
Wastewaters, with or without Hospital Contribution, Harbour MDR, Carbapenemase-Producing, but Not Hypervirulent Klebsiella pneumoniae


 and
and
Abstract
:1. Introduction
2. Results and Discussion
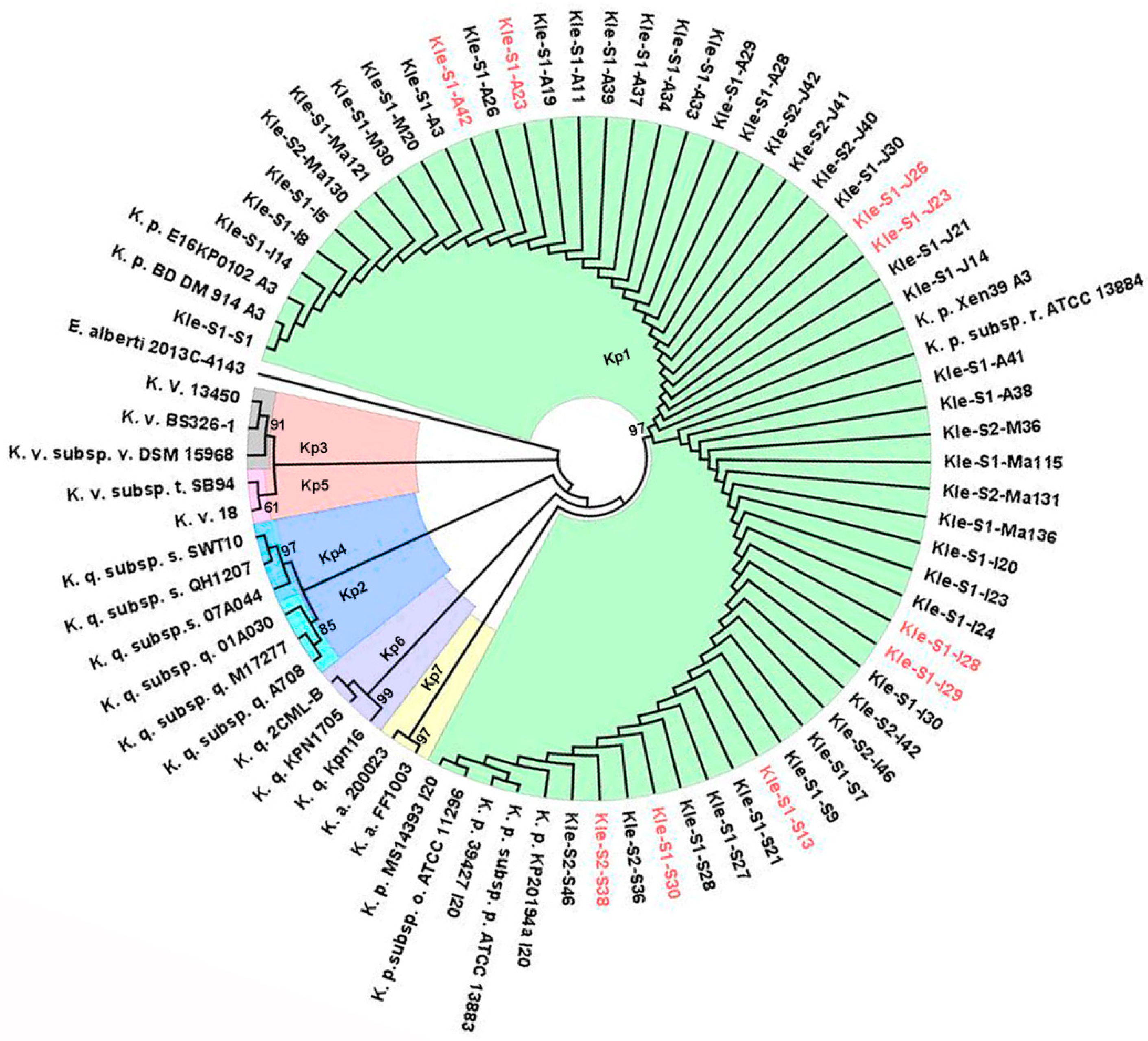
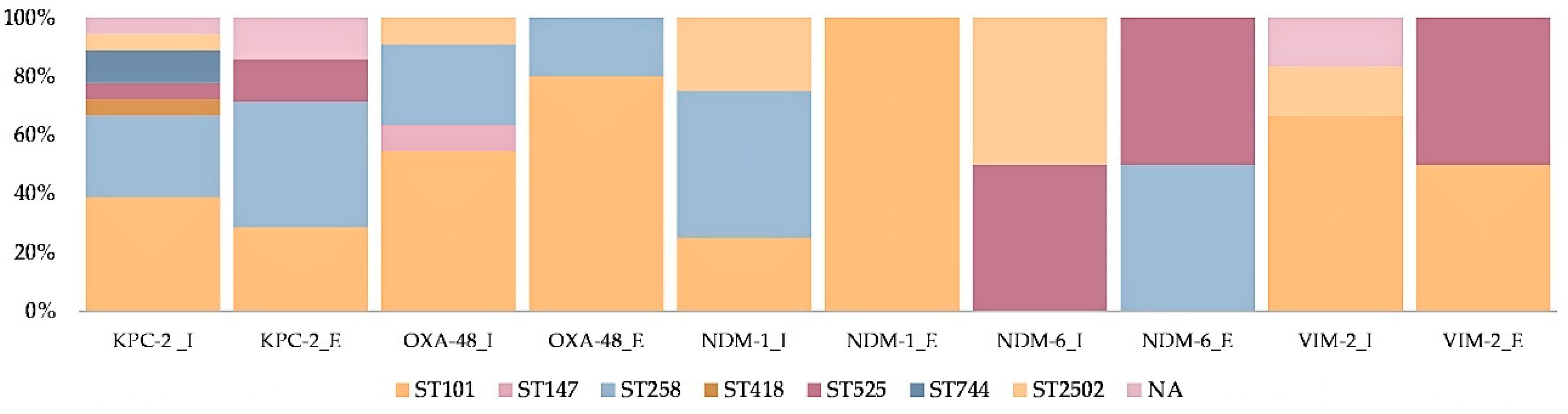
2.1. Carbapenemase Diversity and Hypervirulence of Klebsiella Pneumoniae in the Investigated Wastewaters
2.2. Klebsiella Pneumoniae Sequence Types
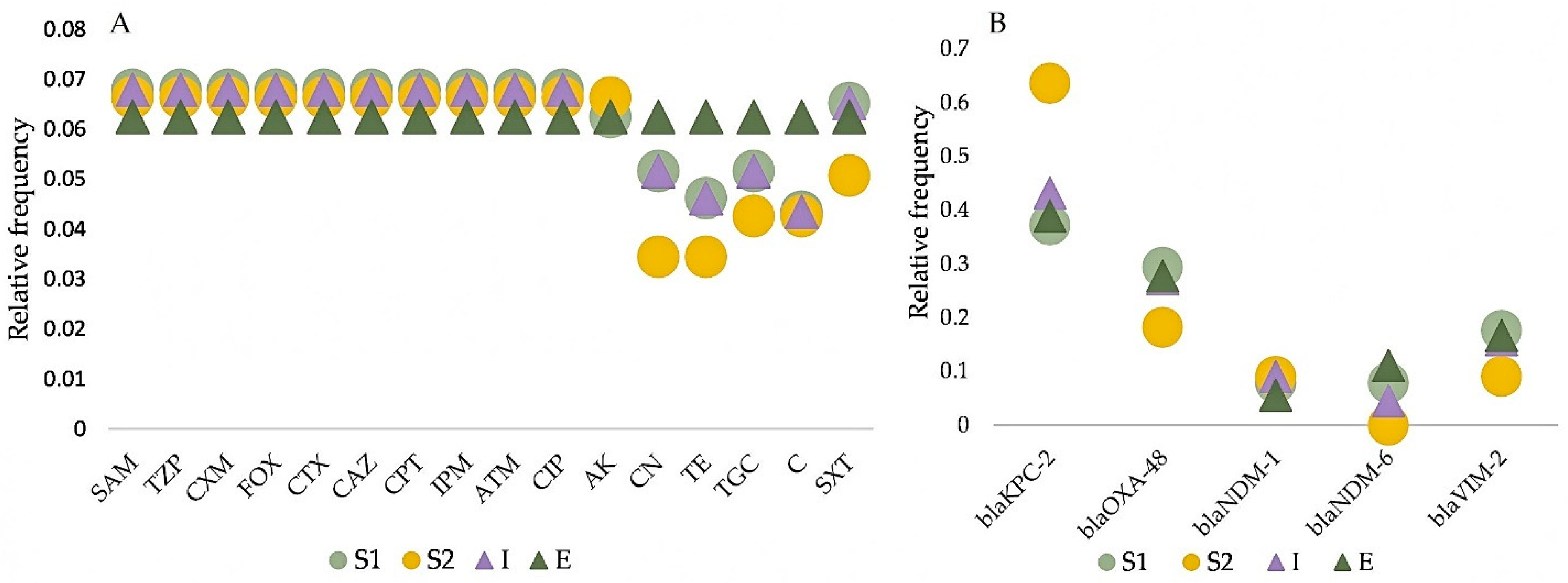
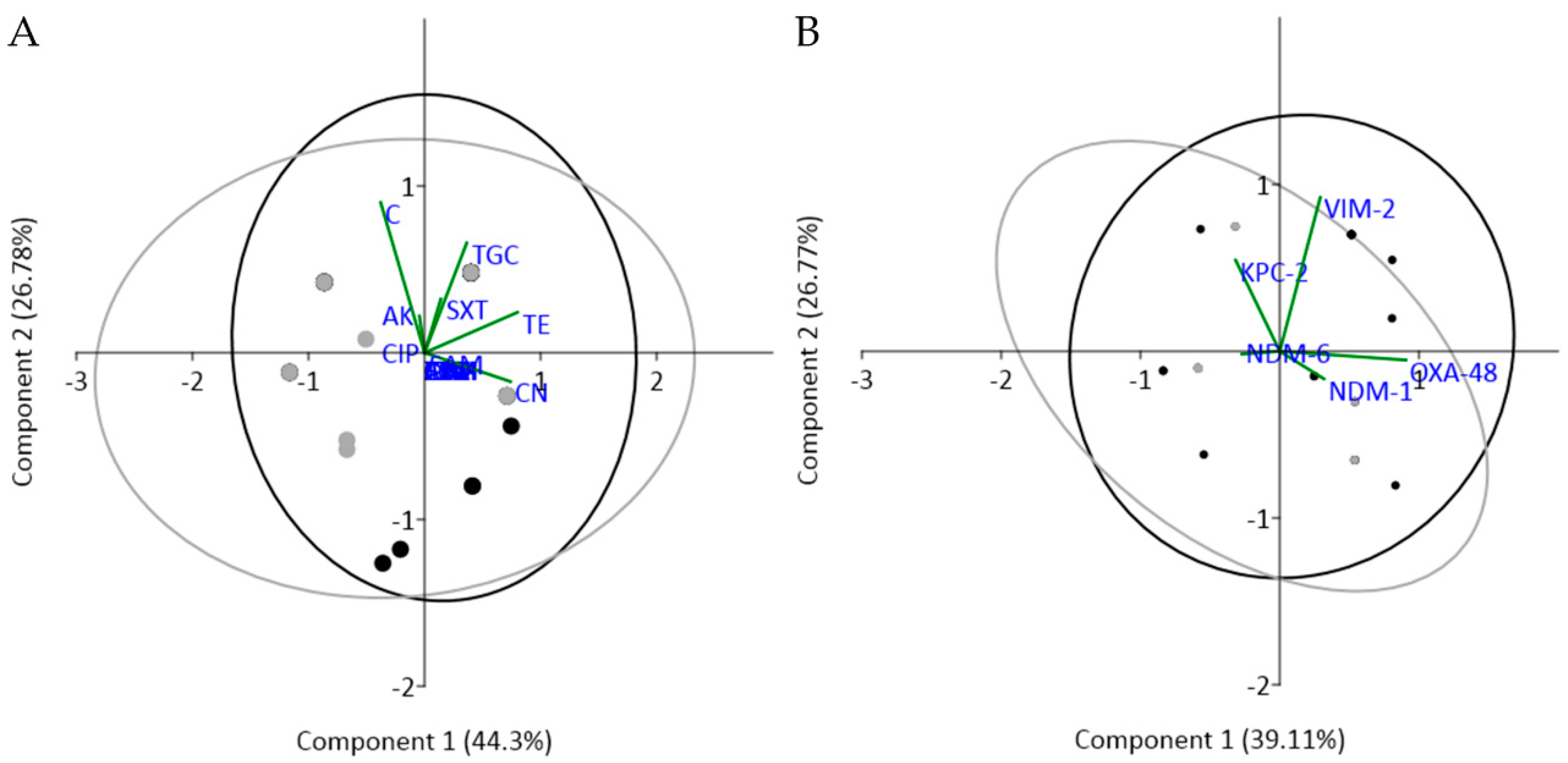
2.3. Antimicrobial Susceptibility Profiling and Comparative Analysis among Wastewater Types
2.4. Study Limitations
3. Materials and Methods
3.1. Sampling and Strain Isolation
3.2. Phenotypic Investigation of Carbapenemase Activity and Hypervirulence
3.3. Molecular Identification of Strains

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence 5′–3′ | Amplicon Size (bp) | Reference |
|---|---|---|---|
| dnaJ | DJF: CNG+GYG+ATYTGTAYGTWCAGGT DJR: T+CRT+CRA+ARAAYTTYTTNACNC | 385 | [50] |
| blaKPC | KPC Fm: CGTCTAGTTCTGCTGTCTTG KPC Rm: CTTGTCATCCTTGTTAGGCG | 232 | [56] |
| blaOXA-48 | OXA-48 F: GCGTGGTTAAGGATGAACAC OXA-48 R: CATCAAGTTCAACCCAACCG | 438 | |
| blaNDM | NDM F: GGTTTGGCGATCTGGTTTTC NDM R: CGGAATGGCTCATCACGATC | 621 | |
| blaVIM | VIM F: GATGGTGTTTGGTCGCATA VIM R: CGAATGCGCAGCACCAG | 390 | |
| iucA | F1: AATCAATGGCTATTCCCGCTG R1: CGCTTCACTTCTTTCACTGACAGG | 239 | [57] |
| iroB | F1: ATCTCATCATCTACCCTCCGCTC R1: GGTTCGCCGTCGTTTTCAA | 235 | |
| peg-344 | F: CTTGAAACTATCCCTCCAGTC R: CCAGCGAAAGAATAACCCC | 508 | |
| prmpA | F: GAGTAGTTAATAAATCAATAGCAAT R: CAGTAGGCATTGCAGCA | 332 | |
| prmpA2 | F: GTGCAATAAGGATGTTACATTA R: GGATGCCCTCCTCCTG | 430 |
3.4. Molecular Investigation of Hypervirulence Genes and the Diversity of Carbapenemases
3.5. Antimicrobial Susceptibility Testing
3.6. Identification of Sequence Types (STs)
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bardhan, T.; Chakraborty, M.; Bhattacharjee, B. Prevalence of colistin-resistant, carbapenem-hydrolyzing proteobacteria in hospital water bodies and out-falls of west Bengal, India. Int. J. Environ. Res. Public Health 2020, 17, 1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics 2017. Available online: http://www.who.int/medicines/publications. (accessed on 19 February 2021).
