
Dynamic Adaptive Response of Pseudomonas aeruginosa to Clindamycin/Rifampicin-Impregnated Catheters

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Continuous Culture
2.2. Measurement of Growth Curves
2.3. Antimicrobial Susceptibility Testing
2.4. Detection of Mutations
2.5. Determination of Efflux Pump Activity
2.6. Invasion Assay
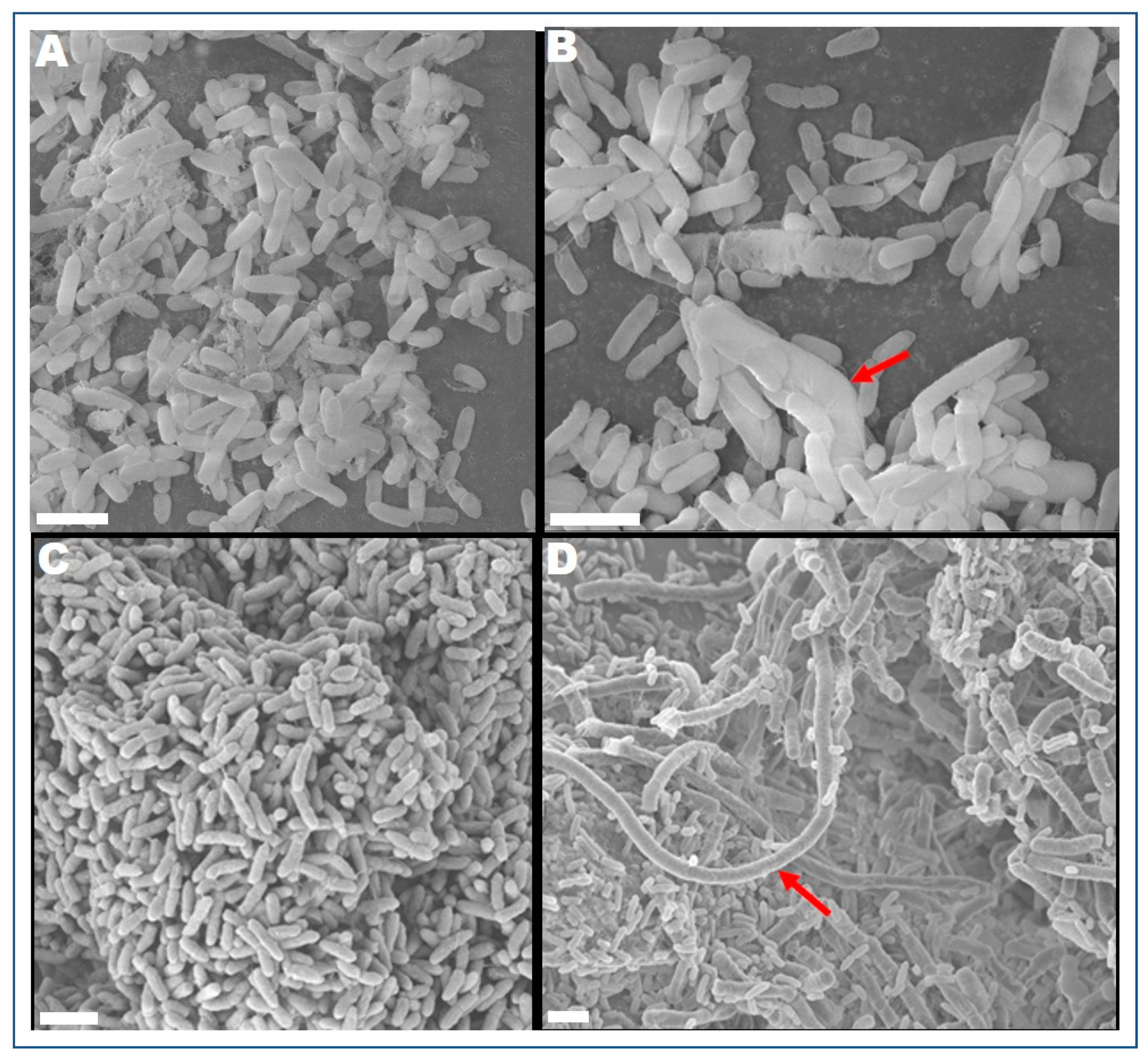
2.7. Field Emission Scanning Electron Microscope (FESEM)
2.8. Protein Extraction and Liquid Chromatography-Tandem Mass Spectrometry
3. Results and Discussion
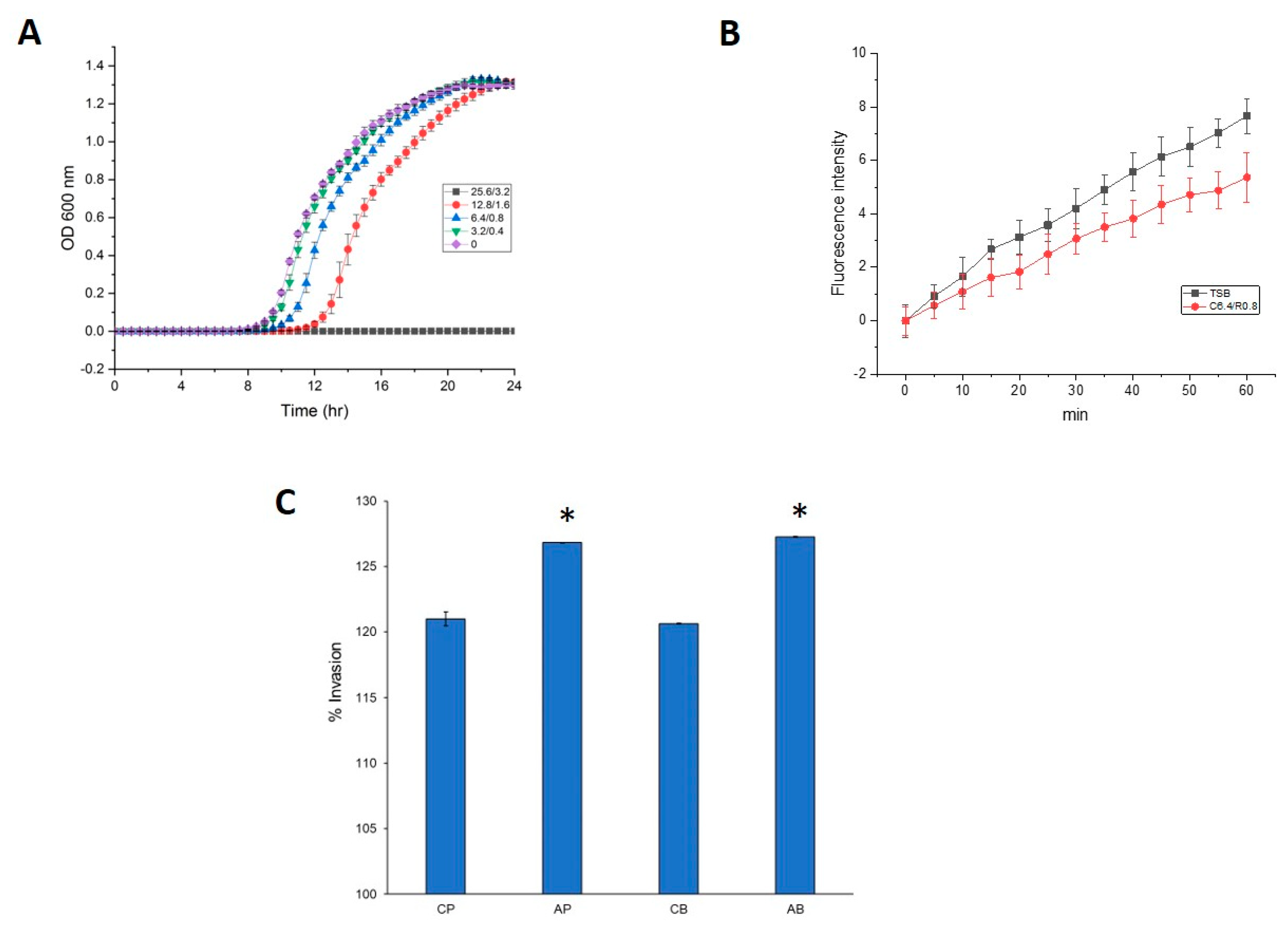
3.1. Cell Morphology and Antimicrobial Susceptibility of P. aeruginosa PAO1
3.2. Proteomic Insights into the Global Cellular Response to Clindamycin/Rifampicin
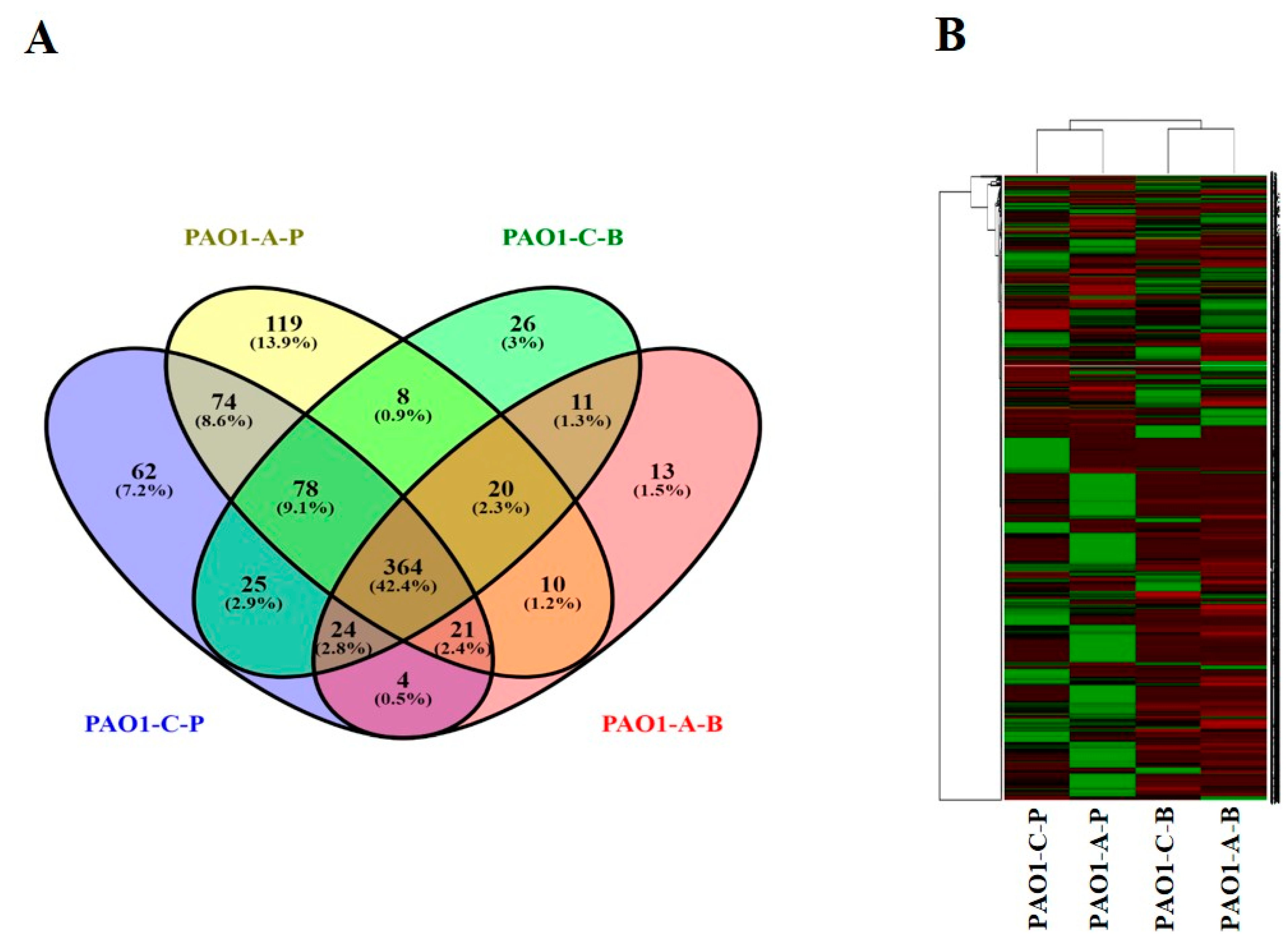
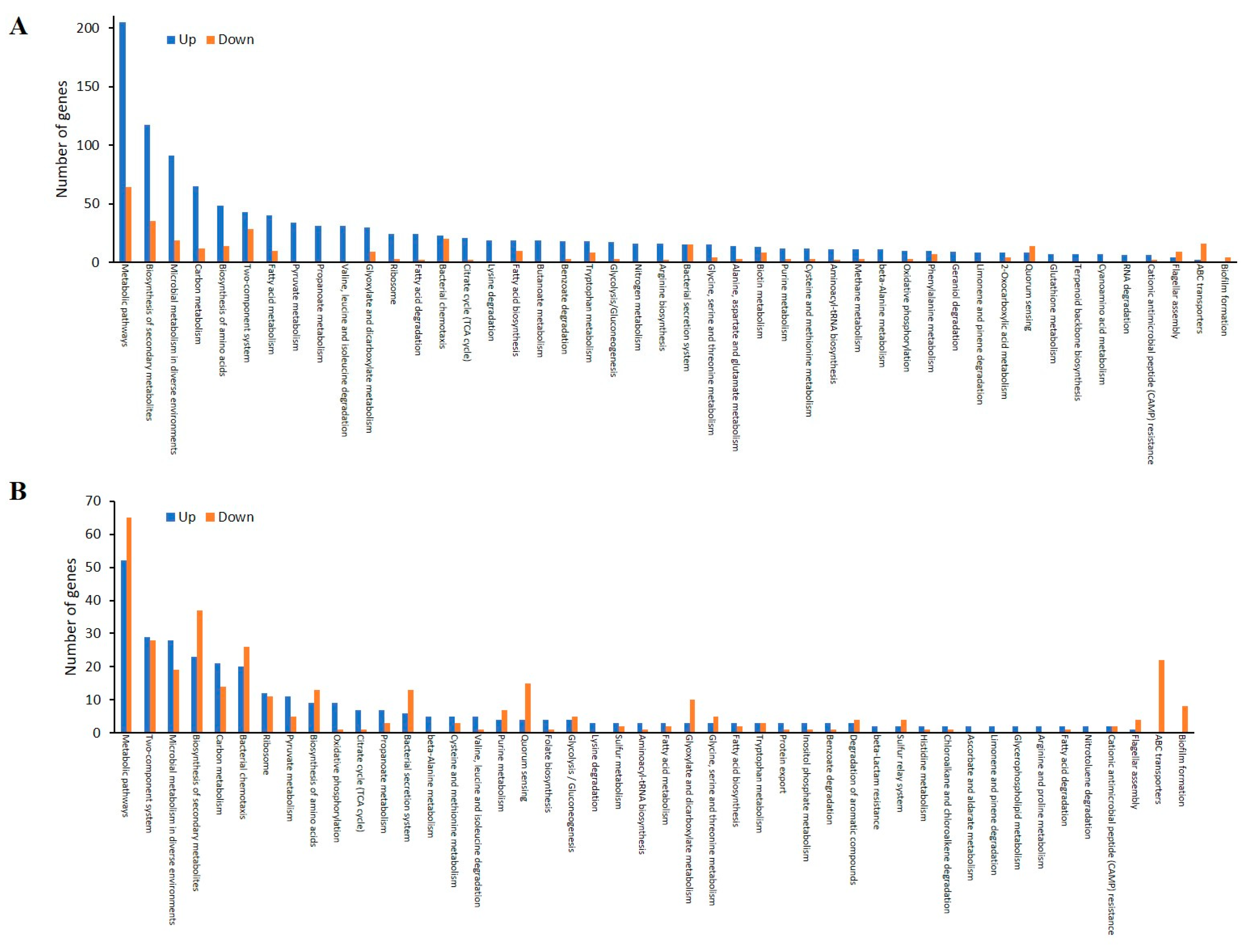
3.2.1. Global Overview of the Proteome Data
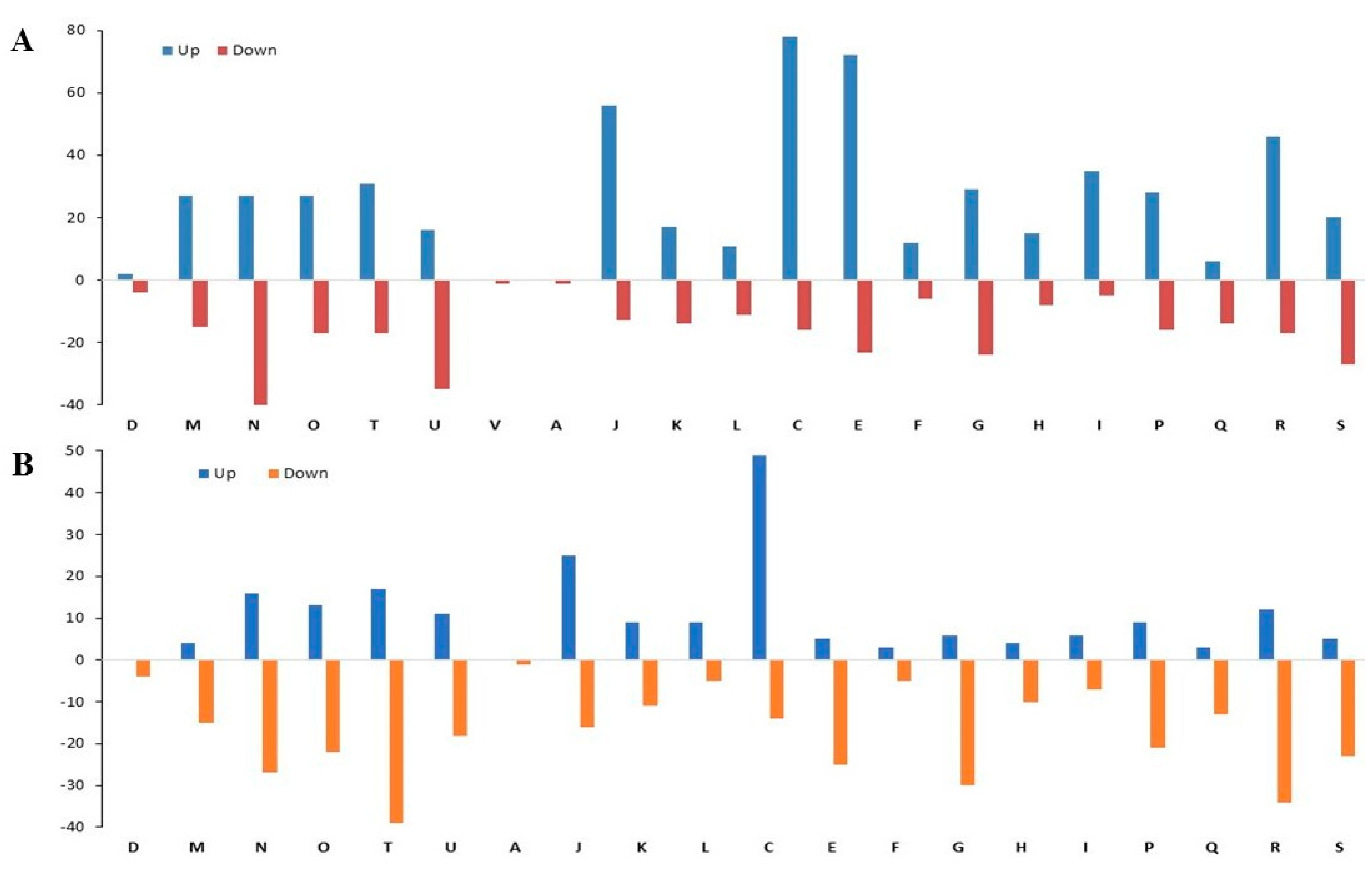
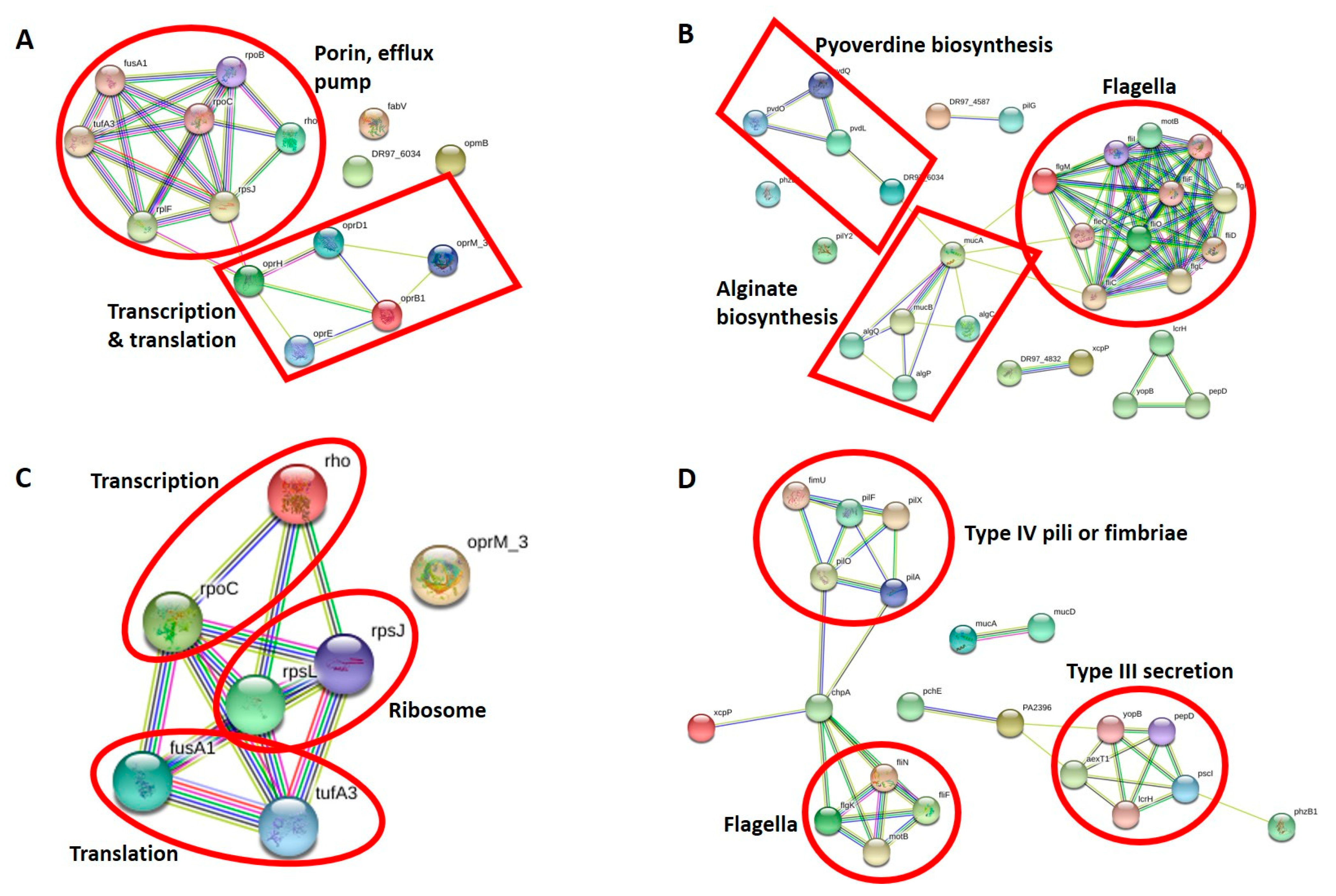
3.2.2. Antibiotic Resistance Proteins of CR-IC-Treated Planktonic Cells
3.2.3. Virulence Proteins of CR-IC-Treated Planktonic Cells
3.2.4. Antibiotic Resistance Proteins of CR-IC-Treated Biofilm Cells
3.2.5. Virulence Proteins of CR-IC-Treated Biofilm Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darouiche, R.O. Device-Associated Infections: A Macroproblem that Starts with Microadherence. Clin. Infect. Dis. 2001, 33, 1567–1572. [Google Scholar] [CrossRef]
- Von Eiff, C.; Jansen, B.; Kohnen, W.; Becker, K. Infections associated with medical devices: Pathogenesis, management and prophylaxis. Drugs 2005, 65, 179–214. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, S.J.; Records, A.R.; Orr, M.W.; Linden, S.B.; Lee, V.T. Catheter-Associated Urinary Tract Infection by Pseudomonas aeruginosa Is Mediated by Exopolysaccharide-Independent Biofilms. Infect. Immun. 2014, 82, 2048–2058. [Google Scholar] [CrossRef] [Green Version]
- Pascual, A. Pathogenesis of catheter-related infections: Lessons for new designs. Clin. Microbiol. Infect. 2002, 8, 256–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murga, R.; Miller, J.M.; Donlan, R.M. Biofilm Formation by Gram-Negative Bacteria on Central Venous Catheter Connectors: Effect of Conditioning Films in a Laboratory Model. J. Clin. Microbiol. 2001, 39, 2294–2297. [Google Scholar] [CrossRef] [Green Version]
- System, N. National Nosocomial Infections Surveillance (NNIS) System Report, Data Summary from January 1990-May 1999, Issued June 1999. Am. J. Infect. Control 1999, 27, 520–532. [Google Scholar] [CrossRef]
- Cross, A.; Allen, J.R.; Burke, J.; Ducel, G.; Harris, A.; John, J.; Johnson, D.; Lew, M.; MacMillan, B.; Meers, P.; et al. Nosocomial infections due to Pseudomonas aeruginosa: Review of recent trends. Rev. Infect. Dis. 1983, 5 (Suppl. 5), S837–S845. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, L.R.; Isabella, V.M.; Lewis, K. Pseudomonas aeruginosa Biofilms in Disease. Microb. Ecol. 2014, 68, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olejnickova, K.; Hola, V.; Ruzicka, F. Catheter-related infections caused byPseudomonas aeruginosa: Virulence factors involved and their relationships. Pathog. Dis. 2014, 72, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Lister, P.D.; Wolter, D.J.; Hanson, N.D. Antibacterial-resistant Pseudomonas aeruginosa: Clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 2009, 22, 582–610. [Google Scholar] [CrossRef] [Green Version]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Breidenstein, E.B.; Hancock, R.E. Creeping baselines and adaptive resistance to antibiotics. Drug Resist. Updat. 2011, 14, 1–21. [Google Scholar] [CrossRef]
