
A C-Type Lectin Highly Expressed in Portunus trituberculatus Intestine Functions in AMP Regulation and Prophenoloxidase Activation

Abstract
:1. Introduction
2. Results
2.1. Cloning and Characterization of PtCLec2 cDNA
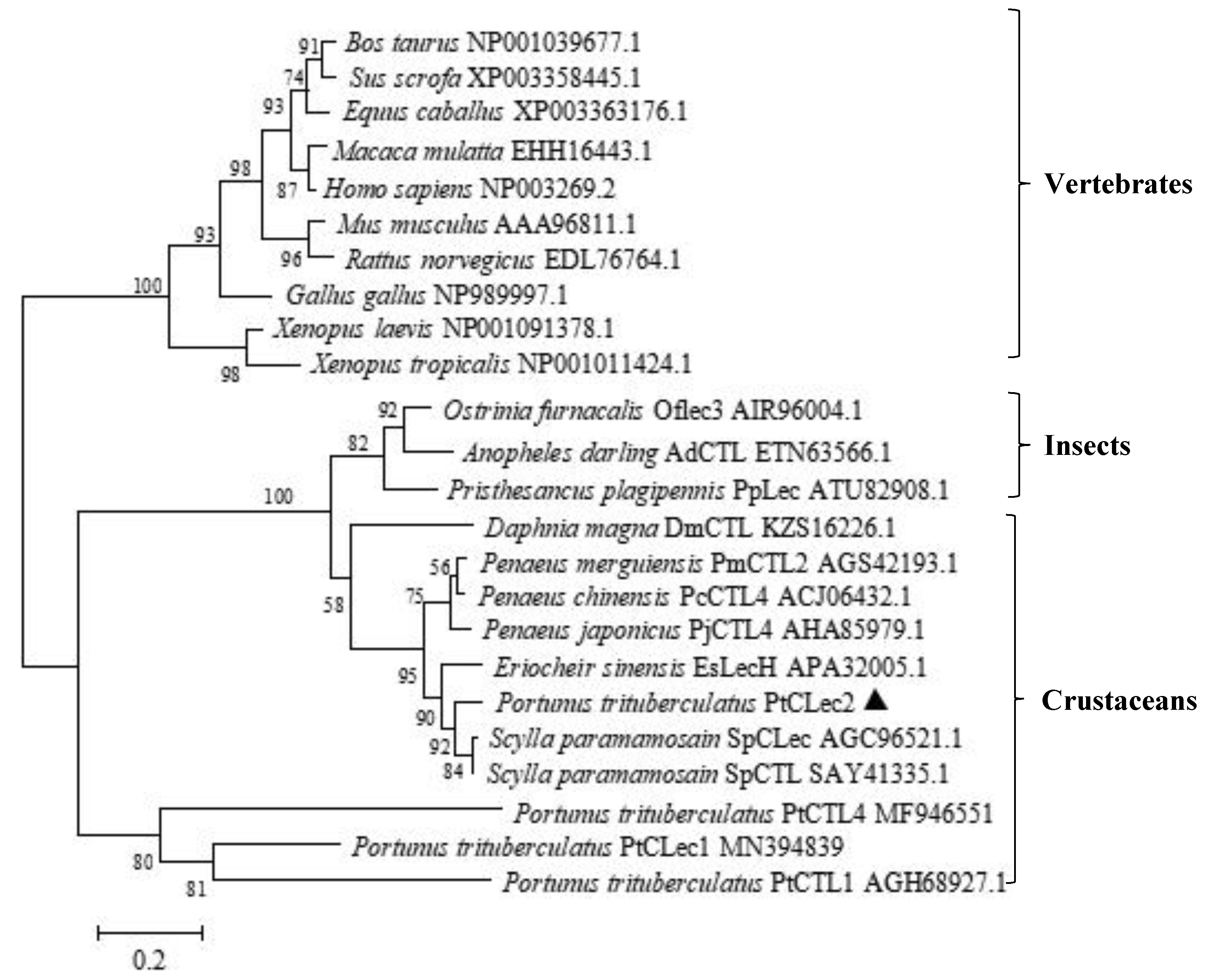
2.2. Multiple Alignment and Phylogenetic Analysis of PtCLec2
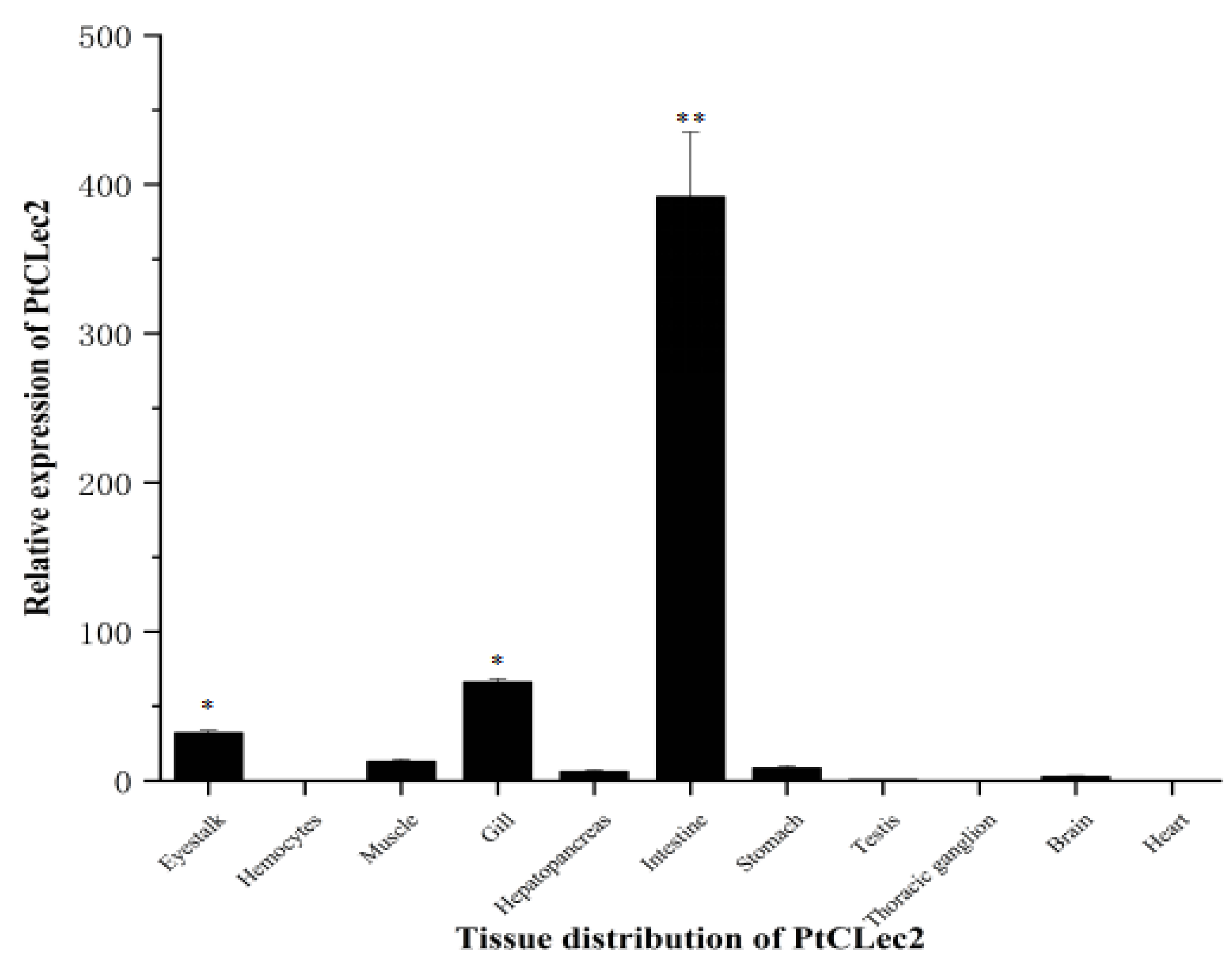
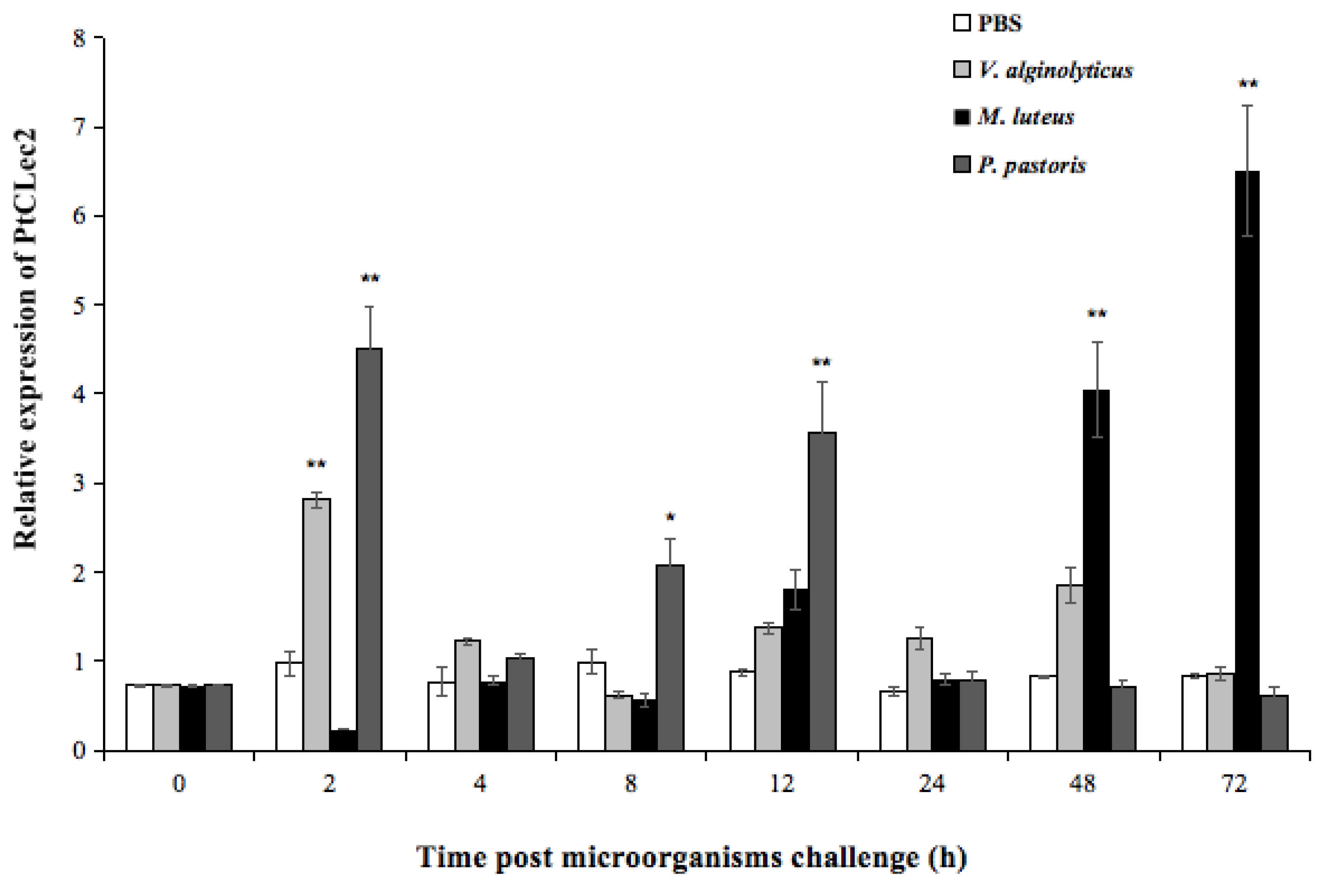
2.3. Quantitative Analysis of PtCLec2 Transcripts
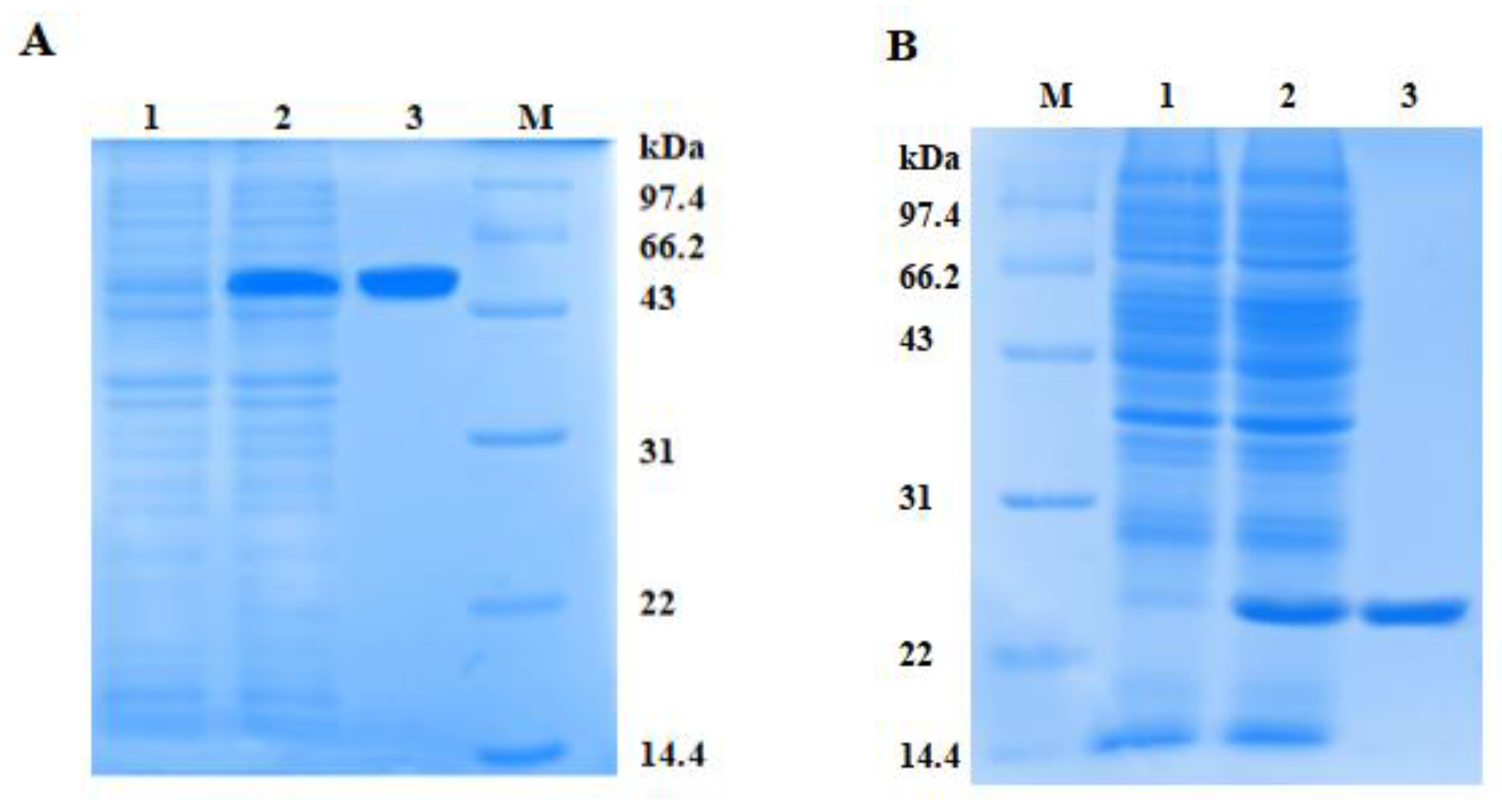
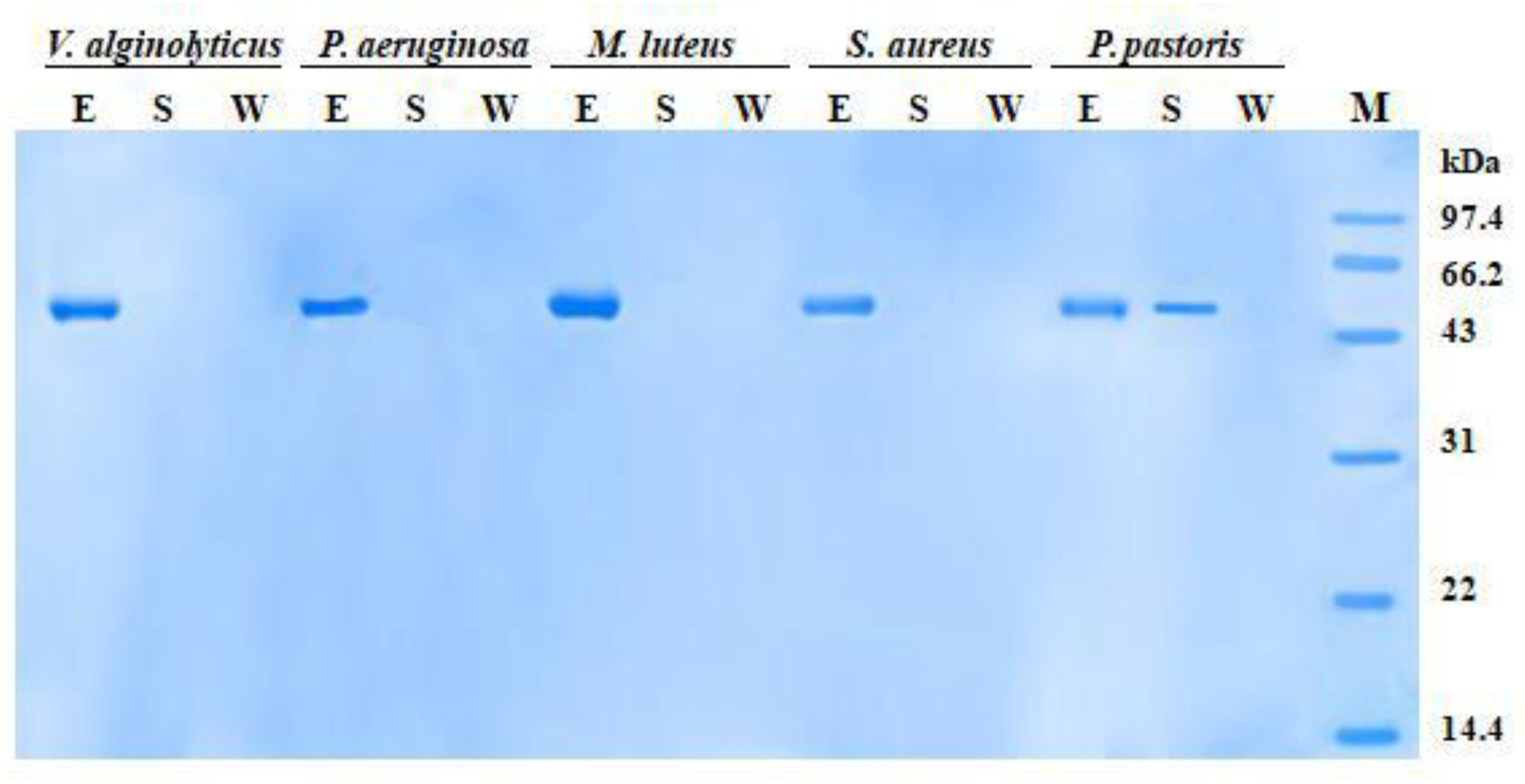
2.4. Expression and Purification of rPtCLec2
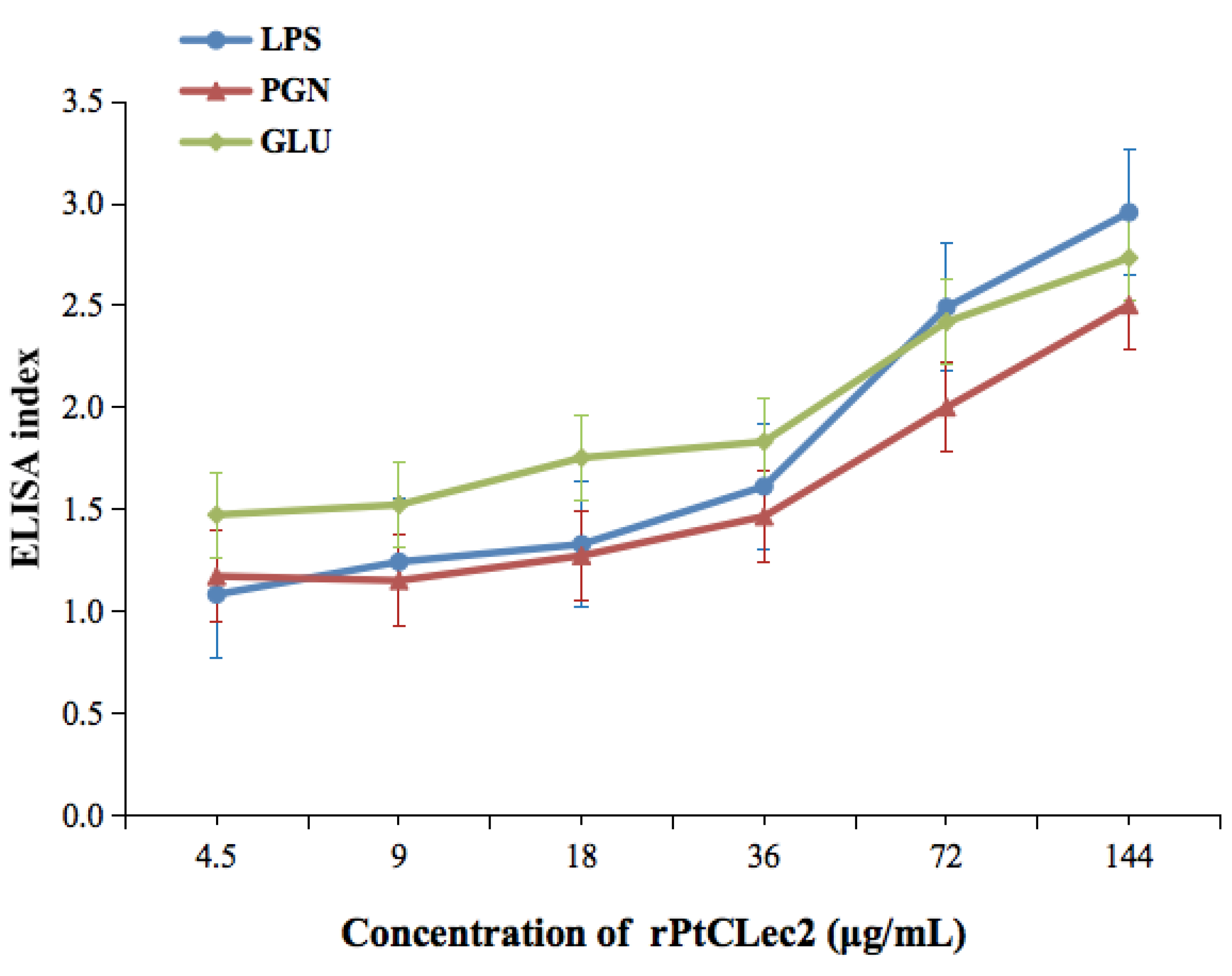
2.5. PAMP-Binding Activity of rPtCLec2
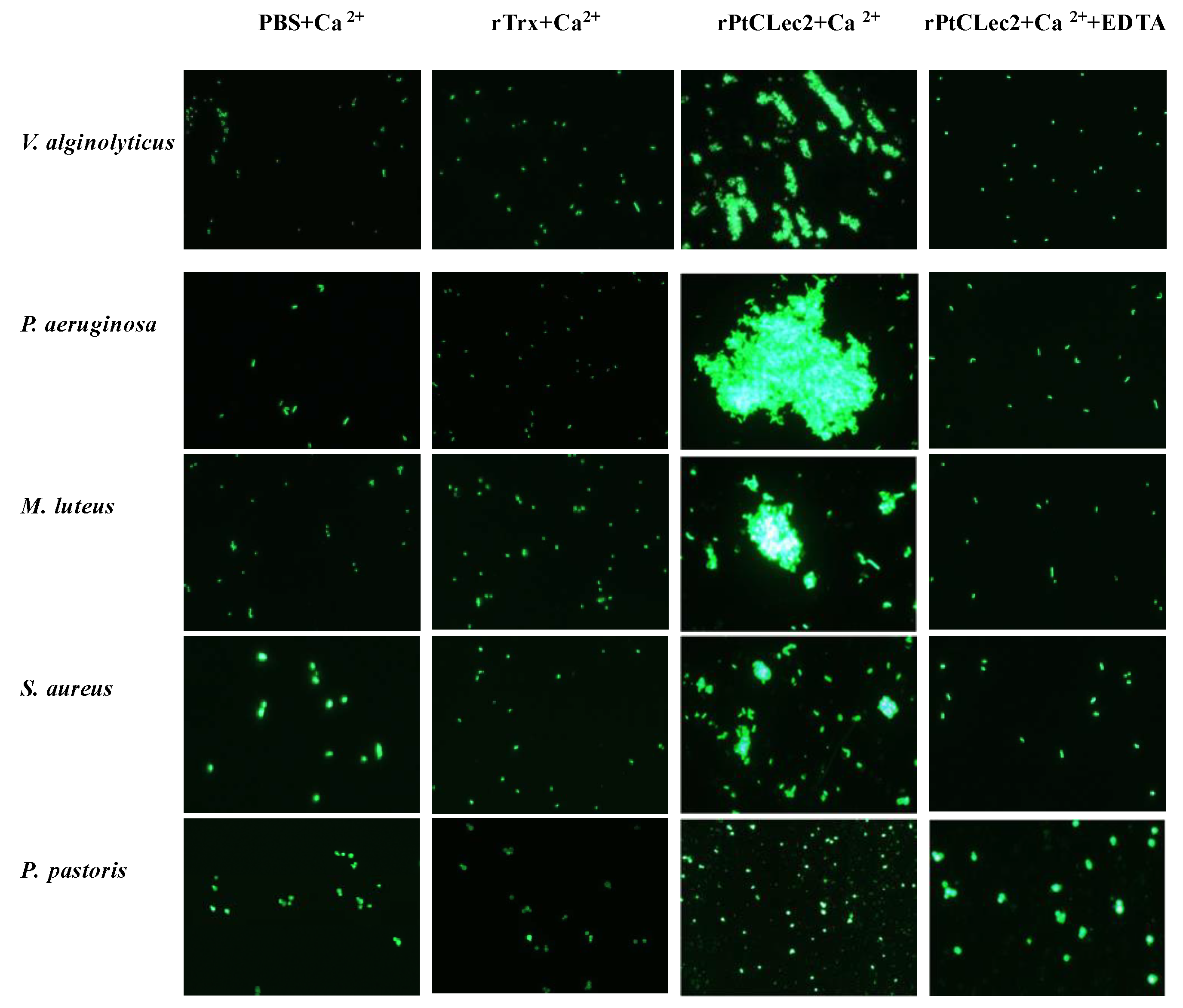
2.6. Microorganism-Binding Activity of rPtCLec2
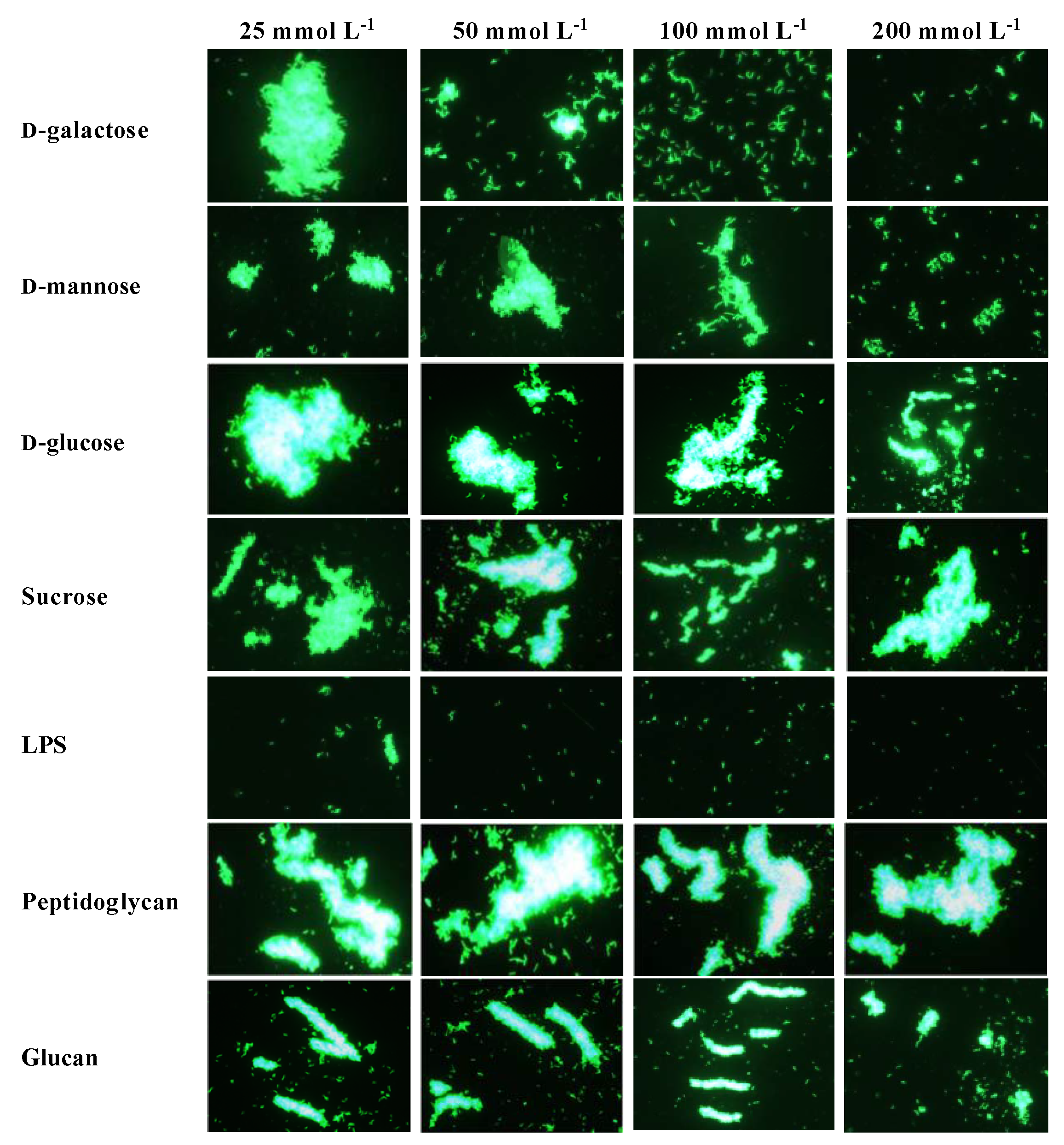
2.7. Agglutinating Activity of rPtCLec2 to Microorganisms and Erythrocytes
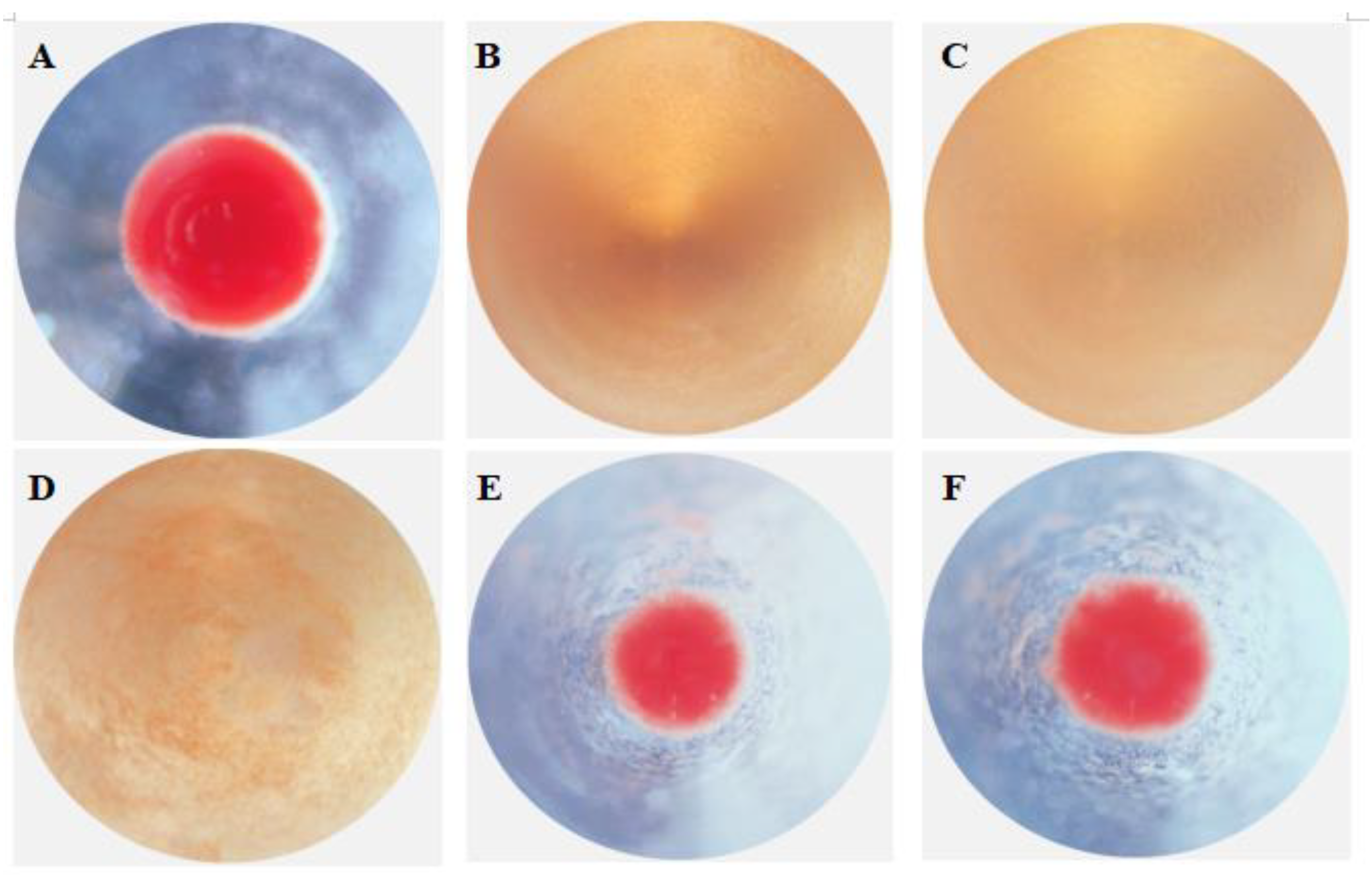
2.8. Antibacterial Activity of rPtCLec2
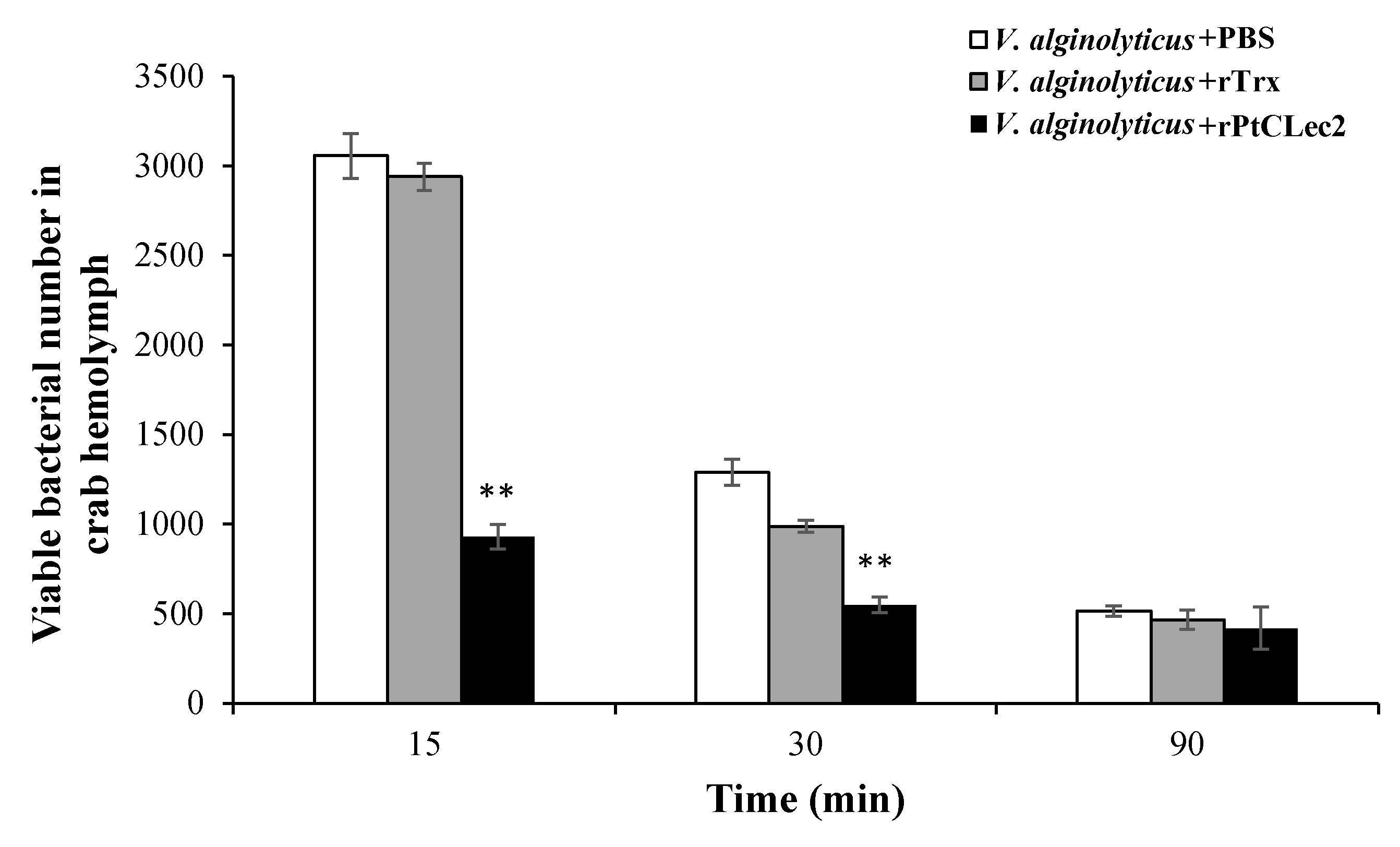
2.9. In Vivo V. alginolyticus Clearance Activity of rPtCLec2
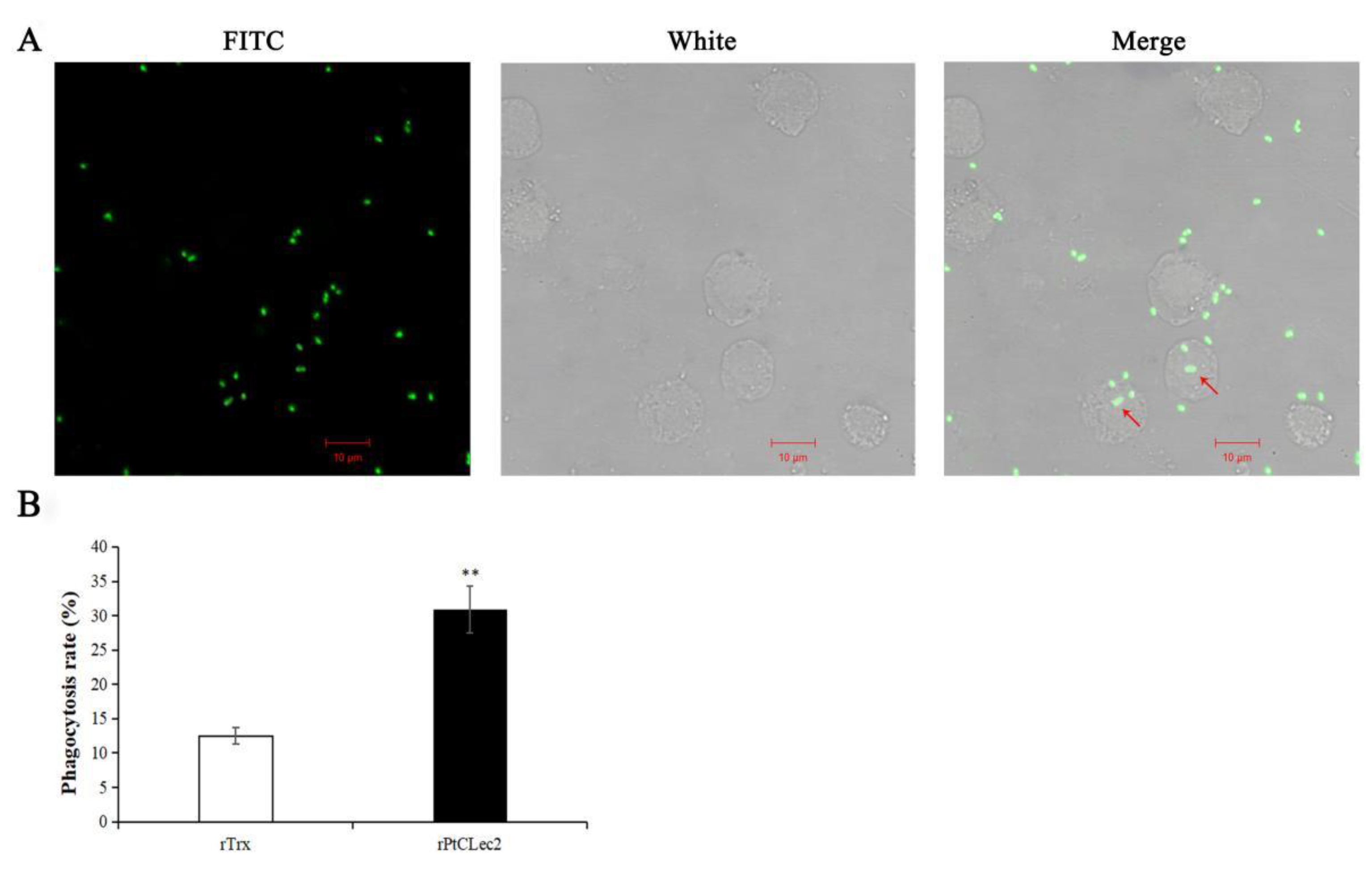
2.10. Phagocytic Activity of rPtCLec2
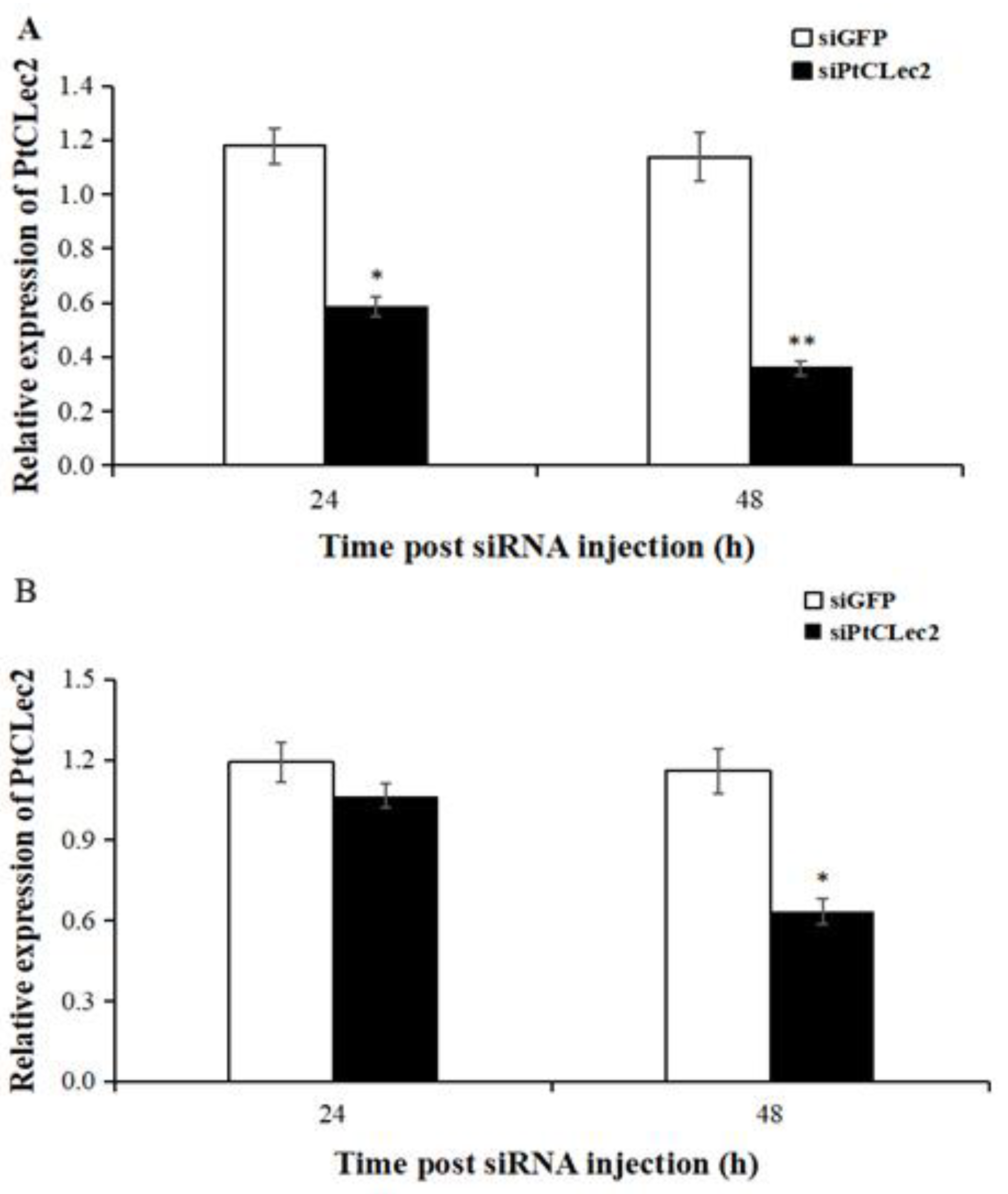
2.11. Gene Knockdown of PtCLec2
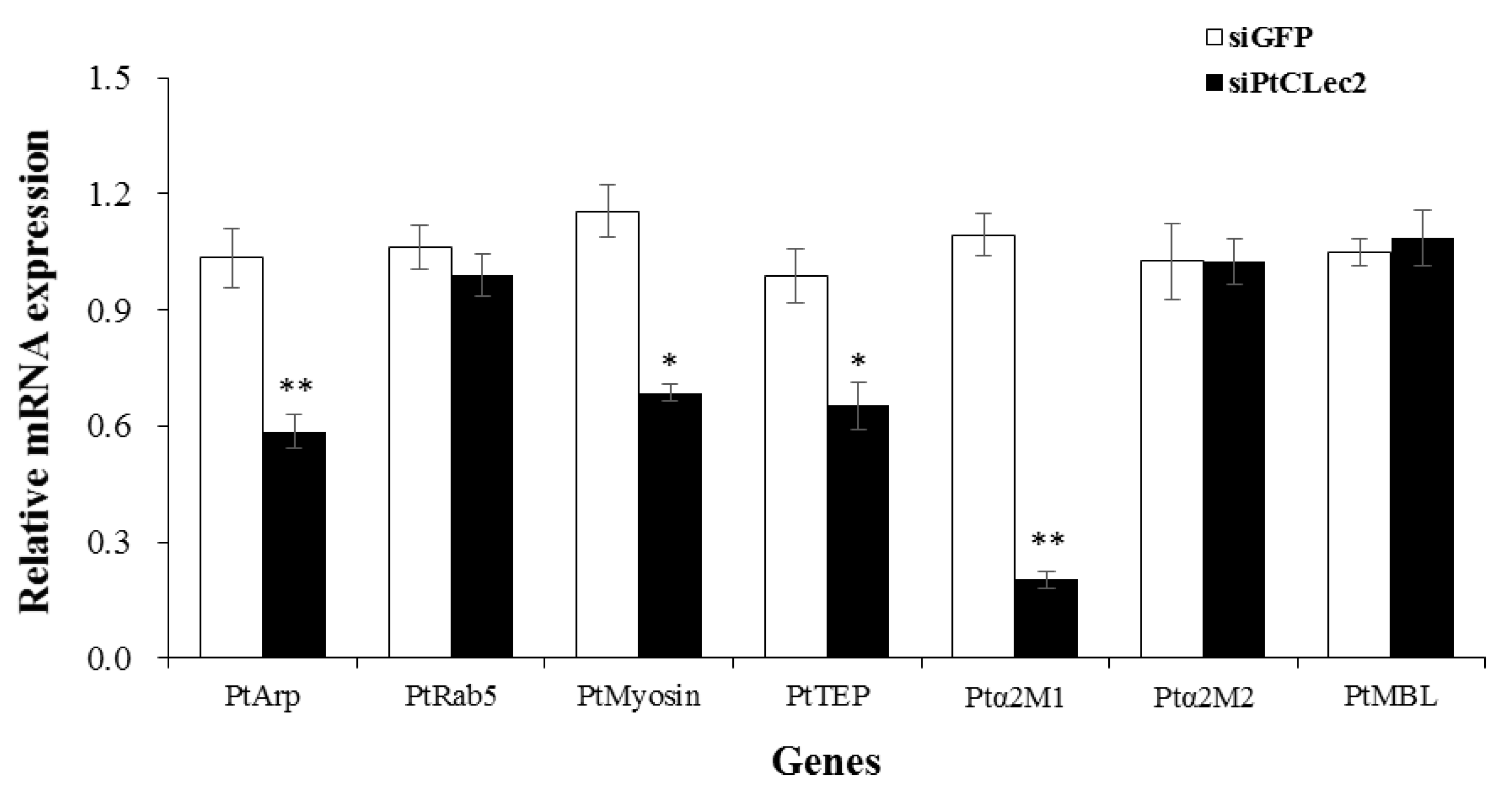
2.12. PtCLec2 Regulates the Expression of Genes Involved in Phagocytosis and Complement System
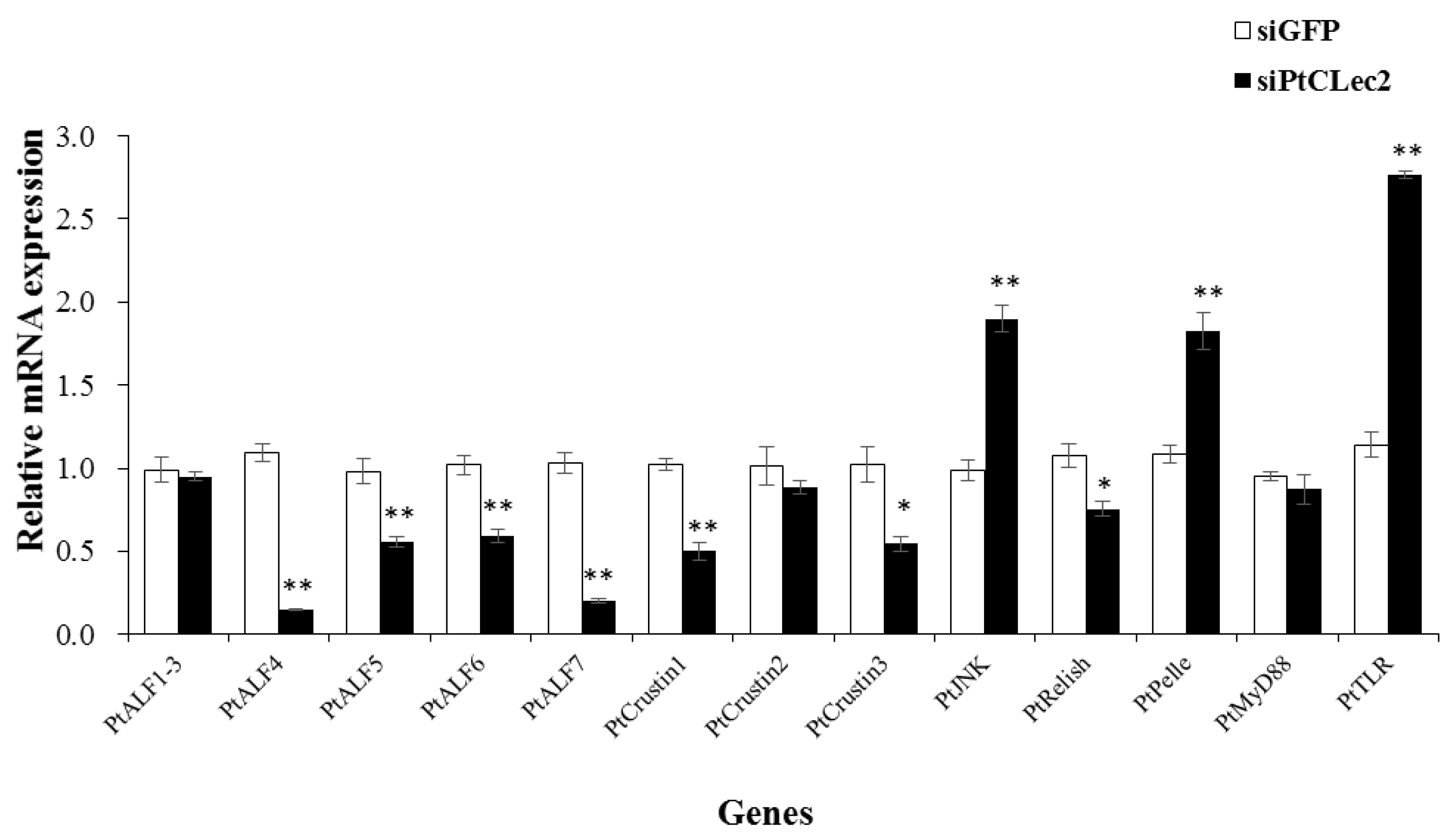
2.13. PtCLec2 May Regulate AMP Expression via Immune Pathways
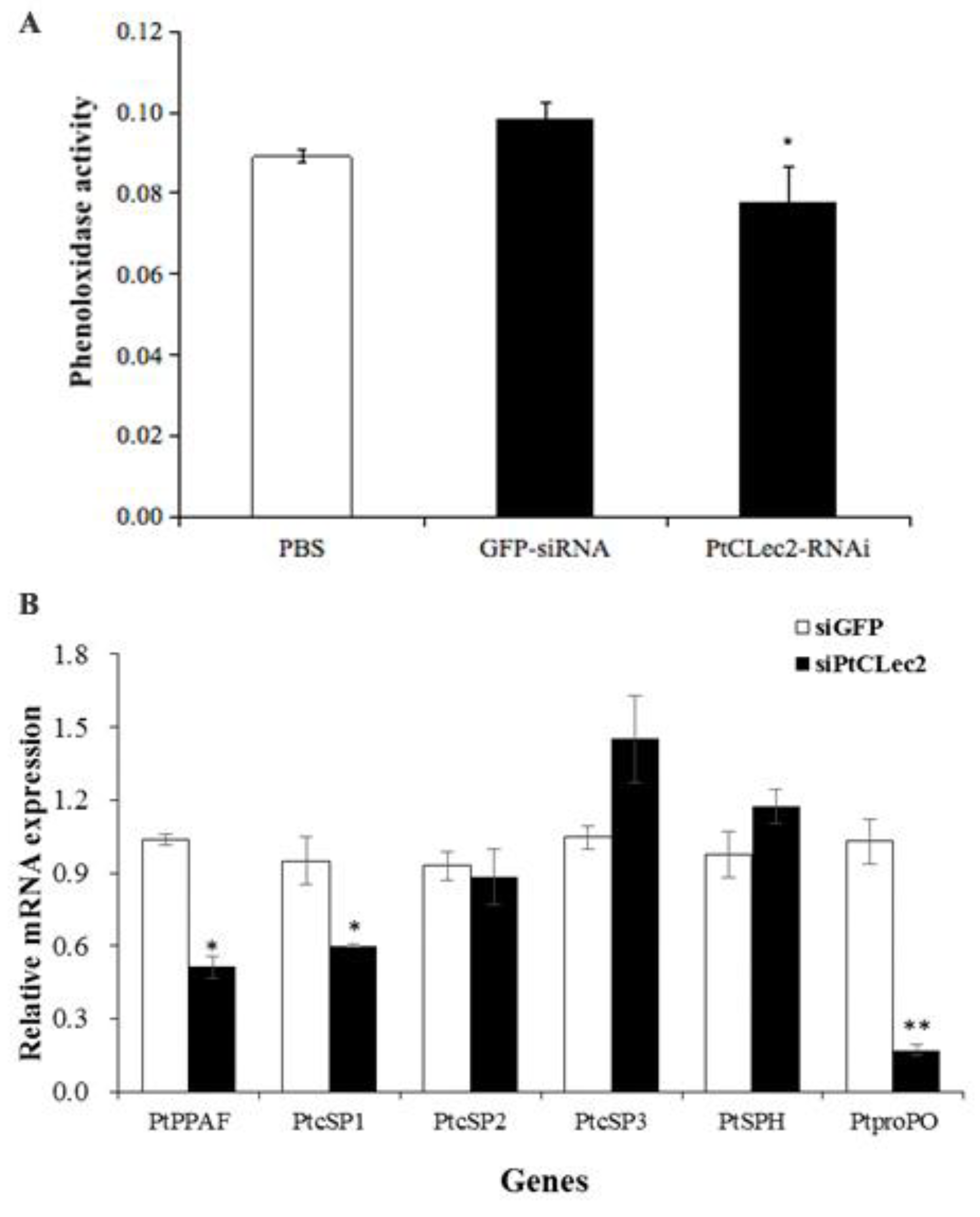
2.14. Effect of PtCLec2 Interference on proPO System
3. Discussion
4. Materials and Methods
4.1. Gene Cloning and Bioinformatic Analysis
4.2. Experimental Animals, Tissue Collection, and Immune Challenge
4.3. The Temporal mRNA Expression of PtCLec2
4.4. Expression and Purification of Recombinant PtCLec2
4.5. PAMP-Binding Assay
4.6. Microbial-Binding Assay
4.7. Microbial Agglutination and Inhibition Assay
4.8. Hemagglutination Assay
4.9. Antimicrobial Activity
4.10. In Vivo Bacterial Clearance Assay
4.11. Phagocytosis Assay
4.12. RNA Interference Assay
4.13. Expression of Immune-Related Genes in the PtCLec2-Knockdown Crabs
4.14. Phenoloxidase (PO) Activity in the PtCLec2-Knockdown Crabs
