
A Complementary Herbal Product for Controlling Giardiasis

 , ,
, ,  , ,
, ,  and
and 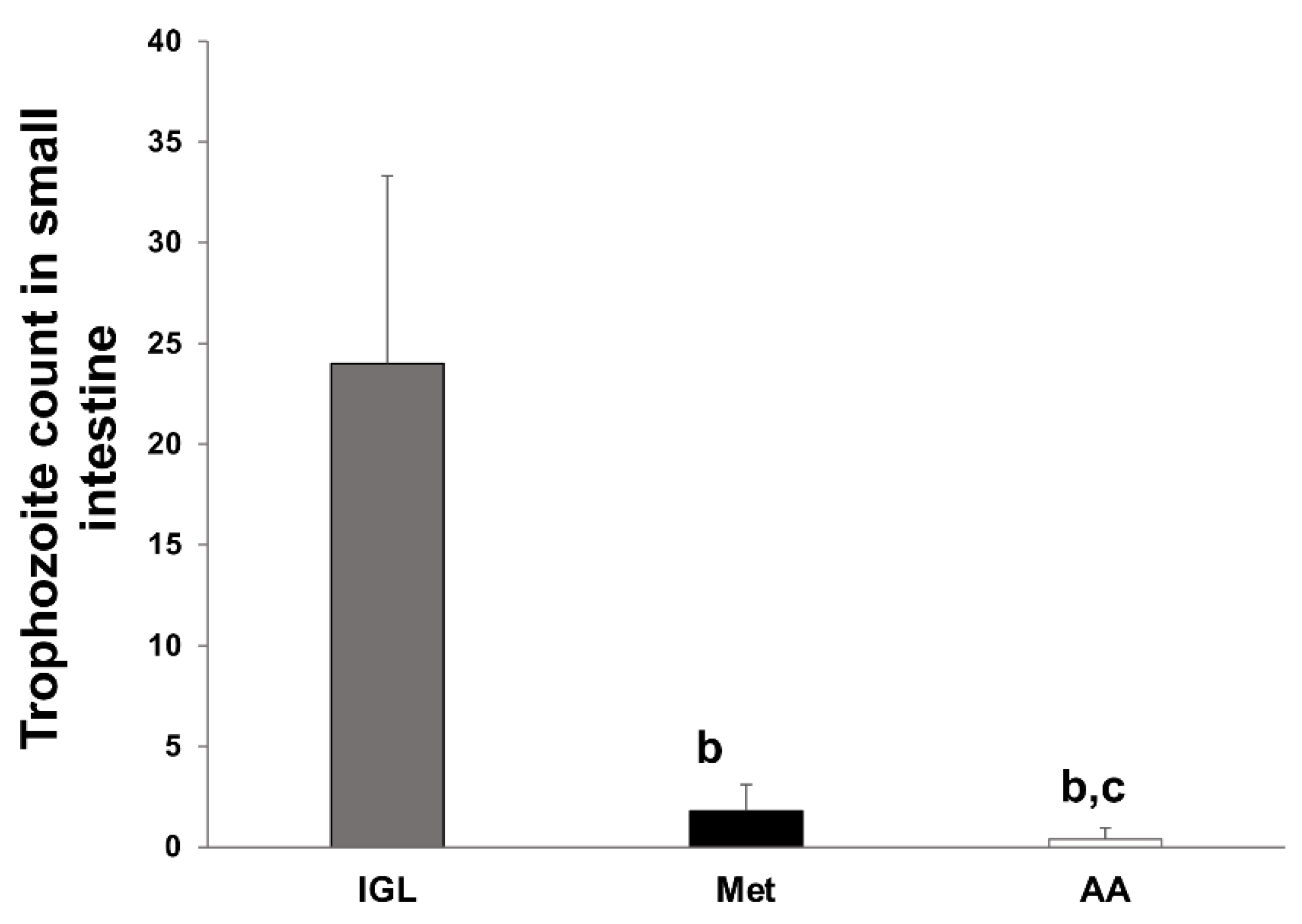{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Plant Material
2.3. Preparation of A. annua Extract
2.4. Preparation of Parasite for Infection
2.5. Animal Experiments
2.6. Assessment of the Efficacy of the Extract
2.6.1. Determination of Trophozoites Count in Different Groups of Infected Hamsters
2.6.2. Quantification of Proinflammatory Cytokines and NO
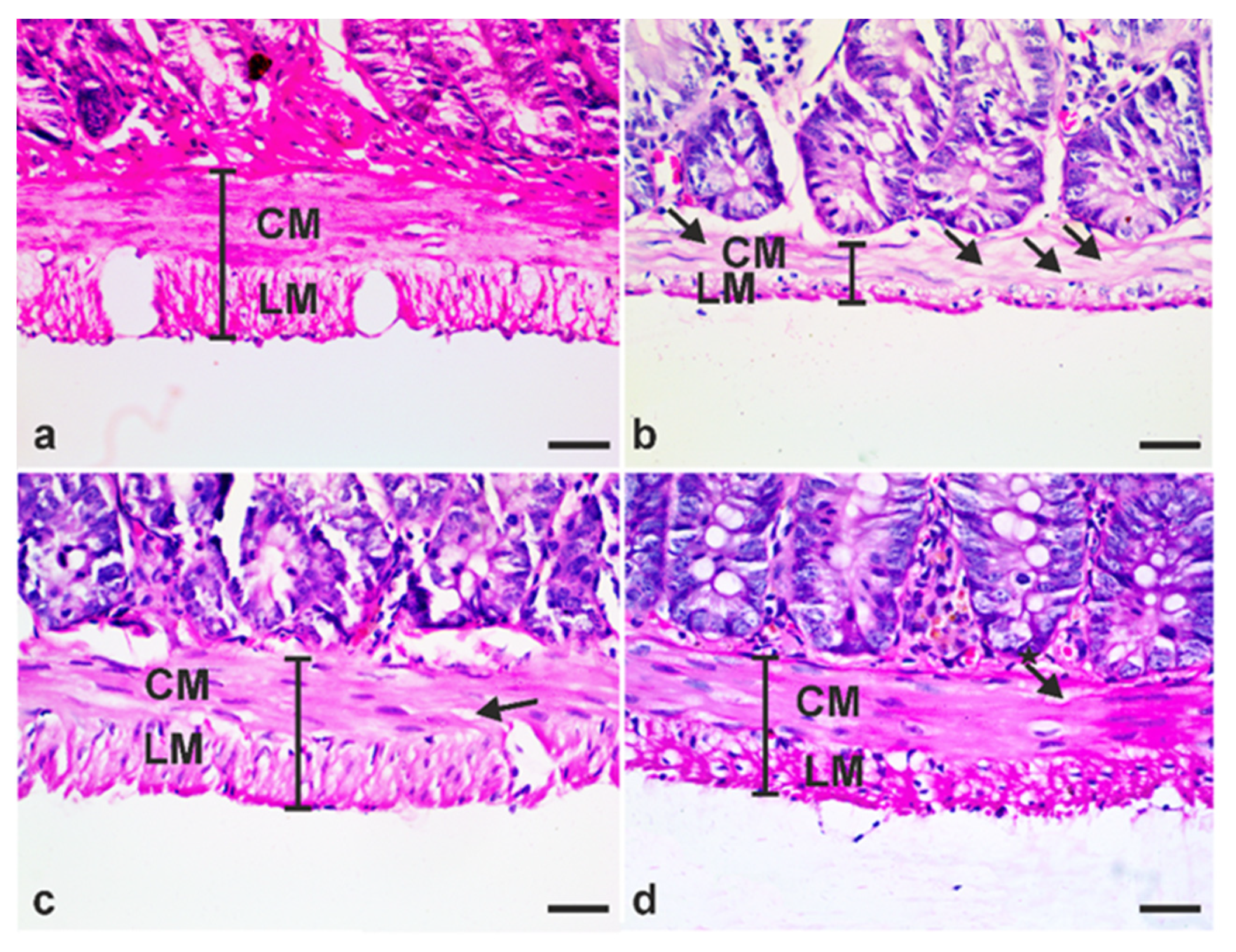
2.7. Histopathological Studies
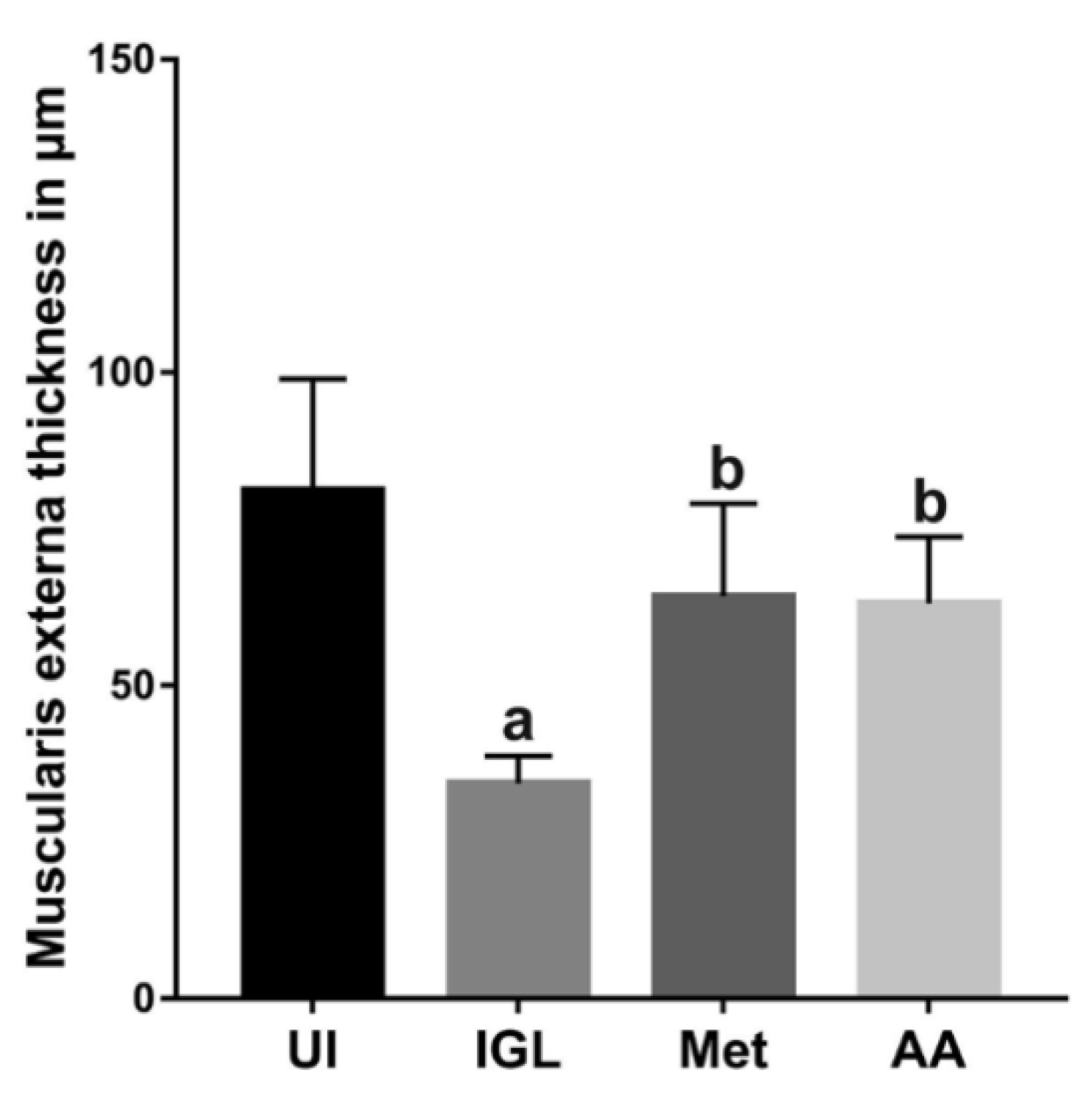
2.7.1. Assessment of Villi Length and Muscularis Externa Thickness
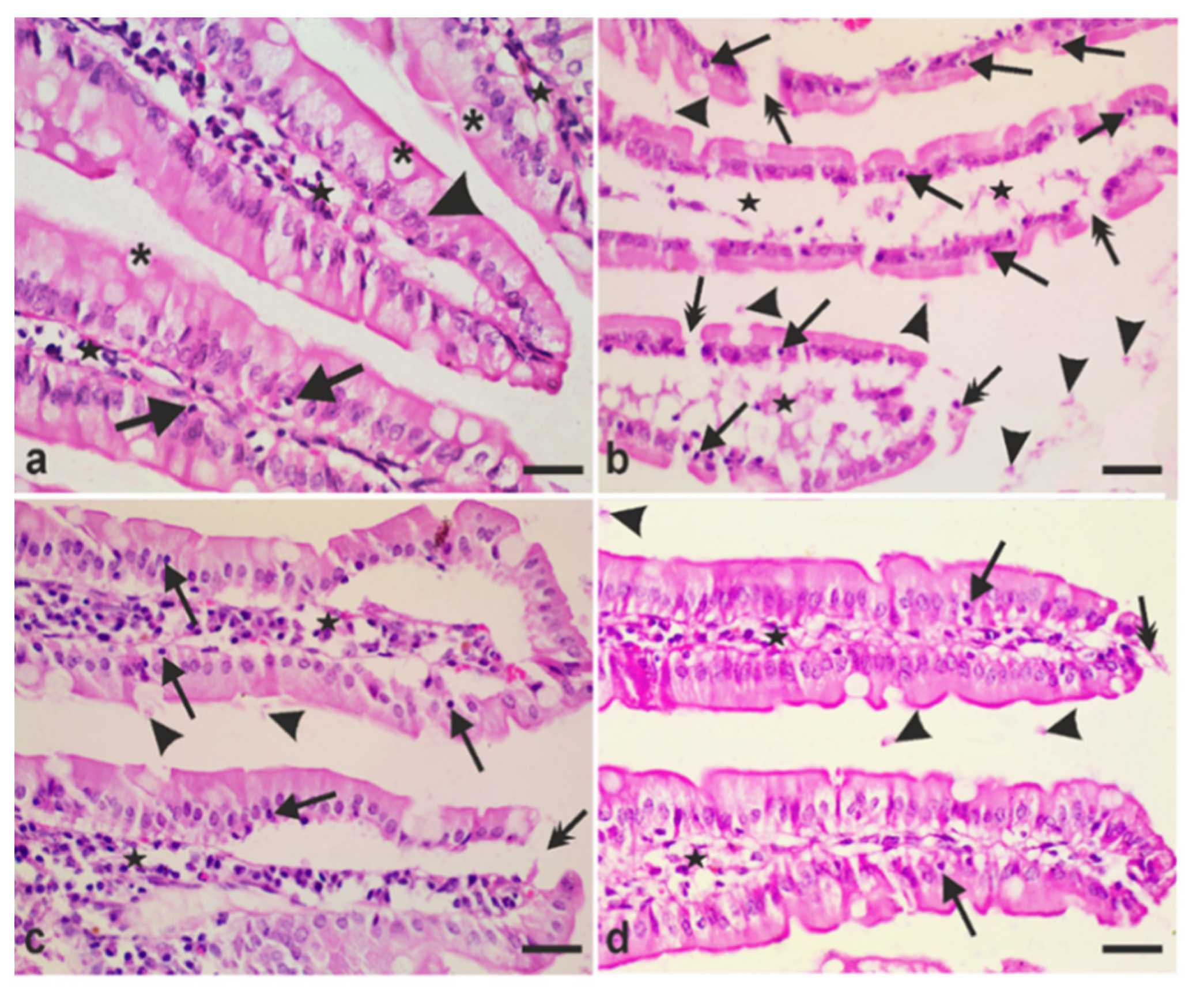
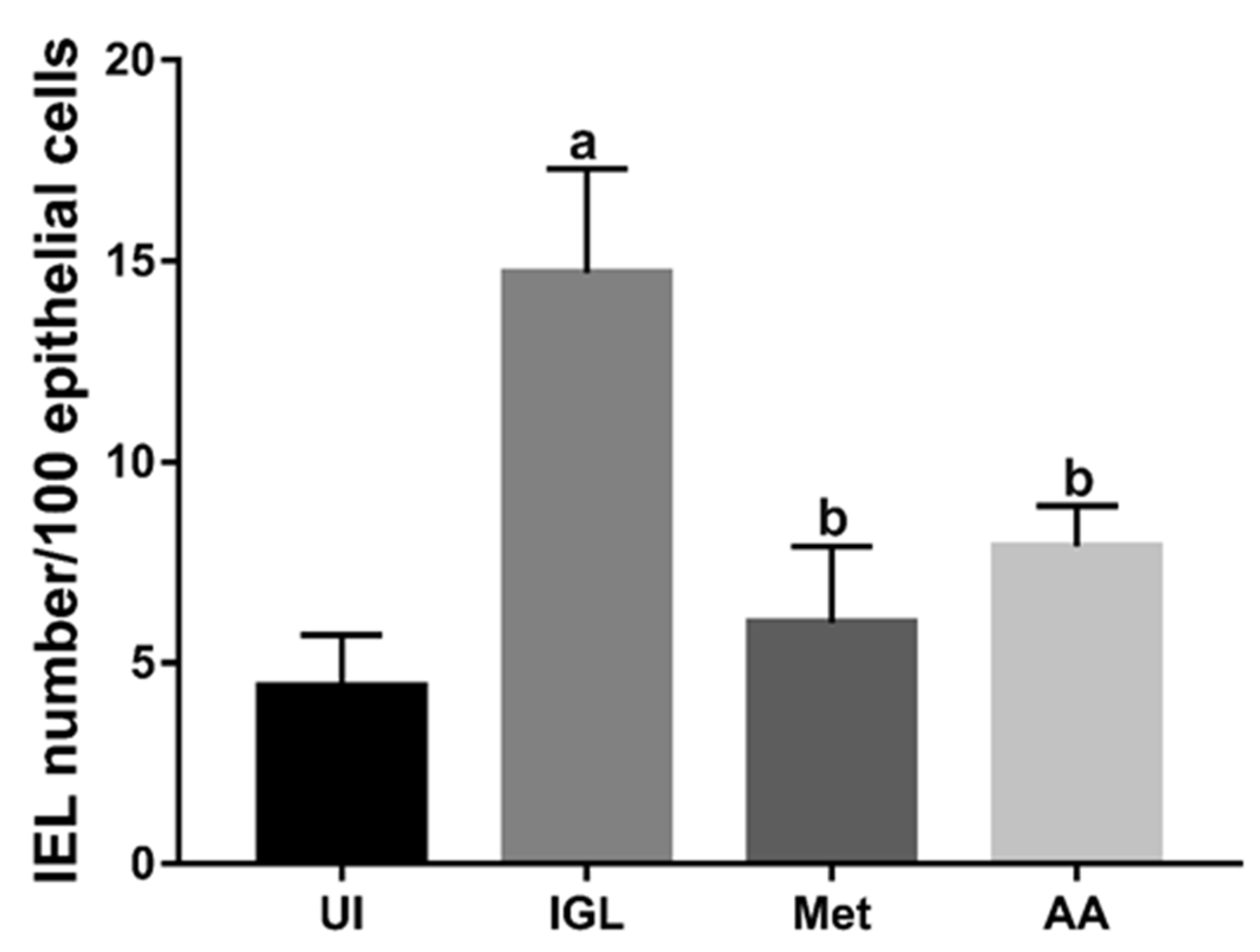
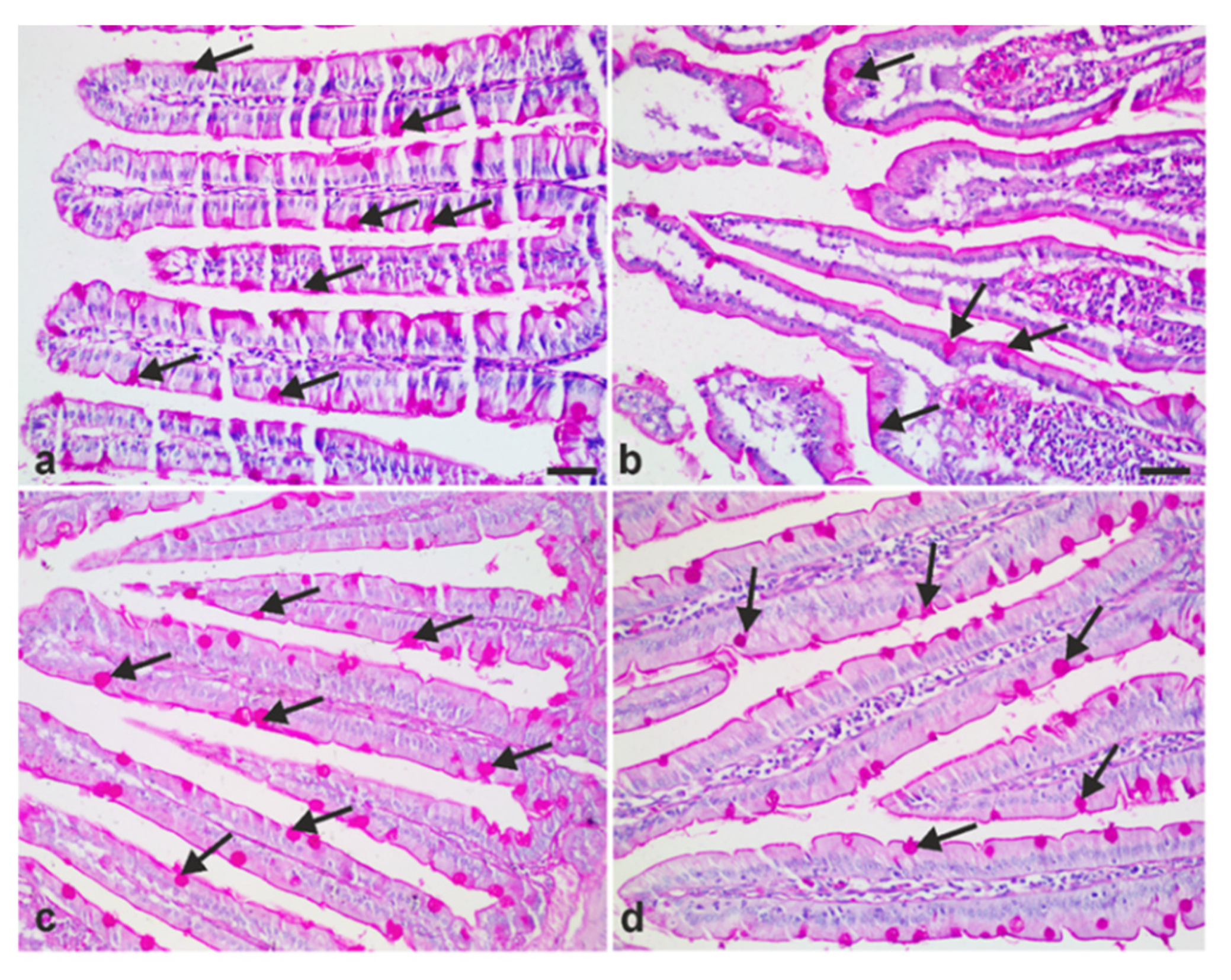
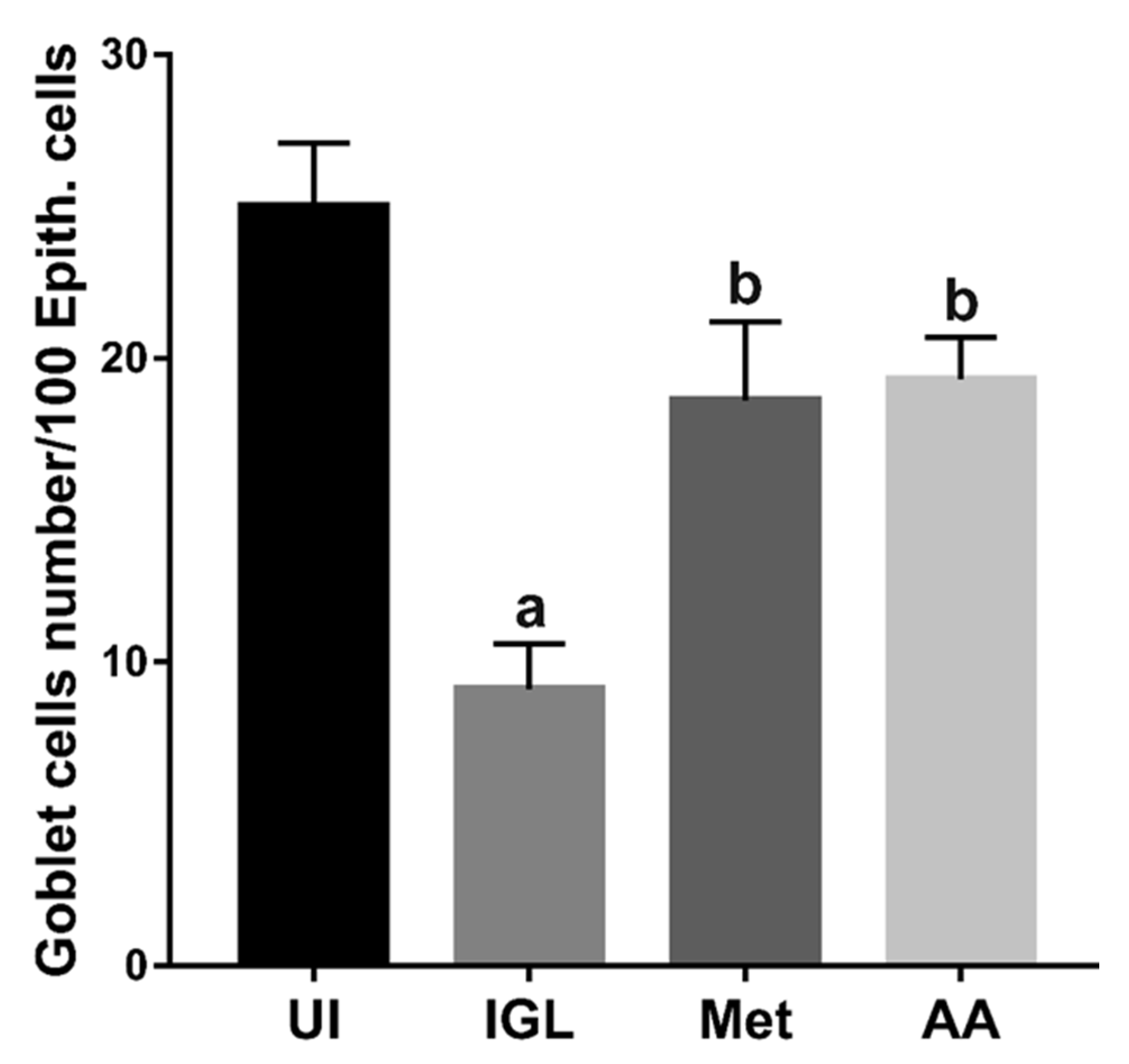
2.7.2. Determination of Intraepithelial Lymphocytes (IEL) and Goblet Cell Numbers
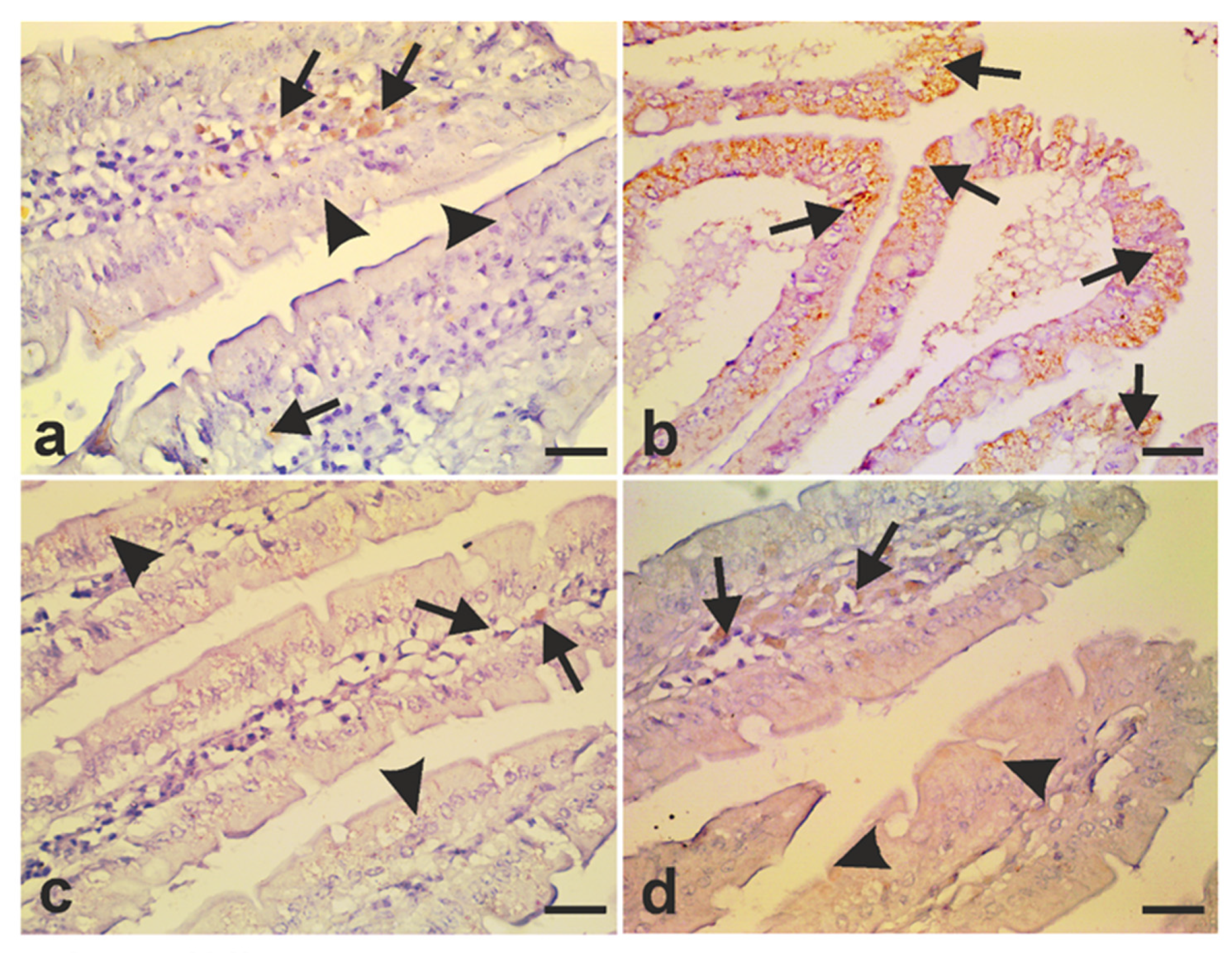
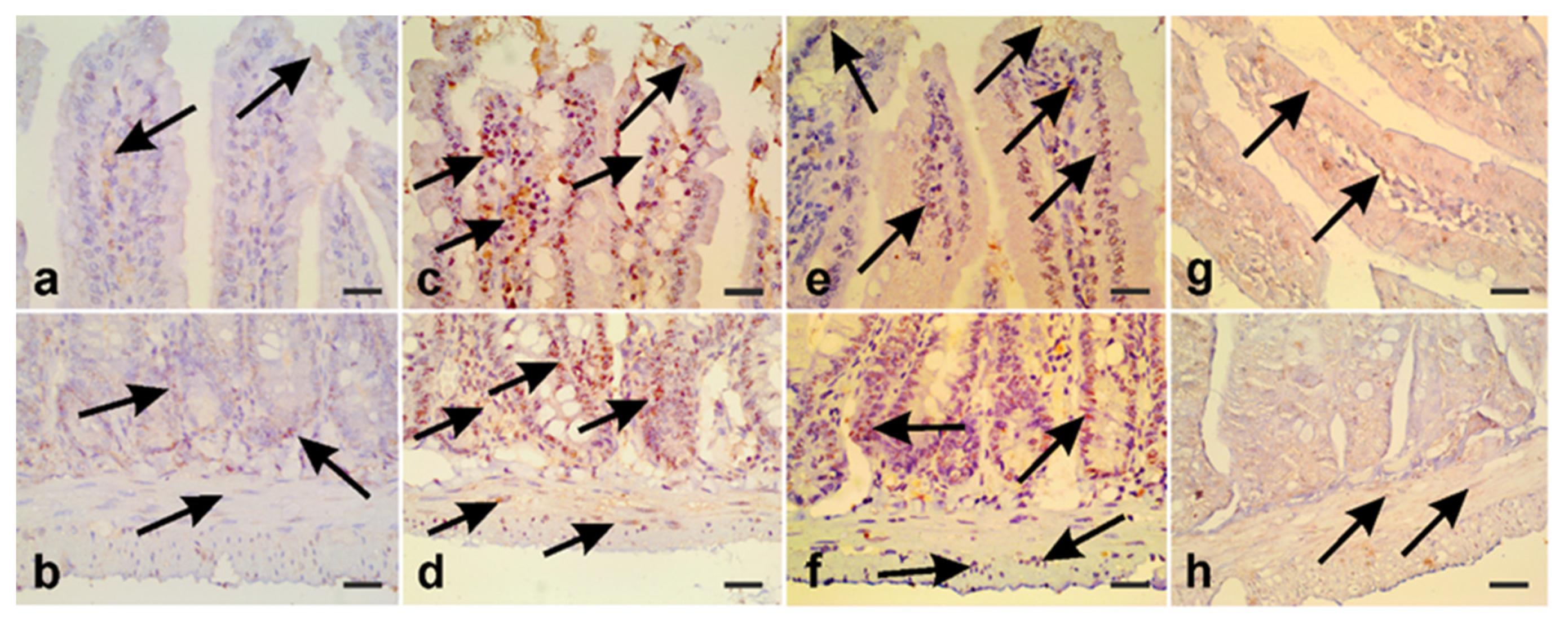
2.7.3. Immunohistochemistry
2.8. Statistical Analysis
3. Results
3.1. A. annua Treatment Reduces G. lamblia Trophozoite Count in the Intestine
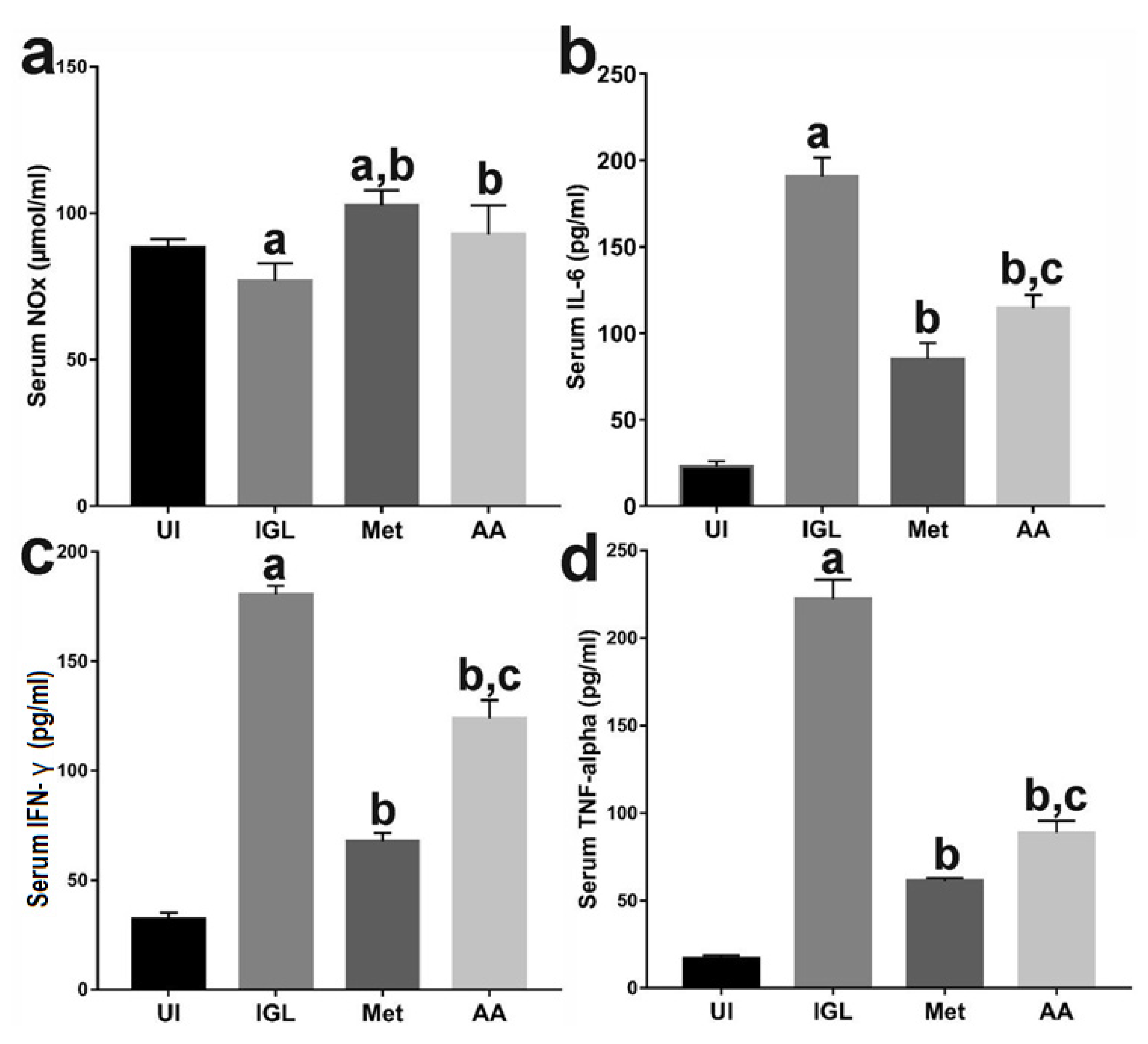
3.2. A. annua Treatment Induced NO and Reduced Proinflammatory Cytokines Production
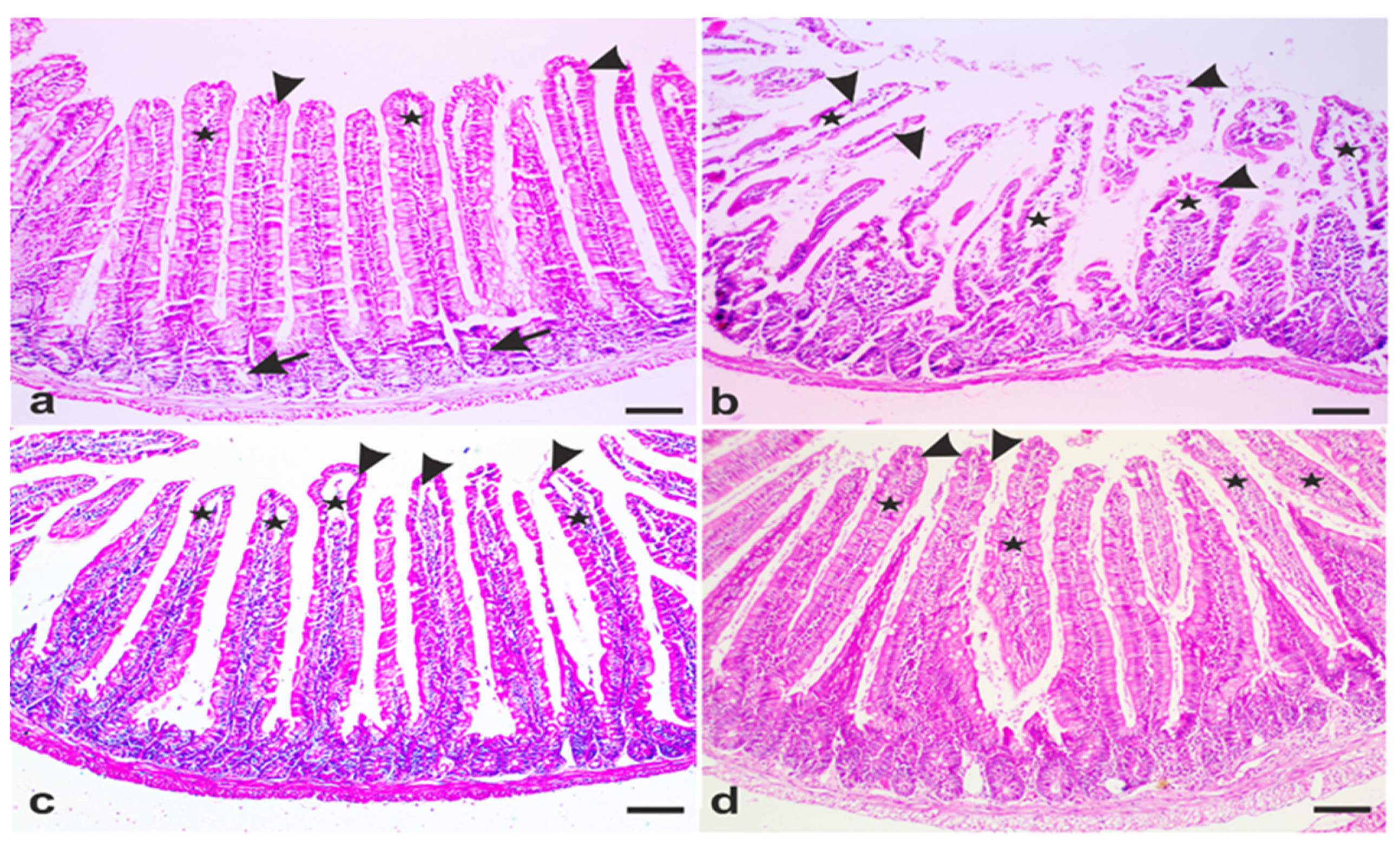
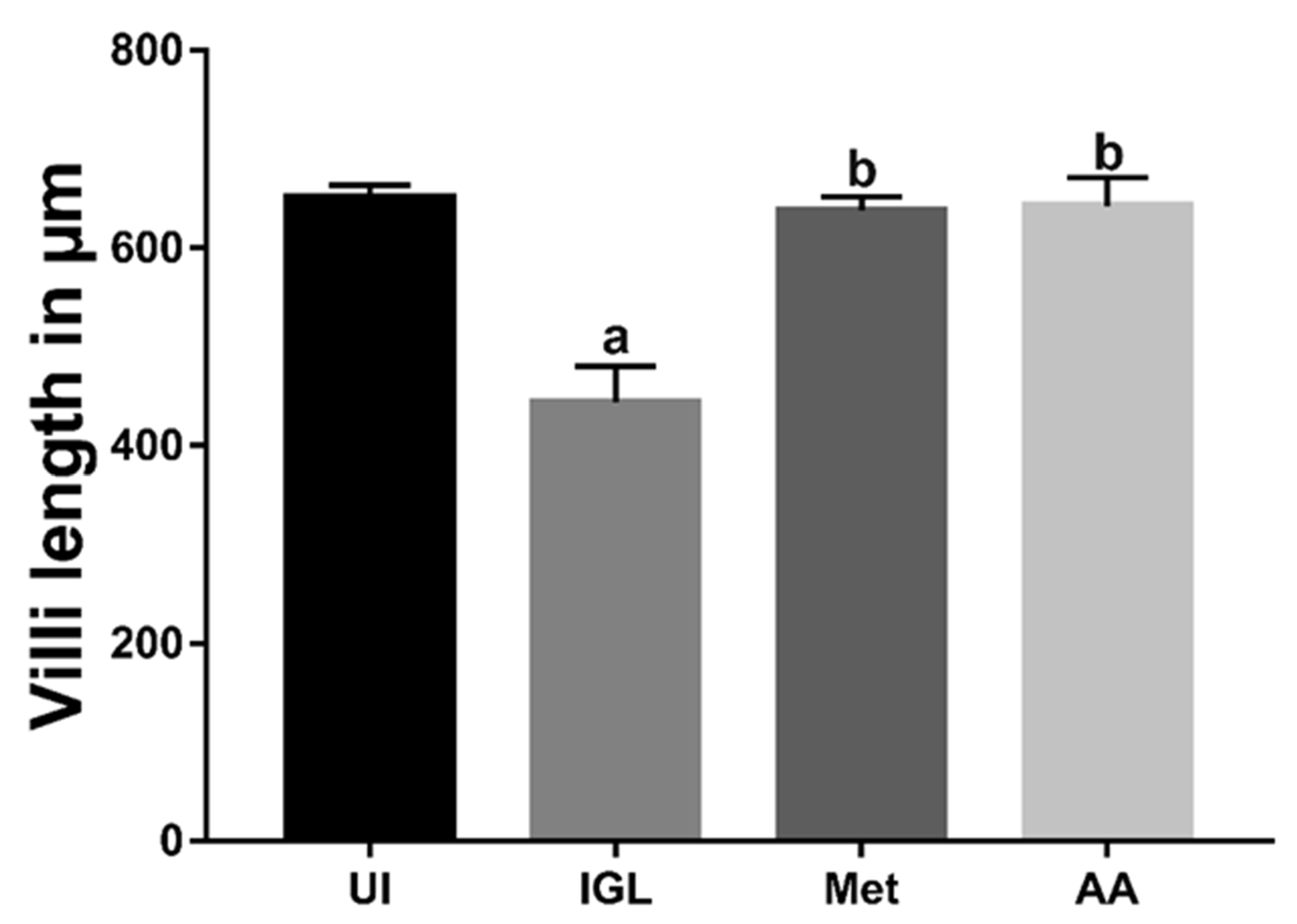
3.3. A. annua Treatment Restored Villi Structure of G. lamblia-Infected Animals
3.4. A. annua Treatment Reduced IEL Numbers in G. lamblia-Infected Animals, Restored the Goblet Cell Numbers, and Reversed Infection-Induced Pathological Effect on Muscularis Externa
3.5. A. annua Treatment Modulates iNOS Expression and Protects Intestinal Cells from Apoptosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| A. annua | Artemisia annua |
| HPF | High power field |
| IEL | Intraepithelial Lymphocytes |
| IL-6 | Interlukin-6 |
| IFN-γ | Interferon-γ |
| iNOS | Inducible nitric oxide synthase |
| NF-kB | Nuclear factor-kappa B |
| nNOS | Neuronal nitric oxide synthase |
| NO | Nitric oxide |
| PAS | Periodic Acid Schiff reagent |
| TNF-α | Tumor necrosis factor-α |
| TNFR1 | TNF-α receptor 1 |
References
- Gardner, T.B.; Hill, D.R. Treatment of giardiasis. Clin. Microbiol. Rev. 2001, 14, 114–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam, R.D. Biology of Giardia lamblia. Clin. Microbiol. Rev. 2001, 14, 447–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ankarklev, J.; Jerlstrom-Hultqvist, J.; Ringqvist, E.; Troell, K.; Svard, S.G. Behind the smile: Cell biology and disease mechanisms of Giardia species. Nat. Rev. Microbiol. 2010, 8, 413–422. [Google Scholar] [CrossRef]
- Thompson, R.C. Giardiasis as a re-emerging infectious disease and its zoonotic potential. Int. J. Parasitol. 2000, 30, 1259–1267. [Google Scholar] [CrossRef]
- Foronda, P.; Bargues, M.D.; Abreu-Acosta, N.; Periago, M.V.; Valero, M.A.; Valladares, B.; Mas-Coma, S. Identification of genotypes of Giardia intestinalis of human isolates in Egypt. Parasitol. Res. 2008, 103, 1177–1181. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, S.M.; El-Bahy, M.M.; Abd Elaziz, J.; El-Dardiry, M.A. Detection of protozoal parasites in the stools of diarrhoeic patients using different techniques. J. Egypt Soc. Parasitol. 2006, 36, 7–22. [Google Scholar]
- Baiomy, A.M.; Mohamed, K.A.; Ghannam, M.A.; Shahat, S.A.; Al-Saadawy, A.S. Opportunistic parasitic infections among immunocompromised Egyptian patients. J. Egypt Soc. Parasitol. 2010, 40, 797–808. [Google Scholar]