- Machado, I.; Coquet, L.; Jouenne, T.; Pereira, M.O. Proteomic approach to Pseudomonas aeruginosa adaptive resistance to benzalkonium chloride. J. Proteom. 2013, 89, 273–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secer, H.I.; Kural, C.; Kaplan, M.; Kilic, A.; Duz, B.; Gönül, E.; Izci, Y. Comparison of the Efficacies of Antibiotic-Impregnated and Silver-Impregnated Ventricular Catheters on the Prevention of Infections. Pediatr. Neurosurg. 2008, 44, 444–447. [Google Scholar] [CrossRef]
- Munson, E.L.; Heard, S.O.; Doern, G.V. In Vitro Exposure of Bacteria to Antimicrobial Impregnated-Central Venous Catheters Does Not Directly Lead to the Emergence of Antimicrobial Resistance. Chest 2004, 126, 1628–1635. [Google Scholar] [CrossRef]
- Sampath, L.A.; Tambe, S.M.; Modak, S.M. In Vitro and In Vivo Efficacy of Catheters Impregnated With Antiseptics or Antibiotics: Evaluation of the Risk of Bacterial Resistance to the Antimicrobials in the Catheters. Infect. Control Hosp. Epidemiol. 2001, 22, 640–646. [Google Scholar] [CrossRef]
- Shakir, Z.; Khan, S.; Sung, K.; Khare, S.; Khan, A.; Steele, R.; Nawaz, M. Molecular Characterization of Fluoroquinolone-Resistant Aeromonas spp. Isolated from Imported Shrimp. Appl. Environ. Microbiol. 2012, 78, 8137–8141. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.; Sung, K.; Kweon, O.; Khan, S.; Nawaz, S.; Steele, R. Characterisation of novel mutations involved in quinolone resistance in Escherichia coli isolated from imported shrimp. Int. J. Antimicrob. Agents 2015, 45, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Moggridge, S.; Sorensen, P.H.; Morin, G.B.; Hughes, C.S. Extending the Compatibility of the SP3 Paramagnetic Bead Processing Approach for Proteomics. J. Proteome Res. 2018, 17, 1730–1740. [Google Scholar] [CrossRef]
- Jean-Toussaint, R.; Tian, Y.; Chaudhuri, A.D.; Haughey, N.J.; Sacan, A.; Ajit, S.K. Proteome characterization of small extracellular vesicles from spared nerve injury model of neuropathic pain. J. Proteom. 2020, 211, 103540. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, L.C.; Mergulhão, F.J. SEM Analysis of Surface Impact on Biofilm Antibiotic Treatment. Scanning 2017, 2017, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- She, P.; Luo, Z.; Chen, L.; Wu, Y. Efficacy of levofloxacin against biofilms of Pseudomonas aeruginosa isolated from patients with respiratory tract infections in vitro. Microbiologyopen 2019, 8, e00720. [Google Scholar] [CrossRef] [PubMed]
- López-Causapé, C.; Cabot, G.; Del Barrio-Tofiño, E.; Oliver, A. The Versatile Mutational Resistome of Pseudomonas aeruginosa. Front. Microbiol. 2018, 9, 685. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Winsor, G.L.; Griffiths, E.J.; Lo, R.; Dhillon, B.K.; Shay, J.A.; Brinkman, F.S.L. Enhanced annotations and features for comparing thousands ofPseudomonasgenomes in the Pseudomonas genome database. Nucleic Acids Res. 2016, 44, D646–D653. [Google Scholar] [CrossRef] [Green Version]