- Papp-Wallace, K.; Endimiani, A.; Taracila, M.; Bonomo, R. Carbapenems: Past, present, and future. Antimicrob. Agents. Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meletis, G. Carbapenem resistance: Overview of the problem and future perspectives. Ther. Adv. Infect. Dis. 2015, 3, 1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codjoe, F.S.; Donkor, E.S. Carbapenem resistance: A review. J. Med. Sci. 2017, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Effah, C.Y.; Sun, T.; Liu, S.; Wu, Y. Klebsiella pneumoniae: An increasing threat to public health. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 1. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.; Passet, V.; Rakotondrasoa, A.; Diallo, T.A.; Criscuolo, A.; Brisse, S. Description of Klebsiella africanensis sp. nov., Klebsiella variicola subsp. tropicalensis subsp. nov. and Klebsiella variicola subsp. variicola subsp. nov. Res. Microbiol. 2019, 170. [Google Scholar] [CrossRef] [Green Version]
- Brisse, S.; Passet, V.; Grimont, P.A.D. Description of Klebsiella quasipneumoniae sp. nov., a novel species isolated from human infections, with two subspecies Klebsiella quasipneumoniae subsp. quasipneumoniae subsp. nov. and Klebsiella quasipneumoniae subsp. similipneumoniae subsp. nov., and demonstration that Klebsiella singaporensis is a junior heterotypic synonym of Klebsiella variicola. Int. J. Syst. Evol. Microbiol. 2014, 64, 3146–3152. [Google Scholar] [CrossRef] [Green Version]
- Ballash, G.A.; Lee, S.; Mollenkopf, D.F.; Mathys, D.A.; Albers, A.L.; Sechrist, E.; Feicht, S.M.; Van Balen Rubio, J.C.; Sullivan, S.M.P.; Lee, J.; et al. Pulsed electric field application reduces carbapenem- and colistin-resistant microbiota and blaKPC spread in urban wastewater. J. Environ. Manag. 2020, 265, e110529. [Google Scholar] [CrossRef]
- Ebomah, K.E.; Okoh, A.I. Detection of carbapenem-resistance genes in Klebsiella species recovered from selected environmental niches in the Eastern Cape Province, South Africa. Antibiotics 2020, 9, 425. [Google Scholar] [CrossRef]
- Van Duin, D.; Doi, Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence 2017, 8, 460–469. [Google Scholar] [CrossRef]
- Alexander, J.; Hembach, N.; Schwartz, T. Evaluation of antibiotic resistance dissemination by wastewater treatment plant effluents with different catchment areas in Germany. Sci. Rep. 2020, 10, 8952. [Google Scholar] [CrossRef] [PubMed]