- Abdel-Hafeez, E.H.; Ahmad, A.K.; Ali, B.A.; Moslam, F.A. Opportunistic parasites among immunosuppressed children in Minia District, Egypt. Korean J. Parasitol. 2012, 50, 57–62. [Google Scholar] [CrossRef]
- Feng, Y.; Xiao, L. Zoonotic potential and molecular epidemiology of Giardia species and giardiasis. Clin. Microbiol. Rev. 2011, 24, 110–140. [Google Scholar] [CrossRef] [Green Version]
- Eppig, C.; Fincher, C.L.; Thornhill, R. Parasite prevalence and the worldwide distribution of cognitive ability. Proc. Biol. Sci. 2010, 277, 3801–3808. [Google Scholar] [CrossRef]
- Balcioglu, I.C.; Limoncu, E.; Ertan, P.; Yereli, K.; Ozbilgin, A.; Onag, A. Incidence of giardiasis among siblings in Turkey. Pediatr. Int. 2003, 45, 311–313. [Google Scholar] [CrossRef]
- Veenemans, J.; Mank, T.; Ottenhof, M.; Baidjoe, A.; Mbugi, E.V.; Demir, A.Y.; Wielders, J.P.; Savelkoul, H.F.; Verhoef, H. Protection against diarrhea associated with Giardia intestinalis Is lost with multi-nutrient supplementation: A study in Tanzanian children. PLoS Negl. Trop. Dis. 2011, 5, e1158. [Google Scholar] [CrossRef]
- Gonçalves, A.C.M.; Gabbay, Y.B.; Mascarenhas, J.D.; Yassaka, M.B.; Moran, L.C.; Fraga, V.D.; Castro, E.; Franco, C.; Machado, R.L.D.; Rossit, A.R.B. Calicivirus and Giardia lamblia are associated with diarrhea in human immunodeficiency virus-seropositive patients from southeast Brazil. Am. J. Trop. Med. Hyg. 2009, 81, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Reynoso, R.; Ponce, M.; Rosas, L.E.; Ramos, A.; Mart, M.N.; Gonz, A. The invasive potential of Giardia intestinalis in an in vivo model. Sci. Rep. 2015, 16, 1–8. [Google Scholar] [CrossRef]
- Panarelli, N.C.; Gobara, N.; Hoda, R.S.; Chaump, M.; Jessurun, J.; Yantiss, R.K. Cytology Preparations of Formalin Fixative Aid Detection of Giardia in Duodenal Biopsy Samples. Am. J. Surg. Pathol. 2017, 41, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Ventura, L.L.; Oliveira, D.R.; Viana, J.C.; Santos, J.F.; Caliari, M.V.; Gomes, M.A. Impact of protein malnutrition on histological parameters of experimentally infected animals with Giardia lamblia. Exp. Parasitol. 2013, 133, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Panaro, M.A.; Cianciulli, A.; Mitolo, V.; Mitolo, C.I.; Acquafredda, A.; Brandonisio, O.; Cavallo, P. Caspase-dependent apoptosis of the HCT-8 epithelial cell line induced by the parasite Giardia intestinalis. FEMS Immunol. Med. Microbiol. 2007, 51, 302–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, A.C.; Teoh, D.A.; Scott, K.G.-E.; Meddings, J.B.; Macnaughton, W.K.; Buret, A.G. Strain-dependent induction of enterocyte apoptosis by Giardia lamblia disrupts epithelial barrier function in a caspase-3-dependent manner. Infect. Immun. 2002, 70, 3673–3680. [Google Scholar] [CrossRef] [Green Version]
- Pavanelli, M.F.; Colli, C.M.; Bezagio, R.C.; Gois, M.B.; de Melo, G.d.A.N.; de Almeida Araujo, E.J.; Sant’Ana, D.d.M.G. Assemblages A and B of Giardia duodenalis reduce enteric glial cells in the small intestine in mice. Parasitol. Res. 2018, 117, 2025–2033. [Google Scholar] [CrossRef]
- Zhou, P.; Li, E.; Zhu, N.; Robertson, J.; Nash, T.; Singer, S.M. Role of interleukin-6 in the control of acute and chronic Giardia lamblia infections in mice. Infect. Immun. 2003, 71, 1566–1568. [Google Scholar] [CrossRef] [Green Version]
