
Evaluation of Microalgae Antiviral Activity and Their Bioactive Compounds




Abstract
:1. Introduction
2. Microalgae Antiviral Compounds
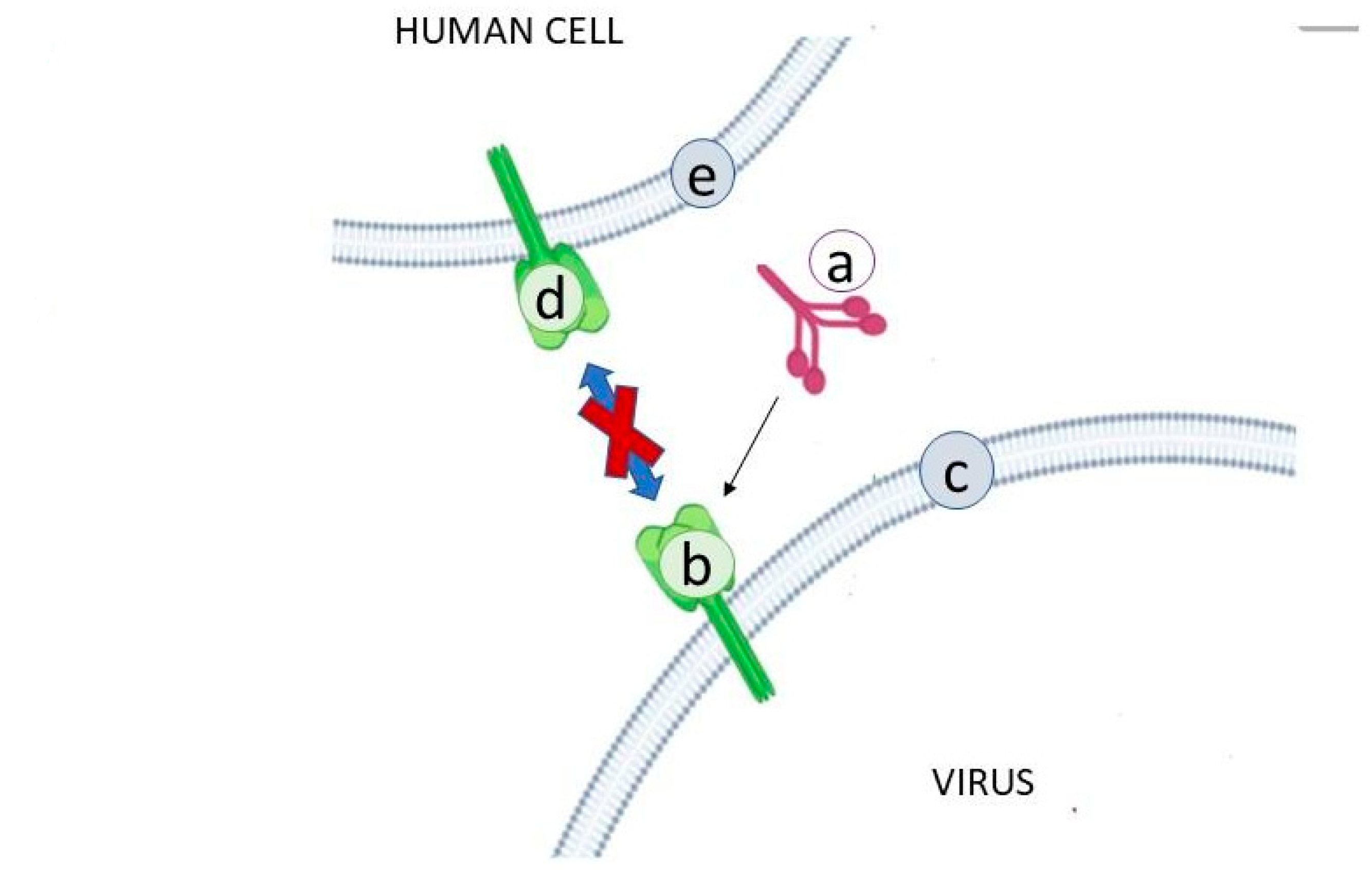
2.1. Lectins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
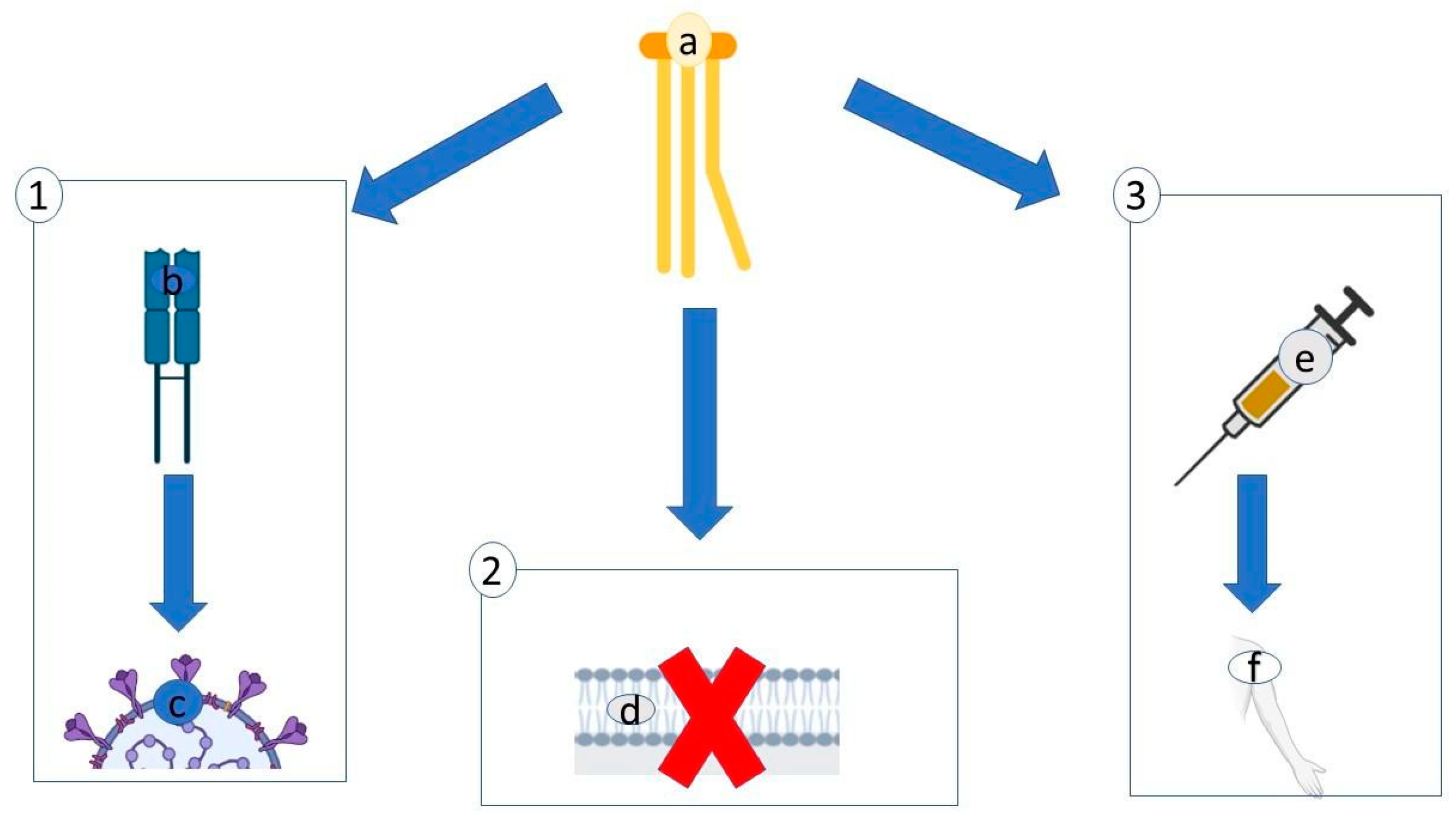{kind=link}
{kind=link}
| Lectins Interact Directly with the High-Glycan Structure of Viral Envelope Glycoproteins | |||
| Name | Organism | Virus | References |
| Agglutinin OAA | Oscillatoria agardhii | HIV1, HIV2 | [31,32,33] |
| CV-N | Nostoc ellipsosporum | HIV1, HSV1, HCV, IAV, IBV | [35,36,39,40,41,42,50,52] |
| Microvirin | Microcystis aeruginosa | HIV1, HIV2 | [54,55] |
| MVL | Microcystin viridis | HIV1, HIV2, HCV | [57,58,59,60] |
| Scytovirin | Scytonema varium | HIV1, ZEBOV | [61,62,63] |
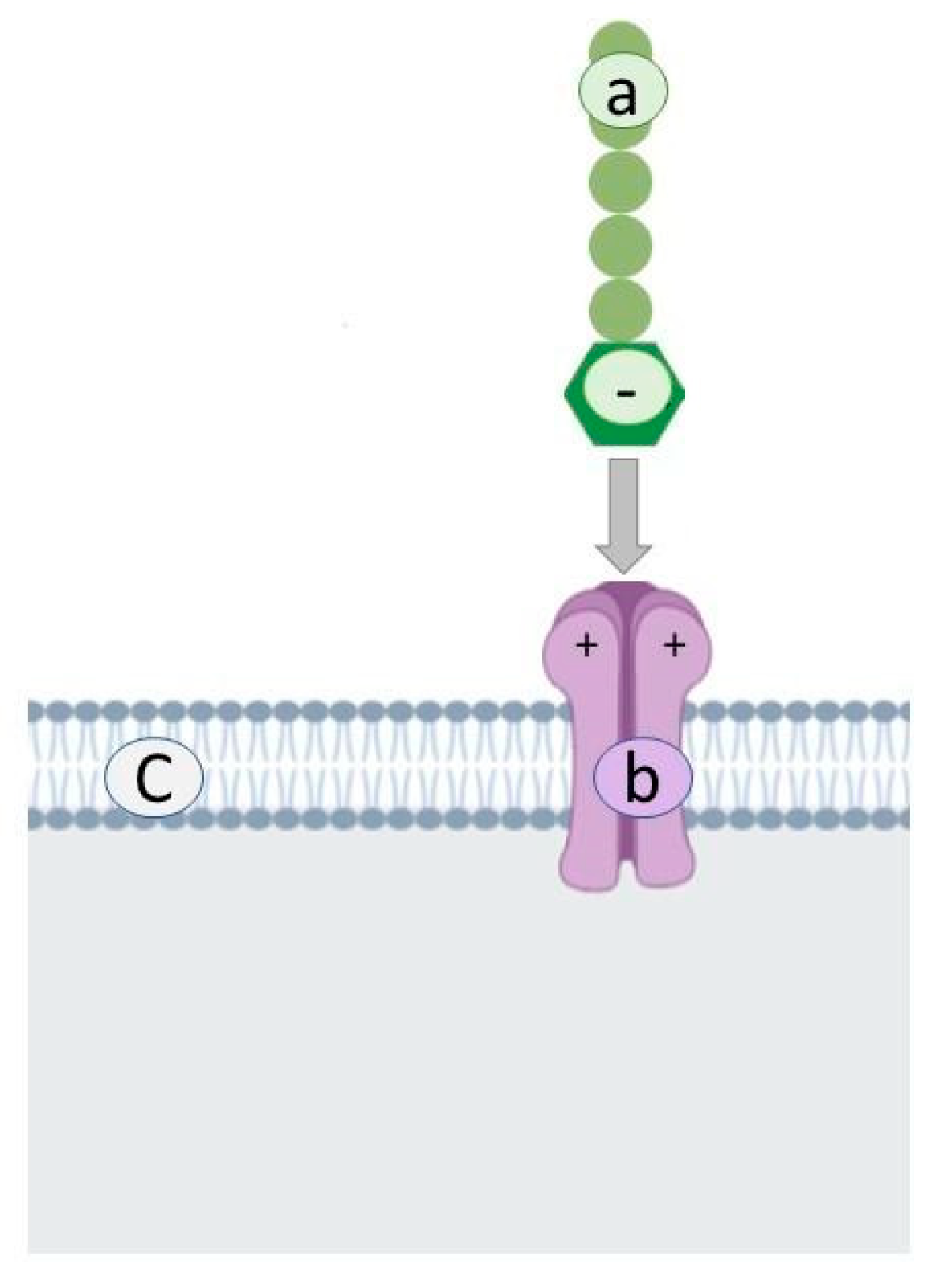
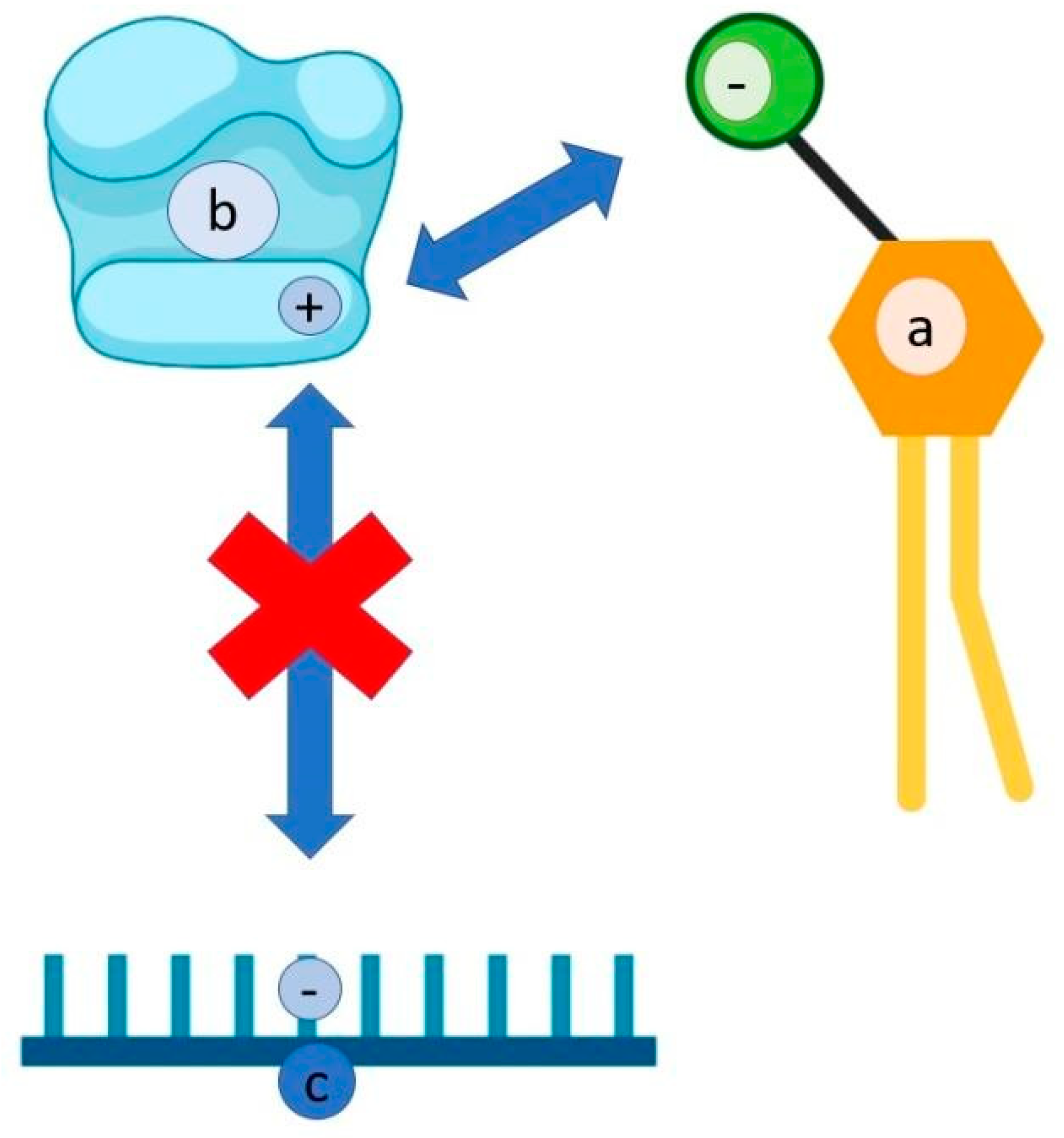
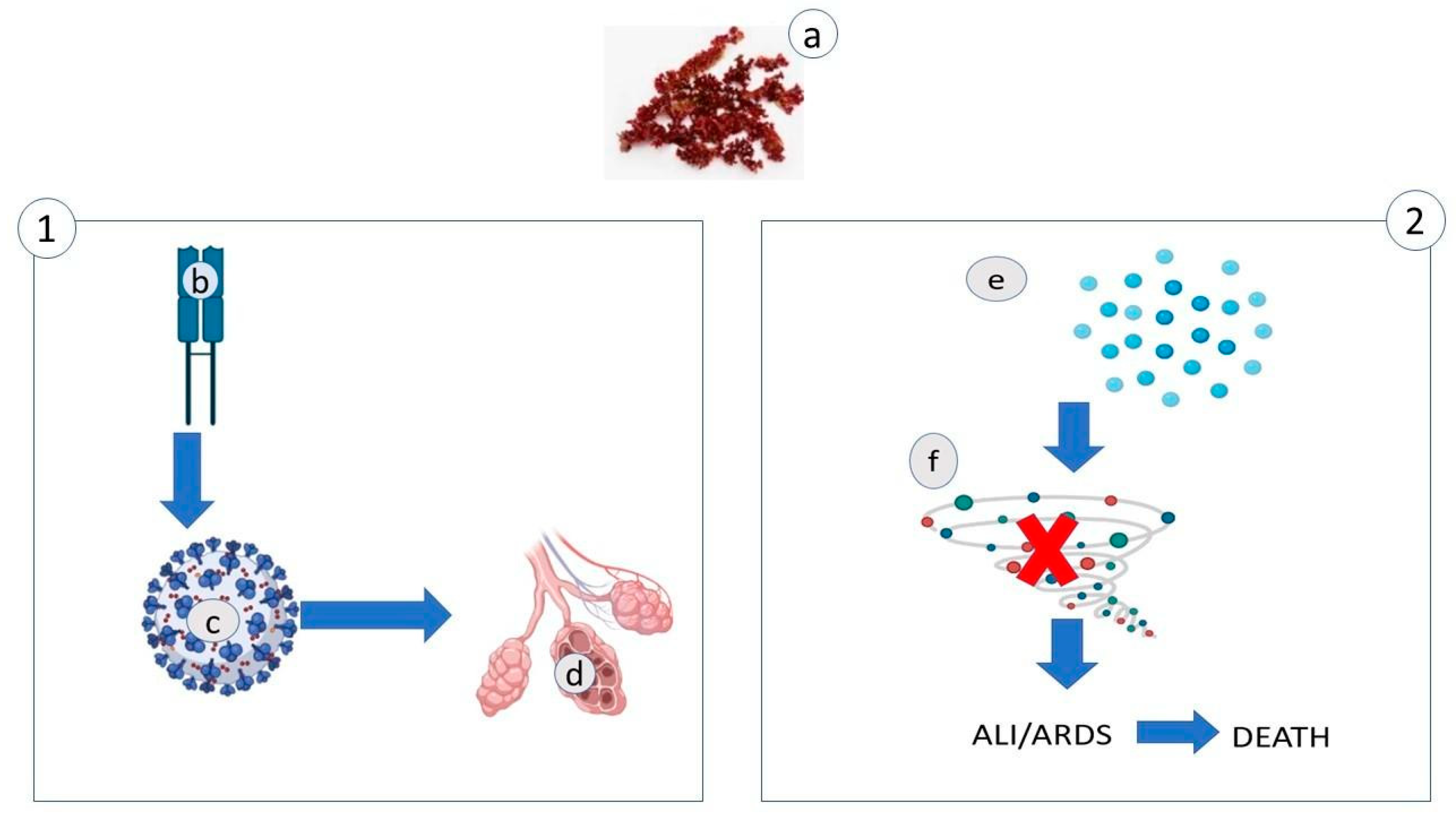
2.2. Polysaccharides
| Polysaccharides Interact with the Positively Charged Domains of the Virus Envelope Glycoprotein and Create a Non-Reversible Complex | |||
| Sulphate Polysaccharides: | |||
| Name | Organism | Virus | References |
| Calcium-spirulan (Ca-SP) | Spirulina | HIV1, HIV2, HSV1, HSV2, HCMV, MuV, IAV | [20,66] |
| Naviculan | Navicula directa | HSV, IAV, HIV | |
| TK-V3 | Spirulina platensis | HSV1, ECTV, VV | [72] |
| / | Arthsospira fusiforme | HSV1, HSV2 | [73] |
| / | Porphyridium sp. | HSV, MuLV, | [72,74,75,77,78,79] |
| / | Porphyridium cruentum | HH3, VV, ASFV, VHSV | [80] |
| / | Cochlodinium polykrikoides | HIV1, HSV, IBV, HPIVs, MuV | [81] |
| KGO3 | Gyrodinium impudium | IAV, EMCV, | [82,83] |
| / | Chlorella autotrophica | VHSV, ASFV | [80] |
| Acid Polysaccharides | |||
| Nostoflan | Nostoc flagelliforme | HSV, IAV | [84,85,86] |
| Chlorella pyrenoidosa | VSV | [88] | |
2.3. Pigments
| Name | Organism | Virus | Action | References |
|---|---|---|---|---|
| Pheophorbide a | Dunaliella Primolecta, Lyngbya | HSV1 | Bonds to virus cell receptors, effects post-entry steps | [89,90,91] |
| Carotenoids extracts | Dunaliella salina | Su-HV1, PRV | Inhibition of plaque formation and downregulation of gene and protein expression | [94,95,96] |
| Astaxanthin | Haematococcus pluvialis | WSSV, IHNV | Antioxidant action | [96,98] |
| Allophycocyanin | Spirulina platentis | EV71 | Delay of viral RNA synthesis in vitro | [99] |
| Phycocyanin | Spirulina platentis | IAV | Downregulation of expression of inflammatory factors | [102] |
| Not identified pigment | Ankistrodesmus convolotus, Spirulina | EBV | Inhibition of some proteins involved in the lytic cycle | [103,104] |
2.4. Others Microalgae Compounds with Antiviral Effects
2.4.1. Peptides and Proteins
2.4.2. Flavonoids and Polyphenols
2.4.3. Glycolipids
| Name | Typology | Organism | Virus | Action | References |
|---|---|---|---|---|---|
| Ichthyopeptin A | Peptide | Microcystis ichthyoblabe | IAV | Inhibition of proteins of virus cycle | [107,108] |
| SM | Peptide | Spirulina maxima | HIV1 |
Inhibition of the reverse transcriptase of the virus and p24 antigen production | [105] |
| Not identified protein | protein | Nannochloropsis oculata | NNV | Increase in α actin activity and immunity system | [109] |
| Not identified protein | protein | Spirulina platensis | NPV | Decrease in mortality | [105] |
| Not identified flavonoids | flavonoid | Geitlerinema sp. | HCV | Reduction in ATPase activity | [111,112] |
| Marennine | polyphenol | Haslea ostrearia |
HSV, HIV |
Inhibition of virus invasion and replication. | [113,114,115,116,117,119] |
| Monogalactosyldiacilglyceride | glycolipid | Coccomixa sp. | HSV2 | Change of virus shape causing lysis. | [122] |
| Sulfoquinovosyldiacyglycerol | glycolipid | Phormidium sp.
Lyngbya sp. | HIV, HSV | Inhibition of RNA polymerase. | [123] |
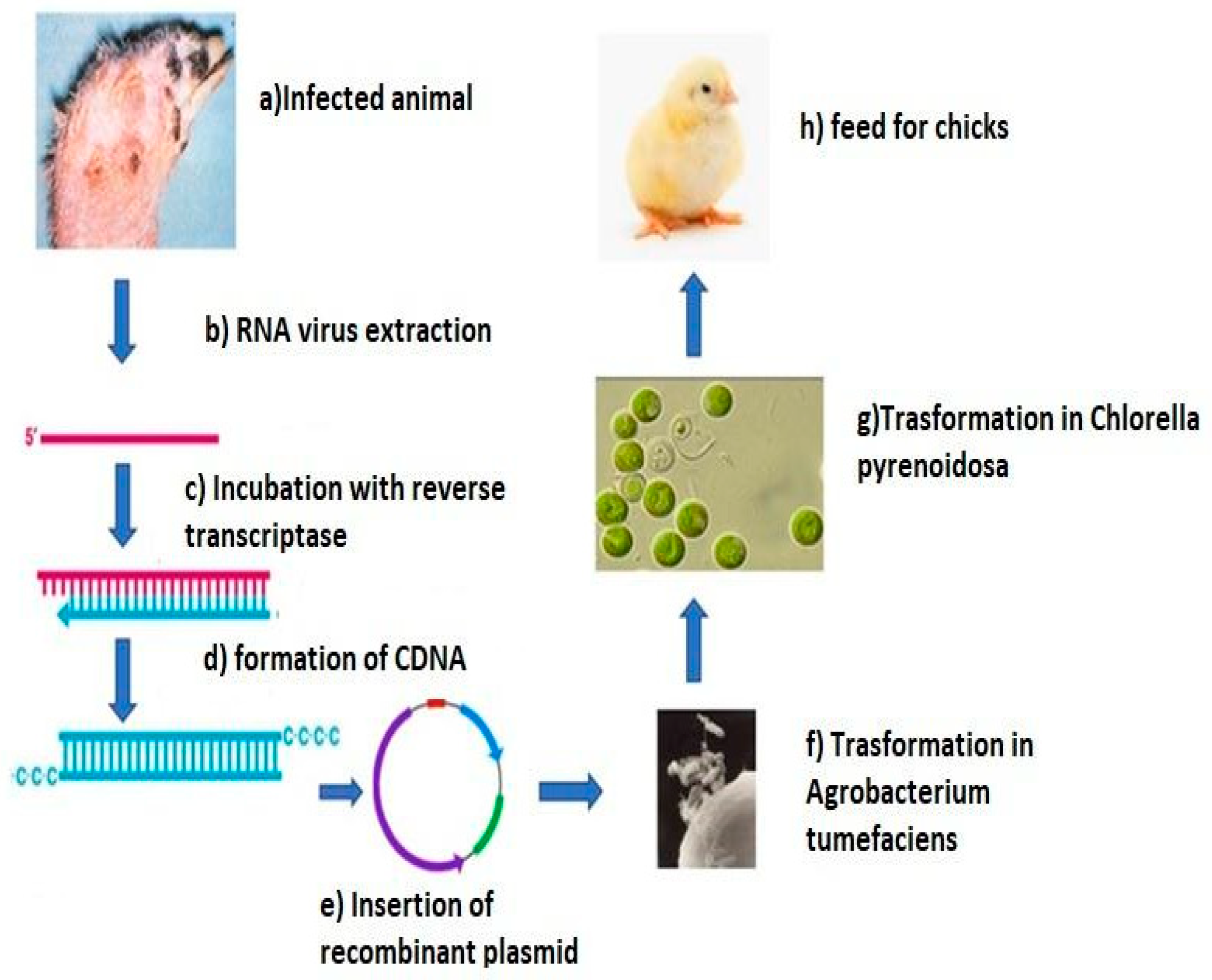
3. Antiviral Bioengineering Perspectives Using Microalgae
| Genetic Engineering | |||
| Microalgae | Uses | Virus | References |
| Phaeodactylum tricornutum | Expression of a recombinant antibody | HBV | [128,129] |
| Chlamydomonas reinhardtii | Expression of RNA interfering | YHV | [131] |
| Chlorella pyrenoidosa | Expression of antigenic protein | IBDV | [132] |
| Schizochytrium sp. | Expression of antigenic protein | ZIKV | [133] |
4. An Overview of the Antiviral Effects of a Supplementary Microalgae Diet and Its Possible Action on SARS-CoV-2
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jassim, S.A.A.; Naji, M.A. Novel antiviral agents: A medicinal plant perspective. J. Appl. Microbiol. 2003, 95, 412–427. [Google Scholar] [CrossRef] [Green Version]
