
Next-Generation Human Liver Models for Antimalarial Drug Assays


Abstract
:1. Global Impact of Malaria and Urgent Needs
2. Intrahepatic Development of Plasmodium Parasites: An Unfilled Gap in Malaria Biology
3. Limitations in the Treatment of Liver-Stage Malaria
4. Current In Vitro Hepatic Models for Human Malaria and Unaddressed Issues
5. Current Toolbox for Assaying the Plasmodicidal Activity of Liver-Stage Malaria
6. Three-Dimensional Cell Culture Is Emerging as a Next-Generation Malaria Model
6.1. Spheroids
6.1.1. Human Hepatoma and Immortalized Hepatic Cell-Derived Spheroids
6.1.2. Human Primary Hepatocyte-Derived Spheroids
6.2. Organoids
6.2.1. Single Cell Type-Derived Hepatic Organoids
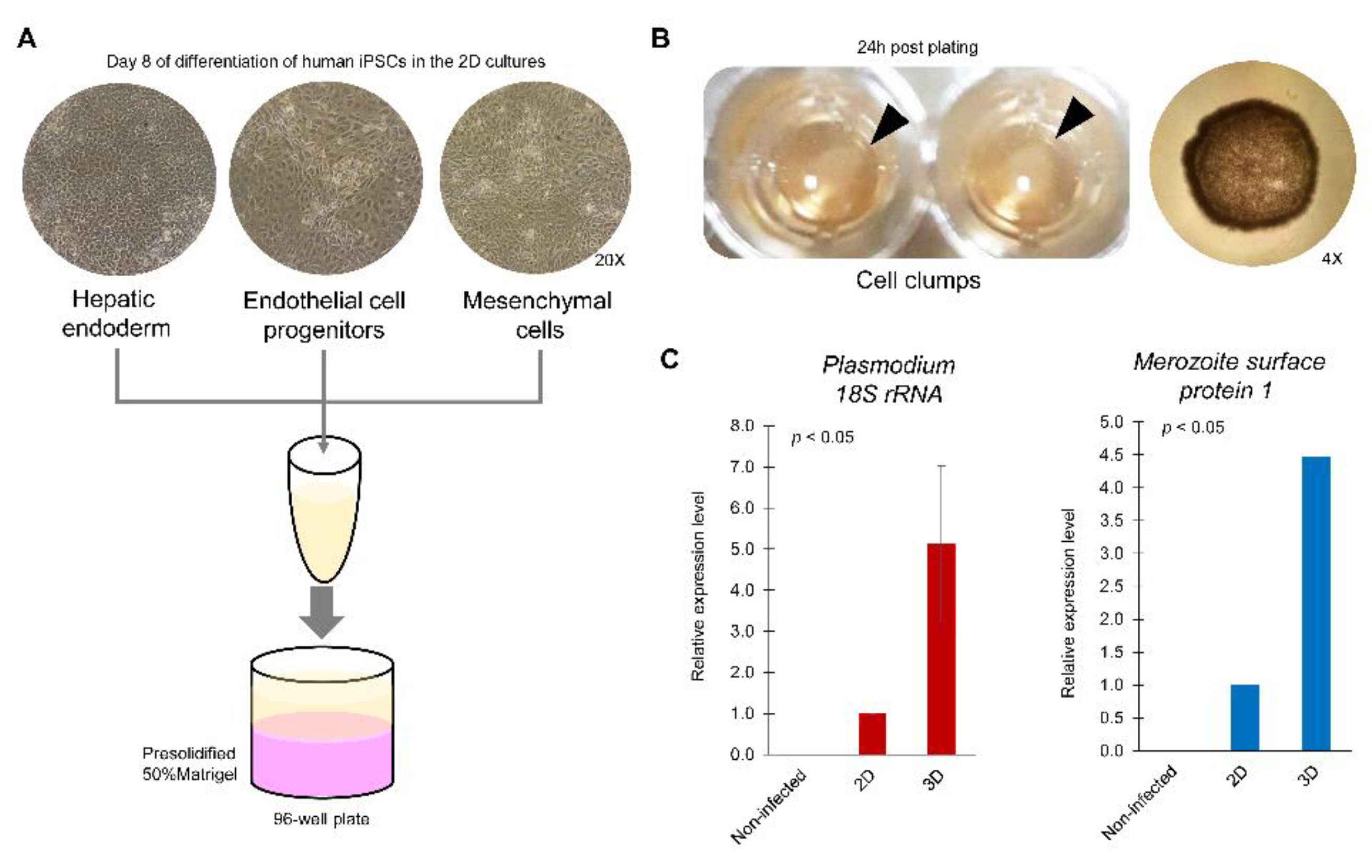
6.2.2. Unknown Cell Type
6.2.3. Multiple Cell Types for Heterogenous Organoids
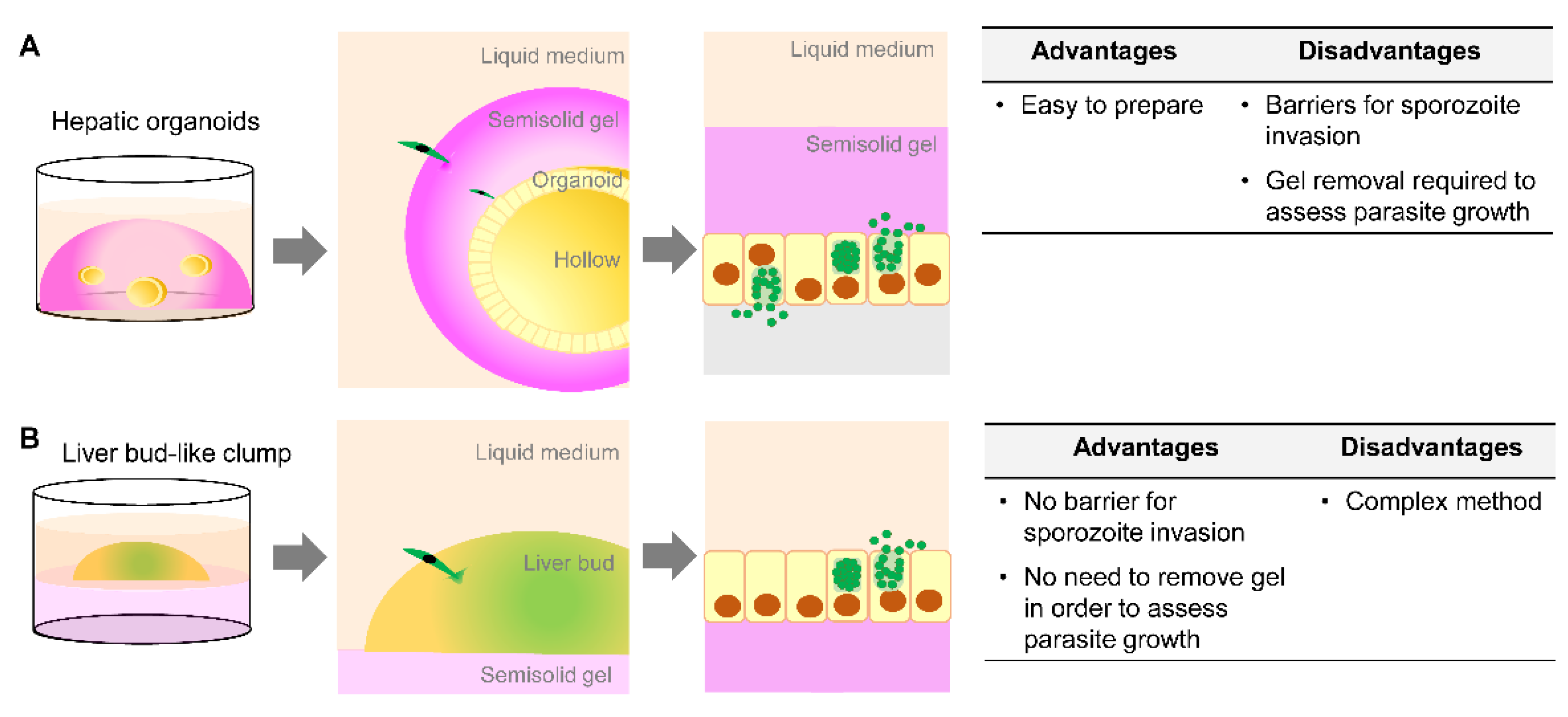
7. Opportunity for the Use of Organoids in Drug Development for Liver-Stage Human Malaria
8. Challenges in the Preclinical Use of Organoids as a Liver-Stage Malaria Model
9. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2020: 20 Years of Global Progress and Challenges; World Health Organization: Geneva, Switzerland, 2020; pp. 18–37. [Google Scholar]
- Lalloo, D.G.; Olukoya, P.; Olliaro, P. Malaria in adolescence: Burden of disease, consequences, and opportunities for intervention. Lancet Infect. Dis. 2006, 6, 780–793. [Google Scholar] [CrossRef]
- Wells, T.N.; Burrows, J.N.; Baird, J.K. Targeting the hypnozoite reservoir of plasmodium vivax: The hidden obstacle to malaria elimination. Trends Parasitol. 2010, 26, 145–151. [Google Scholar] [CrossRef]
- Lacerda, M.V.G.; Llanos-Cuentas, A.; Krudsood, S.; Lon, C.; Saunders, D.L.; Mohammed, R.; Yilma, D.; Batista Pereira, D.; Espino, F.E.J.; Mia, R.Z.; et al. Single-dose tafenoquine to prevent relapse of plasmodium vivax malaria. N. Engl. J. Med. 2019, 380, 215–228. [Google Scholar] [CrossRef]
- Llanos-Cuentas, A.; Lacerda, M.V.G.; Hien, T.T.; Velez, I.D.; Namaik-Larp, C.; Chu, C.S.; Villegas, M.F.; Val, F.; Monteiro, W.M.; Brito, M.A.M.; et al. Tafenoquine versus primaquine to prevent relapse of plasmodium vivax malaria. N. Engl. J. Med. 2019, 380, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Arrowsmith, J.; Miller, P. Trial watch: Phase ii and phase iii attrition rates 2011–2012. Nat. Rev. Drug Discov. 2013, 12, 569. [Google Scholar] [CrossRef]
- White, N.J.; Pukrittayakamee, S.; Hien, T.T.; Faiz, M.A.; Mokuolu, O.A.; Dondorp, A.M. Malaria. Lancet 2014, 383, 723–735. [Google Scholar] [CrossRef]
- Ejigiri, I.; Sinnis, P. Plasmodium sporozoite-host interactions from the dermis to the hepatocyte. Curr. Opin. Microbiol. 2009, 12, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Deslyper, G.; Doherty, D.G.; Carolan, J.C.; Holland, C.V. The role of the liver in the migration of parasites of global significance. Parasites Vectors 2019, 12, 531. [Google Scholar] [CrossRef] [Green Version]