- Raneri, M.; Pinatel, E.; Peano, C.; Rampioni, G.; Leoni, L.; Bianconi, I.; Jousson, O.; Dalmasio, C.; Ferrante, P.; Briani, F. Pseudomonas aeruginosa mutants defective in glucose uptake have pleiotropic phenotype and altered virulence in non-mammal infection models. Sci. Rep. 2018, 8, 16912. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Luo, Y.-F.; Williams, B.J.; Blackwell, T.S.; Xie, C.-M. Structure and function of OprD protein in Pseudomonas aeruginosa: From antibiotic resistance to novel therapies. Int. J. Med. Microbiol. 2012, 302, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Samanta, S.; Bodrenko, I.; Acosta-Gutierrez, S.; D’Agostino, T.; Pathania, M.; Ghai, I.; Schleberger, C.; Bumann, D.; Wagner, R.; Winterhalter, M.; et al. Getting Drugs through Small Pores: Exploiting the Porins Pathway in Pseudomonas aeruginosa. ACS Infect. Dis. 2018, 4, 1519–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevalier, S.; Bouffartigues, E.; Bodilis, J.; Maillot, O.; Lesouhaitier, O.; Feuilloley, M.G.J.; Orange, N.; Dufour, A.; Cornelis, P. Structure, function and regulation of Pseudomonas aeruginosa porins. FEMS Microbiol. Rev. 2017, 41, 698–722. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, D.L.; Velayudhan, J.; Kenney, T.F.; Therrien, J.H.; Sutherland, J.L.; Barker, L.M.; Baker, W.R. Fosfomycin Enhances the Active Transport of Tobramycin in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2012, 56, 1529–1538. [Google Scholar] [CrossRef] [Green Version]
- Ranieri, M.R.M.; Chan, D.C.K.; Yaeger, L.N.; Rudolph, M.; Karabelas-Pittman, S.; Abdo, H.; Chee, J.; Harvey, H.; Nguyen, U.; Burrows, L.L. Thiostrepton Hijacks Pyoverdine Receptors to Inhibit Growth of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Fito-Boncompte, L.; Chapalain, A.; Bouffartigues, E.; Chaker, H.; Lesouhaitier, O.; Gicquel, G.; Bazire, A.; Madi, A.; Connil, N.; Véron, W.; et al. Full virulence of Pseudomonas aeruginosa requires OprF. Infect. Immun. 2011, 79, 1176–1186. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Sun, L.; Ali, F.; Guo, Z.; Zhang, L.; Lin, W.; Lin, X. Proteomic Analysis of Alterations in Aeromonas hydrophila Outer Membrane Proteins in Response to Oxytetracycline Stress. Microb. Drug Resist. 2018, 24, 1067–1074. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.-M.; Yang, J.-N.; Peng, X.-X.; Li, H. A Novel Negative Regulation Mechanism of Bacterial Outer Membrane Proteins in Response to Antibiotic Resistance. J. Proteome Res. 2010, 9, 5952–5959. [Google Scholar] [CrossRef]
- Kesavan, D.; Vasudevan, A.; Wu, L.; Chen, J.; Su, Z.; Wang, S.; Xu, H. Integrative analysis of outer membrane vesicles proteomics and whole-cell transcriptome analysis of eravacycline induced Acinetobacter baumannii strains. BMC Microbiol. 2020, 20, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laborda, P.; Alcalde-Rico, M.; Blanco, P.; Martínez, J.L.; Hernando-Amado, S. Novel Inducers of the Expression of Multidrug Efflux Pumps That Trigger Pseudomonas aeruginosa Transient Antibiotic Resistance. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Meng, J.; Jia, M.; Ma, X.; He, G.; Yu, J.; Wang, R.; Bai, H.; Hou, Z.; Luo, X. oprM as a new target for reversion of multidrug resistance in Pseudomonas aeruginosa by antisense phosphorothioate oligodeoxynucleotides. FEMS Immunol. Med. Microbiol. 2010, 60, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Substrate specificities of MexAB-OprM, MexCD-OprJ, and MexXY-oprM efflux pumps in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3322–3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillis, R.J.; White, K.G.; Choi, K.-H.; Wagner, V.E.; Schweizer, H.P.; Iglewski, B.H. Molecular Basis of Azithromycin-Resistant Pseudomonas aeruginosa Biofilms. Antimicrob. Agents Chemother. 2005, 49, 3858–3867. [Google Scholar] [CrossRef] [Green Version]