- Wagner, E.K.; Hewlett, M.J. Basic Virology; Blackwell Science: Malden, MA, USA, 2001. [Google Scholar]
- Chen, C.H.; Matthews, T.J.; McDanal, C.B.; Bolognesi, D.P.; Greenberg, M.L. A molecular clasp in the human immunodeficiency virus (HIV) type 1 TM protein determines the anti-HIV activity of gp41 derivatives: Implication for viral fusion. J. Virol. 1995, 69, 3771–3775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Clercq, E. Current lead natural products for the chemotherapy of human immunodeficiency virus (HIV) infection. Med. Res. Rev. 2000, 20, 323–349. [Google Scholar] [CrossRef]
- Drosten, C.; Günther, S.; Preiser, W.; van der Werf, S.; Brodt, H.-R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.M.; et al. Identification of a novel coronavirus in patients with Severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murrell, S.; Wu, S.C.; Butler, M. Review of dengue virus and the development of a vaccine. Biotechnol. Adv. 2011, 29, 239–247. [Google Scholar] [CrossRef]
- Loutfy, M.R.; Wu, W.; Letchumanan, M.; Bondy, L.; Antoniou, T.; Margolese, S.; Zhang, Y.; Rueda, S.; McGee, F.; Peck, R.; et al. Correction: Systematic review of HIV transmission between heterosexual sero-discordant couples where the HIV-positive partner is fully suppressed on antiretroviral therapy. PLoS ONE 2013, 8, e55747. [Google Scholar] [CrossRef]
- Ahmadi, A.; Zorofchian Moghadamtousi, S.; Abubakar, S.; Zandi, K. Antiviral potential of algae polysaccharides isolated from marine sources: A review. BioMed Res. Int. 2015, 30, 1967–1976. [Google Scholar] [CrossRef] [Green Version]
- Yasuhara-Bell, J.; Lu, Y. Marine compounds and their antiviral activities. Antivir. Res. 2010, 86, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K. Recent advances in microalgal bioscience in Japan, with special reference to utilization of biomass and metabolites: A review. J. Appl. Phycol. 1996, 8, 487–502. [Google Scholar] [CrossRef]
- Priyadarshani, I.; Rath, B. Commercial and industrial applications of micro algae: A review. Res. Artic. J. Algal Biomass Util. 2012, 2229–6905. [Google Scholar] [CrossRef]
- Carbone, D.A.; Olivieri, G.; Pollio, A.; Melkonian, M. Biomass and phycobiliprotein production of Galdieria sulphuraria, immobilized on a twin-layer porous substrate photobioreactor. Appl. Microbiol. Biotechnol. 2020, 104, 3109–3119. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. Commercial production of microalgae: Ponds, tanks, and fermenters. Prog. Ind. Microbiol. 1999, 35, 313–321. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Carbone, D.A.; Olivieri, G.; Pollio, A.; Pinto, G.; Melkonian, M. Growth and biomass productivity of Scenedesmus vacuolatus on a twin layer system and a comparison with other types of cultivations. Appl. Microbiol. Biotechnol. 2017, 101, 8321–8329. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, I.; Komiyama, K. Acidic polysaccharide CH-1 isolated from Chlorella pyrenoidosa and the use thereof. U.S. Patent 4533548A, 7 August 1985. [Google Scholar]
- Reshef, V.; Mizrachi, E.; Maretzki, T.; Silberstein, C.; Loya, S.; Hizi, A.; Carmeli, S. New acylated sulfoglycolipids and digalactolipids and related known glycolipids from cyanobacteria with a potential to inhibit the reverse transcriptase of HIV-1. J. Nat. Prod. 1997, 60, 1251–1260. [Google Scholar] [CrossRef]
- Ayehunie, S.; Belay, A.; Baba, T.W.; Ruprecht, R.M. Inhibition of HIV-1 replication by an aqueous extract of Spirulina platensis (Arthrospira platensis). J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1998, 18, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Hayashi, K.; Maeda, M.; Kojima, I. Calcium Spirulan, an inhibitor of enveloped virus replication, from a blue-green alga Spirulina platensis. J. Nat. Prod. 1996, 59, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Huskens, D.; Schols, D. Algal lectins as potential HIV microbicide candidates. Mar. Drugs 2012, 10, 1476–1497. [Google Scholar] [CrossRef] [Green Version]
- Muir, J. Managing to harvest? Perspectives on the potential of aquaculture. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 191–218. [Google Scholar] [CrossRef] [Green Version]
- Bostock, J.; McAndrew, B.; Richards, R.; Jauncey, K.; Telfer, T.; Lorenzen, K.; Little, D.; Ross, L.; Handisyde, N.; Gatward, I.; et al. Aquaculture: Global status and trends. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2897–2912. [Google Scholar] [CrossRef]