- Price, R.N.; Tjitra, E.; Guerra, C.A.; Yeung, S.; White, N.J.; Anstey, N.M. Vivax malaria: Neglected and not benign. Am. J. Trop. Med. Hyg. 2007, 77, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Flint, M.; Maidens, C.; Loomis-Price, L.D.; Shotton, C.; Dubuisson, J.; Monk, P.; Higginbottom, A.; Levy, S.; McKeating, J.A. Characterization of hepatitis c virus e2 glycoprotein interaction with a putative cellular receptor, cd81. J. Virol. 1999, 73, 6235–6244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzoni, G.; Marinach, C.; Topcu, S.; Briquet, S.; Grand, M.; Tolle, M.; Gransagne, M.; Lescar, J.; Andolina, C.; Franetich, J.F.; et al. Plasmodium p36 determines host cell receptor usage during sporozoite invasion. eLife 2017, 6, e25903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingdale, M.R.; Collins, W.E.; Campbell, C.C.; Schwartz, A.L. In vitro culture of two populations (dividing and nondividing) of exoerythrocytic parasites of plasmodium vivax. Am. J. Trop. Med. Hyg. 1985, 34, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Karnasuta, C.; Pavanand, K.; Chantakulkij, S.; Luttiwongsakorn, N.; Rassamesoraj, M.; Laohathai, K.; Webster, H.K.; Watt, G. Complete development of the liver stage of plasmodium falciparum in a human hepatoma cell line. Am. J. Trop. Med. Hyg. 1995, 53, 607–611. [Google Scholar] [CrossRef]
- Hollingdale, M.R.; Nardin, E.H.; Tharavanij, S.; Schwartz, A.L.; Nussenzweig, R.S. Inhibition of entry of plasmodium falciparum and p. Vivax sporozoites into cultured cells; an in vitro assay of protective antibodies. J. Immunol. 1984, 132, 909–913. [Google Scholar] [PubMed]
- Mazier, D.; Beaudoin, R.L.; Mellouk, S.; Druilhe, P.; Texier, B.; Trosper, J.; Miltgen, F.; Landau, I.; Paul, C.; Brandicourt, O.; et al. Complete development of hepatic stages of plasmodium falciparum in vitro. Science 1985, 227, 440–442. [Google Scholar] [CrossRef]
- March, S.; Ng, S.; Velmurugan, S.; Galstian, A.; Shan, J.; Logan, D.J.; Carpenter, A.E.; Thomas, D.; Sim, B.K.; Mota, M.M.; et al. A microscale human liver platform that supports the hepatic stages of plasmodium falciparum and vivax. Cell Host Microbe 2013, 14, 104–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, A.; Maher, S.P.; Conway, A.J.; Ubalee, R.; Chaumeau, V.; Andolina, C.; Kaba, S.A.; Vantaux, A.; Bakowski, M.A.; Thomson-Luque, R.; et al. A comprehensive model for assessment of liver stage therapies targeting plasmodium vivax and plasmodium falciparum. Nat. Commun. 2018, 9, 1837. [Google Scholar] [CrossRef] [Green Version]
- Sattabongkot, J.; Yimamnuaychoke, N.; Leelaudomlipi, S.; Rasameesoraj, M.; Jenwithisuk, R.; Coleman, R.E.; Udomsangpetch, R.; Cui, L.; Brewer, T.G. Establishment of a human hepatocyte line that supports in vitro development of the exo-erythrocytic stages of the malaria parasites plasmodium falciparum and p. Vivax. Am. J. Trop. Med. Hyg. 2006, 74, 708–715. [Google Scholar] [CrossRef] [Green Version]
- Kaushansky, A.; Douglass, A.N.; Arang, N.; Vigdorovich, V.; Dambrauskas, N.; Kain, H.S.; Austin, L.S.; Sather, D.N.; Kappe, S.H. Malaria parasites target the hepatocyte receptor epha2 for successful host infection. Science 2015, 350, 1089–1092. [Google Scholar] [CrossRef] [Green Version]
