
Novel Soil-Derived Beta-Lactam, Chloramphenicol, Fosfomycin and Trimethoprim Resistance Genes Revealed by Functional Metagenomics

Abstract
:1. Introduction
2. Results and Discussion
2.1. Screening of Soil Metagenomic Libraries Enabled the Discovery of Eight Novel ARGs
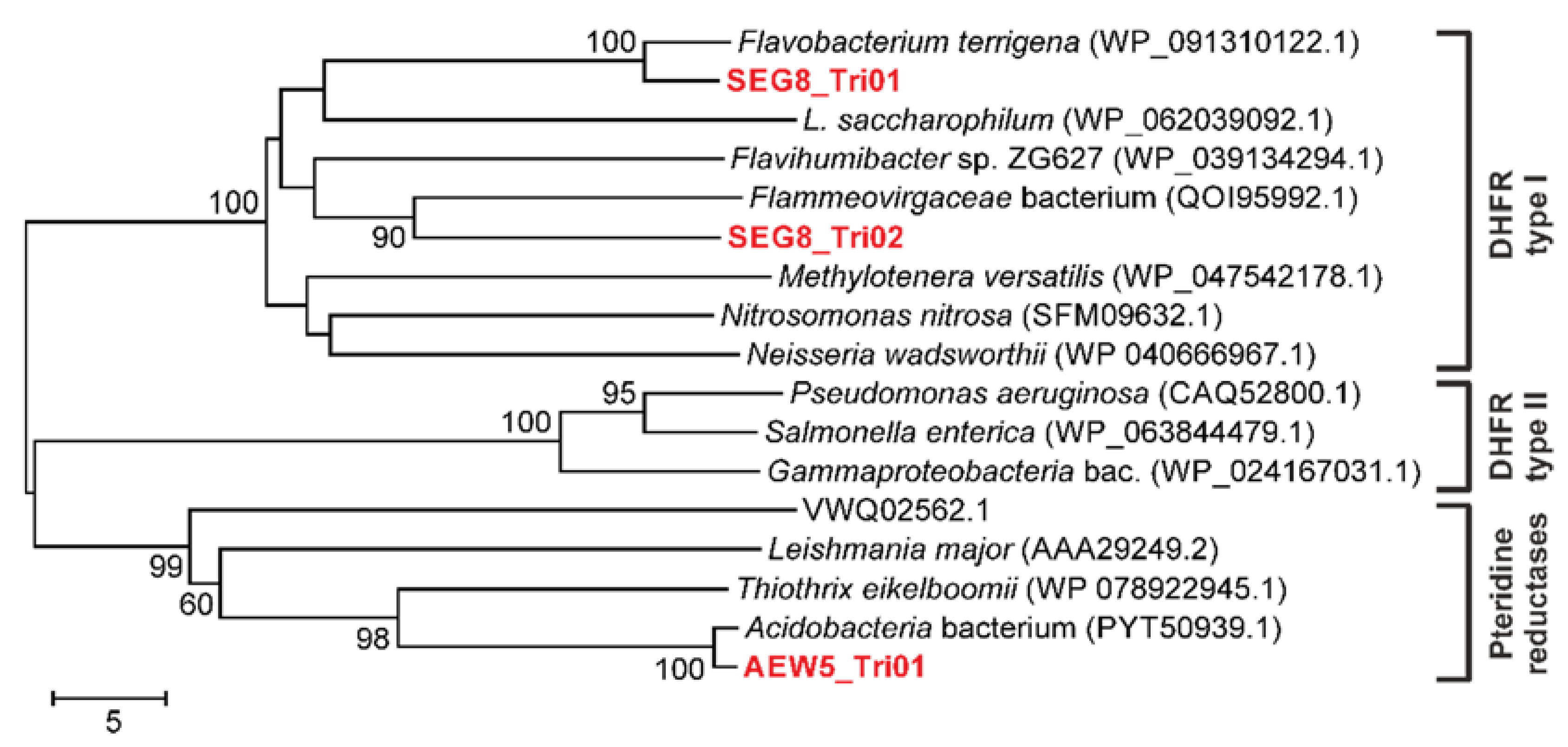
2.2. Trimethoprim Resistance Mediated by Soil-Derived DHFRs and a Pteridine Reductase
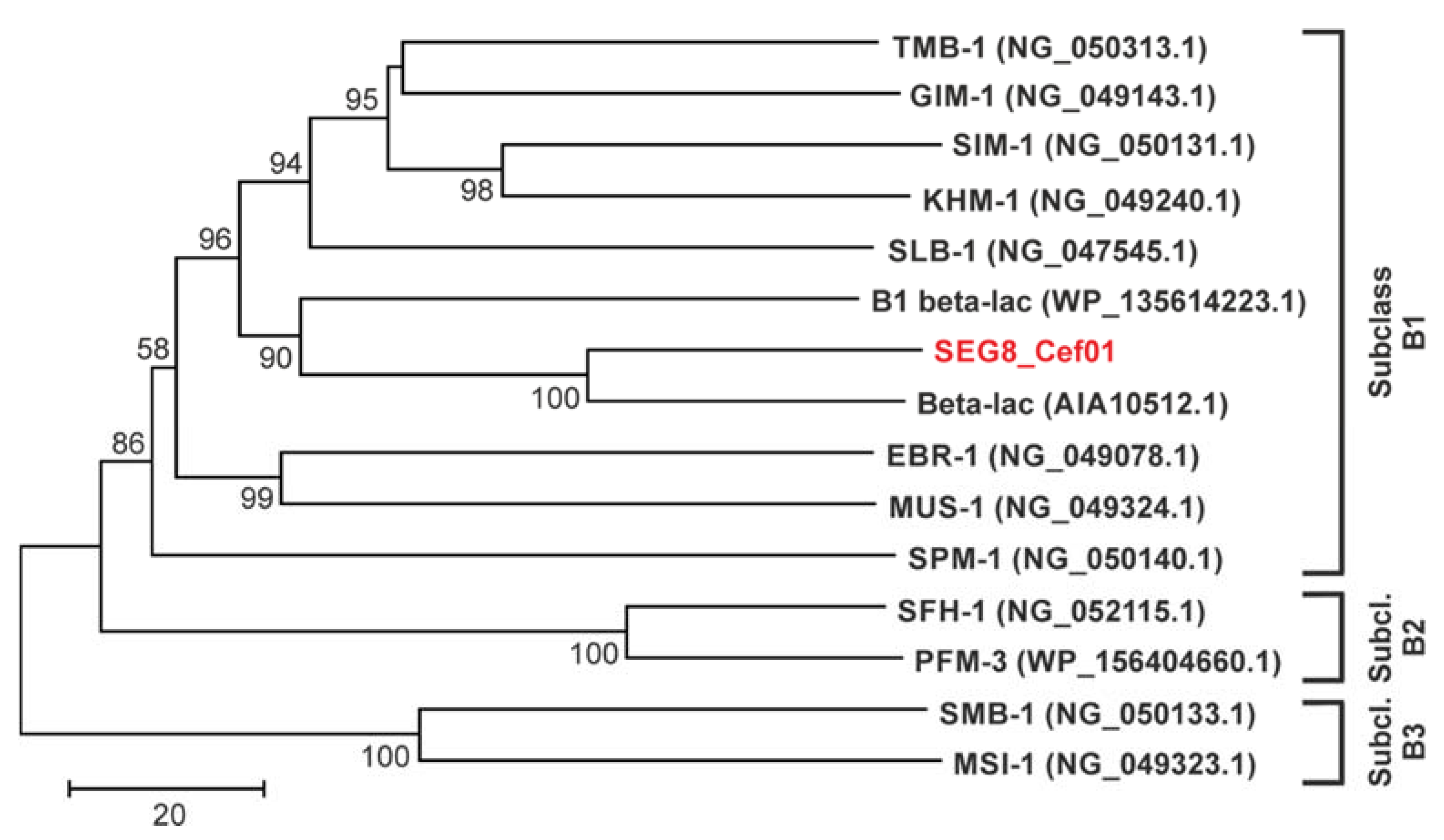
2.3. Identification of a Novel Subclass B1 Metallo-Beta-Lactamase
2.4. Efflux-Mediated Chloramphenicol Resistance Encoded by SEW5_chl01
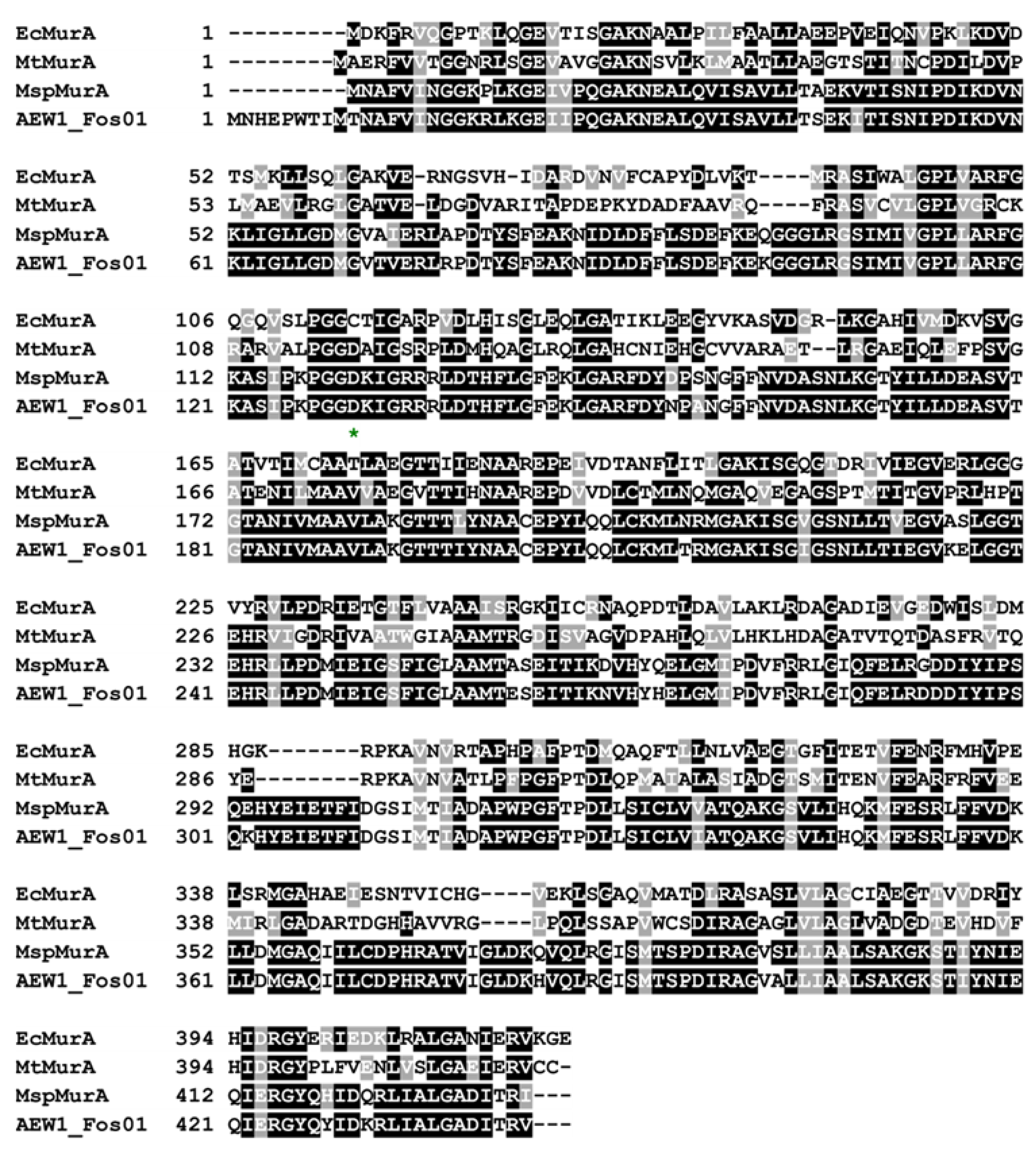
2.5. AEW1_Fos01 Confers Resistance to the Last Resort Antibiotic Fosfomycin
2.6. AEW4_Amp01 and SEG8_Amp01 Evoke Resistance toward Different Penicillins
3. Materials and Methods
3.1. Soil Metagenomic Libraries
3.2. Function-Based Metagenomic Library Screening
3.3. Sequence Analysis
3.4. Subcloning of Putative ARGs
3.5. Determination of Minimum Inhibitory Concentrations (MICs)
3.6. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dos Santos, D.F.; Istvan, P.; Quirino, B.F.; Kruger, R.H. Functional metagenomics as a tool for identification of new antibiotic resistance genes from natural environments. Microb. Ecol. 2017, 73, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Castillo Villamizar, G.A.; Nacke, H.; Boehning, M.; Herz, K.; Daniel, R. Functional metagenomics reveals an overlooked diversity and novel features of soil-derived bacterial phosphatases and phytases. mBio 2019, 10, e01966-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De, R. Metagenomics: Aid to combat antimicrobial resistance in diarrhea. Gut Pathog. 2019, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.H.; van Engelen, K.; Gordon, S.; Renaud, J.; Topp, E. Novel antibiotic resistance determinants from agricultural soil exposed to antibiotics widely used in human medicine and animal farming. Appl. Environ. Microbiol. 2017, 83, e00989-17. [Google Scholar] [CrossRef] [Green Version]
- Allen, H.K.; An, R.; Handelsman, J.; Moe, L.A. A response regulator from a soil metagenome enhances resistance to the β-lactam antibiotic carbenicillin in Escherichia coli. PLoS ONE 2015, 10, e0120094. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. β-lactams and β-lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- Lima, L.M.; da Silva, B.N.M.; Barbosa, G.; Barreiro, E.J. β-lactam antibiotics: An overview from a medicinal chemistry perspective. Eur. J. Med. Chem. 2020, 208, 112829. [Google Scholar] [CrossRef]