- Pazda, M.; Kumirska, J.; Stepnowski, P.; Mulkiewicz, E. Antibiotic resistance genes identified in wastewater treatment plant systems—A review. Sci. Total Environ. 2019, 697. [Google Scholar] [CrossRef] [PubMed]
- Marti, E.; Variatza, E.; Balcazar, J.L. The role of aquatic ecosystems as reservoirs of antibiotic resistance. Trends Microbiol. 2014, 22, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Cornejova, T.; Venglovsky, J.; Gregova, G.; Kmetova, M.; Kmet, V. Extended spectrum beta-lactamases in Escherichia coli from municipal wastewater. Ann. Agric. Environ. Med. 2015, 22, 447–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, N.; O’Connor, L.; Mahon, B.; Varley, Á.; McGrath, E.; Ryan, P.; Cormican, M.; Brehony, C.; Jolley, K.A.; Maiden, M.C.; et al. Hospital effluent: A reservoir for carbapenemase-producing Enterobacterales? Sic. Tot. Environ. 2019, 672, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Furlan, J.P.R.; Savazzi, E.A.; Stehling, E.G. Genomic insights into multidrug-resistant and hypervirulent Klebsiella pneumoniae co-harboring metal resistance genes in aquatic environments. Ecotoxicol. Environ. Saf. 2020, 201. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.A.; Hellmark, B.; Ehricht, R.; Söderquist, B.; Jass, J. Related carbapenemase-producing Klebsiella isolates detected in both a hospital and associated aquatic environment in Sweden. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 2241–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, H.; Sib, E.; Gajdiss, M.; Klanke, U.; Lenz-Plet, F.; Barabasch, V.; Albert, C.; Schallenberg, A.; Timm, C.; Zacharias, N.; et al. Dissemination of multi-resistant Gram-negative bacteria into German wastewater and surface waters. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [PubMed]
- Popa, L.I.; Gheorghe, I.; Barbu, I.C.; Surleac, M.; Paraschiv, S.; Marutescu, L.; Popa, M.; Pircalabioru, G.G.; Talapan, D.; Nita, M.; et al. Multidrug resistant Klebsiella pneumoniae ST101 clone survival chain from inpatients to hospital effluent after chlorine treatment. Front. Microbiol. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Braun, S.D.; Dorneanu, O.S.; Vremeră, T.; Reißig, A.; Monecke, S.; Ehricht, R. Carbapenemase-producing Enterobacteriaceae: A 2-year surveillance in a hospital in Iaşi, Romania. Future Microbiol. 2016. [Google Scholar] [CrossRef]