- Pamp, S.J.; Gjermansen, M.; Johansen, H.K.; Tolker-Nielsen, T. Tolerance to the antimicrobial peptide colistin in Pseudomonas aeruginosa biofilms is linked to metabolically active cells, and depends on the pmr and mexAB-oprM genes. Mol. Microbiol. 2008, 68, 223–240. [Google Scholar] [CrossRef]
- Yang, L.; Chen, L.; Shen, L.; Surette, M.; Duan, K. Inactivation of MuxABC-OpmB transporter system in Pseudomonas aeruginosa leads to increased ampicillin and carbenicillin resistance and decreased virulence. J. Microbiol. 2011, 49, 107–114. [Google Scholar] [CrossRef]
- Bayston, R.; Ashraf, W.; Ortori, C. Does release of antimicrobial agents from impregnated external ventricular drainage catheters affect the diagnosis of ventriculitis? J. Neurosurg. 2016, 124, 375–381. [Google Scholar] [CrossRef]
- Dinos, G.P. The macrolide antibiotic renaissance. Br. J. Pharmacol. 2017, 174, 2967–2983. [Google Scholar] [CrossRef]
- Ding, Y.; Teo, J.W.P.; Drautz-Moses, D.I.; Schuster, S.C.; Givskov, M.; Yang, L. Acquisition of resistance to carbapenem and macrolide-mediated quorum sensing inhibition by Pseudomonas aeruginosa via ICETn43716385. Commun. Biol. 2018, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Lin, L.; Yao, Z.; Li, W.; Sun, L.; Zhang, D.; Luo, J.; Lin, W. An integrated quantitative and targeted proteomics reveals fitness mechanisms of Aeromonas hydrophila under oxytetracycline stress. J. Proteome Res. 2015, 14, 1515–1525. [Google Scholar] [CrossRef]
- Zhu, J.-H.; Wang, B.-W.; Pan, M.; Zeng, Y.-N.; Rego, H.; Javid, B. Rifampicin can induce antibiotic tolerance in mycobacteria via paradoxical changes in rpoB transcription. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Giddey, A.; De Kock, E.; Nakedi, K.C.; Garnett, S.; Nel, A.J.M.; Soares, N.C.; Blackburn, J.M. A temporal proteome dynamics study reveals the molecular basis of induced phenotypic resistance in Mycobacterium smegmatis at sub-lethal rifampicin concentrations. Sci. Rep. 2017, 7, srep43858. [Google Scholar] [CrossRef] [Green Version]
- Morita, Y.; Tomida, J.; Kawamura, Y. Responses of Pseudomonas aeruginosa to antimicrobials. Front. Microbiol. 2014, 4, 422. [Google Scholar] [CrossRef] [Green Version]
- Tart, A.H.; Wolfgang, M.C.; Wozniak, D.J. The Alternative Sigma Factor AlgT Represses Pseudomonas aeruginosa Flagellum Biosynthesis by Inhibiting Expression of fleQ. J. Bacteriol. 2005, 187, 7955–7962. [Google Scholar] [CrossRef] [Green Version]
- Kawamura-Sato, K.; Iinuma, Y.; Hasegawa, T.; Horii, T.; Yamashino, T.; Ohta, M. Effect of Subinhibitory Concentrations of Macrolides on Expression of Flagellin in Pseudomonas aeruginosa and Proteus mirabilis. Antimicrob. Agents Chemother. 2000, 44, 2869–2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diepold, A.; Armitage, J.P. Type III secretion systems: The bacterial flagellum and the injectisome. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bader, M.W.; Navarre, W.; Shiau, W.; Nikaido, H.; Frye, J.; McClelland, M.; Fang, F.; Miller, S.I. Regulation of Salmonella typhimurium virulence gene expression by cationic antimicrobial peptides. Mol. Microbiol. 2003, 50, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Yim, G.; de la Cruz, F.; Spiegelman, G.B.; Davies, J. Transcription Modulation of Salmonella enterica Serovar Typhimurium Promoters by Sub-MIC Levels of Rifampin. J. Bacteriol. 2006, 188, 7988–7991. [Google Scholar] [CrossRef] [Green Version]