- Mitchell, C.A.; Ramessar, K.; O’Keefe, B.R. Antiviral lectins: Selective inhibitors of viral entry. Antivir. Res. 2017, 142, 37–54. [Google Scholar] [CrossRef]
- Botos, I.; Wlodawer, A. Cyanovirin-N: A sugar-binding antiviral protein with a new twist. Cell. Mol. Life Sci. 2003, 60, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Weiss, R.A. How does HIV cause AIDS? Science 1993, 260, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Douek, D.C.; Roederer, M.; Koup, R.A. Emerging concepts in the immunopathogenesis of AIDS. Annu. Rev. Med. 2009, 60, 471–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, C.K.; Spellman, M.W.; Riddle, L.; Harris, R.J.; Thomas, J.N.; Gregory, T.J. Assignment of intrachain disulfide bonds and characterization of potential glycosylation sites of the type 1 recombinant human immunodeficiency virus envelope glycoprotein (gp120) expressed in Chinese hamster ovary cells. J. Biol. Chem. 1990, 265, 10373–10382. [Google Scholar] [CrossRef]
- Kwong, P.D.; Wyatt, R.; Robinson, J.; Sweet, R.W.; Sodroski, J.; Hendrickson, W.A. Structure of an HIV gp 120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature 1998, 393, 648–659. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Murakami, M.; Miyazawa, K.; Hori, K. Purification and characterization of a novel lectin from a freshwater cyanobacterium, Oscillatoria agardhii. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2000, 125, 169–177. [Google Scholar] [CrossRef]
- Sato, Y.; Okuyama, S.; Hori, K. Primary Structure and Carbohydrate Binding Specificity of a Potent Anti-HIV Lectin isolated from the filamentous cyanobacterium Oscillatoria agardhii. J. Biol. Chem. 2007, 282, 11021–11029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Hori, K. Cloning, expression, and characterization of a novel anti-HIV lectin from the cultured cyanobacterium, Oscillatoria agardhii. Fish. Sci. 2009, 75, 743–753. [Google Scholar] [CrossRef]
- Kehr, J.C.; Zilliges, Y.; Springer, A.; Disney, M.D.; Ratner, D.D.; Bouchier, C.; Seeberger, P.H.; de Marsac, N.T.; Dittmann, E. A mannan binding lectin is involved in cell-cell attachment in a toxic strain of Microcystis aeruginosa. Mol. Microbiol. 2006, 59, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Boyd, M.R.; Gustafson, K.R.; McMahon, J.B.; Shoemaker, R.H.; O’Keefe, B.R.; Mori, T.; Gulakowski, R.J.; Wu, L.; Rivera, M.I.; Laurencot, C.M.; et al. Discovery of cyanovirin-N, a novel human immunodeficiency virus inactivating protein that binds viral surface envelope glycoprotein gp120: Potential applications to microbicide development. Antimicrob. Agents Chemother. 1997, 41, 1521–1530. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, K.R.; Sowder, R.C.; Henderson, L.E.; Cardellina, J.H.; McMahon, J.B.; Rajamani, U.; Pannell, L.K.; Boyd, M.R. Isolation, primary sequence determination, and disulfide bond structure of cyanovirin-N, an anti-HIV (Human immunodeficiency virus) protein from the cyanobacterium Nostoc ellipsosporum. Biochem. Biophys. Res. Commun. 1997, 238, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Bewley, C.A.; Louis, J.M.; Gustafson, K.R.; Boyd, M.R.; Gronenborn, A.M.; Clore, G.M.; Wlodawer, A. Crystal structure of cyanovirin-N, a potent HIV-inactivating protein, shows unexpected domain swapping. J. Mol. Biol. 1999, 288, 403–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrientos, L.G.; Louis, J.M.; Botos, I.; Mori, T.; Han, Z.; O’Keefe, B.R.; Boyd, M.R.; Wlodawer, A. Reconciliation of X-ray and NMR structures. Structure 2002, 10, 673–686. [Google Scholar] [CrossRef] [Green Version]
- Férir, G.; Huskens, D.; Noppen, S.; Koharudin, L.M.I.; Gronenborn, A.M.; Schols, D. Broad anti-HIV activity of the Oscillatoria agardhii agglutinin homologue lectin family. J. Antimicrob. Chemother. 2014, 69, 2746–2758. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Chen, W.; Guo, C.; Qian, C.; Liu, G.; Ge, F.; Huang, Y.; Kitazato, K.; Wang, Y.; Xiong, S. Soluble cytoplasmic expression, rapid purification, and characterization of cyanovirin-N as a His-SUMO fusion. Appl. Microbiol. Biotechnol. 2010, 85, 1051–1060. [Google Scholar] [CrossRef]
- Agarwal, R.; Trivedi, J.; Mitra, D. High yield production of recombinant cyanovirin-N (antiviral lectin) exhibiting significant anti-HIV activity, from a rationally selected Escherichia coli strain. Process Biochem. 2020, 93, 1–11. [Google Scholar] [CrossRef]
- Barrientos, L.G.; Gronenborn, A.M. The highly specific carbohydrate-binding protein cyanovirin-N: Structure, anti-HIV/Ebola activity and possibilities for therapy. Mini Rev. Med. Chim. 2005, 5, 220–231. [Google Scholar] [CrossRef]