- Eiamphungporn, W.; Schaduangrat, N.; Malik, A.A.; Nantasenamat, C. Tackling the antibiotic resistance caused by class A β-lactamases through the use of β-lactamase inhibitory protein. Int. J. Mol. Sci. 2018, 19, 2222. [Google Scholar] [CrossRef] [Green Version]
- Wilke, M.S.; Lovering, A.L.; Strynadka, N.C. Beta-lactam antibiotic resistance: A current structural perspective. Curr. Opin. Microbiol. 2005, 8, 525–533. [Google Scholar] [CrossRef]
- Armalytė, J.; Skerniškytė, J.; Bakienė, E.; Krasauskas, R.; Šiugždinienė, R.; Kareivienė, V.; Kerzienė, S.; Klimienė, I.; Sužiedėlienė, E.; Ružauskas, M. Microbial diversity and antimicrobial resistance profile in microbiota from soils of conventional and organic farming systems. Front. Microbiol. 2019, 10, 892. [Google Scholar] [CrossRef] [PubMed]
- Silver, L.L. Fosfomycin: Mechanism and resistance. Cold Spring Har. Perspect. Med. 2017, 7, a025262. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-García, A.; Blázquez, J.; Rodríguez-Rojas, A. Molecular mechanisms and clinical impact of acquired and intrinsic fosfomycin resistance. Antibiotics 2013, 2, 217–236. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.C.M. Potential of fosfomycin in treating multidrug-resistant infections in children. J. Paediatr. Child Health 2020, 56, 864–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, R.; Mustapha, M.M.; Tomich, A.D.; Callaghan, J.D.; McElheny, C.L.; Mettus, R.T.; Shanks, R.M.Q.; Sluis-Cremer, N.; Doi, Y. Widespread fosfomycin resistance in Gram-negative bacteria attributable to the chromosomal fosA gene. mBio 2017, 8, e00749-17. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Pankow, C.A.; Oh, M.; Heath, L.S.; Zhang, L.; Du, P.; Xia, K.; Pruden, A. Effect of antibiotic use and composting on antibiotic resistance gene abundance and resistome risks of soils receiving manure-derived amendments. Environ. Int. 2019, 128, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Hu, Y.; Lu, N.; Li, J.; Wang, Z.; Chen, Q.; Zhu, B. Identification of a novel fosfomycin-resistant UDP-N-acetylglucosamine enolpyruvyl transferase (MurA) from a soil metagenome. Biotechnol. Lett. 2013, 35, 273–278. [Google Scholar] [CrossRef]