- Surleac, M.; Czobor Barbu, I.; Paraschiv, S.; Popa, L.I.; Gheorghe, I.; Marutescu, L.; Popa, M.; Sarbu., I.; Talapan, D.; Nita, M.; et al. Whole genome sequencing snapshot of multi-drug resistant Klebsiella pneumoniae strains from hospitals and receiving wastewater treatment plants in Southern Romania. PLoS ONE 2020, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurfluh, K.; Bagutti, C.; Brodmann, P.; Alt, M.; Schulze, J.; Fanning, S.; Stephan, R.; Nüesch-Inderbinen, M. Wastewater is a reservoir for clinically relevant carbapenemase- and 16s rRNA methylase-producing Enterobacteriaceae. Int. J. Antimicrob. Agents. 2017, 50, 436–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Mohamed, A.M.; Faiz, A.; Ahmad, J. Enterobacterial infection in Saudi Arabia: First record of Klebsiella pneumoniae with triple carbapenemase genes resistance. J. Infect. Dev. Ctries 2019, 13, 334–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedenić, B.; Sardelić, S. Carbapenemases-Provisional chapter. Web Sci. 2018. [Google Scholar] [CrossRef] [Green Version]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents. Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.U.; Maryam, L.; Zarrilli, R. Structure, genetics and worldwide spread of New Delhi Metallo-β-lactamase (NDM): A threat to public health. BMC Microbiol. 2017, 17, 101. [Google Scholar] [CrossRef] [Green Version]
- Gamal, D.; Fernández-Martínez, M.; El-Defrawy, I.; Ocampo-Sosa, A.A.; Martínez-Martínez, L. First identification of NDM-5 associated with OXA-181 in Escherichia coli from Egypt. Emerg. Microbes. Infect. 2016, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Bahramian, A.; Shariati, A.; Azimi, T.; Sharahi, J.Y.; Bostanghadiri, N.; Gachkar, L.; Ghalavand, Z.; Chirani, A.S.; Erfanimanesh, S.; Hashemi, A. First report of New Delhi metallo-β-lactamase-6 (NDM-6) among Klebsiella pneumoniae ST147 strains isolated from dialysis patients in Iran. Infect. Genet. Evol. 2019, 142–145. [Google Scholar] [CrossRef]