- Tiwari, V.; Shukla, S.Y.; Shukla, D. A sugar binding protein cyanovirin-N blocks herpes simplex virus type-1 entry and cell fusion. Antivir. Res. 2009, 84, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Kachko, A.; Loesgen, S.; Shahzad-Ul-Hussan, S.; Tan, W.; Zubkova, I.; Takeda, K.; Wells, F.; Rubin, S.; Bewley, C.A.; Major, M.E. Inhibition of Hepatitis C virus by the cyanobacterial protein Microcystis viridis lectin: Mechanistic differences between the high-mannose specific lectins MVL, CV-N, and GNA. Mol. Pharm. 2013, 10, 4590–4602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubuisson, J.; Cosset, F.L. Virology and cell biology of the Hepatitis C virus life cycle. An update. J. Hepatol. 2014, 61, S3–S13. [Google Scholar] [CrossRef] [Green Version]
- Garrison, A.R.; Giomarelli, B.G.; Lear-Rooney, C.M.; Saucedo, C.J.; Yellayi, S.; Krumpe, L.R.H.; Rose, M.; Paragas, J.; Bray, M.; Olinger, G.G.; et al. The cyanobacterial lectin scytovirin displays potent in vitro and in vivo activity against Zaire Ebola virus. Antivir. Res. 2014, 112, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Stumpf, M.P.H.; Laidlaw, Z.; Jansen, V.A.A. Herpes viruses hedge their bets. Proc. Natl. Acad. Sci. USA 2002, 99, 15234–15237. [Google Scholar] [CrossRef] [Green Version]
- Thompson, W.W.; Shay, D.K.; Weintraub, E.; Cox, N.; Anderson, L.J.; Fukuda, K. Mortality associated with influenza and respiratory syncytial virus in the United States. J. Am. Med. Assoc. 2003, 289, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Perrone, L.A.; Plowden, J.K.; García-Sastre, A.; Katz, J.M.; Tumpey, T.M. H5N1 and 1918 Pandemic influenza virus infection. Results in early and excessive infiltration of macrophages and neutrophils in the lungs of mice. PLoS Pathog. 2008, 4, 1000115. [Google Scholar] [CrossRef] [Green Version]
- Zambon, M.C. Epidemiology and pathogenesis of influenza. J. Antimicrob. Chemother. 1999, 44, 3–9. [Google Scholar] [CrossRef]
- Giomarelli, B.; Provvedi, R.; Meacci, F.; Maggi, T.; Medaglini, D.; Pozzi, G.; Mori, T.; McMahon, J.B.; Gardella, R.; Boyd, M.R. The microbicide cyanovirin-N expressed on the surface of commensal bacterium Streptococcus gordonii captures HIV-1. AIDS 2002, 16, 1351–1356. [Google Scholar] [CrossRef]
- Mori, T.; Barrientos, L.G.; Han, Z.; Gronenborn, A.M.; Turpin, J.A.; Boyd, M.R. Functional homologs of cyanovirin-N amenable to mass production in prokaryotic and eukaryotic hosts. Protein Expr. Purif. 2002, 26, 42–49. [Google Scholar] [CrossRef]
- Sexton, A.; Drake, P.M.; Mahmood, N.; Harman, S.J.; Shattock, R.J.; Ma, J.K. Transgenic plant production of cyanovirin-N, an HIV microbicide. FASEB J. 2006, 20, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Colleluori, D.M.; Tien, D.; Kang, F.; Pagliei, T.; Kuss, R.; McCormick, T.; Watson, K.; McFadden, K.; Chaiken, I.; Buckheit, R.W.; et al. Expression, purification, and characterization of recombinant cyanovirin-N for vaginal anti-HIV microbicide development. Protein Expr. Purif. 2005, 39, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lagenaur, L.A.; Simpson, D.A.; Essenmacher, K.P.; Frazier-Parker, C.L.; Liu, Y.; Tsai, D.; Rao, S.S.; Hamer, D.H.; Parks, T.P.; et al. Engineered vaginal lactobacillus strain for mucosal delivery of the human immunodeficiency virus inhibitor cyanovirin-N. Antimicrob. Agents Chemother. 2006, 50, 3250–3259. [Google Scholar] [CrossRef] [Green Version]
- Zilliges, Y.; Kehr, J.C.; Mikkat, S.; Bouchier, C.; De Marsac, N.T.; Börner, T.; Dittmann, E. An extracellular glycoprotein is implicated in cell-cell contacts in the toxic cyanobacterium Microcystis aeruginosa PCC 7806. J. Bacteriol. 2008, 190, 2871–2879. [Google Scholar] [CrossRef] [Green Version]
- Botos, I.; O’Keefe, B.R.; Shenoy, S.R.; Cartner, L.K.; Ratner, D.M.; Seeberger, P.H.; Boyd, M.R.; Wlodawer, A. Structures of the complexes of a potent anti-HIV protein cyanovirin-N and high mannose oligosaccharides. J. Biol. Chem. 2002, 277, 34336–34342. [Google Scholar] [CrossRef] [Green Version]
- Huskens, D.; Férir, G.; Vermeire, K.; Kehr, J.C.; Balzarini, J.; Dittmann, E.; Schols, D. Microvirin, a novel α (1,2)-mannose-specific lectin isolated from Microcystis aeruginosa, has anti-HIV-1 activity comparable with that of cyanovirin-N but a much higher safety profile. J. Biol. Chem. 2010, 285, 24845–24854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, M.; Ogawa, T.; Muramoto, K.; Kamio, Y.; Jimbo, M.; Kamiya, H. Isolation and characterization of a mannan-binding lectin from the freshwater cyanobacterium (blue-green algae) Microcystis viridis. Biochem. Biophys. Res. Commun. 1999, 265, 703–708. [Google Scholar] [CrossRef]
