. Studies on antibacterial activity and antibacterial mechanism of a novel polysaccharide from Streptomyces virginia H03. Food Control 2010, 21, 1257–1262. [Google Scholar] [CrossRef]
- Wang, D.; Yu, L.; Xiang, H.; Fan, J.; He, L.; Guo, N.; Feng, H.; Deng, X. Global transcriptional profiles ofStaphylococcus aureustreated with berberine chloride. FEMS Microbiol. Lett. 2010, 279, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Chu, M.; Ding, R.; Chu, Z.Y.; Zhang, M.B.; Liu, X.Y.; Xie, S.H.; Zhai, Y.J.; Wang, Y.D. Role of berberine in anti-bacterial as a high-affinity LPS antagonist binding to TLR4/MD-2 receptor. BMC Complement. Altern. Med. 2014, 14, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.G.; Yan, L.; Jing, Y.Y.; Xu, L.H.; Liang, Y.D.; Wei, H.X.; Hu, B.; Pan, H.; Zha, Q.B.; Ouyang, D.Y.; et al. Berberine augments ATP-induced inflammasome activation in macrophages by enhancing AMPK signaling. Oncotarget 2016, 8, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Feng, L.; Xu, F.; Sun, Y.; Ma, Y.; Zhang, X.; Liu, H.; Xu, G.; Wu, X.; Shen, Y.; et al. Berberine inhibits palmitate-induced NLRP3 inflammasome activation by triggering autophagy in macrophages: A new mechanism linking berberine to insulin resistance improvement. Biomed. Pharmacother. 2017, 89, 864–874. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shou, J.-W.; Li, X.-Y.; Zhao, Z.-X.; Fu, J.; He, C.-Y.; Feng, R.; Ma, C.; Wen, B.-Y.; Guo, F.; et al. Berberine-induced bioactive metabolites of the gut microbiota improve energy metabolism. Metabolism 2017, 70, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.; Yang, X.J.; Ma, Y.N.; Zheng, F.; Song, X.P.; Zhou, L. Structural Modification of Sanguinarine and Chelerythrine and Their in vitro Acaricidal Activity against Psoroptes cuniculi. Chem. Pharm. Bull. 2012, 60, 1508–1513. [Google Scholar] [CrossRef] [Green Version]
- Yamada, H. Natural products of commercial potential as medicines. Curr. Opin. Biotechnol. 1991, 2, 203–210. [Google Scholar] [CrossRef]
- Vuong, C.; Yeh, A.J.; Cheung, G.Y.C.; Otto, M. Investigational drugs to treat methicillin-resistant Staphylococcus aureus. Expert Opin. Investig. Drugs 2016, 25, 73–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Liu, D.; Tian, X.; Koseki, S.; Chen, S.; Ye, X.; Ding, T. Novel antibacterial modalities against methicillin resistant Staphylococcus aureus derived from plants. Crit. Rev. Food Sci. Nutr. 2018, 59, S153–S161. [Google Scholar] [CrossRef]
- Belwal, T.; Pandey, A.; Bhatt, I.D.; Rawal, R.S. Optimized microwave assisted extraction (MAE) of alkaloids and polyphenols from Berberis roots using multiple-component analysis. Sci. Rep. 2020, 10, 917. [Google Scholar] [CrossRef]
- Djarot, N.F.U.P.; Utami, N.F.; Veonicha, A.R.N.; Rahmadini, A.; Iman, A.N. Antibacterial Activity Tests of Staphylococcus Aureus and Phytochemical Screening in Family Asteraceae, Clusiaceae, Phyllanthaceae. J. Southwest Jiaotong Univ. 2020, 55. [Google Scholar] [CrossRef]
- Chen, Y.W.; Liu, H.G. Recent Advances in Pharmacokinetics of Alkaloids. Med. Rev. 2009, 15, 3489–3491. [Google Scholar]
- Shi, Y.J. Effect of medicinal excipients on intestinal absorption of alkaloids from Coptidis rhizoma. J. Nanjing For. Univ. 2013, 4, 94. [Google Scholar]
- Wang, X.F.; Leng, X.H.; Chen, H.Y.; Guo, H.Y.; Li, J. Fingerprint analysis of total alkaloid extracts from different parts of Sophora alopecuroides. Chin. J. Exp. Tradit. Med. Form. 2016, 17, 49–54. [Google Scholar]
- Zhao, C.F. Studies on the differences of Chemical constituents and contents in different parts of Sophora flavescens. Chin. Acad. Chin. Med. Sci. 2015, 2, 157. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Y.; Li, X.; Zhang, C.; Lv, L.; Gao, B.; Li, M. Research Progress on Antibacterial Activities and Mechanisms of Natural Alkaloids: A Review. Antibiotics 2021, 10, 318. https://doi.org/10.3390/antibiotics10030318
Yan Y, Li X, Zhang C, Lv L, Gao B, Li M. Research Progress on Antibacterial Activities and Mechanisms of Natural Alkaloids: A Review. Antibiotics. 2021; 10(3):318. https://doi.org/10.3390/antibiotics10030318
Chicago/Turabian StyleYan, Yumei, Xing Li, Chunhong Zhang, Lijuan Lv, Bing Gao, and Minhui Li. 2021. "Research Progress on Antibacterial Activities and Mechanisms of Natural Alkaloids: A Review" Antibiotics 10, no. 3: 318. https://doi.org/10.3390/antibiotics10030318
APA StyleYan, Y., Li, X., Zhang, C., Lv, L., Gao, B., & Li, M. (2021). Research Progress on Antibacterial Activities and Mechanisms of Natural Alkaloids: A Review. Antibiotics, 10(3), 318. https://doi.org/10.3390/antibiotics10030318