- Xanthopoulou, K.; Urrutikoetxea-Gutiérrez, M.; Vidal-Garcia, M.; Diaz de Tuesta Del Arco, J.L.; Sánchez-Urtaza, S.; Wille, J.; Seifert, H.; Higgins, P.G.; Gallego, L. First report of New Delhi Metallo-β-Lactamase-6 (NDM-6) in a clinical Acinetobacter baumannii Isolate from Northern Spain. Front. Microbiol. 2020, 11, 589253. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Peirano, G.; Devinney, R.; Bradford, P.A.; Motyl, M.R.; Adams, M.D.; Chen, L.; Kreiswirth, B.; Pitout, J.D.D. Genomic epidemiology of global VIM-producing Enterobacteriaceae. J. Antimicrob. Chemother. 2017, 72, 2249–2258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitout, J.D.D.; Peirano, G.; Kock, M.M.; Strydom, K.A.; Matsumura, Y. The global ascendency of OXA-48-Type carbapenemases. Clin. Microbiol. Rev. 2019, 33, e00102–e00119. [Google Scholar] [CrossRef] [PubMed]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2014, 41, 252–275. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Holt, M.K.E. Klebsiella pneumoniae as a key trafficker of drug resistance genes from environmental to clinically important bacteria. Curr. Opin. Microbiol. 2018, 45, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Roe, C.C.; Vazquez, A.J.; Esposito, E.P.; Zarrilli, R.; Sahl, J.W. Diversity, virulence and antimicrobial resistance in isolates from the newly emerging Klebsiella pneumoniae ST101 lineage. Front. Microbiol. 2019, 2, 542. [Google Scholar] [CrossRef] [PubMed]
- Can, F.; Menekse, S.; Ispir, P.; Atac, N.; Albayrak, O.; Demir, T.; Karaaslan, D.C.; Karahan, S.N.; Kapmaz, M.; Kurt Azap, O.; et al. Ergonul, O. Impact of the ST101 clone on fatality among patients with colistin-resistant Klebsiella pneumoniae infection. J. Antimicrob. Chemother. 2018, 73, 1235–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messaoudi, A.; Haenni, M.; Bouallègue, O.; Saras, E.; Chatre, P.; Chaouch, C.; Boujâafar, N.; Mansour, W.; Madec, J.Y. Dynamics and molecular features of OXA-48-like-producing Klebsiella pneumoniae lineages in a Tunisian hospital. J. Glob. Antimicrob. Resist. 2020, 20, 87–93. [Google Scholar] [CrossRef]