- Shahzad-ul-Hussan, S.; Gustchina, E.; Ghirlando, R.; Clore, G.M.; Bewley, C.A. Solution structure of the monovalent lectin microvirin in complex with Man α (1-2) Man provides a basis for anti-HIV activity with low toxicity. J. Biol. Chem. 2011, 286, 20788–20796. [Google Scholar] [CrossRef] [Green Version]
- Bokesch, H.R.; O’Keefe, B.R.; McKee, T.C.; Pannell, L.K.; Patterson, G.M.L.; Gardella, R.S.; Sowder, R.C.; Turpin, J.; Watson, K.; Buckheit, R.W. A potent novel anti-HIV protein from the cultured cyanobacterium Scytonema varium. Biochemistry 2003, 42, 2578–2584. [Google Scholar] [CrossRef] [PubMed]
- Wood, A. Synthesis of a new scytovirin protein derivative, Sd1-Sd2-Sd2, to decrease Hiv-1 binding affinity. Ph.D. Thesis, University of Mississippi, Oxford, MS, USA, 2013. [Google Scholar]
- Takebe, Y.; Saucedo, C.J.; Lund, G.; Uenishi, R.; Hase, S.; Tsuchiura, T.; Kneteman, N.; Ramessar, K.; Tyrrell, D.L.J.; Shirakura, M.; et al. Antiviral lectins from red and blue-green algae show potent in vitro and in vivo activity against Hepatitis C virus. PLoS ONE 2013, 8, e64449. [Google Scholar] [CrossRef] [Green Version]
- Breitenbach, L.C.; dos Santos, M.; de Olivieria, W.; de Moura, E. Lectin as antimicrobial agents. J. Appl. Microbiol. 2018, 125, 5. [Google Scholar] [CrossRef] [Green Version]
- Neyts, J.; Snoeck, R.; Schols, D.; Balzarini, J.; Esko, J.D.; Van Schepdael, A.; De Clercq, E. Sulfated polymers inhibit the interaction of human cytomegalovirus with cell surface heparan sulfate. Virology 1992, 189, 48–58. [Google Scholar] [CrossRef]
- Lüscher-Mattli, M. Polyanions. A lost chance in the fight against HIV and other virus diseases? Antivir. Chem. Chemother. 2000, 11, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.B.; Srisomporn, P.; Hayashi, K.; Tanaka, T.; Sankawa, U.; Hayashi, T. Effects of structural modification of calcium spirulan, a sulfated polysaccharide from Spirulina Platensis, on antiviral activity. Chem. Pharm. Bull. 2001, 49, 108–110. [Google Scholar] [CrossRef] [Green Version]
- Damonte, E.; Matulewicz, M.; Cerezo, A. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2012, 11, 2399–2419. [Google Scholar] [CrossRef] [PubMed]
- Harden, E.A.; Falshaw, R.; Carnachan, S.M.; Kern, E.R.; Prichard, M.N. Virucidal activity of polysaccharide extracts from four algal species against herpes simplex virus. Antivir. Res. 2009, 83, 282–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, M.; Schols, D.; Pauwels, R.; Nakashima, H.; De Clercq, E. Sulfated polysaccharides as potent inhibitors of HIV-induced syncytium formation: A new strategy towards AIDS chemotherapy. J. Acquir. Immune Defic. Syndr. 1990, 3, 493–499. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E.; Herdewijn, P. Strategies in the design of antiviral drugs. In Pharmaceutical Sciences Encyclopedia; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 1–56. [Google Scholar]
- Huleihel, M.; Ishanu, V.; Tal, J.; Arad, S. Antiviral effect of red microalgal polysaccharides on Herpes simplex and Varicella zoster viruses. J. Appl. Phycol. 2001, 13, 127–134. [Google Scholar] [CrossRef]
- Radonic, L.M.; Zimmermann, J.M.; Zavallo, D.; López, N.; López Bilbao, M. Rooting in Km selective media as efficient in vitro selection method for sunflower genetic transformation. Electron. J. Biotechnol. 2006, 9, 712–713. [Google Scholar] [CrossRef] [Green Version]
- Sharaf, M.; Amara, A.; Aboul-Enein, A.; Helmi, S.; Ballot, A.; Astani, A.; Schnitzler, P. Molecular authentication and characterization of the antiherpetic activity of the cyanobacterium Arthrospira fusiformis. Pharmazie 2010, 65, 132–136. [Google Scholar] [CrossRef]
- Malis Arad, S.; Adda, M.; Cohen, E. The potential of production of sulfated polysaccharides from Porphyridium. Plant Soil 1985, 89, 117–127. [Google Scholar] [CrossRef]
- Sun, L.; Wang, C.; Shi, Q.; Ma, C. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef]
- Gardeva, E.; Toshkova, R.; Yossifova, L.; Minkova, K.; Gigova, L. Cytotoxic and apoptogenic potential of red microalgal polysaccharides. Biotechnol. Biotechnol. Equip. 2012, 26, 3167–3172. [Google Scholar] [CrossRef]