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Janeway, C.A.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medzhitov, R.; Janeway, C.A. Decoding the patterns of self and nonself by the innate immune system. Science 2002, 296, 298–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelensky, A.N.; Gready, J.E. The C-type lectin-like domain superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; Song, C.; Ning, J.; Cui, Z. Characterization and functional analysis of a novel mannose-binding lectin from the swimming crab Portunus trituberculatus. Fish Shellfish Immunol. 2019, 89, 448–457. [Google Scholar] [CrossRef]
- Luo, T.; Yang, H.; Li, F.; Zhang, X.; Xu, X. Purification, characterization and cDNA cloning of a novel lipopolysaccharide-binding lectin from the shrimp Penaeus monodon. Dev. Comp. Immunol. 2006, 30, 607–617. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; Yin, Z.X.; Xu, X.P.; Weng, S.P.; Rao, X.Y. A novel C-type lectin from the shrimp Litopenaeus vannamei possesses anti-white spot syndrome virus activity. J. Virol. 2009, 83, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.W.; Wang, J.X. Diversity and multiple functions of lectins in shrimp immunity. Dev. Comp. Immunol. 2013, 39, 27–38. [Google Scholar] [CrossRef]
- Wang, L.; Huang, M.; Zhang, H.; Song, L. The immune role of C-type lectins in molluscs. Invertebr. Surviv. J. 2011, 8, 241–246. [Google Scholar]
- Wang, X.W.; Vasta, G.R.; Wang, J.X. The functional relevance of shrimp C-type lectins in host-pathogen interactions. Dev. Comp. Immunol. 2020, 109, 103708. [Google Scholar] [CrossRef] [PubMed]
- Gross, P.S.; Bartlett, T.C.; Browdy, C.L.; Chapman, R.W.; Warr, G.W. Immune gene discovery by expressed sequence tag analysis of hemocytes and hepatopancreas in the Pacific White Shrimp, Litopenaeus vannamei, and the Atlantic White Shrimp, L. setiferus. Dev. Comp. Immunol. 2001, 25, 565–577. [Google Scholar] [CrossRef]
- Hoving, J.C.; Wilson, G.J.; Brown, G.D. Signaling C-type lectin receptors, microbial recognition and immunity. Cell. Microbiol. 2014, 16, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.W.; Wang, Y.; Wang, X.W.; Wang, L.; Mu, Y.; Wang, J.X. A C-type lectin with an immunoglobulin-like domain promotes phagocytosis of hemocytes in crayfish Procambarus clarkii. Sci. Rep. 2016, 6, 29924. [Google Scholar] [CrossRef] [Green Version]