- Pewkliang, Y.; Rungin, S.; Lerdpanyangam, K.; Duangmanee, A.; Kanjanasirirat, P.; Suthivanich, P.; Sa-Ngiamsuntorn, K.; Borwornpinyo, S.; Sattabongkot, J.; Patrapuvich, R.; et al. A novel immortalized hepatocyte-like cell line (imhc) supports in vitro liver stage development of the human malarial parasite plasmodium vivax. Malar. J. 2018, 17, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, S.; Schwartz, R.E.; March, S.; Galstian, A.; Gural, N.; Shan, J.; Prabhu, M.; Mota, M.M.; Bhatia, S.N. Human ipsc-derived hepatocyte-like cells support plasmodium liver-stage infection in vitro. Stem Cell Rep. 2015, 4, 348–359. [Google Scholar] [CrossRef] [Green Version]
- Silvie, O.; Rubinstein, E.; Franetich, J.F.; Prenant, M.; Belnoue, E.; Renia, L.; Hannoun, L.; Eling, W.; Levy, S.; Boucheix, C.; et al. Hepatocyte cd81 is required for plasmodium falciparum and plasmodium yoelii sporozoite infectivity. Nat. Med. 2003, 9, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.D.; Hannus, M.; Prudencio, M.; Martin, C.; Goncalves, L.A.; Portugal, S.; Epiphanio, S.; Akinc, A.; Hadwiger, P.; Jahn-Hofmann, K.; et al. Host scavenger receptor sr-bi plays a dual role in the establishment of malaria parasite liver infection. Cell Host Microbe 2008, 4, 271–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvie, O.; Charrin, S.; Billard, M.; Franetich, J.F.; Clark, K.L.; van Gemert, G.J.; Sauerwein, R.W.; Dautry, F.; Boucheix, C.; Mazier, D.; et al. Cholesterol contributes to the organization of tetraspanin-enriched microdomains and to cd81-dependent infection by malaria sporozoites. J. Cell Sci. 2006, 119, 1992–2002. [Google Scholar] [CrossRef] [Green Version]
- Yalaoui, S.; Huby, T.; Franetich, J.F.; Gego, A.; Rametti, A.; Moreau, M.; Collet, X.; Siau, A.; van Gemert, G.J.; Sauerwein, R.W.; et al. Scavenger receptor bi boosts hepatocyte permissiveness to plasmodium infection. Cell Host Microbe 2008, 4, 283–292. [Google Scholar] [CrossRef] [Green Version]
- McCall, M.B.B.; Wammes, L.J.; Langenberg, M.C.C.; van Gemert, G.J.; Walk, J.; Hermsen, C.C.; Graumans, W.; Koelewijn, R.; Franetich, J.F.; Chishimba, S.; et al. Infectivity of plasmodium falciparum sporozoites determines emerging parasitemia in infected volunteers. Sci. Transl. Med. 2017, 9, eaag2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, X.; House, B.L.; Zyzak, M.D.; Richie, T.L.; Gerbasi, V.R. Towards an optimized inhibition of liver stage development assay (ilsda) for plasmodium falciparum. Malar. J. 2013, 12, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sa-ngiamsuntorn, K.; Wongkajornsilp, A.; Kasetsinsombat, K.; Duangsa-ard, S.; Nuntakarn, L.; Borwornpinyo, S.; Akarasereenont, P.; Limsrichamrern, S.; Hongeng, S. Upregulation of cyp 450s expression of immortalized hepatocyte-like cells derived from mesenchymal stem cells by enzyme inducers. BMC Biotechnol. 