- Kavitha, M.D.; Kathiresan, S.; Bhattacharya, S.; Sarada, R. Culture media optimization of Porphyridium purpureum: Production potential of biomass, total lipids, arachidonic and eicosapentaenoic acid. J. Food Sci. Technol. 2016, 53, 2270–2278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardeva, E.; Toshkova, R.; Minkova, K.; Gigova, L. Cancer protective action of polysaccharide, derived from red microalga porphyridium cruentum: A biological background. Biotechnol. Biotechnol. Equip. 2009, 23, 783–787. [Google Scholar] [CrossRef]
- Talyshinsky, M.M.; Souprun, Y.Y.; Huleihel, M.M. Anti-viral activity of red microalgal polysaccharides against retroviruses. Cancer Cell Int. 2002, 2, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabregas, J.; García, D.; Fernandez-Alonso, M.; Rocha, A.I.; Gómez-Puertas, P.; Escribano, J.M.; Otero, A.; Coll, J.M. In vitro inhibition of the replication of Haemorrhagic septicaemia virus (VHSV) and African swine fever virus (ASFV) by extracts from marine microalgae. Antivir. Res. 1999, 44, 67–73. [Google Scholar] [CrossRef]
- Yim, J.H.; Kim, S.J.; Ahn, S.H.; Lee, C.K.; Rhie, K.T.; Lee, H.K. Antiviral effects of sulfated exopolysaccharide from the marine microalga Gyrodinium impudicum strain KG03. Mar. Biotechnol. 2004, 6, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Hasui, M.; Matsuda, M.; Okutani, K.; Shigeta, S. In vitro antiviral activities of sulfated polysaccharides from a marine microalga (Cochlodinium polykrikoides) against human immunodeficiency virus and other enveloped viruses. Int. J. Biol. Macromol. 1995, 17, 293–297. [Google Scholar] [CrossRef]
- Schmidt, A.C.; Schaap-Nutt, A.; Bartlett, E.J.; Schomacker, H.; Boonyaratanakornkit, J.; Karron, R.A.; Collins, P.L. Progress in the development of Human parainfluenza virus vaccines. Expert Rev. Respir. Med. 2011, 5, 515–526. [Google Scholar] [CrossRef]
- Ligas, M.W.; Johnson, D.C. A herpes simplex virus mutant in which glycoprotein D sequences are replaced by beta-galactosidase sequences binds to but is unable to penetrate into cells. J. Virol. 1988, 62, 1486–1494. [Google Scholar] [CrossRef] [Green Version]
- Kanekiyo, K.; Hayashi, K.; Takenaka, H.; Lee, J.-B.; Hayashi, T. Anti-Herpes simplex virus target of an acidic polysaccharide, nostoflan, from the edible blue-green alga Nostoc flagelliforme. Biol. Pharm. Bull. 2007, 30, 1573–1575. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T. Studies on evaluation of natural products for antiviral effects and their applications. Yakugaku Zasshi 2008, 128, 61–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, K. A natural sulfated polysaccharide, calcium spirulan, isolated from Spirulina platensis: In vitro and ex vivo evaluation of anti-Herpes simplex virus and anti-Human immunodeficiency virus activities. AIDS Res. Hum. Retrovir. 1996, 12, 1463–1471. [Google Scholar] [CrossRef] [Green Version]
- Plaza, M.; Herrero, H.; Cifuentas, A. Innovative natural functional ingredients from microalgae. J. Agric. 2009, 7159–7170. [Google Scholar] [CrossRef] [PubMed]
- Xodo, L.; Rapozzi, V.; Zacchigna, M.; Drioli, S.; Zorzet, S. The chlorophyll catabolite pheophorbide a as a photosensitizer for the photodynamic therapy. Curr. Med. Chem. 2012, 19, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Murakami, A.; Koshimizu, K.; Ohigashi, H. Identification of pheophorbide a and its related compounds as possible anti-tumor promoters in the leaves of Neptunia oleracea. Biosci. Biotechnol. Biochem. 1996, 60, 1028–1030. [Google Scholar] [CrossRef] [PubMed]
- Saide, A.; Lauritano, C.; Ianora, A. Pheophorbide a: State of the art. Mar. Drugs 2020, 18, 257. [Google Scholar] [CrossRef] [PubMed]
- Bouslama, L.; Hayashi, K.; Lee, J.B.; Ghorbel, A.; Hayashi, T. Potent virucidal effect of pheophorbide a and pyropheophorbide a on enveloped viruses. J. Nat. Med. 2011, 65, 229–233. [Google Scholar] [CrossRef]
- Ratnoglik, S.L.; Aoki, C.; Sudarmono, P.; Komoto, M.; Deng, L.; Shoji, I.; Fuchino, H.; Kawahara, N.; Hotta, H. Antiviral activity of extracts from Morinda citrifolia leaves and chlorophyll catabolites, pheophorbide a and pyropheophorbide a, against Hepatitis C virus. Microbiol. Immunol. 2014, 58, 188–194. [Google Scholar] [CrossRef]
- Ohta, S.; Ono, F.; Shiomi, Y.; Nakao, T.; Aozasa, O.; Nagate, T.; Kitamura, K.; Yamaguchi, S.; Nishi, M.; Miyata, H. Anti-Herpes simplex virus substances produced by the marine green alga Dunaliella primolecta. J. Appl. Phycol. 1998, 10, 349–356. [Google Scholar] [CrossRef]
- Concise Review of Veterinary Virology. Available online: https://www.libyanvet.com/concisereviewofveterinaryvirology.htm (accessed on 19 April 2021).