- Kwankaew, P.; Praparatana, R.; Runsaeng, P.; Utarabhand, P. An alternative function of C-type lectin comprising low-density lipoprotein receptor domain from Fenneropenaeus merguiensis to act as a binding receptor for viral protein and vitellogenin. Fish and Shellfish Immunol. 2018, 74, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Yang, L.; Wang, Z.; Zuo, H.; Weng, S.; He, J.; Xu, X. A novel C-type lectin with microbiostatic and immune regulatory functions from Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 93, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.J.; Lan, J.F.; Zhao, X.F.; Vasta, G.R.; Wang, J.X. Binding of a C-type lectin’s coiled-coil domain to the Domeless receptor directly activates the JAK/STAT pathway in the shrimp immune response to bacterial infection. PLoS Pathog. 2017, 13, e1006626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Z.; Li, D.; Li, X.; Zhang, X.; Zhu, Y.; Li, W.; Wang, Q. A single CRD C-type lectin from Eriocheir sinensis (EsLecB) with microbial-binding, antibacterial prophenoloxidase activation and hem-encapsulation activities. Fish Shellfish Immunol. 2016, 50, 175–190. [Google Scholar] [CrossRef]
- Wang, X.W.; Zhang, H.W.; Li, X.; Zhao, X.F.; Wang, J.X. Characterization of a C-type lectin (PcLec2) as an upstream detector in the prophenoloxidase activating system of red swamp crayfish. Fish Shellfish Immunol. 2011, 30, 241–247. [Google Scholar] [CrossRef]
- Kong, H.J.; Park, E.M.; Nam, B.H.; Kim, Y.O.; Kim, W.J.; Park, H.J. A C-type lectin like-domain (CTLD)-containing protein (PtLP) from the swimming crab Portunus trituberculatus. Fish Shellfish Immunol. 2008, 25, 311–314. [Google Scholar] [CrossRef]
- Lu, J.; Yu, Z.; Mu, C.; Li, R.; Song, W.; Wang, C. Characterization and functional analysis of a novel C-type lectin from the swimming crab Portunus trituberculatus. Fish Shellfish Immunol. 2017, 64, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Mu, C.; Wu, Y.; Ye, F.; Wang, D.; Sun, C.; Lv, Z.; Han, B.; Wang, C.; Xu, X.W. The functional characterization and comparison of two single CRD containing C-type lectins with novel and typical key motifs from Portunus trituberculatus. Fish Shellfish Immunol. 2017, 70, 398–407. [Google Scholar] [CrossRef]
- Zhang, X.; Lu, J.; Mu, C.; Li, R.; Song, W.; Ye, Y.; Shi, C.; Liu, L.; Wang, C. Molecular cloning of a C-type lectin from Portunus trituberculatus, which might be involved in the innate immune response. Fish Shellfish Immunol. 2018, 76, 216–223. [Google Scholar] [CrossRef]
- Wang, X.W.; Zhang, X.W.; Xu, W.T.; Zhao, X.F.; Wang, J.X. A novel C-type lectin (FcLec4) facilitates the clearance of Vibrio anguillarum in vivo in Chinese white shrimp. Dev. Comp. Immunol. 2009, 33, 1039–1047. [Google Scholar] [CrossRef]