- Schwarz, S.; Kehrenberg, C.; Doublet, B.; Cloeckaert, A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 2004, 28, 519–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crofts, T.S.; Sontha, P.; King, A.O.; Wang, B.; Biddy, B.A.; Zanolli, N.; Gaumnitz, J.; Dantas, G. Discovery and characterization of a nitroreductase capable of conferring bacterial resistance to chloramphenicol. Cell Chem. Biol. 2019, 26, 559–570. [Google Scholar] [CrossRef]
- Van Goethem, M.W.; Pierneef, R.; Bezuidt, O.K.I.; Van De Peer, Y.; Cowan, D.A.; Makhalanyane, T.P. A reservoir of ‘historical’ antibiotic resistance genes in remote pristine Antarctic soils. Microbiome 2018, 6, 40. [Google Scholar] [CrossRef] [Green Version]
- Dinos, G.P.; Athanassopoulos, C.M.; Missiri, D.A.; Giannopoulou, P.C.; Vlachogiannis, I.A.; Papadopoulos, G.E.; Papaioannou, D.; Kalpaxis, D.L. Chloramphenicol derivatives as antibacterial and anticancer agents: Historic problems and current solutions. Antibiotics 2016, 5, 20. [Google Scholar] [CrossRef]
- Vecchione, J.J.; Alexander, B.; Sello, J.K. Two distinct major facilitator superfamily drug efflux pumps mediate chloramphenicol resistance in Streptomyces coelicolor. Antimicrob. Agents Chemother. 2009, 53, 4673–4677. [Google Scholar] [CrossRef] [Green Version]
- Sköld, O. Resistance to trimethoprim and sulfonamides. Vet. Res. 2001, 32, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, R.; van der Linden, M.; Chhatwal, G.S.; Nitsche-Schmitz, D.P. Factors that cause trimethoprim resistance in Streptococcus pyogenes. Antimicrob. Agents Chemother. 2014, 58, 2281–2288. [Google Scholar] [CrossRef] [Green Version]
- McGarvey, K.M.; Queitsch, K.; Fields, S. Wide variation in antibiotic resistance proteins identified by functional metagenomic screening of a soil DNA library. Appl. Environ. Microbiol. 2012, 78, 1708–1714. [Google Scholar] [CrossRef] [Green Version]
- Torres-Cortés, G.; Millán, V.; Ramírez-Saad, H.C.; Nisa-Martínez, R.; Toro, N.; Martínez-Abarca, F. Characterization of novel antibiotic resistance genes identified by functional metagenomics on soil samples. Environ. Microbiol. 2011, 13, 1101–1114. [Google Scholar] [CrossRef]