- Bienz, M.; Dai, W.J.; Welle, M.; Gottstein, B.; Müller, N. Interleukin-6-deficient mice are highly susceptible to Giardia lamblia infection but exhibit normal intestinal immunoglobulin A responses against the parasite. Infect. Immun. 2003, 71, 1569–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.Y.; Kim, J.; Noh, H.J.; Kim, H.P.; Park, S.J. Giardia lamblia binding immunoglobulin protein triggers maturation of dendritic cells via activation of TLR4-MyD88-p38 and ERK1/2 MAPKs. Parasite Immunol. 2014, 36, 627–646. [Google Scholar] [CrossRef] [PubMed]
- Kamda, J.D.; Nash, T.E.; Singer, S.M. Giardia duodenalis: Dendritic cell defects in IL-6 deficient mice contribute to susceptibility to intestinal infection. Exp. Parasitol. 2012, 130, 288–291. [Google Scholar] [CrossRef] [Green Version]
- Eckmann, L.; Laurent, F.; Langford, T.D.; Hetsko, M.L.; Smith, J.R.; Kagnoff, M.F.; Gillin, F.D. Nitric oxide production by human intestinal epithelial cells and competition for arginine as potential determinants of host defense against the lumen-dwelling pathogen Giardia lamblia. J. Immunol. 2000, 164, 1478–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovic, P.J.; Zeh, H.J., 3rd; Ochoa, J.B. Arginine and immunity. J. Nutr. 2007, 137, 1681S–1686S. [Google Scholar] [CrossRef] [Green Version]
- Das, P.; Lahiri, A.; Lahiri, A.; Chakravortty, D. Modulation of the arginase pathway in the context of microbial pathogenesis: A metabolic enzyme moonlighting as an immune modulator. PLoS Pathog. 2010, 6, e1000899. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.M.; Upcroft, J.A.; Edwards, M.R.; Upcroft, P. Anaerobic bacterial metabolism in the ancient eukaryote Giardia duodenalis. Int. J. Parasitol. 1998, 28, 149–164. [Google Scholar] [CrossRef]
- Maloney, J.; Keselman, A.; Li, E.; Singer, S.M. Macrophages expressing arginase 1 and nitric oxide synthase 2 accumulate in the small intestine during Giardia lamblia infection. Microbes Infect. 2015, 17, 462–467. [Google Scholar] [CrossRef] [Green Version]
- Stadelmann, B.; Hanevik, K.; Andersson, M.K.; Bruserud, O.; Svärd, S.G. The role of arginine and arginine-metabolizing enzymes during Giardia—host cell interactions in vitro. BMC Microbiol. 2013, 13, 256. [Google Scholar] [CrossRef] [Green Version]
- Dupouy-Camet, J. New drugs for the treatment of human parasitic protozoa. Parassitologia 2004, 46, 81–84. [Google Scholar]
- Lalle, M. Giardiasis in the post genomic era: Treatment, drug resistance and novel therapeutic perspectives. Infect Disord. Drug Targets 2010, 10, 283–294. [Google Scholar] [CrossRef]
- Huang, D.B.; White, A.C. An updated review on Cryptosporidium and Giardia. Gastroenterol. Clin. N. Am. 2006, 35, 291–314. [Google Scholar] [CrossRef]
- Upcroft, J.A.; Dunn, L.A.; Wright, J.M.; Benakli, K.; Upcroft, P.; Vanelle, P. 5-Nitroimidazole drugs effective against metronidazole-resistant Trichomonas vaginalis and Giardia duodenalis. Antimicrob. Agents Chemother. 2006, 50, 344–347. [Google Scholar] [CrossRef] [Green Version]
- Long, K.Z.; Rosado, J.L.; Montoya, Y.; de Lourdes Solano, M.; Hertzmark, E.; DuPont, H.L.; Santos, J.I. Effect of vitamin A and zinc supplementation on gastrointestinal parasitic infections among Mexican children. Pediatrics 2007, 120, e846–e855. [Google Scholar] [CrossRef] [PubMed]