2011, 11, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si-Tayeb, K.; Noto, F.K.; Nagaoka, M.; Li, J.; Battle, M.A.; Duris, C.; North, P.E.; Dalton, S.; Duncan, S.A. Highly efficient generation of human hepatocyte-like cells from induced pluripotent stem cells. Hepatology 2010, 51, 297–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, R.E.; Fleming, H.E.; Khetani, S.R.; Bhatia, S.N. Pluripotent stem cell-derived hepatocyte-like cells. Biotechnol. Adv. 2014, 32, 504–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, R.E.; Trehan, K.; Andrus, L.; Sheahan, T.P.; Ploss, A.; Duncan, S.A.; Rice, C.M.; Bhatia, S.N. Modeling hepatitis c virus infection using human induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2012, 109, 2544–2548. [Google Scholar] [CrossRef] [Green Version]
- Shlomai, A.; Schwartz, R.E.; Ramanan, V.; Bhatta, A.; de Jong, Y.P.; Bhatia, S.N.; Rice, C.M. Modeling host interactions with hepatitis b virus using primary and induced pluripotent stem cell-derived hepatocellular systems. Proc. Natl. Acad. Sci. USA 2014, 111, 12193–12198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, A.M.; Mikolajczak, S.A.; Wilson, E.M.; Grompe, M.; Kaushansky, A.; Camargo, N.; Bial, J.; Ploss, A.; Kappe, S.H. Complete plasmodium falciparum liver-stage development in liver-chimeric mice. J. Clin. Investig. 2012, 122, 3618–3628. [Google Scholar] [CrossRef]
- Kaushansky, A.; Austin, L.S.; Mikolajczak, S.A.; Lo, F.Y.; Miller, J.L.; Douglass, A.N.; Arang, N.; Vaughan, A.M.; Gardner, M.J.; Kappe, S.H. Susceptibility to plasmodium yoelii preerythrocytic infection in balb/c substrains is determined at the point of hepatocyte invasion. Infect. Immun. 2015, 83, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Soulard, V.; Bosson-Vanga, H.; Lorthiois, A.; Roucher, C.; Franetich, J.F.; Zanghi, G.; Bordessoulles, M.; Tefit, M.; Thellier, M.; Morosan, S.; et al. Plasmodium falciparum full life cycle and plasmodium ovale liver stages in humanized mice. Nat. Commun. 2015, 6, 7690. [Google Scholar] [CrossRef] [Green Version]
- Shan, J.; Schwartz, R.E.; Ross, N.T.; Logan, D.J.; Thomas, D.; Duncan, S.A.; North, T.E.; Goessling, W.; Carpenter, A.E.; Bhatia, S.N. Identification of small molecules for human hepatocyte expansion and ips differentiation. Nat. Chem. Biol. 2013, 9, 514–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Pybus, B.S.; Marcsisin, R.; Logan, T.; Luong, T.L.; Sousa, J.; Matlock, N.; Collazo, V.; Asher, C.; Carroll, D.; et al. An lc-ms based study of the metabolic profile of primaquine, an 8-aminoquinoline antiparasitic drug, with an in vitro primary human hepatocyte culture model. Eur. J. Drug Metab. Pharmacokinet. 2014, 39, 139–146. [Google Scholar] [CrossRef]
- Pybus, B.S.; Sousa, J.C.; Jin, X.; Ferguson, J.A.; Christian, R.E.; Barnhart, R.; Vuong, C.; Sciotti, R.J.; Reichard, G.A.; Kozar, M.P.; et al. Cyp450 phenotyping and accurate mass identification of metabolites of the 8-aminoquinoline, anti-malarial drug primaquine. Malar. J. 2012, 11, 259. [Google Scholar] [CrossRef] [Green Version]