- Talukdar, J.; Dasgupta, S.; Nagle, V.; Bhadra, B. COVID-19: Potential of microalgae derived natural astaxanthin as adjunctive supplement in alleviating cytokine storm. SSRN Electron. J. 2020. [Google Scholar] [CrossRef]
- Li, W.; Jia, X.; Shen, C.; Zhang, M.; Xu, J.; Shang, Y.; Zhu, K.; Hu, M.; Yan, Q.; Qin, D.; et al. A KSHV microRNA enhances viral latency and induces angiogenesis by targeting GRK2 to activate the CXCR2/AKT pathway. Oncotarget 2016, 7, 32286–32305. [Google Scholar] [CrossRef]
- Wang, H.; Dai, A.; Liu, F.; Guan, Y. Effects of dietary astaxanthin on the immune response, resistance to White spot syndrome virus and transcription of antioxidant enzyme genes in Pacific white shrimp Litopenaeus vannamei. Iran. J. Fish. Sci. 2015, 14, 699–718. [Google Scholar] [CrossRef]
- Shih, S.-R.; Tsai, K.-N.; Li, Y.-S.; Chueh, C.-C.; Chan, E.-C. Inhibition of Enterovirus 71-induced apoptosis by allophycocyanin isolated from a blue-green alga spirulina platensis. J. Med. Virol. 2003, 70, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Geiler, J.; Michaelis, M.; Naczk, P.; Leutz, A.; Langer, K.; Doerr, H.W.; Cinatl, J. N-acetyl-l-cysteine (NAC) inhibits virus replication and expression of pro-inflammatory molecules in A549 cells infected with highly pathogenic H5N1 Influenza A virus. Biochem. Pharmacol. 2010, 79, 413–420. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Qiao, H.; Liu, T.; Yang, Z.; Xu, L.; Xu, Y.; Ge, H.M.; Tan, R.X.; Li, E. Inhibition of Herpes simplex virus infection by oligomeric stilbenoids through ROS generation. Antivir. Res. 2012, 95, 30–36. [Google Scholar] [CrossRef]
- Chen, Y.H.; Chang, G.K.; Kuo, S.M.; Huang, S.Y.; Hu, I.C.; Lo, Y.L.; Shih, S.R. Well-tolerated Spirulina extract inhibits influenza virus replication and reduces virus-induced mortality. Sci. Rep. 2016, 6, 24253. [Google Scholar] [CrossRef] [Green Version]
- Kok, Y.Y.; Chu, W.L.; Phang, S.M.; Mohamed, S.M.; Naidu, R.; Lai, P.J.; Ling, S.N.; Mak, J.W.; Lim, P.K.C.; Balraj, P.; et al. Inhibitory activities of microalgal extracts against Epstein-Barr virus DNA release from lymphoblastoid cells. J. Zhejiang Univ. Sci. B 2011, 12, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Kok, Y.; Chu, W.; Lai, P.; Tan, B.K.; Ling, S.N.; Shar, M.; Mak, J.W.; Lim, K.C.P.; Rakesh, N.; Pauline, B.; et al. Antiviral activity of microalgae extracts against Epstein-Barr virus in lymphoblastoid cell lines. In Proceedings of the 6th Asia-Pacific Conference on Algal Biotechnology, Makati City, Philippines, 12–15 October 2006. [Google Scholar]
- Jang, I.S.; Park, S.J. A Spirulina maxima-derived peptide inhibits HIV-1 infection in a human T cell line MT4. Fish. Aquat. Sci. 2016, 19, 37. [Google Scholar] [CrossRef] [Green Version]
- Andavan, G.S.B.; Lemmens-Gruber, R. Cyclodepsipeptides from marine sponges: Natural agents for drug research. Mar. Drugs 2010, 8, 810–834. [Google Scholar] [CrossRef] [Green Version]
- Zainuddin, E.N.; Mundt, S.; Wegner, U.; Mentel, R. Cyanobacteria: A potential source of antiviral substances against Influenza virus. Med. Microbiol. Immunol. 2002, 191, 181–182. [Google Scholar] [CrossRef] [PubMed]
- Zainuddin, E.N.; Mentel, R.; Wray, V.; Jansen, R.; Nimtz, M.; Lalk, M.; Mundt, S. Cyclic depsipeptides, ichthyopeptins A and B, from Microcystis ichthyoblabe. J. Nat. Prod. 2007, 70, 1084–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanuhar, U.; Khumaidi, A. The application of pigment-protein fraction from Nannochloropsis oculata on β-actin response of Cromileptes altivelis infected with viral nervous necrosis. J. Akuakultur Indones. 2017, 16, 22. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, A.D. Pharmacological activities of flavonoids: A review. Int. J. Pharm. Sci. Nanotechnol. 2011, 4, 1394–1398. [Google Scholar] [CrossRef]
- Zaenal Mustopa, A.; Umami, R.N.; Putri, P.H.; Susilaningsih, D.; Farida, H. Identification of bioactive compound from microalga BTM 11 as Hepatitis C virus RNA helicase inhibitor. J. Biol. Indones. 2016, 11, 810–834. [Google Scholar]
- Zaenal Mustopa, A.; Ridwan Lembaga Bahasa Lia, M.; Sukmarini, L. Purification and characterization of polysaccharide from microalgae btm 11 as inhibitor of Hepatitis c virus RNA helicase. Indones. J. Pharm. 2015, 49, 108–110. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Mouget, J.L.; Rosa, P.; Vachoux, C.; Tremblin, G. Enhancement of marennine production by blue light in the diatom Haslea ostrearia. J. Appl. Phycol. 2005, 17, 437–445. [Google Scholar] [CrossRef]
- Gastineau, R.; Turcotte, F.; Pouvreau, J.-B.; Morançais, M.; Fleurence, J.; Windarto, E.; Prasetiya, F.; Arsad, S.; Jaouen, P.; Babin, M.; et al. Marennine, Promising Blue Pigments from a Widespread Haslea Diatom Species Complex. Mar. Drugs 2014, 12, 3161–3189. [Google Scholar] [CrossRef] [Green Version]
- Gastineau, R.; Pouvreau, J.B.; Hellio, C.; Morançais, M.; Fleurence, J.; Gaudin, P.; Bourgougnon, N.; Mouget, J.L. Biological activities of purified marennine, the blue pigment responsible for the greening of oysters. J. Agric. Food Chem. 2012, 60, 3599–3605. [Google Scholar] [CrossRef]
- Vandanjon, L. Etude d’un procédé de valorisation d’une microalgue marine: Concentration et purification par techniques à membranes d’un pigment naturel produit par la diatomée Hhaslea ostrearia. Ph.D. Thesis, University of Nantes, Nantes, France, 1997. [Google Scholar]
- Chentir, H.; Hamdi, H.; Doumandji, A.; Sadok, H.; Ouada, M. Enhancement of extracellular polymeric substances (EPS) production in Spirulina by two step cultivation process and partial characterization of their polysaccharides moiety. Int. J. Biol. Macromol. 2017, 72, 197–206. [Google Scholar] [CrossRef]
- Olicard, C.; Renault, T.; Torhy, C.; Benmansour, A.; Bourgougnon, N. Putative antiviral activity in hemolymph from adult Pacific oysters, Crassostrea gigas. Antivir. Res. 2005, 66, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Satoh, A.; Kato, M.; Yamato, K.; Ishibashi, M.; Sekiguchi, H.; Kurano, N.; Miyachi, S. Characterization of the lipid accumulation in a new microalgal species, Pseudochoricystis ellipsoidea (trebouxiophyceae). J. Jpn. Inst. Energy 2010, 89, 909–913. [Google Scholar] [CrossRef] [Green Version]
- Inès, M.; Dhouha, G. Glycolipid biosurfactants: Potential related biomedical and biotechnological applications. Carbohydr. Res. 2015, 416, 59–69. [Google Scholar] [CrossRef]
- Loya, S.; Reshef, V.; Mizrachi, E.; Silberstein, C.; Rachamim, Y.; Carmeli, S.; Hizi, A. The inhibition of the reverse transcriptase of HIV-1 by the natural sulfoglycolipids from cyanobacteria: Contribution of different moieties to their high potency. J. Nat. Prod. 1998, 61, 891–895. [Google Scholar] [CrossRef]
- Matsumoto, K.; Sakai, H.; Takeuchi, R.; Tsuchiya, K.; Ohta, K.; Sugawara, F.; Abe, M.; Sakaguchi, K. Effective form of sulfoquinovosyldiacyglycerol (SQDG) vesicles for DNA polymerase inhibition. Colloids Surf. B Biointerfaces 2005, 46, 175–181. [Google Scholar] [CrossRef]
- Chirasuwan, N.; Chaiklahan, R.; Ruengjitchatchawalya, M.; Bunnag, B.; Tanticharoen, M. Anti HSV-1 Activity of Spirulina platensis polysaccharide. Kasetsart J. Nat. Sci. 2007, 41, 311–318. [Google Scholar]
- Mayfield, S.; Golden, S.S. Photosynthetic bio-manufacturing:fFood, fuel, and medicine for the 21st century. Photosynth. Res. 2015, 123, 225–226. [Google Scholar] [CrossRef] [Green Version]
- Gimpel, J.A.; Henríquez, V.; Mayfield, S.P. In metabolic engineering of eukaryotic microalgae potential and challenges come with great diversity. Front. Microbiol. 2015, 6, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Guihéneuf, F.; Khan, A.; Tran, L.S.P. Genetic engineering: A promising tool to engender physiological, biochemical, and molecular stress resilience in green microalgae. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Hempel, F.; Maier, U.G. An engineered diatom acting like a plasma cell secreting human IgG antibodies with high efficiency. Microb. Cell Factories 2012, 11, 126. [Google Scholar] [CrossRef] [Green Version]
- Hempel, F.; Lau, J.; Klingl, A.; Maier, U.G. Algae as protein factories: Expression of a human antibody and the respective antigen in the diatom Phaeodactylum tricornutum. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.T.; Islam, M.J.; Parihar, A.; Islam, R.; Jerin, T.J.; Dhote, R.; Ali, M.A.; Laura, F.K.; Halim, M.A. Immunoinformatics and molecular modeling approach to design universal multi-epitope vaccine for SARS-CoV-2. Inform. Med. Unlocked 2021, 24, 100578. [Google Scholar] [CrossRef]
- Somchai, P.; Jitrakorn, S.; Thitamadee, S.; Meetam, M.; Saksmerprome, V. Use of microalgae Chlamydomonas reinhardtii for production of double-stranded RNA against Shrimp virus. Aquac. Rep. 2016, 3, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.H.; Johnson, A.M.A.; Kumar, J.K.; Naveen, T.; Devi, M.C. Heterologous expression of Infectious bursal disease virus VP2 gene in Chlorella pyrenoidosa as a model system for molecular farming. Plant CellTissue Organ Cult. 2017, 131, 119–126. [Google Scholar] [CrossRef]