- Soscia, C.; Hachani, A.; Bernadac, A.; Filloux, A.; Bleves, S. Cross Talk between Type III Secretion and Flagellar Assembly Systems in Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 3124–3132. [Google Scholar] [CrossRef] [Green Version]
- Filloux, A. Protein Secretion Systems in Pseudomonas aeruginosa: An Essay on Diversity, Evolution, and Function. Front. Microbiol. 2011, 2, 155. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Weng, X.; Jiang, R.; Wu, J.; Mi, Z. Transcriptomic response of uropathogenic Escherichia coli to Taraxacum mongolicum. Int. J. Clin. Exp. Med. 2017, 10, 12030–12043. [Google Scholar]
- Losada, L.; Shea, A.A.; DeShazer, D. A MarR family transcriptional regulator and subinhibitory antibiotics regulate type VI secretion gene clusters in Burkholderia pseudomallei. Microbiology 2018, 164, 1196–1211. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ye, M.; Li, X.; Li, J.; Deng, Z.; Yao, Y.-F.; Ou, H.-Y. Identification and Characterization of an Antibacterial Type VI Secretion System in the Carbapenem-Resistant Strain Klebsiella pneumoniae HS11286. Front. Cell. Infect. Microbiol. 2017, 7, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penesyan, A.; Nagy, S.S.; Kjelleberg, S.; Gillings, M.R.; Paulsen, I. Rapid microevolution of biofilm cells in response to antibiotics. NPJ Biofilms Microb. 2019, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, D.J.; Keyser, R. Effects of Subinhibitory Concentrations of Macrolide Antibiotics on Pseudomonas aeruginosa. Chest 2004, 125, 62S–69S. [Google Scholar] [CrossRef] [PubMed]
- Nalca, Y.; Jänsch, L.; Bredenbruch, F.; Geffers, R.; Buer, J.; Häussler, S. Quorum-Sensing Antagonistic Activities of Azithromycin in Pseudomonas aeruginosa PAO1: A Global Approach. Antimicrob. Agents Chemother. 2006, 50, 1680–1688. [Google Scholar] [CrossRef] [Green Version]
- Swatton, J.E.; Davenport, P.W.; Maunders, E.A.; Griffin, J.L.; Lilley, K.S.; Welch, M. Impact of Azithromycin on the Quorum Sensing-Controlled Proteome of Pseudomonas aeruginosa. PLoS ONE 2016, 11, e0147698. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Ma, L.Z. Biofilm matrix and its regulation in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2013, 14, 20983–21005. [Google Scholar] [CrossRef]
- Liu, X.; Shen, B.; Du, P.; Wang, N.; Wang, J.; Li, J.; Sun, A. Transcriptomic analysis of the response of Pseudomonas fluorescens to epigallocatechin gallate by RNA-seq. PLoS ONE 2017, 12, e0177938. [Google Scholar] [CrossRef] [Green Version]
- Springer, B.; Kidan, Y.G.; Prammananan, T.; Ellrott, K.; Bottger, E.C.; Sander, P. Mechanisms of streptomycin resistance: Selection of mutations in the 16S rRNA gene conferring resistance. Antimicrob. Agents Chemother. 2001, 45, 2877–2884. [Google Scholar] [CrossRef] [Green Version]