- Nacke, H.; Will, C.; Herzog, S.; Nowka, B.; Engelhaupt, M.; Daniel, R. Identification of novel lipolytic genes and gene families by screening of metagenomic libraries derived from soil samples of the German Biodiversity Exploratories. FEMS Microbiol. Ecol. 2011, 78, 188–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willms, I.M.; Kamran, A.; Aßmann, N.F.; Krone, D.; Bolz, S.H.; Fiedler, F.; Nacke, H. Discovery of novel antibiotic resistance determinants in forest and grassland soil metagenomes. Front. Microbiol. 2019, 10, 460. [Google Scholar] [CrossRef] [PubMed]
- Mullany, P. Functional metagenomics for the investigation of antibiotic resistance. Virulence 2014, 5, 443–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, D.A.; Bolin, J.T.; Burridge, J.M.; Filman, D.J.; Volz, K.W.; Kraut, J. Dihydrofolate reductase. The stereochemistry of inhibitor selectivity. J. Biol. Chem. 1985, 260, 392–399. [Google Scholar] [CrossRef]
- Sundqvist, M.; Geli, P.; Andersson, D.I.; Sjölund-Karlsson, M.; Runehagen, A.; Cars, H.; Abelson-Storby, K.; Cars, O.; Kahlmeter, G. Little evidence for reversibility of trimethoprim resistance after a drastic reduction in trimethoprim use. J. Antimicrob. Chemother. 2010, 65, 350–360. [Google Scholar] [CrossRef]
- González-Plaza, J.J.; Šimatović, A.; Milaković, M.; Bielen, A.; Wichmann, F.; Udiković-Kolić, N. Functional repertoire of antibiotic resistance genes in antibiotic manufacturing effluents and receiving freshwater sediments. Front. Microbiol. 2018, 8, 2675. [Google Scholar] [CrossRef]
- Howell, E.E. Searching sequence space: Two different approaches to dihydrofolate reductase catalysis. Chembiochem 2005, 6, 590–600. [Google Scholar] [CrossRef]
- Tulloch, L.B.; Martini, V.P.; Iulek, J.; Huggan, J.K.; Lee, J.H.; Gibson, C.L.; Smith, T.K.; Suckling, C.J.; Hunter, W.N. Structure-based design of pteridine reductase inhibitors targeting African sleeping sickness and the leishmaniases. J. Med. Chem. 2010, 53, 221–229. [Google Scholar] [CrossRef]
- Forsberg, K.J.; Patel, S.; Gibson, M.K.; Lauber, C.L.; Knight, R.; Fierer, N.; Dantas, G. Bacterial phylogeny structures soil resistomes across habitats. Nature 2014, 509, 612–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojica, M.F.; Bonomo, R.A.; Fast, W. B1-Metallo-β-Lactamases: Where do we stand? Curr. Drug Targets 2016, 17, 1029–1050. [Google Scholar] [CrossRef]