- Pybus, B.S.; Marcsisin, S.R.; Jin, X.; Deye, G.; Sousa, J.C.; Li, Q.; Caridha, D.; Zeng, Q.; Reichard, G.A.; Ockenhouse, C.; et al. The metabolism of primaquine to its active metabolite is dependent on cyp 2d6. Malar. J. 2013, 12, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khetani, S.R.; Bhatia, S.N. Microscale culture of human liver cells for drug development. Nat. Biotechnol. 2008, 26, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Biagini, G.A.; Fisher, N.; Shone, A.E.; Mubaraki, M.A.; Srivastava, A.; Hill, A.; Antoine, T.; Warman, A.J.; Davies, J.; Pidathala, C.; et al. Generation of quinolone antimalarials targeting the plasmodium falciparum mitochondrial respiratory chain for the treatment and prophylaxis of malaria. Proc. Natl. Acad. Sci. USA 2012, 109, 8298–8303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, J.W.; Pybus, B.S.; Yadava, A.; Tosh, D.; Sousa, J.C.; McCarthy, W.F.; Deye, G.; Melendez, V.; Ockenhouse, C.F. Primaquine failure and cytochrome p-450 2d6 in plasmodium vivax malaria. N. Engl. J. Med. 2013, 369, 1381–1382. [Google Scholar] [CrossRef] [PubMed]
- Antonova-Koch, Y.; Meister, S.; Abraham, M.; Luth, M.R.; Ottilie, S.; Lukens, A.K.; Sakata-Kato, T.; Vanaerschot, M.; Owen, E.; Jado, J.C.; et al. Open-source discovery of chemical leads for next-generation chemoprotective antimalarials. Science 2018, 362, eaat9446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meister, S.; Plouffe, D.M.; Kuhen, K.L.; Bonamy, G.M.; Wu, T.; Barnes, S.W.; Bopp, S.E.; Borboa, R.; Bright, A.T.; Che, J.; et al. Imaging of plasmodium liver stages to drive next-generation antimalarial drug discovery. Science 2011, 334, 1372–1377. [Google Scholar] [CrossRef] [Green Version]
- Brunschwig, C.; Lawrence, N.; Taylor, D.; Abay, E.; Njoroge, M.; Basarab, G.S.; Le Manach, C.; Paquet, T.; Cabrera, D.G.; Nchinda, A.T.; et al. Uct943, a next-generation plasmodium falciparum pi4k inhibitor preclinical candidate for the treatment of malaria. Antimicrob. Agents Chemother. 2018, 62, e00012-18. [Google Scholar] [CrossRef] [Green Version]
- Rubashkin, M.G.; Ou, G.; Weaver, V.M. Deconstructing signaling in three dimensions. Biochemistry 2014, 53, 2078–2090. [Google Scholar] [CrossRef]
- Arez, F.; Rebelo, S.P.; Fontinha, D.; Simao, D.; Martins, T.R.; Machado, M.; Fischli, C.; Oeuvray, C.; Badolo, L.; Carrondo, M.J.T.; et al. Flexible 3d cell-based platforms for the discovery and profiling of novel drugs targeting plasmodium hepatic infection. ACS Infect. Dis. 2019, 5, 1831–1842. [Google Scholar] [CrossRef]
- Chua, A.C.Y.; Ananthanarayanan, A.; Ong, J.J.Y.; Wong, J.Y.; Yip, A.; Singh, N.H.; Qu, Y.; Dembele, L.; McMillian, M.; Ubalee, R.; et al. Hepatic spheroids used as an in vitro model to study malaria relapse. Biomaterials 2019, 216, 119221. [Google Scholar] [CrossRef]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, J.F.; Wiegerinck, C.L.; de Jonge, H.R.; Bronsveld, I.; Janssens, H.M.; de Winter-de Groot, K.M.; Brandsma, A.M.; de Jong, N.W.; Bijvelds, M.J.; Scholte, B.J.; et al. A functional cftr assay using primary cystic fibrosis intestinal organoids. Nat. Med. 2013, 19, 939–945. [Google Scholar] [CrossRef]