- Park, A.J.; Murphy, K.; Krieger, J.R.; Brewer, D.; Taylor, P.; Habash, M.; Khursigara, C.M. A temporal examination of the planktonic and biofilm proteome of whole cell Pseudomonas aeruginosa PAO1 using quantitative mass spectrometry. Mol. Cell Proteom. 2014, 13, 1095–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrows, L.L. Pseudomonas aeruginosaTwitching Motility: Type IV Pili in Action. Annu. Rev. Microbiol. 2012, 66, 493–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Sass, A.; Van Acker, H.; Wille, J.; Verhasselt, B.; Van Nieuwerburgh, F.; Kaever, V.; Crabbe, A.; Coenye, T. Coumarin Reduces Virulence and Biofilm Formation in Pseudomonas aeruginosa by Affecting Quorum Sensing, Type III Secretion and C-di-GMP Levels. Front. Microbiol. 2018, 9, 1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus Tag | Gene | Product | Fold Ratio |
|---|---|---|---|
| PA0156 | triA | Multidrug efflux system, membrane fusion component | 0.97 |
| PA0291 | oprE | Outer membrane low permeability porin, OprD family | 0.00 |
| PA0425 | mexA | Multidrug efflux system, membrane fusion component | 0.71 |
| PA0427 | oprM | Multidrug efflux system, outer membrane factor lipoprotein | 18.38 |
| PA0958 | oprD | Outer membrane low permeability porin, OprD family | 13.16 |
| PA1178 | oprH | PhoP/Q and low Mg2+ inducible outer membrane protein H1 precursor | 1000.00 |
| PA1777 | oprF | Nonspecific porin and structural outer membrane protein OprF | 1.41 |
| PA2071 | fusA2 | Translation elongation factor G | 1.37 |
| PA2291 | oprB2 | Probable glucose-sensitive porin | 1000.00 |
| PA2398 | fpvA | Ferripyoverdine receptor | 1000.00 |
| PA2525 | opmB | Outer membrane factor (OMF) lipoprotein associated with MdtABC efflux system | 0.00 |
| PA2950 | fabV | Enoyl-[acyl-carrier-protein] reductase [NADH] | 6.63 |
| PA3186 | oprB | Glucose/carbohydrate outer membrane porin OprB precursor | 1000.00 |
| PA4248 | rplF | LSU ribosomal protein L6p (L9e) | 3.21 |
| PA4264 | rpsJ | SSU ribosomal protein S10p (S20e) | 19.09 |
| PA4266 | fusA1 | Translation elongation factor G | 3.45 |
| PA4269 | rpoC | DNA-directed RNA polymerase beta subunit | 6.81 |
| PA4270 | rpoB | DNA-directed RNA polymerase beta subunit | 13.15 |
| PA4277 | tufB | Translation elongation factor Tu | 3.46 |
| PA4974 | Outer membrane channel TolC (OpmH) | 1.04 | |
| PA5239 | rho | Transcription termination factor Rho | 6.61 |
| Locus Tag | Gene | Product | Fold Ratio |
|---|---|---|---|
| PA0070 | Autotransporter adhesin | 0.46 | |
| PA0083 | tssB1 | T6SS component TssB (ImpB/VipA) | 0.28 |
| PA0408 | pilG | Twitching motility protein PilG | 0.00 |
| PA0409 | pilH | Twitching motility protein PilH | 0.79 |
| PA0411 | pilJ | Twitching motility protein PilJ | 0.74 |
| PA0413 | chpA | Still frameshift probable component of chemotactic signal transduction system | 1.00 |
| PA0763 | mucA | Sigma factor RpoE negative regulatory protein RseA | 1000.00 |
| PA0764 | mucB | Sigma factor RpoE negative regulatory protein RseB precursor | 0.00 |
| PA0766 | mucD | HtrA protease/chaperone protein | 0.72 |
| PA1086 | flgK | Flagellar hook-associated protein FlgK | 0.00 |
| PA1087 | flgL | Flagellar hook-associated protein FlgL | 0.25 |
| PA1092 | fliC | Flagellin protein FlaB | 7.01 |
| PA1094 | fliD | Flagellar cap protein FliD | 0.13 |
| PA1096 | Two-component system sensor histidine kinase | 0.00 | |
| PA1097 | fleQ | Flagellar regulatory protein FleQ | 0.21 |
| PA1101 | fliF | Flagellar M-ring protein FliF | 0.00 |
| PA1103 | fliH | Flagellar assembly protein FliH | 0.06 |
| PA1104 | fliI | Flagellum-specific ATP synthase FliI | 0.00 |
| PA1445 | fliO | Flagellar biosynthesis protein FliO | 0.00 |
| PA1707 | pcrH | Type III secretion chaperone protein for YopD (SycD) | 1000.00 |
| PA1708 | popB | Type III secretion host injection protein (YopB) | 0.00 |