- Márquez-Escobar, V.A.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Expression of a Zika virus antigen in microalgae: Towards mucosal vaccine development. J. Biotechnol. 2018, 282, 86–91. [Google Scholar] [CrossRef]
- Soni, R.A.; Sudhakar, K.; Rana, R.S. Spirulina from growth to nutritional product: A review. Trends Food Sci. Technol. 2017, 69, 157–171. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.H.; Balachandran, P.; Christensen, O.; Pugh, N.D.; Tamta, H.; Sufka, K.J.; Wu, X.; Walsted, A.; Schjørring-Thyssen, M.; Enevold, C.; et al. Enhancement of natural killer cell activity in healthy subjects by Immulina®, a Spirulina extract enriched for Braun-type lipoproteins. Planta Med. 2010, 76, 1802–1808. [Google Scholar] [CrossRef] [Green Version]
- Appel, K.; Munoz, E.; Navarrete, C.; Cruz-Teno, C.; Biller, A.; Thiemann, E. Immunomodulatory and inhibitory effect of Immulina®, and Immunloges® in the Ig-E mediated activation of RBL-2H3 Cells. A new role in allergic inflammatory responses. Plants 2018, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Kefayat, A.; Ghahremani, F.; Safavi, A.; Hajiaghababa, A.; Moshtaghian, J. Spirulina extract enriched for Braun-type lipoprotein (Immulina®) for inhibition of 4T1 breast tumors’ growth and metastasis. Phytother. Res. 2020, 34, 368–378. [Google Scholar] [CrossRef]
- Pugh, N.; Ross, S.A.; Eisohly, H.N.; Eisohly, M.A.; Pasco, D.S. Isolation of three high molecular weight polysaccharide preparations with potent immunostimulatory activity from Spirulina platensis, Aphanizomenon flos-aquae and Chlorella pyrenoidosa. Planta Med. 2001, 67, 737–742. [Google Scholar] [CrossRef] [Green Version]
- Teas, J.; Hebert, J.R.; Fitton, J.H.; Zimba, P.V. Algae: A poor man’s HAART? Med Hypotheses 2004, 62, 507–510. [Google Scholar] [CrossRef]
- Ngo-Matip, M.-E.; Pieme, A.; Azabji-Kenfack, M.; Biapa, N.; Germaine, N.; Heike, E.; Moukette Moukette, B.; Emmanuel, K.; Philippe, S.; Mbofung, C.M.; et al. Effects of Spirulina platensis supplementation on lipid profile in HIV-infected antiretroviral naïve patients in Yaounde-Cameroon: A randomized trial study. Lipids Health Dis. 2011, 13, 191. [Google Scholar] [CrossRef] [Green Version]
- Riss, J.; Décordé, K.; Sutra, T.; Delage, M.; Baccou, J.C.; Jouy, N.; Brune, J.P.; Oréal, H.; Cristol, J.P.; Rouanet, J.M. Phycobiliprotein C-phycocyanin from Spirulina platensis is powerfully responsible for reducing oxidative stress and NADPH oxidase expression induced by an atherogenic diet in hamsters. J. Agric. Food Chem. 2007, 55, 7962–7967. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, Y.J.; Ryu, H.K.; Kim, M.H.; Chung, H.W.; Kim, W.Y. A randomized double-blind, placebo-controlled study to establish the effects of Spirulina in elderly Koreans. Ann. Nutr. Metab. 2008, 52, 322–328. [Google Scholar] [CrossRef]
- Siqueira, A.S.; Lima, A.R.J.; de Souza, R.C.; Santos, A.S.; Vianez Júnior, J.L.S.G.; Gonçalves, E.C. In silico analysis of the cyanobacterial lectin scytovirin: New insights into binding properties. Mol. Biol. Rep. 2017, 44, 353–358. [Google Scholar] [CrossRef]
- Bhat, V.B.; Madyastha, K.M. Scavenging of peroxynitrite by phycocyanin and phycocyanobilin from Spirulina platensis: Protection against oxidative damage to DNA. Biochem. Biophys. Res. Commun. 2001, 285, 262–266. [Google Scholar] [CrossRef]
- Belay, A. The potential application of Spirulina (Arthrospira) as a nutritional and therapeutic supplement in health management. J. Am. Nutraceutical Assoc. 2002, 5, 27–48. [Google Scholar]
- Simpore, J.; Zongo, F.; Kabore, F.; Dansou, D.; Bere, A.; Nikiema, J.-B.; Pignatelli, S.; Biondi, D.M.; Ruberto, G.; Musumeci, S. Nutrition rehabilitation of HIV-infected and HIV-negative undernourished children utilizing Spirulina. Ann. Nutr. Metab. 2005, 49, 373–380. [Google Scholar] [CrossRef]
- Yakoot, M.; Salem, A. Spirulina platensis versus silymarin in the treatment of chronic Hepatitis C virus infection. A pilot randomized, comparative clinical trial. BMC Gastroenterol. 2012, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C.; Grimble, R.F. Polyunsaturated fatty acids, inflammation and immunity. Eur. J. Clin. Nutr. 2002, 56, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Forster, P.; Forster, L.; Renfrew, C.; Forster, M. Phylogenetic network analysis of SARS-CoV-2 genomes. Proc. Natl. Acad. Sci. USA 2020, 117, 9241–9243. [Google Scholar] [CrossRef] [Green Version]
- Thormar, H.; Isaacs, C.E.; Brown, H.R.; Barshatzky, M.R.; Pessolano, T. Inactivation of enveloped viruses and killing of cells by fatty acids and monoglycerides. Antimicrob. Agents Chemother. 1987, 31, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Subhash, G.V.; Raja, G.; Kumar, K.; Sapre, A.; Dasgupta, S. Possible Prevention of Covid 19 By Usinglinoleic Acid (C18) Rich Algae Oil; AIJR Preprints; 2020; Available online: https://preprints.aijr.org/index.php/ap/preprint/view/36 (accessed on 19 April 2021).
- Maria De Andrade, L.; De Andrade, C.J.; Dias, M.; Nascimento, C.A.O. Chlorella and Spirulina: Microalgae as sources of functional foods, nutraceuticals, and food supplements; an overview. Food Process. Technol. 2018, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Kate, B.N.; Banecjee, U.C. Bioactive compounds from cyanobacteria and microalgae: An overview. Crit. Rev. Biotechnol. 2005, 25, 73–95. [Google Scholar] [CrossRef]
- Choi, H.D.; Kim, J.H.; Chang, M.J.; Kyu-Youn, Y.; Shin, W.G. Effects of astaxanthin on oxidative stress in overweight and obese adults. Phytother. Res. 2011, 25, 1813–1818. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Chyun, J.H.; Kim, Y.K.; Line, L.L.; Chew, B.P. Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutr. Metab. 2010, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, K.S.; Hung, I.F.; Chan, P.P.; Lung, K.; Tso, E.; Liu, R.; Ng, Y.; Chu, M.Y.; Chung, T.W.; Tam, A.R. Gastrointestinal manifestations of SARS-CoV-2 infection and virus load in fecal samples from the Hong Kong cohort and systematic review and meta-analysis. Gastroenterology 2020, 159, 81–95. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.; Yeoh, Y.K.; Li, A.Y.; Zhan, H.; Wan, Y.; Chung, A.; Cheung, C.P.; Chen, N. Alterations in Gut Microbiota of patients with COVID-19 during time of hospitalization. Gastroenterology 2020, 159, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Xu, J.L.; Wang, Z. Microalgae Biotechnology for Food, Health and High Values Products; Springer: Singapore, 2020. [Google Scholar]
- Fields, F.J.; Lejzerowicz, F.; Schroeder, D.; Ngoi, S.M.; Tran, M.; McDonald, D.; Jiang, L.; Chang, J.T.; Knight, R.; Mayfield, S. Effects of the microalgae Chlamydomonas on gastrointestinal health. J. Funct. Foods 2020, 65, 103738. [Google Scholar] [CrossRef]
- Chen, X.; Han, W.; Wang, G.; Zhao, X. Application prospect of polysaccharides in the development of anti-novel coronavirus drugs and vaccines. Int. J. Biol. Macromoules 2020, 164, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Critchley, A.T. The COVID 19 novel coronavirus pandemic 2020: Seaweeds to the rescue? Why does substantial, supporting research about the antiviral properties of seaweed polysaccharides seem to go unrecognized by the pharmaceutical community in these desperate times? J. Appl. Phycol. 2020, 32, 1875–1877. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Parra-Saldivar, R.; Bilal, M.; Afroze, C.A.; Ahmed, M.N.; Iqbal, H.M.N.; Xu, J. Algae-derived bioactive molecules for the potential treatment of sars-cov-2. Molecules 2021, 26, 2134. [Google Scholar] [CrossRef] [PubMed]
- Koide, S.S. Chitin-chitosan: Properties, benefits and risks. Nutr. Res. 1998, 18, 1091–1101. [Google Scholar] [CrossRef]
- Raja, R.; Hemaiswarya, S. Microalgae and immune potential. In Dietary Components and Immune Function; Watson, R.R., Zibadi, S., Preedy, V.R., Eds.; Humana Press, Springer Science + Business Media: New York, NY, USA, 2010; pp. 515–527. [Google Scholar] [CrossRef]
- Rechter, S.; König, T.; Auerochs, S.; Thulke, S.; Walter, H.; Dörnenburg, H.; Walter, C.; Marschall, M. Antiviral activity of Arthrospira-derived spirulan-like substances. Antivir. Res. 2006, 72, 197–206. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carbone, D.A.; Pellone, P.; Lubritto, C.; Ciniglia, C. Evaluation of Microalgae Antiviral Activity and Their Bioactive Compounds. Antibiotics 2021, 10, 746. https://doi.org/10.3390/antibiotics10060746
Carbone DA, Pellone P, Lubritto C, Ciniglia C. Evaluation of Microalgae Antiviral Activity and Their Bioactive Compounds. Antibiotics. 2021; 10(6):746. https://doi.org/10.3390/antibiotics10060746
Chicago/Turabian StyleCarbone, Dora Allegra, Paola Pellone, Carmine Lubritto, and Claudia Ciniglia. 2021. "Evaluation of Microalgae Antiviral Activity and Their Bioactive Compounds" Antibiotics 10, no. 6: 746. https://doi.org/10.3390/antibiotics10060746
APA StyleCarbone, D. A., Pellone, P., Lubritto, C., & Ciniglia, C. (2021). Evaluation of Microalgae Antiviral Activity and Their Bioactive Compounds. Antibiotics, 10(6), 746. https://doi.org/10.3390/antibiotics10060746