- Utarabhand, P.; Thepnarong, S.; Runsaeng, P. Lipopolysaccharide-specific binding C-type lectin with one CRD domain from Fenneropenaeus merguiensis (FmLC4) functions as a pattern recognition receptor in shrimp innate immunity. Fish Shellfish Immunol. 2017, 69, 236–246. [Google Scholar] [CrossRef]
- Qin, Y.; Jiang, S.; Huang, J.; Zhou, F.; Yang, Q.; Jiang, S.; Yang, L. C-type lectin response to bacterial infection and ammonia nitrogen stress in tiger shrimp (Penaeus monodon). Fish Shellfish Immunol. 2019, 90, 188–198. [Google Scholar] [CrossRef]
- Feng, J.; Huang, X.; Jin, M.; Zhang, Y.; Li, T.; Hui, K.; Ren, Q. A C-type lectin (MrLec) with high expression in intestine is involved in innate immune response of Macrobrachium rosenbergii. Fish Shellfish Immunol. 2016, 59, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Engel, P.; Moran, N.A. The gut microbiota of insects diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef]
- Song, X.; Xin, X.; Wang, H.; Li, H.; Zhang, H.; Jia, Z.; Liu, C.; Jiang, S.; Wang, L.; Song, L. A single-CRD C-type lectin (CgCLec-3) with novel DIN motif exhibits versatile immune functions in Crassostrea gigas. Fish Shellfish Immunol. 2019, 92, 772–781. [Google Scholar] [CrossRef]
- Zhang, X.W.; Man, X.; Huang, X.; Wang, Y.; Song, Q.S.; Hui, K.M.; Zhang, H.W. Identification of a C-type lectin possessing both antibacterial and antiviral activities from red swamp crayfish. Fish Shellfish Immunol. 2018, 77, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Zhang, D.; Li, F.; Wang, M.; Huang, M.; Song, L. A novel C-type lectin from crab Eriocheir sinensis functions as pattern recognition receptor enhancing cellular encapsulation. Fish Shellfish Immunol. 2013, 34, 832–842. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, L.; Huang, M.M.; Song, L. A galectin from Eriocheir sinensis functions as pattern recognition receptor enhancing microbe agglutination and haemocytes encapsulation. Fish Shellfish Immunol. 2016, 55, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Lis, H.; Sharon, N. Lectins: carbohydrate-specific proteins that mediate cellular recognition. Chem. Rev. 1998, 98, 637–674. [Google Scholar] [CrossRef]