- Huch, M.; Gehart, H.; van Boxtel, R.; Hamer, K.; Blokzijl, F.; Verstegen, M.M.; Ellis, E.; van Wenum, M.; Fuchs, S.A.; de Ligt, J.; et al. Long-term culture of genome-stable bipotent stem cells from adult human liver. Cell 2015, 160, 299–312. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.; Vela, I.; Sboner, A.; Iaquinta, P.J.; Karthaus, W.R.; Gopalan, A.; Dowling, C.; Wanjala, J.N.; Undvall, E.A.; Arora, V.K.; et al. Organoid cultures derived from patients with advanced prostate cancer. Cell 2014, 159, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Boj, S.F.; Hwang, C.I.; Baker, L.A.; Chio, I.I.; Engle, D.D.; Corbo, V.; Jager, M.; Ponz-Sarvise, M.; Tiriac, H.; Spector, M.S.; et al. Organoid models of human and mouse ductal pancreatic cancer. Cell 2015, 160, 324–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubert, C.G.; Rivera, M.; Spangler, L.C.; Wu, Q.; Mack, S.C.; Prager, B.C.; Couce, M.; McLendon, R.E.; Sloan, A.E.; Rich, J.N. A three-dimensional organoid culture system derived from human glioblastomas recapitulates the hypoxic gradients and cancer stem cell heterogeneity of tumors found in vivo. Cancer Res. 2016, 76, 2465–2477. [Google Scholar] [CrossRef] [Green Version]
- Mun, S.J.; Ryu, J.S.; Lee, M.O.; Son, Y.S.; Oh, S.J.; Cho, H.S.; Son, M.Y.; Kim, D.S.; Kim, S.J.; Yoo, H.J.; et al. Generation of expandable human pluripotent stem cell-derived hepatocyte-like liver organoids. J. Hepatol. 2019, 71, 970–985. [Google Scholar] [CrossRef]
- Kretzschmar, K.; Clevers, H. Organoids: Modeling development and the stem cell niche in a dish. Dev. Cell 2016, 38, 590–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huch, M.; Koo, B.K. Modeling mouse and human development using organoid cultures. Development 2015, 142, 3113–3125. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Clevers, H. Growing self-organizing mini-guts from a single intestinal stem cell: Mechanism and applications. Science 2013, 340, 1190–1194. [Google Scholar] [CrossRef] [Green Version]
- Pampaloni, F.; Reynaud, E.G.; Stelzer, E.H. The third dimension bridges the gap between cell culture and live tissue. Nat. Rev. Mol. Cell Biol. 2007, 8, 839–845. [Google Scholar] [CrossRef]
- Chen, Y.F.; Tseng, C.Y.; Wang, H.W.; Kuo, H.C.; Yang, V.W.; Lee, O.K. Rapid generation of mature hepatocyte-like cells from human induced pluripotent stem cells by an efficient three-step protocol. Hepatology 2012, 55, 1193–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmelzer, E.; Zhang, L.; Bruce, A.; Wauthier, E.; Ludlow, J.; Yao, H.L.; Moss, N.; Melhem, A.; McClelland, R.; Turner, W.; et al. Human hepatic stem cells from fetal and postnatal donors. J. Exp. Med. 2007, 204, 1973–1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broutier, L.; Andersson-Rolf, A.; Hindley, C.J.; Boj, S.F.; Clevers, H.; Koo, B.K.; Huch, M. Culture and establishment of self-renewing human and mouse adult liver and pancreas 3d organoids and their genetic manipulation. Nat. Protoc. 2016, 11, 1724–1743. [Google Scholar] [CrossRef]