- Palzkill, T. Metallo-β-lactamase structure and function. Ann. N. Y. Acad. Sci. 2013, 1277, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Gudeta, D.D.; Bortolaia, V.; Pollini, S.; Docquier, J.D.; Rossolini, G.M.; Amos, G.C.; Wellington, E.M.; Guardabassi, L. Expanding the repertoire of carbapenem-hydrolyzing metallo-ß-lactamases by functional metagenomic analysis of soil microbiota. Front. Microbiol. 2016, 7, 1985. [Google Scholar] [CrossRef]
- Pradel, N.; Delmas, J.; Wu, L.F.; Santini, C.L.; Bonnet, R. Sec- and Tat-dependent translocation of beta-lactamases across the Escherichia coli inner membrane. Antimicrob. Agents Chemother. 2009, 53, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.P.; Kumar, S.; Varela, M.F. Identification, cloning, and functional characterization of EmrD-3, a putative multidrug efflux pump of the major facilitator superfamily from Vibrio cholerae O395. Arch. Microbiol. 2009, 191, 903–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqua, M.; Grossi, M.; Zennaro, A.; Fanelli, G.; Micheli, G.; Barras, F.; Colonna, B.; Prosseda, G. The varied role of efflux pumps of the MFS family in the interplay of bacteria with animal and plant cells. Microorganisms 2019, 7, 285. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Deng, Z.; Yan, A. Bacterial multidrug efflux pumps: Mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 2014, 453, 254–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Lekshmi, M.; Parvathi, A.; Ojha, M.; Wenzel, N.; Varela, M.F. Functional and structural roles of the major facilitator superfamily bacterial multidrug efflux pumps. Microorganisms 2020, 8, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feßler, A.T.; Wang, Y.; Wu, C.; Schwarz, S. Mobile macrolide resistance genes in staphylococci. Plasmid 2018, 99, 2–10. [Google Scholar] [CrossRef]
- Slipski, C.J.; Zhanel, G.G.; Bay, D.C. Biocide selective TolC-independent efflux pumps in Enterobacteriaceae. J. Membr. Biol. 2018, 251, 15–33. [Google Scholar] [CrossRef]
- Thompson, M.K.; Keithly, M.E.; Sulikowski, G.A.; Armstrong, R.N. Diversity in fosfomycin resistance proteins. Perspec. Sci. 2015, 4, 17–23. [Google Scholar] [CrossRef] [Green Version]
- López-Montesinos, I.; Horcajada, J.P. Oral and intravenous fosfomycin in complicated urinary tract infections. Rev. Esp. Quimioter. 2019, 32 (Suppl. S1), 37–44. [Google Scholar]