- Huang, M.; Wang, L.; Yang, J.; Zhang, H.; Wang, L.; Song, L. A four-CRD C-type lectin from Chlamys farreri mediating nonself-recognition with broader spectrum and opsonization. Dev. Comp. Immunol. 2013, 39, 363–369. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, H.; Jiang, S.; Wang, L.; Liu, R.; Yi, Q.; Song, L. An EPD/WSD motifs containing C-type lectin from Argopectens irradians recognizes and binds microbes with broad spectrum. Fish Shellfish Immunol. 2015, 43, 287–293. [Google Scholar] [CrossRef]
- Tian, Y.Y.; Liu, Y.; Zhao, X.F.; Wang, J.X. Characterization of a C-type lectin from the cotton bollworm, Helicoverpa armigera. Dev. Comp. Immunol. 2009, 33, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.K.; Guo, X.N.; Li, S.; Wu, M.H.; Zhu, Y.T.; Yu, A.Q. Association of a hepatopancreas-specific C-type lectin with the antibacterial response of Eriocheir sinensis. PLoS ONE 2013, 8, e76132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runsaeng, P.; Puengyam, P.; Utarabhand, P. A mannose-specific C-type lectin from Fenneropenaeus merguiensis exhibited antimicrobial activity to mediate shrimp innate immunity. Mol. Immunol. 2017, 92, 87–98. [Google Scholar] [CrossRef]
- Zhu, Y.T.; Zhang, X.; Wang, S.C.; Li, W.W.; Wang, Q. Antimicrobial functions of EsLecH, a C-type lectin, via JNK pathway in the Chinese mitten crab, Eriocheir sinensis. Dev. Comp. Immunol. 2016, 61, 225–235. [Google Scholar] [CrossRef]
- Wongpanya, R.; Sengprasert, P.; Amparyup, P.; Tassanakajon, A. A novel C-type lectin in the black tiger shrimp Penaeus monodon functions as a pattern recognition receptor by binding and causing bacterial agglutination. Fish Shellfish Immunol. 2017, 60, 103–113. [Google Scholar] [CrossRef]
- Zhang, X.W.; Xu, W.T.; Wang, X.W.; Mu, Y.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A novel C-type lectin with two CRD domains from Chinese shrimp Fenneropenaeus chinensis functions as a pattern recognition protein. Mol. Immunol. 2009, 46, 1626–1637. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Liu, X.; Yang, J.; Wang, S.; Sun, G.; Yang, J. Critical roles of sea cucumber C-type lectin in non-self recognition and bacterial clearance. Fish Shellfish Immunol. 2015, 45, 791–799. [Google Scholar] [CrossRef]