- Arena, F.; Di Pilato, V.; Vannetti, F.; Fabbri, L.; Antonelli, A.; Coppi, M.; Pupillo, R.; Macchi, C.; Rossolini, G.M. Population structure of KPC carbapenemase-producing Klebsiella pneumoniae in a long-term acute-care rehabilitation facility: Identification of a new lineage of clonal group 101, associated with local hyperendemicity. Microb. Genom. 2020, 6. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Ragheb, S.M.; Tawfick, M.M.; El-Kholy, A.A.; Abdulall, A.K. Phenotypic and genotypic features of Klebsiella pneumoniae harboring carbapenemases in Egypt: OXA-48-Like carbapenemases as an Investigated Model. Antibiotics 2020, 9, 852. [Google Scholar] [CrossRef]
- Lv, L.; Wan, M.; Wang, C.; Gao, X.; Yang, Q.; Partridge, S.R.; Wang, Y.; Zong, Z.; Doi, Y.; Shen, J.; et al. Emergence of a plasmid-encoded resistance-nodulation-division efflux pump conferring resistance to multiple drugs, including tigecycline, in Klebsiella pneumoniae. Mbio 2020, 11, e02930-19. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Choi, Q.; Kwon, G.C.; Koo, S.H. Molecular epidemiology and mechanisms of tigecycline resistance in carbapenem-resistant Klebsiella pneumoniae isolates. J. Clin. Lab. Anal. 2020, 34, 23506. [Google Scholar] [CrossRef]
- Elazhary, M.A.S.Y.; Saheb, S.A.; Roy, R.S.; Lagace, A. A simple procedure for the preliminary identification of aerobic Gram-negative intestinal bacteria with special reference to the Enterobacteriaceae. Can. J. Comp. Med. 1973, 37, 43–46. [Google Scholar]
- Jing, X.; Zhou, H.; Min, X.; Zhang, X.; Yang, Q.; Du, S.; Li, Y.; Yu, F.; Jia, M.; Zhan, Y.; et al. The simplified carbapenem inactivation method (sCIM) for simple and accurate detection of carbapenemase-producing Gram-negative bacilli. Front. Microbiol. 2018, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Workneh, M.; Yee, R.; Simner, P.J. Phenotypic methods for detection of carbapenemase production in carbapenem-resistant organisms: What method should your laboratory choose? Clin. Microbiol. News 2019, 41, 11–22. [Google Scholar] [CrossRef]
- Shon, A.S.; Bajwa, R.P.; Russo, T.A. Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: A new and dangerous breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadano, Y. String test. BMJ Case. Rep. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.T.; Chuang, Y.P.; Shun, C.T.; Chang, S.C.; Wang, J.T. A novel virulence gene in Klebsiella pneumoniae strains causing primary liver abscess and septic metastatic complications. J. Exp. Med. 2004, 199, 697–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nhung, P.H.; Shah, M.M.; Ohkusu, K.; Noda, M.; Hata, H.; Sun, X.S.; Iihara, H.; Goto, K.; Masaki, T.; Miyasaka, J.; et al. The dnaJ gene as a novel phylogenetic marker for identification of Vibrio species. Syst. Appl. Microbiol. 2007, 30, 309–315. [Google Scholar] [CrossRef] [PubMed]