- Sofowora, A.; Ogunbodede, E.; Onayade, A. The role and place of medicinal plants in the strategies for disease prevention. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 210–229. [Google Scholar] [CrossRef]
- Bora, K.S.; Sharma, A. The genus Artemisia: A comprehensive review. Pharm. Biol. 2011, 49, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Valderrábano, J.; Calvete, C.; Uriarte, J. Effect of feeding bioactive forages on infection and subsequent development of Haemonchus contortus in lamb faeces. Vet. Parasitol. 2010, 172, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Daddy, N.B.; Kalisya, L.M.; Bagire, P.G.; Watt, R.L.; Towler, M.J.; Weathers, P.J. Artemisia annua dried leaf tablets treated malaria resistant to ACT and i.v. artesunate: Case reports. Phytomedicine 2017, 32, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.F.; Luthria, D.L.; Sasaki, T.; Heyerick, A. Flavonoids from Artemisia annua L. as antioxidants and their potential synergism with artemisinin against malaria and cancer. Molecules 2010, 15, 3135–3170. [Google Scholar] [CrossRef] [Green Version]
- Chukwurah, P.N.; Brisibe, E.A.; Osuagwu, A.N.; Okoko, T. Protective capacity of Artemisia annua as a potent antioxidant remedy against free radical damage. Asian Pac. J. Trop. Biomed. 2014, 4, S92–S98. [Google Scholar] [CrossRef] [Green Version]
- Mishina, Y.V.; Krishna, S.; Haynes, R.K.; Meade, J.C. Artemisinins inhibit Trypanosoma cruzi and Trypanosoma brucei rhodesiense in vitro growth. Antimicrob. Agents Chemother. 2007, 51, 1852–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, J.F.; Peaden, P.; Keiser, J. In vitro trematocidal effects of crude alcoholic extracts of Artemisia annua, A. absinthium, Asimina triloba, and Fumaria officinalis: Trematocidal plant alcoholic extracts. Parasitol. Res. 2011, 109, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, T.C.; Silva, D.A.; Rostkowska, C.; Bela, S.R.; Ferro, E.A.; Magalhaes, P.M.; Mineo, J.R. Toxoplasma gondii: Effects of Artemisia annua L. on susceptibility to infection in experimental models in vitro and in vivo. Exp. Parasitol. 2009, 122, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Sen, R.; Bandyopadhyay, S.; Dutta, A.; Mandal, G.; Ganguly, S.; Saha, P.; Chatterjee, M. Artemisinin triggers induction of cell-cycle arrest and apoptosis in Leishmania donovani promastigotes. J. Med. Microbiol. 2007, 56, 1213–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesa, L.E.; Vasquez, D.; Lutgen, P.; Velez, I.D.; Restrepo, A.M.; Ortiz, I.; Robledo, S.M. In vitro and in vivo antileishmanial activity of Artemisia annua L. leaf powder and its potential usefulness in the treatment of uncomplicated cutaneous leishmaniasis in humans. Rev. Soc. Bras. Med. Trop. 2017, 50, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Jiao, J.; Yang, Y.; Liu, M.; Li, J.; Cui, Y.; Yin, S.; Tao, J. Artemisinin and Artemisia annua leaves alleviate Eimeria tenella infection by facilitating apoptosis of host cells and suppressing inflammatory response. Vet. Parasitol. 2018, 254, 172–177. [Google Scholar] [CrossRef]
- Golami, S.; Rahimi-Esboei, B.; Mousavi, P.; Marhaba, Z.; Youssefi, M.R.; Rahimi, M.T. Survey on efficacy of chloroformic extract of Artemisia annua against Giardia lamblia trophozoite and cyst in vitro. J. Parasite Dis. 2016, 40, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Tariq, K.A.; Chishti, M.Z.; Ahmad, F.; Shawl, A.S. Anthelmintic activity of extracts of Artemisia absinthium against ovine nematodes. Vet. Parasitol. 2009, 160, 83–88. [Google Scholar] [CrossRef]