- Singh, V.K.; Vaish, M.; Johansson, T.R.; Baum, K.R.; Ring, R.P.; Singh, S.; Shukla, S.K.; Moskovitz, J. Significance of four methionine sulfoxide reductases in Staphylococcus aureus. PLoS ONE 2015, 10, e0117594. [Google Scholar] [CrossRef] [PubMed]
- Chouchani, C.; El Salabi, A.; Marrakchi, R.; Ferchichi, L.; Walsh, T.R. First report of mefA and msrA/msrB multidrug efflux pumps associated with blaTEM-1 β-lactamase in Enterococcus faecalis. Int. J. Infect. Dis. 2012, 16, e104–e109. [Google Scholar] [CrossRef] [Green Version]
- Venter, H.; Mowla, R.; Ohene-Agyei, T.; Ma, S. RND-type drug efflux pumps from Gram-negative bacteria: Molecular mechanism and inhibition. Front. Microbiol. 2015, 6, 377. [Google Scholar] [CrossRef]
- Spengler, G.; Kincses, A.; Gajdács, M.; Amaral, L. New roads leading to old destinations: Efflux pumps as targets to reverse multidrug resistance in bacteria. Molecules 2017, 22, 468. [Google Scholar] [CrossRef] [Green Version]
- Nikaido, H.; Takatsuka, Y. Mechanisms of RND multidrug efflux pumps. Biochim. Biophys. Acta 2009, 1794, 769–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belfort, M.; Bonocora, R.P. Homing endonucleases: From genetic anomalies to programmable genomic clippers. Methods Mol. Biol. 2014, 1123, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, M.; Bossdorf, O.; Gockel, S.; Hänsel, F.; Hemp, A.; Hessenmöller, D.; Korte, G.; Nieschulze, J.; Pfeiffer, S.; Prati, D.; et al. Implementing large-scale and long-term functional biodiversity research: The Biodiversity Exploratories. Basic Appl. Ecol. 2010, 11, 473–485. [Google Scholar] [CrossRef]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-lactamase database (BLDB)—Structure and function. J. Enzyme Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef] [PubMed]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Leplae, R.; Lima-Mendez, G.; Toussaint, A. ACLAME: A CLAssification of Mobile genetic Elements, update 2010. Nucleic Acids Res. 2010, 38, D57–D61. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Library | Number of Clones | Average Insert Size (kb) | Insert Frequency (%) | Estimated Library Size (Gb) | Plasmids of Positive Clones |
|---|---|---|---|---|---|
| AEW4 | 54320 | 3.4 | 80 | 0.15 | pLAEW4_amp01 |
| SEG8 * | 559000 | 4.8 | 86 | 2.30 | pLSEG8_amp01, pLSEG8_cef01, pLSEG8_tri01-02 |
| SEW5 * | 166040 | 4.0 | 95 | 0.63 | pLSEW5_chl01 |
| AEW1 * | 129748 | 6.7 | 91 | 0.79 | pLAEW1_fos01 |