- McLean, K.; Rosenthal, C.A.; Sengupta, D.; Owens, J.; Cookson, B.T.; Hoffman, N.G.; Salipante, S.J. Improved species-level clinical identification of Enterobacteriaceae through broad-range dnaJ PCR and sequencing. J. Clin. Microbiol. 2019, 23, 1–22. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis acrosscomputing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordmann, P.; Naas, T.; Poirel, L. Global spread of carbapenemase producing Enterobacteriaceae. Emerg. Infect. Dis. 2011, 17, 10. [Google Scholar] [CrossRef]
- Russo, T.A.; Olson, R.; Fang, C.T.; Stoesser, N.; Miller, M.; MacDonald, U.; Hutson, A.; Barker, J.H.; La Hoz, R.M.; Johnson, J.R. Identification of biomarkers for differentiation of hypervirulent Klebsiella pneumoniae from classical K. pneumoniae. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [Green Version]
- Hudzicki, J. Kirby-Bauer disk diffusion susceptibility test protocol. ASM 2009, 1–23. Available online: https://asm.org/Protocols/Kirby-Bauer-Disk-Diffusion-Susceptibility-Test-Pro (accessed on 1 March 2020).
- Osei Sekyere, J.; Govinden, U.; Bester, L.A.; Essack, S.Y. Colistin and tigecycline resistance in carbapenemase-producing Gram-negative bacteria: Emerging resistance mechanisms and detection methods. J. Appl. Microbiol. 2016, 121, 601–617. [Google Scholar] [CrossRef] [PubMed]
- Sader, H.S.; Ferraro, M.J.; Reller, L.B.; Schreckenberger, P.C.; Swenson, J.M.; Jones, R.N. Reevaluation of clinical and laboratory standards institute disk diffusion breakpoints for tetracyclines for testing Enterobacteriaceae. J. Clin. Microbiol. 2007, 45, 1640–1643. [Google Scholar] [CrossRef] [Green Version]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.A.; Brisse, S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J. Clin. Microbiol. 2005, 43, 4178–4182. [Google Scholar] [CrossRef] [Green Version]
- Institut Pasteur. Genomic-Based Strain Taxonomy and nomenclature Platform of Institut Pasteur. Available online: https://bigsdb.pasteur.fr/klebsiella/primers_used.html (accessed on 19 February 2021).