- Dyab, A.K.; Yones, D.A.; Ibraheim, Z.Z.; Hassan, T.M. Anti-giardial therapeutic potential of dichloromethane extracts of Zingiber officinale and Curcuma longa in vitro and in vivo. Parasitol. Res. 2016, 115, 2637–2645. [Google Scholar] [CrossRef]
- Aly, E.M.; Sabry, H.Y.; Fahmy, Z.H.; Zalat, R.S. Ef fi cacy of combination therapy (metronidazole and/or artemether) in experimental giardiasis and its impact on nonenzymatic oxidative stress biomarkers. Parasitol. United J. 2014, 68–74. [Google Scholar] [CrossRef]
- Squires, J.M.; Ferreira, J.F.; Lindsay, D.S.; Zajac, A.M. Effects of artemisinin and Artemisia extracts on Haemonchus contortus in gerbils (Meriones unguiculatus). Vet. Parasitol. 2011, 175, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Laurence, D.R.; Bacharach, A.L. Evaluation of Drug Activities: Pharmacometrics; Academic Press: London, UK, 1964. [Google Scholar]
- Fathy, F.M. Effect of mirazid (Commiphora molmol) on experimental giardiasis. J. Egypt Soc. Parasitol. 2011, 41, 155–177. [Google Scholar] [PubMed]
- Rayan, P.; Stenzel, D.; McDonnell, P.A. The effects of saturated fatty acids on Giardia duodenalis trophozoites in vitro. Parasitol. Res. 2005, 97, 191–200. [Google Scholar] [CrossRef]
- Nuttall, K.L.; Gordon, W.H.; Ash, K.O. Inductively coupled plasma mass spectrometry for trace element analysis in the clinical laboratory. Ann. Clin. Lab. Sci. 1995, 25, 264–271. [Google Scholar] [PubMed]
- Drury, R.A.B.; Wallington, E.A.; Cameron, R. Carleton’s Histological Technique (Oxford Medical Publications), 5th ed.; Oxford University Press: Oxford, UK, 1980. [Google Scholar]
- Chott, A.; Gerdes, D.; Spooner, A.; Mosberger, I.; Kummer, J.A.; Ebert, E.C.; Blumberg, R.S.; Balk, S.P. Intraepithelial lymphocytes in normal human intestine do not express proteins associated with cytolytic function. Am. J. Pathol. 1997, 151, 435–442. [Google Scholar] [PubMed]
- Pacheco, F.T.F.; Carvalho, S.S.; Cardoso, L.S.; Andrade, L.S.; das Chagas, G.M.T.; Gomes, D.C.; Merces, C.F.; Rocha, F.C.; Silva, L.K.; Soares, N.M.; et al. Immune response markers in sera of children infected with Giardia duodenalis AI and AII subassemblages. Immunobiology 2019, 224, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Mello, R.d.O.; da Silva, C.M.G.; Fonte, F.P.; Silva, D.L.F.; Pereira, J.A.; Margarido, N.F.; Martinez, C.A.R. Evaluation of the number of goblet cells in crypts of the colonic mucosa with and without fecal transit. Rev. Col. Bras. Cir. 2012, 39, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Mokrzycka, M.; Kolasa, A.; Kosierkiewicz, A.; Wiszniewska, B. Inducible nitric oxide synthase in duodenum of children with Giardia lamblia infection. Folia Histochem. Cytobiol. 2010, 48, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.S.; Choi, W.J.; Lee, S.; Kim, W.J.; Lee, D.C.; Sohn, U.D.; Shin, H.S.; Kim, W. Anti-inflammatory, Antioxidant and Antimicrobial Effects of Artemisinin Extracts from Artemisia annua L. Korean J. Physiol. Pharmacol. 2015, 19, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Calzada, F.; Yépez-Mulia, L.; Aguilar, A. In vitro susceptibility of Entamoeba histolytica and Giardia lamblia to plants used in Mexican traditional medicine for the treatment of gastrointestinal disorders. J. Ethnopharmacol. 2006, 108, 367–370. [Google Scholar] [CrossRef]