| PA1709 | popD | Type III secretion host injection and negative regulator protein (YopD) | 1000.00 |
| PA1900 | phzB2 | Phenazine biosynthesis protein PhzB | 1000.00 |
| PA2385 | pvdQ | Acyl-homoserine lactone acylase PvdQ | 24.33 |
| PA2392 | pvdP | Pyoverdine biosynthesis related protein PvdP | 0.00 |
| PA2393 | Putative dipeptidase, pyoverdin biosynthesis PvdM | 7.82 | |
| PA2395 | pvdO | PvdO, pyoverdine responsive serine/threonine kinase (predicted by OlgaV) | 2.22 |
| PA2396 | pvdF | Pyoverdine synthetase PvdF, N5-hydroxyornithine formyltransferase | 1.05 |
| PA2398 | fpvA | Outer membrane ferripyoverdine receptor FpvA | 1000.00 |
| PA2412 | MbtH-like NRPS chaperone | 1000.00 | |
| PA2424 | pvdL | Pyoverdine chromophore precursor synthetase PvdL | 0.00 |
| PA3101 | xcpT | General secretion pathway protein G | 0.32 |
| PA3104 | xcpP | General secretion pathway protein C | 0.00 |
| PA3115 | fimV | Probable type IV pilus assembly FimV-related transmembrane protein | 0.43 |
| PA3351 | flgM | Negative regulator of flagellin synthesis FlgM (anti-sigma28) | 0.44 |
| PA3352 | Flagellar biosynthesis protein FlgN | 4.30 | |
| PA3805 | pilF | Type IV pilus biogenesis protein PilF | 0.56 |
| PA4211 | phzB1 | Phenazine biosynthesis protein PhzB | 1000.00 |
| PA4230 | pchB | Isochorismate pyruvate-lyase | 0.69 |
| PA4525 | pilA | Type IV pilin PilA | 1.21 |
| PA4555 | pilY2 | Type IV fimbrial biogenesis protein PilY2 | 0.00 |
| PA4953 | motB | Flagellar motor rotation protein MotB | 15.08 |
| PA5042 | pilO | Type IV pilus biogenesis protein PilO | 0.90 |
| PA5253 | algP | Alginate regulatory protein AlgP | 0.07 |
| PA5255 | algQ | Alginate regulatory protein AlgQ | 1000.00 |
| PA5322 | algC | Phosphoglucomutase | 1000.00 |
| Locus Tag | Gene | Product | Fold Ratio |
|---|---|---|---|
| PA0156 | Multidrug efflux system, membrane fusion component | 0.97 | |
| PA0427 | oprM | Multidrug efflux system, outer membrane factor lipoprotein | 1000.00 |
| PA1777 | oprF | Nonspecific porin and structural outer membrane protein OprF | 1.29 |
| PA4248 | rplF | LSU ribosomal protein L6p (L9e) | 1.69 |
| PA4264 | rpsJ | SSU ribosomal protein S10p (S20e) | 2.17 |
| PA4266 | fusA1 | Translation elongation factor G | 4.60 |
| PA4268 | rpsL | SSU ribosomal protein S12p (S23e) | 1000.00 |
| PA4269 | rpoC | DNA-directed RNA polymerase beta subunit | 2.68 |
| PA4277 | tufB | Translation elongation factor Tu | 2.06 |
| PA4974 | Outer membrane channel TolC (OpmH) | 1.13 | |
| PA5239 | rho | Transcription termination factor Rho | 0.08 |
| Locus Tag | Gene | Product | Fold Ratio |
|---|---|---|---|
| PA0044 | exoT | Hypothetical protein | 1000.00 |
| PA0070 | Autotransporter adhesin | 0.69 | |
| PA0083 | T6SS component TssB (ImpB/VipA) | 0.00 | |
| PA0408 | pilG | Twitching motility protein PilG | 0.77 |
| PA0409 | pilH | Twitching motility protein PilH | 0.66 |
| PA0411 | pilJ | Twitching motility protein PilJ | 0.92 |
| PA0413 | chpA | Still frameshift probable component of chemotactic signal transduction system | 0.27 |
| PA0763 | mucA | Sigma factor RpoE negative regulatory protein RseA | 0.00 |
| PA0766 | mucD | HtrA protease/chaperone protein | 2.88 |
| PA1086 | flgK | Flagellar hook-associated protein FlgK | 0.40 |
| PA1087 | flgL | Flagellar hook-associated protein FlgL | 0.95 |
| PA1092 | fliC | Flagellin protein FlaB | 1.67 |
| PA1094 | fliD | Flagellar cap protein FliD | 0.74 |