- Zhang, H.; Song, X.; Wang, L.; Kong, P.; Yang, J.; Liu, L.; Qiu, L.; Zhang, Y.; Qiu, L.; Song, L. AiCTL-6, a novel C-type lectin from bay scallop Argopecten irradians with a long C-type lectin-like domain. Fish Shellfish Immunol. 2011, 30, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Song, X.; Zhao, J.; Mu, C.; Wang, L.; Zhang, H.; Zhou, Z.; Liu, X.; Song, L. A C-type lectin (AiCTL-3) from bay scallop Argopecten irradians with mannose/galactose binding ability to bind various bacteria. Gene 2013, 531, 31–38. [Google Scholar] [CrossRef]
- Su, Y.; Liu, Y.; Gao, F.; Cui, Z. A novel C-type lectin with a YPD motif from Portunus trituberculatus (PtCLec1) mediating pathogen recognition and opsonization. Dev. Comp. Immunol. 2020, 106, 103609. [Google Scholar] [CrossRef] [PubMed]
- Phanthipha, R.; Pattamaporn, K.; Prapaporn, U. FmLC6: An ultimate dual-CRD C-type lectin from Fenneropenaeus merguiensis mediated its roles in shrimp defense immunity towards bacteria and virus. Fish Shellfish Immunol. 2018, 80, 200–213. [Google Scholar]
- Hoffmann, J.A.; Kafatos, F.C.; Janeway, C.A.; Ezekowitz, R.A.B. Phylogenetic perspectives in innate immunity. Science 1999, 284, 1313–1318. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Wang, L.; Sun, W.; Zhang, M.; Ma, H.; Zhang, Y.; Zhang, X.; Li, S. C-type lectin B (SpCTL-B) regulates the expression of antimicrobial peptides and promotes phagocytosis in mud crab Scylla paramamosain. Dev. Comp. Immunol. 2018, 84, 213–229. [Google Scholar] [CrossRef]
- Jia, Z.; Zhang, H.; Jiang, S.; Wang, M.; Wang, L.; Song, L. Comparative study of two single CRD C-type lectins, CgCLec-4 and CgCLec-5, from pacific oyster Crassostrea gigas. Fish Shellfish Immunol. 2016, 59, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Li, T.; Jin, M.; Yin, S.; Wang, W.; Ren, Q. Identification of a Macrobrachium nipponense C-type lectin with a close evolutionary relationship to vertebrate lectins. Mol. Immunol. 2017, 87, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Ponprateep, S.; Vatanavicharn, T.; Chu, F.L.; Tassanakajon, A.; Rimphanitchayakit, V. Alpha-2-macroglobulin is a modulator of prophenoloxidase system in pacific white shrimp Litopenaeus vannamai. Fish Shellfish Immunol. 2016, 62, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. Disulfide-bond reshuffling in the evolution of an ape placental ribonuclease. Mol. Biol. Evol. 2007, 24, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2-ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Söderhäll, K. Characterization of a pattern recognition protein, a masquerade-like protein, in the freshwater crayfish Pacifastacus leniusculus. J. Immunol. 