| AEW5 * | 90300 | 5.2 | 89 | 0.42 | pLAEW5_tri01 |
| Gene | No. of Encoded Amino Acids | Closest Similar Protein, Accession no. (no. of Encoded Amino Acids), Organism | E-Value | Identity to Closest Similar Protein (Blast), no. of aa Similar/Total no. (%) | Percent Identity to Closest Similar Protein (Clustal, Full Length) |
|---|---|---|---|---|---|
| AEW4_amp01 | 343 | Efflux RND transporter periplasmic subunit, PYM22253 (469), Candidatus Rokubacteria bacterium | 1e-14 | 102/359 (28%) | 20.833 |
| SEG8_amp01 | 220 | Bifunctional methionine sulfoxide reductase, MBI1189218 (321), Tepidisphaera sp. | 4e-112 | 158/216 (73%) | 48.160 |
| SEG8_cef01 | 251 | NDM-CcrA beta lactamase, AIA10512 (261), uncultured bacterium | 9e-119 | 162/248 (65%) | 62.069 |
| SEW5_chl01 | 402 | Bcr/CflA subfamily drug resistance transporter, AIA10695 (410), uncultured bacterium | 0.0 | 370/402 (92%) | 90.244 |
| AEW1_fos01 | 443 | UDP-N-acetylglucosamine 1-carboxyvinyltransferase (MurA), HAL80920 (434), Mucilaginibacter sp. | 0.0 | 406/433 (94%) | 91.648 |
| AEW5_tri01 | 263 | Pteridine reductase, PYT50939 (263), Acidobacteria bacterium | 2e-171 | 236/263 (90%) | 89.734 |
| SEG8_tri01 | 165 | Dihydrofolate reductase, WP_091310122 (162), Flavobacterium terrigena | 3e-108 | 147/162 (91%) | 89.091 |
| SEG8_tri02 | 166 | Dihydrofolate reductase, QOI95992 (167), Flammeovirgaceae bacterium | 4e-88 | 117/165 (71%) | 70.060 |
| Plasmid | Minimum Inhibitory Concentration (µg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| CHL | FOS | RIF | TET | TRI | CPH | CFX | CFT | |
| Cloning vector | 1 | 4 | 4 | ≤0.25 | ≤0.25 | 4 | 2 | ≤0.125 |
| pCR4_SEG8_cef01 | 1 | ≤1 | 4 | ≤0.25 | ≤0.25 | 16 | 8 | 64 |
| pCR4_SEW5_chl01 | 64 | 4 | 4 | 0.5 | ≤0.25 | 4 | 2 | ≤0.125 |
| pCR4_AEW1_fos01 | 1 | ≥512 | 4 | ≤0.25 | ≤0.25 | 4 | 2 | ≤0.125 |
| pCR4_AEW5_tri01 | 1 | 4 | 4 | ≤0.25 | ≥128 | 4 | 2 | ≤0.125 |
| pCR4_SEG8_tri01 | 1 | 4 | 4 | ≤0.25 | ≥128 | 4 | 2 | ≤0.125 |
| pCR4_SEG8_tri02 | 1 | 4 | 4 | ≤0.25 | ≥128 | 4 | 2 | ≤0.125 |
| Plasmid | Minimum Inhibitory Concentration (µg/mL) | |||||
|---|---|---|---|---|---|---|
| AMP | CRB | PIP | CPH | CFX | CFT | |
| Cloning vector (pHSG398) | ≤2 | ≤2 | ≤2 | 4 | 2 | ≤0.125 |
| pHSG398_AEW4_amp01 | ≥1024 | ≥1024 | 512 | 4 | 2 | ≤0.125 |
| pHSG398_SEG8_amp01 | ≥1024 | ≥1024 | 512 | 4 | 2 | ≤0.125 |