- Rahimi-Esboei, B.; Ebrahimzadeh, M.A.; Gholami, S.; Falah-Omrani, V. Anti-giardial activity of Sambucus ebulus. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2047–2050. [Google Scholar] [PubMed]
- Alin, M.H.; Bjorkman, A. Concentration and time dependency of artemisinin efficacy against Plasmodium falciparum in vitro. Am. J. Trop. Med. Hyg. 1994, 50, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.; Assreuy, J. Role of nitric oxide and superoxide in Giardia lamblia killing. Braz. J. Med. Biol. Res. 1997, 30, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Ringqvist, E.; Palm, J.E.; Skarin, H.; Hehl, A.B.; Weiland, M.; Davids, B.J.; Reiner, D.S.; Griffiths, W.J.; Eckmann, L.; Gillin, F.D.; et al. Release of metabolic enzymes by Giardia in response to interaction with intestinal epithelial cells. Mol. Biochem. Parasitol. 2008, 159, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Ding, J.; Yang, C.; Gao, Y.; Li, X.; Chen, X.; Peng, Y.; Fang, J.; Xiao, S. Immunomodulatory and anti-inflammatory properties of artesunate in experimental colitis. Curr. Med. Chem. 2012, 19, 4541–4551. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Zhou, P.; Singer, S.M. Neuronal nitric oxide synthase is necessary for elimination of Giardia lamblia infections in mice. J. Immunol. 2006, 176, 516–521. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Li, E.; Shea-Donohue, T.; Singer, S.M. Tumour necrosis factor alpha contributes to protection against Giardia lamblia infection in mice. Parasite Immunol. 2007, 29, 367–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matowicka-Karna, J.; Dymicka-Piekarska, V.; Kemona, H. IFN-gamma, IL-5, IL-6 and IgE in patients infected with Giardia intestinalis. Folia Histochem. Cytobiol. 2009, 47, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, X.; Gong, P.; Xia, F.; Li, L.; Yang, Z.; Li, J. TLR2(-/-) Mice Display Decreased Severity of Giardiasis via Enhanced Proinflammatory Cytokines Production Dependent on AKT Signal Pathway. Front. Immunol. 2017, 8, 1186. [Google Scholar] [CrossRef] [Green Version]
- Solaymani-Mohammadi, S.; Singer, S.M. Host immunity and pathogen strain contribute to intestinal disaccharidase impairment following gut infection. J. Immunol. 2011, 187, 3769–3775. [Google Scholar] [CrossRef]
- Kuhn, K.A.; Manieri, N.A.; Liu, T.-C.; Stappenbeck, T.S. IL-6 stimulates intestinal epithelial proliferation and repair after injury. PLoS ONE 2014, 9, e114195. [Google Scholar] [CrossRef] [Green Version]
- Zong, X.; Hu, W.; Song, D.; Li, Z.; Du, H.; Lu, Z.; Wang, Y. Porcine lactoferrin-derived peptide LFP-20 protects intestinal barrier by maintaining tight junction complex and modulating inflammatory response. Biochem. Pharmacol. 2016, 104, 74–82. [Google Scholar] [CrossRef]
- Cabrera-Licona, A.; Solano-Gonzalez, E.; Fonseca-Linan, R.; Bazan-Tejeda, M.L.; Raul, A.-G.; Bermudez-Cruz, R.M.; Ortega-Pierres, G. Expression and secretion of the Giardia duodenalis variant surface protein 9B10A by transfected trophozoites causes damage to epithelial cell monolayers mediated by protease activity. Exp. Parasitol. 2017, 179, 49–64. [Google Scholar] [CrossRef]
- Scott, K.G.; Yu, L.C.; Buret, A.G. Role of CD8+ and CD4+ T lymphocytes in jejunal mucosal injury during murine giardiasis. Infect. Immun. 2004, 72, 3536–3542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youakim, A.; Ahdieh, M. Interferon-gamma decreases barrier function in T84 cells by reducing ZO-1 levels and disrupting apical actin. Am. J. Physiol. 1999, 276, G1279–G1288. [Google Scholar] [CrossRef]