- Akbari, S.; Sevinc, G.G.; Ersoy, N.; Basak, O.; Kaplan, K.; Sevinc, K.; Ozel, E.; Sengun, B.; Enustun, E.; Ozcimen, B.; et al. Robust, long-term culture of endoderm-derived hepatic organoids for disease modeling. Stem Cell Rep. 2019, 13, 627–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkeaw, K.; Tubsuwan, A.; Tongkrajang, N.; Whangviboonkij, N. Generation of human liver organoids from pluripotent stem cell-derived hepatic endoderms. PeerJ 2020, 8, e9968. [Google Scholar] [CrossRef] [PubMed]
- Takebe, T.; Sekine, K.; Suzuki, Y.; Enomura, M.; Tanaka, S.; Ueno, Y.; Zheng, Y.W.; Taniguchi, H. Self-organization of human hepatic organoid by recapitulating organogenesis in vitro. Transplant. Proc. 2012, 44, 1018–1020. [Google Scholar] [CrossRef] [PubMed]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.R.; Ueno, Y.; Zheng, Y.W.; Koike, N.; et al. Vascularized and functional human liver from an ipsc-derived organ bud transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Schlaermann, P.; Toelle, B.; Berger, H.; Schmidt, S.C.; Glanemann, M.; Ordemann, J.; Bartfeld, S.; Mollenkopf, H.J.; Meyer, T.F. A novel human gastric primary cell culture system for modelling helicobacter pylori infection in vitro. Gut 2016, 65, 202–213. [Google Scholar] [CrossRef] [Green Version]
- Wroblewski, L.E.; Piazuelo, M.B.; Chaturvedi, R.; Schumacher, M.; Aihara, E.; Feng, R.; Noto, J.M.; Delgado, A.; Israel, D.A.; Zavros, Y.; et al. Helicobacter pylori targets cancer-associated apical-junctional constituents in gastroids and gastric epithelial cells. Gut 2015, 64, 720–730. [Google Scholar] [CrossRef] [Green Version]
- DesRochers, T.M.; Kimmerling, E.P.; Jandhyala, D.M.; El-Jouni, W.; Zhou, J.; Thorpe, C.M.; Leong, J.M.; Kaplan, D.L. Effects of shiga toxin type 2 on a bioengineered three-dimensional model of human renal tissue. Infect. Immun. 2015, 83, 28–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, K.; Kawabata, K.; Nagamoto, Y.; Kishimoto, K.; Tashiro, K.; Sakurai, F.; Tachibana, M.; Kanda, K.; Hayakawa, T.; Furue, M.K.; et al. 3d spheroid culture of hesc/hipsc-derived hepatocyte-like cells for drug toxicity testing. Biomaterials 2013, 34, 1781–1789. [Google Scholar] [CrossRef]
- Haque, A.; Hexig, B.; Meng, Q.; Hossain, S.; Nagaoka, M.; Akaike, T. The effect of recombinant e-cadherin substratum on the differentiation of endoderm-derived hepatocyte-like cells from embryonic stem cells. Biomaterials 2011, 32, 2032–2042. [Google Scholar] [CrossRef] [PubMed]
- Vlachogiannis, G.; Hedayat, S.; Vatsiou, A.; Jamin, Y.; Fernandez-Mateos, J.; Khan, K.; Lampis, A.; Eason, K.; Huntingford, I.; Burke, R.; et al. Patient-derived organoids model treatment response of metastatic gastrointestinal cancers. Science 2018, 359, 920–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, T.D.; Eastwood, J.P.; Shay, M.A.; Drake, J.F.; Sonnerup, B.U.O.; Fujimoto, M.; Cassak, P.A.; Oieroset, M.; Burch, J.L.; Torbert, R.B.; et al. Publisher correction: Electron magnetic reconnection without ion coupling in earth’s turbulent magnetosheath. Nature 2019, 569, E9. [Google Scholar] [CrossRef] [Green Version]
- Hofer, M.; Lutolf, M.P. Engineering organoids. Nat. Rev. Mater. 2021, 6, 402–420. [Google Scholar] [CrossRef] [PubMed]