- Pub MLST. Public Databases for Molecular Typing and Microbial Genome Diversity. Available online: https://pubmlst.org/ (accessed on 19 February 2021).
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, D.P. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Real Statistics. Real Statistics Using Excel. Available online: https://www.real-statistics.com/ (accessed on 19 February 2021).




| Combination of CRG Types in the K. pneumoniae Isolates | Prevalence of CRG Patterns (n = 33) | ||
|---|---|---|---|
| Total | S1_I/S1_E | S2_I/S2_E | |
| blaKPC | 10 | 4/1 | 4/1 |
| blaOXA-48 | 4 | 2/1 | 1/0 |
| blaNDM | 1 | 1/0 | 0/0 |
| blaVIM | 0 | 0/0 | 0/0 |
| blaKPC/blaNDM | 2 | 1/1 | 0/0 |
| blaKPC/blaOXA-48 | 2 | 1/1 | 0/0 |
| blaKPC/blaVIM | 2 | 1/0 | 1/0 |
| blaOXA-48/blaNDM | 1 | 1/0 | 0/0 |
| blaOXA-48/blaVIM | 1 | 0/1 | 0/0 |
| blaKPC/blaOXA-48/blaNDM | 3 | 1/1 | 1/0 |
| blaKPC/blaOXA-48/blaVIM | 5 | 4/1 | 0/0 |
| blaKPC/blaNDM/blaVIM | 1 | 1/0 | 0/0 |
| blaKPC/blaOXA-48/blaNDM/blaVIM | 1 | 1/0 | 0/0 |
| Group | N | AMR | CRG | ||||
|---|---|---|---|---|---|---|---|
| p | SI | GSI | p | SI | GSI | ||
| S1 | 25 | 14.72 | 0.177 | 0.936 (99.88%) | 2.04 | 0.551 | 0.731 (91.41%) |
| S2 | 8 | 14.00 | 0.204 | 0.934 (99.68%) | 1.375 | 0.339 | 0.545 (68.18%) |
| I | 24 | 13.44 | 0.212 | 0.936 (99.88%) | 1.833 | 0.509 | 0.704 (87.94%) |
| E | 9 | 16 | 0 | 0.938 (100%) | 2.000 | 0.546 | 0.728 (91.04%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teban-Man, A.; Farkas, A.; Baricz, A.; Hegedus, A.; Szekeres, E.; Pârvu, M.; Coman, C. Wastewaters, with or without Hospital Contribution, Harbour MDR, Carbapenemase-Producing, but Not Hypervirulent Klebsiella pneumoniae. Antibiotics 2021, 10, 361. https://doi.org/10.3390/antibiotics10040361
Teban-Man A, Farkas A, Baricz A, Hegedus A, Szekeres E, Pârvu M, Coman C. Wastewaters, with or without Hospital Contribution, Harbour MDR, Carbapenemase-Producing, but Not Hypervirulent Klebsiella pneumoniae. Antibiotics. 2021; 10(4):361. https://doi.org/10.3390/antibiotics10040361
Chicago/Turabian StyleTeban-Man, Adela, Anca Farkas, Andreea Baricz, Adriana Hegedus, Edina Szekeres, Marcel Pârvu, and Cristian Coman. 2021. "Wastewaters, with or without Hospital Contribution, Harbour MDR, Carbapenemase-Producing, but Not Hypervirulent Klebsiella pneumoniae" Antibiotics 10, no. 4: 361. https://doi.org/10.3390/antibiotics10040361
APA StyleTeban-Man, A., Farkas, A., Baricz, A., Hegedus, A., Szekeres, E., Pârvu, M., & Coman, C. (2021). Wastewaters, with or without Hospital Contribution, Harbour MDR, Carbapenemase-Producing, but Not Hypervirulent Klebsiella pneumoniae. Antibiotics, 10(4), 361. https://doi.org/10.3390/antibiotics10040361