| Template | Oligonucleotide | Sequence (5′ to 3′) | Annealing Temperature ( °C) |
|---|---|---|---|
| pLAEW4_amp01 | AEW4_amp01_for_150 | CCATGATTACGAATTCTTATTTGTTCGGCGTCTTGC | 60 |
| AEW4_amp01_rev | TACCGAGCTCGAATTCTAATGCACTGACTTCTGCAAG | 60 | |
| pLSEG8_amp01 | SEG8_amp01_for_46 | CCATGATTACGAATTGTTCGACAAGGACCACATC | 60 |
| SEG8_amp01_rev | TACCGAGCTCGAATTTCACTCGTCCTCCGGCCC | 60 | |
| pLSEG8_cef01 | SEG8_cef01_for_150 | GAGATTGCTTCGTCGCTCTG | 59 |
| SEG8_cef01_rev | TCACTTCCTCTGGGCAACATTC | 59 | |
| pLSEW5_chl01 | SEW5_chl01_for_150 | ACCATTTGCTATAATAGCATCTAC | 53 |
| SEW5_chl01_rev | TCATCTCTTTCTCGGTATCAG | 53 | |
| pLAEW1_fos01 | AEW1_fos01_for_150 | CCAAAATAACAGGGGGCG | 55 |
| AEW1_fos01_rev | TCACACCCTCGTAATATCCG | 55 | |
| pLAEW5_tri01 | AEW5_tri01_for_150 | GCGGCGATTGCGAGTAG | 59 |
| AEW5_tri01_rev | TCACAACGTCCGCCCG | 59 | |
| pLSEG8_tri01 | SEG8_tri01_for_150 | ACTTACTCAGAATTTGACATA | 49 |
| SEG8_tri01_rev | TTAATTGGCTCTCACATAAG | 49 | |
| pLSEG8_tri02 | SEG8_tri02_for_145 | CGGACCCCTCAGAAC | 52 |
| SEG8_tri02_rev | TTACTTCTTTTCATAGATAACAAAGTC | 52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Willms, I.M.; Grote, M.; Kocatürk, M.; Singhoff, L.; Kraft, A.A.; Bolz, S.H.; Nacke, H. Novel Soil-Derived Beta-Lactam, Chloramphenicol, Fosfomycin and Trimethoprim Resistance Genes Revealed by Functional Metagenomics. Antibiotics 2021, 10, 378. https://doi.org/10.3390/antibiotics10040378
Willms IM, Grote M, Kocatürk M, Singhoff L, Kraft AA, Bolz SH, Nacke H. Novel Soil-Derived Beta-Lactam, Chloramphenicol, Fosfomycin and Trimethoprim Resistance Genes Revealed by Functional Metagenomics. Antibiotics. 2021; 10(4):378. https://doi.org/10.3390/antibiotics10040378
Chicago/Turabian StyleWillms, Inka Marie, Maja Grote, Melissa Kocatürk, Lukas Singhoff, Alina Andrea Kraft, Simon Henning Bolz, and Heiko Nacke. 2021. "Novel Soil-Derived Beta-Lactam, Chloramphenicol, Fosfomycin and Trimethoprim Resistance Genes Revealed by Functional Metagenomics" Antibiotics 10, no. 4: 378. https://doi.org/10.3390/antibiotics10040378
APA StyleWillms, I. M., Grote, M., Kocatürk, M., Singhoff, L., Kraft, A. A., Bolz, S. H., & Nacke, H. (2021). Novel Soil-Derived Beta-Lactam, Chloramphenicol, Fosfomycin and Trimethoprim Resistance Genes Revealed by Functional Metagenomics. Antibiotics, 10(4), 378. https://doi.org/10.3390/antibiotics10040378