2001, 166, 7319–7326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, C.; Song, X.; Zhao, J.; Wang, L.; Qiu, L.; Zhang, H. A scallop C-type lectin from Argopecten irradians (AiCTL5) with activities of lipopolysaccharide binding and Gram-negative bacteria agglutination. Fish Shellfish Immunol. 2012, 32, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Rathinakumar, R.; Walkenhorst, W.F.; WimLey, W.C. Broad-spectrum antimicrobial peptides by rational combinatorial design and high-throughput screening: the importance of interfacial activity. J. Am. Chem. Soc. 2009, 131, 7609–7617. [Google Scholar] [PubMed] [Green Version]
- Liu, H.; Liu, Y.; Cui, Z. Functional characterization of two clip-domain serine proteases in the swimming crab Portunus trituberculatus. Fish Shellfish Immunol. 2019, 89, 98–107. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | MIC (μM) |
|---|---|
| Gram-negative bacteria | |
| Vibrio alginolyticus | 0.86–1.74 |
| Pseudomonas aeruginosa | 1.74–3.50 |
| Gram-positive bacteria | |
| Micrococcus luteus | 0.43–0.86 |
| Staphylococcus aureus | 0.86–1.74 |
| Fungus | |
| Pichia pastoris | Na |
| Primer Name | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| cDNA cloning | ||
| PtCLec2 | CCGAGAATGCGAATCTACTTGC | TGTTTACTGTGAGCAGCAGCCAC |
| RACE | ||
| PtCLec2-3P1 | CGACCAACGCATCGCCACCAG | |
| PtCLec2-3P2 | CGACATTGCCTGCTACCACAAGAAG | |
| PtCLec2-5P1 | AGCCTGGTGGCGATGCGTTGG | |
| PtCLec2-5P2 | TCAGCCAGTCAACCTTGCTACCTTCC | |
| UPM (long) | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | |
| UPM (short) NUP | CTAATACGACTCACTATAGGGC AAGCAGTGGTATCAACGCAGAGT | |
| Recombinant expression | ||
| RE-PtCLec2 | CGCGGATCCCAAGGCCGTGTGTTGGCG | CCGCTCGAGACCAAAATGTGCTCGTGCGCG |
| qRT-PCR | ||
| PtCLec2-qRT | ACAGGACAACTGGAAGGTAGCAAG | GAGAAGAACCATCCCTTGACACTG |
| PtcSP1-qRT | ACTATGTCCAGCCAGCGTGT | GGAAGGACTCGCGCTCATAG |
| PtcSP2-qRT | TAAGGACATCGGACAGGAGACACT | TAAGGAAGTGAACGCTATCTCT |
| PtcSP3-qRT | AAGCCAGTCGAAATACAGGAG | CAGCATCTCCTTCCCAATTCC |
| PtSPH-qRT | CATCCTTGACCAGCCAGCA | CCCACCCAGACACAACACA |
| PtMyosin-qRT | CGTTGGCGAAGTAGGAGAGT | GAACAAGAGGCGTAATGAGGT |
| PtRab5-qRT | AACCCAGCATCCAGTCACCC | TACCCCTAAGCCCCTCAACC |
| PtArp-qRT | GCTCCACCATCACCACTCTG | TTAGCCATCTCCTTCCCTGC |
| PtproPO-qRT | CCTCTTCTTCACGACACTCAACTG | TCACGAGATAACACAAAACGCC |
| PtPPAF-qRT | GGACAGGACCAAGACCCAGT | GATTTGAGAAGGAACAAGCGTG |