- Troeger, H.; Epple, H.J.; Schneider, T.; Wahnschaffe, U.; Ullrich, R.; Burchard, G.D.; Jelinek, T.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Effect of chronic Giardia lamblia infection on epithelial transport and barrier function in human duodenum. Gut 2007, 56, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.A.; Fisher, B.S.; Estran, C.E. Modeling Long-Term Host Cell-Giardia lamblia Interactions in an In Vitro Co-Culture System. PLoS ONE 2013, 8, e81104. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.M.; Duckworth, C.A.; Watson, A.J.; Frey, M.R.; Miguel, J.C.; Burkitt, M.D.; Sutton, R.; Hughes, K.R.; Hall, L.J.; Caamano, J.H.; et al. A mouse model of pathological small intestinal epithelial cell apoptosis and shedding induced by systemic administration of lipopolysaccharide. Dis. Model. Mech. 2013, 6, 1388–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piguet, P.F.; Vesin, C.; Donati, Y.; Barazzone, C. TNF-induced enterocyte apoptosis and detachment in mice: Induction of caspases and prevention by a caspase inhibitor, ZVAD-fmk. Lab. Investig. 1999, 79, 495–500. [Google Scholar] [PubMed]
- Parker, A.; Vaux, L.; Patterson, A.M.; Modasia, A.; Muraro, D.; Fletcher, A.G.; Byrne, H.M.; Maini, P.K.; Watson, A.J.M.; Pin, C. Elevated apoptosis impairs epithelial cell turnover and shortens villi in TNF-driven intestinal inflammation. Cell Death Dis. 2019, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Han, X.; Wu, S.; Wu, J.; Yang, C.; Li, X. Unexpected mechanism of colitis amelioration by artesunate, a natural product from Artemisia annua L. Inflammopharmacology 2020, 28, 851–868. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.E.M.; Hassanein, E.H.M.; Bakr, A.G.; El-Shoura, E.A.M.; El-Gamal, D.A.; Mahmoud, A.R.; Abd-Elhamid, T.H. Ursodeoxycholic acid abrogates gentamicin-induced hepatotoxicity in rats: Role of NF-kappaB-p65/TNF-alpha, Bax/Bcl-xl/Caspase-3, and eNOS/iNOS pathways. Life Sci. 2020, 254, 117760. [Google Scholar] [CrossRef]
- Abd-Elhamid, T.H.; Elgamal, D.A.; Ali, S.S.; Ali, F.E.M.; Hassanein, E.H.M.; El-Shoura, E.A.M.; Hemeida, R.A.M. Reno-protective effects of ursodeoxycholic acid against gentamicin-induced nephrotoxicity through modulation of NF-kappaB, eNOS and caspase-3 expressions. Cell Tissue Res. 2018, 374, 367–387. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.J.; Schmiech, M.; Hafner, S.; Paetz, C.; Steinborn, C.; Huber, R.; Gaafary, M.E.; Werner, K.; Schmidt, C.Q.; Syrovets, T.; et al. Antitumor activity of an Artemisia annua herbal preparation and identification of active ingredients. Phytomedicine 2019, 62, 152962. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.-D.; Jin, G.-Z.; Piao, G.-C. Protective Effects of the Supernatant of Ethanol Eluate from Artemisia sacrorum Ledeb. against Acetaminophen-Induced Liver Injury in Mice [corrected]. Biol. Pharm. Bull. 2009, 32, 1683–1688. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.R.; Liu, Z.X.; Liu, F.; Pan, L.; Yu, H.P.; Jiang, J.P.; Zhang, J.J.; Liu, L.; Yu, J. Holotransferrin enhances selective anticancer activity of artemisinin against human hepatocellular carcinoma cells. J. Huazhong Univ. Sci. Technol. Med. Sci. 2013, 33, 862–865. [Google Scholar] [CrossRef]