| PA1096 | Two-component system sensor histidine kinase | 1.29 | |
| PA1097 | fleQ | Flagellar regulatory protein FleQ | 0.73 |
| PA1101 | fliF | Flagellar M-ring protein FliF | 0.00 |
| PA1103 | fliH | Flagellar assembly protein FliH | 0.77 |
| PA1444 | fliN | Flagellar motor switch protein FliN | 1000.00 |
| PA1707 | pcrH | Type III secretion chaperone protein for YopD (SycD) | 2.54 |
| PA1708 | popB | Type III secretion host injection protein (YopB) | 0.00 |
| PA1709 | popD | Type III secretion host injection and negative regulator protein (YopD) | 0.41 |
| PA1722 | pscI | Type III secretion cytoplasmic protein (YscI) | 1000.00 |
| PA2385 | pvdQ | Acyl-homoserine lactone acylase PvdQ | 0.96 |
| PA2393 | Putative dipeptidase | 4.00 | |
| PA2395 | pvdO | PvdO, pyoverdine responsive serine/threonine kinase | 0.59 |
| PA2396 | pvdF | Pyoverdine synthetase PvdF, N5-hydroxyornithine formyltransferase | 5.19 |
| PA2412 | MbtH-like NRPS chaperone | 1.69 | |
| PA2413 | pvdH | Pyoverdin biosynthesis protein PvdH | 1.18 |
| PA3101 | xcpT | General secretion pathway protein G | 1.31 |
| PA3101 | xcpT | General secretion pathway protein G | 1.31 |
| PA3104 | xcpP | General secretion pathway protein C | 0.00 |
| PA3115 | fimV | Probable type IV pilus assembly FimV-related transmembrane protein | 0.69 |
| PA3351 | flgM | Negative regulator of flagellin synthesis FlgM (anti-sigma28) | 0.81 |
| PA3352 | Flagellar biosynthesis protein FlgN | 0.07 | |
| PA3805 | pilF | Type IV pilus biogenesis protein PilF | 0.18 |
| PA4211 | phzB1 | Phenazine biosynthesis protein PhzB | 1000.00 |
| PA4226 | pchE | Dihydroaeruginoate synthetase PchE | 0.00 |
| PA4230 | pchB | Isochorismate pyruvate-lyase | 0.75 |
| PA4525 | pilA | Type IV pilin PilA | 21.73 |
| PA4550 | fimU | type 4 fimbrial biogenesis protein FimU | 0.00 |
| PA4553 | pilX | Type IV fimbrial biogenesis protein PilX | 0.00 |
| PA4953 | motB | Flagellar motor rotation protein MotB | 0.00 |
| PA5041 | pilP | Type IV pilus biogenesis protein PilP | 1.31 |
| PA5042 | pilO | Type IV pilus biogenesis protein PilO | 0.00 |
| PA5253 | algP | Alginate regulatory protein AlgP | 1.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sung, K.; Chon, J.; Kweon, O.; Nho, S.; Kim, S.; Park, M.; Paredes, A.; Lim, J.-H.; Khan, S.A.; Phillips, K.S.; et al. Dynamic Adaptive Response of Pseudomonas aeruginosa to Clindamycin/Rifampicin-Impregnated Catheters. Antibiotics 2021, 10, 752. https://doi.org/10.3390/antibiotics10070752
Sung K, Chon J, Kweon O, Nho S, Kim S, Park M, Paredes A, Lim J-H, Khan SA, Phillips KS, et al. Dynamic Adaptive Response of Pseudomonas aeruginosa to Clindamycin/Rifampicin-Impregnated Catheters. Antibiotics. 2021; 10(7):752. https://doi.org/10.3390/antibiotics10070752
Chicago/Turabian StyleSung, Kidon, Jungwhan Chon, Ohgew Kweon, Seongwon Nho, Seongjae Kim, Miseon Park, Angel Paredes, Jin-Hee Lim, Saeed A. Khan, Kenneth Scott Phillips, and et al. 2021. "Dynamic Adaptive Response of Pseudomonas aeruginosa to Clindamycin/Rifampicin-Impregnated Catheters" Antibiotics 10, no. 7: 752. https://doi.org/10.3390/antibiotics10070752
APA StyleSung, K., Chon, J., Kweon, O., Nho, S., Kim, S., Park, M., Paredes, A., Lim, J. -H., Khan, S. A., Phillips, K. S., & Cerniglia, C. E. (2021). Dynamic Adaptive Response of Pseudomonas aeruginosa to Clindamycin/Rifampicin-Impregnated Catheters. Antibiotics, 10(7), 752. https://doi.org/10.3390/antibiotics10070752