| Ptα2M1-qRT | TGTGCCTCCTACCGCCTTCC | GGTGTCCCTCTCTCTCAACTCATTT |
| Ptα2M2-qRT | GTGGTTGGCTACGGGACGGGT | ACGGCAATGTCATCACTGGGGAT |
| PtTEP-qRT | CTCTTCTCGCTGCTTTCCTTCATC | TTTTTGGGACTTTGCCACCACTT |
| PtCrustin1-qRT | GGCAGTTGTGGCTACCATTGT | CGCTCGGTGTAAGGTGGATAG |
| PtCrustin2-qRT | TGGTGATGGCGGTGGCTATG | TCCTTGGCGAACATCCTCT |
| PtCrustin3-qRT | AGTATCTCAGAATCGACCA | CCCTCTTAGTTTCTCTTGTT |
| PtALF1-3-qRT | ACGACGAGGAGGAGAAAGAGG | GGCACTGATGGTGGAAACTGA |
| PtALF4-qRT | GACGCTCTGAAGGACTTTATG | CGCCGAAACGCTTAGAAATAC |
| PtALF5-qRT | TAGTCGTGGTGAGAGGGCAA | CTTTGCTCTCTCATCAGGAC |
| PtALF6-qRT | CATACTCCCGTGAACTCCTA | CAGACCGTGATTTGTGGAG |
| PtALF7-qRT | GCATTTTCTATTTTCCTATCC | GCATGAGTCTTGTATATTTGG |
| PtMBL-qRT | GGCACCGTCGGTCATCCAAC | GCTGGCTGCGACCAAACCTT |
| PtJNK-qRT | AGTGTGGCGGCTCAGCTGTT | CTCCACTCCGACTGCCTCGC |
| PtRelish-qRT | CCAGAGTACGCAAGCCACATCAC | CCGCAGCACCACCTTGTTCAG |
| PtPelle-qRT | ACTCTTGCCTTCCCTTGCTAAC | ACTGACCATGAATCATACCCCTG |
| PtMyD88-qRT | GGTCCTTGAAGCAACAGGTGGTAG | AGTGCTGGCTGACTAGGAGATGAC |
| PtTLR-qRT | CATTGAGGACAGCCACAGGAC | TGGTAGAGAGGTACAGCTTGAGTTC |
| β-actin-qRT | TCACACACTGTCCCCATCTACG | ACCACGCTCGGTCAGGATTTTC |
| RNAi | ||
| PtCLec2-siRNA | GACGCAACAUCACGUACAU | |
| GFP-siRNA | GGAGGACGGCAACAUCCUG | |
| Sequencing | ||
| M13 | CGCCAGGGTTTTCCCAGTCACGAC | GAGCGGATAACAATTTCACACAGG |
| T7 | TAATACGACTCACTATAGGG | GCTAGTTATTGCTCAGCGGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Su, Y.; Zhang, A.; Cui, Z. A C-Type Lectin Highly Expressed in Portunus trituberculatus Intestine Functions in AMP Regulation and Prophenoloxidase Activation. Antibiotics 2021, 10, 541. https://doi.org/10.3390/antibiotics10050541
Liu Y, Su Y, Zhang A, Cui Z. A C-Type Lectin Highly Expressed in Portunus trituberculatus Intestine Functions in AMP Regulation and Prophenoloxidase Activation. Antibiotics. 2021; 10(5):541. https://doi.org/10.3390/antibiotics10050541
Chicago/Turabian StyleLiu, Yuan, Yue Su, Ao Zhang, and Zhaoxia Cui. 2021. "A C-Type Lectin Highly Expressed in Portunus trituberculatus Intestine Functions in AMP Regulation and Prophenoloxidase Activation" Antibiotics 10, no. 5: 541. https://doi.org/10.3390/antibiotics10050541
APA StyleLiu, Y., Su, Y., Zhang, A., & Cui, Z. (2021). A C-Type Lectin Highly Expressed in Portunus trituberculatus Intestine Functions in AMP Regulation and Prophenoloxidase Activation. Antibiotics, 10(5), 541. https://doi.org/10.3390/antibiotics10050541