- Katz, D.; Taylor, D. Parasitic infections of the gastrointestinal tract. Gastroenterol. Clin. N. Am. 2001, 30, 797–815. [Google Scholar] [CrossRef]
- Ebert, E.C. Giardia induces proliferation and interferon gamma production by intestinal lymphocytes. Gut 1999, 44, 342–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deplancke, B.; Gaskins, H.R. Microbial modulation of innate defense: Goblet cells and the intestinal mucus layer. Am. J. Clin. Nutr. 2001, 73, 1131S–1141S. [Google Scholar] [CrossRef] [Green Version]
- Birchenough, G.M.; Johansson, M.E.; Gustafsson, J.K.; Bergstrom, J.H.; Hansson, G.C. New developments in goblet cell mucus secretion and function. Mucosal. Immunol. 2015, 8, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Amat, C.B.; Motta, J.P.; Fekete, E.; Moreau, F.; Chadee, K.; Buret, A.G. Cysteine Protease-Dependent Mucous Disruptions and Differential Mucin Gene Expression in Giardia duodenalis Infection. Am. J. Pathol. 2017, 187, 2486–2498. [Google Scholar] [CrossRef] [Green Version]
- Shukla, G.; Sidhu, R.K. Lactobacillus casei as a probiotic in malnourished Giardia lamblia-infected mice: A biochemical and histopathological study. Can. J. Microbiol. 2011, 57, 127–135. [Google Scholar] [CrossRef]
- Roskens, H.; Erlandsen, S.L. Inhibition of in vitro attachment of Giardia trophozoites by mucin. J. Parasitol. 2002, 88, 869–873. [Google Scholar] [CrossRef]
- Deng, Y.; Liu, Z.; Geng, Y. Anti-allergic effect of Artemisia extract in rats. Exp. Ther. Med. 2016, 12, 1130–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squire, S.A.; Ryan, U. Cryptosporidium and Giardia in Africa: Current and future challenges. Parasites Vectors 2017, 10, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilia, A.R.; Santomauro, F.; Sacco, C.; Bergonzi, M.C.; Donato, R. Essential Oil of Artemisia annua L.: An Extraordinary Component with Numerous Antimicrobial Properties. Evid.-Based Complement Altern. Med. 2014, 2014, 159819. [Google Scholar] [CrossRef] [Green Version]
- Elfawal, M.A.; Towler, M.J.; Reich, N.G.; Golenbock, D.; Weathers, P.J.; Rich, S.M. Dried whole plant Artemisia annua as an antimalarial therapy. PLoS ONE 2012, 7, e52746. [Google Scholar] [CrossRef] [PubMed]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd-Elhamid, T.H.; Abdel-Rahman, I.A.M.; Mahmoud, A.R.; Allemailem, K.S.; Almatroudi, A.; Fouad, S.S.; Abdella, O.H.; Elshabrawy, H.A.; El-Kady, A.M. A Complementary Herbal Product for Controlling Giardiasis. Antibiotics 2021, 10, 477. https://doi.org/10.3390/antibiotics10050477
Abd-Elhamid TH, Abdel-Rahman IAM, Mahmoud AR, Allemailem KS, Almatroudi A, Fouad SS, Abdella OH, Elshabrawy HA, El-Kady AM. A Complementary Herbal Product for Controlling Giardiasis. Antibiotics. 2021; 10(5):477. https://doi.org/10.3390/antibiotics10050477
Chicago/Turabian StyleAbd-Elhamid, Tarek Hamdy, Iman A. M. Abdel-Rahman, Amany Refaat Mahmoud, Khaled S. Allemailem, Ahmad Almatroudi, Samer S. Fouad, Osama H. Abdella, Hatem A. Elshabrawy, and Asmaa M. El-Kady. 2021. "A Complementary Herbal Product for Controlling Giardiasis" Antibiotics 10, no. 5: 477. https://doi.org/10.3390/antibiotics10050477
APA StyleAbd-Elhamid, T. H., Abdel-Rahman, I. A. M., Mahmoud, A. R., Allemailem, K. S., Almatroudi, A., Fouad, S. S., Abdella, O. H., Elshabrawy, H. A., & El-Kady, A. M. (2021). A Complementary Herbal Product for Controlling Giardiasis. Antibiotics, 10(5), 477. https://doi.org/10.3390/antibiotics10050477