- Bunnik, E.M.; Venkat, A.; Shao, J.; McGovern, K.E.; Batugedara, G.; Worth, D.; Prudhomme, J.; Lapp, S.A.; Andolina, C.; Ross, L.S.; et al. Comparative 3d genome organization in apicomplexan parasites. Proc. Natl. Acad. Sci. USA 2019, 116, 3183–3192. [Google Scholar] [CrossRef] [Green Version]
- Hoo, R.; Zhu, L.; Amaladoss, A.; Mok, S.; Natalang, O.; Lapp, S.A.; Hu, G.; Liew, K.; Galinski, M.R.; Bozdech, Z.; et al. Integrated analysis of the plasmodium species transcriptome. EBioMedicine 2016, 7, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models | Infection Rate (%) Based on | Merozoites | Hypnozoites † (dpi) | Applications | References | |||
|---|---|---|---|---|---|---|---|---|
| Sporozoites * | Hepatocytes | Detection (dpi) | Infectivity | |||||
| Hepatocellular carcinoma cell lines | HepG2-A16 | 0.0001 (P.v.) | N/A | 9 | N/A | Yes (5–15) | Development | [13] |
| 0.4–2.5 (P.f.) | N/A | N/A | N/A | N/A | Sporozoite invasion | [15] | ||
| HHS-102 | N/A | 0.009 (P.f.) | 12–13 | Yes | Not applicable | Development | [14] | |
| Primary human hepatocytes | Mazier et al., 1985 | N/A | N/A | 12–13 | Yes | Not applicable | Development | [16] |
| March et al., 2013 | 0.03 (P.f.) 0.013 (P.v.) | 0.18 (P.f.) | 6–10 | Yes (P.f.) | Yes (up to 21) | Development Vaccination Drug testing and screening | [17] | |
| Roth et al., 2018 | 0.6–2 (P.f.) 2–8.3 (P.v.) | N/A | 7–8 (P.f.) 9–11 (P.v.) | Yes | Yes (6–8) | Development Sporozoite invasion Drug testing and screening | [18] | |
| Immortalized cells | HC-04 | N/A | 0.066 (P.f.) 0.041 (P.v.) | 7 (P.f.) 10 (P.v.) | Yes Yes | Yes (28) | Development Sporozoite invasion | [19] [20] |
| imHC | 0.14 ± 0.16 (P.v.) | N/A | 10 (P.v.) | N/A | Yes (14) | Development Drug testing | [21] | |
| Pluripotent stem cells | Hepatocytes | N/A | N/A | 6 (P.f.) 8 (P.v.) | N/A | N/A | Development Drug testing | [22] |
| Cell type | ECM | Well Format | Culture | Drugs | Biomarkers | References |
|---|---|---|---|---|---|---|
| Primary human hepatocytes (PHHs) | Type I collagen (rat tail) | 96-well plates | Coculture with murine embryonic fibroblasts |
| Circumsporozoite protein (P.f.) | [17] |
| 384-well plate | Monoculture |
| GAPDH (P.f.) UIS4 (P.v.) | [18] | ||
| imHCs | Matrigel | 8-well plate | Monoculture |
| Plasmodium HSP70 UIS4 (P.v.) | [21] |
| iPSC-derived hepatocytes | No information | No information | Monoculture |
| HSP70 (P.f.) | [22] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulkeaw, K. Next-Generation Human Liver Models for Antimalarial Drug Assays. Antibiotics 2021, 10, 642. https://doi.org/10.3390/antibiotics10060642
Kulkeaw K. Next-Generation Human Liver Models for Antimalarial Drug Assays. Antibiotics. 2021; 10(6):642. https://doi.org/10.3390/antibiotics10060642
Chicago/Turabian StyleKulkeaw, Kasem. 2021. "Next-Generation Human Liver Models for Antimalarial Drug Assays" Antibiotics 10, no. 6: 642. https://doi.org/10.3390/antibiotics10060642
APA StyleKulkeaw, K. (2021). Next-Generation Human Liver Models for Antimalarial Drug Assays. Antibiotics, 10(6), 642. https://doi.org/10.3390/antibiotics10060642