
Modulation of the Intestinal Microbiota by the Early Intervention with Clostridium Butyricum in Muscovy Ducks


Abstract
:1. Introduction
2. Results
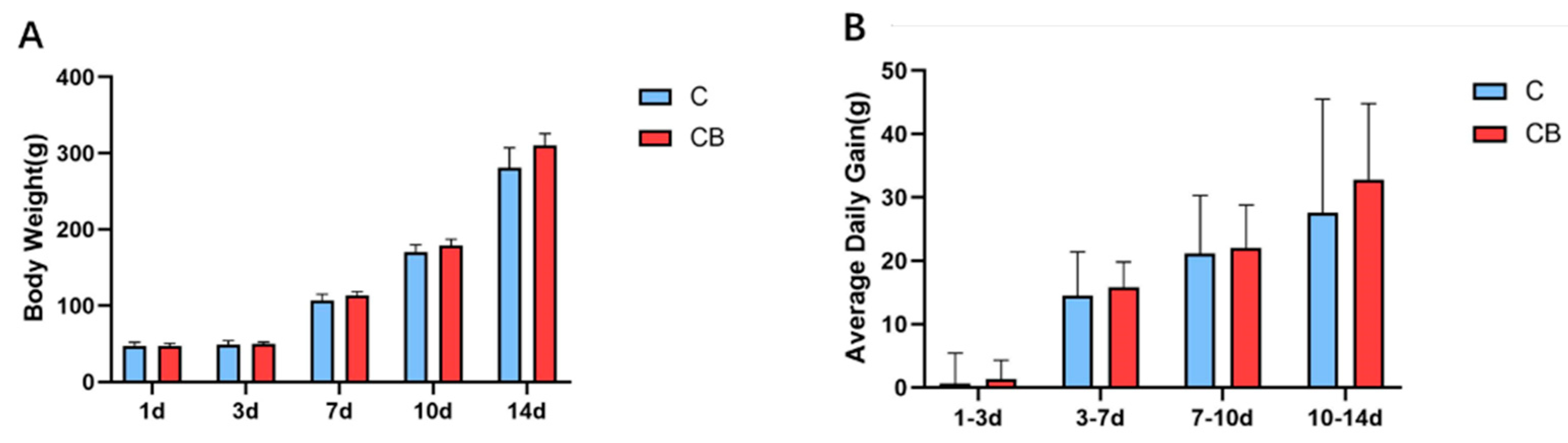
2.1. Growth Performance
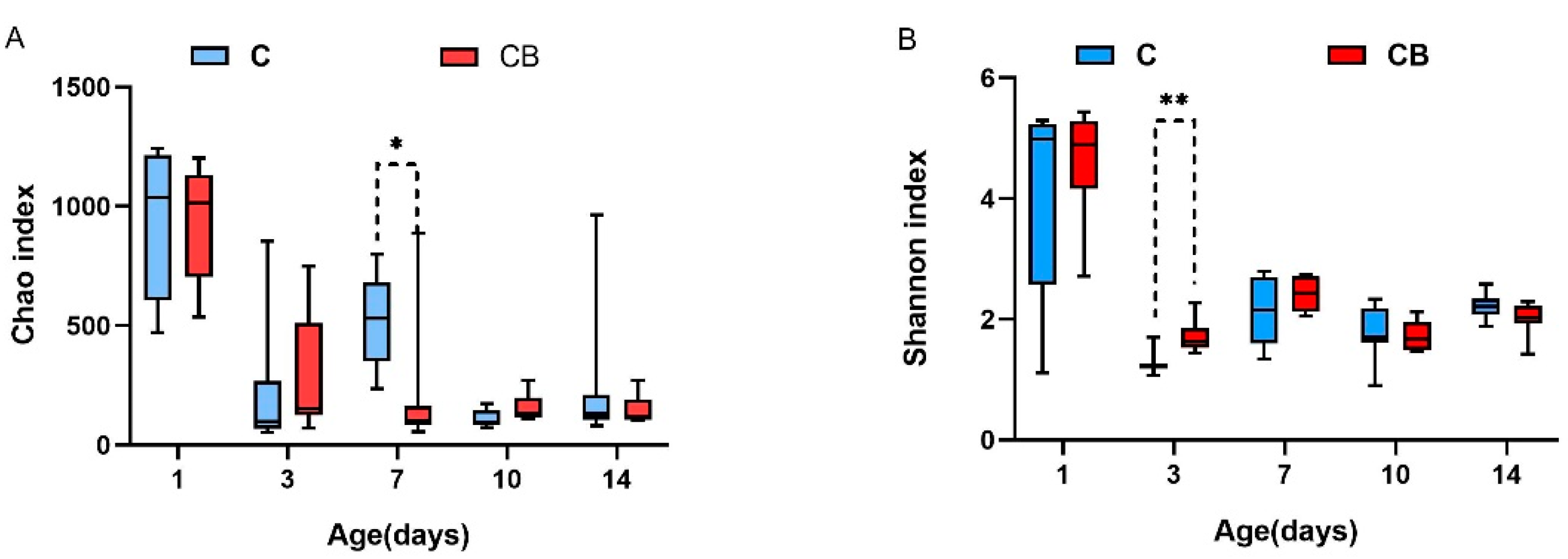
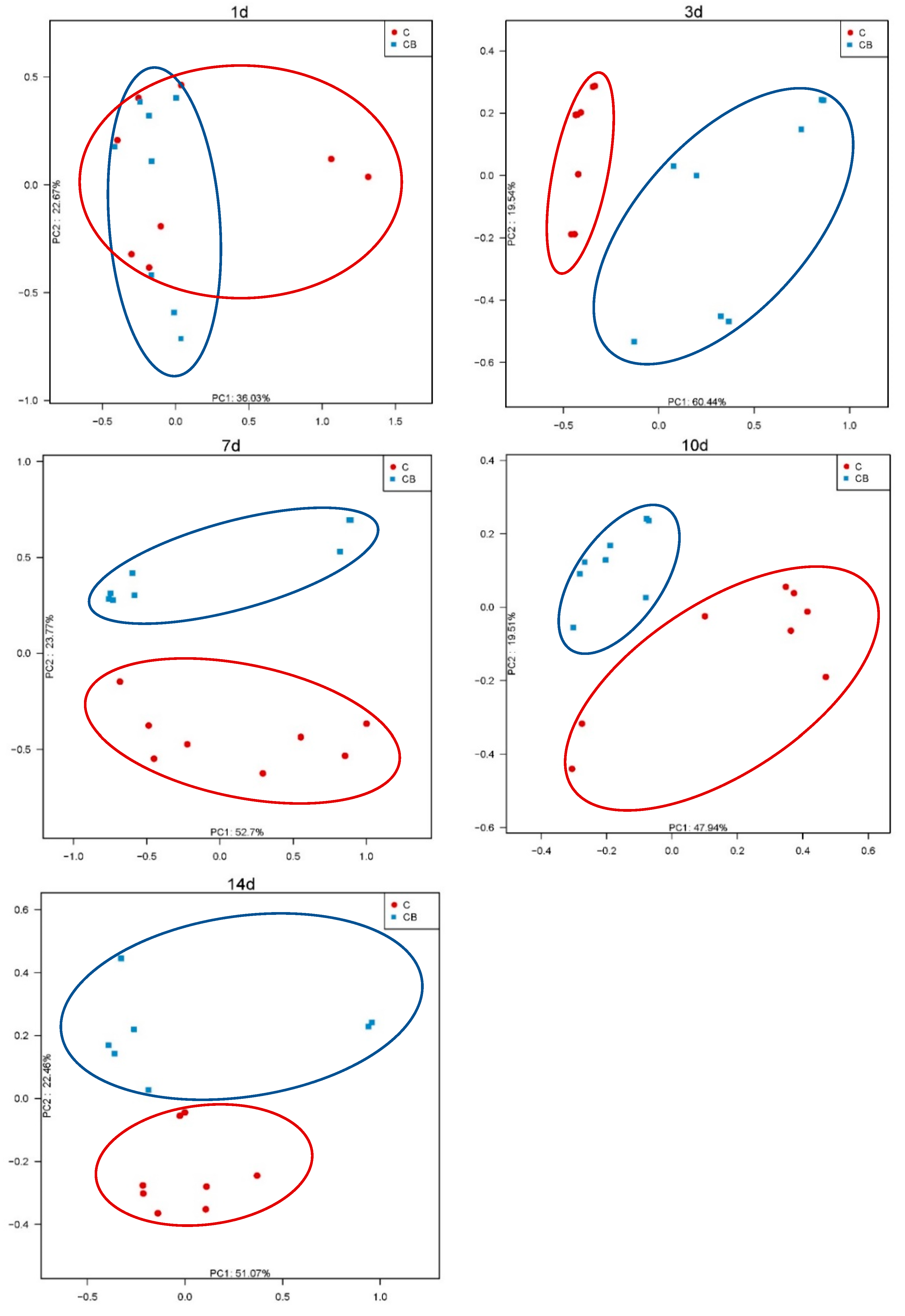
2.2. Diversity and Structure of the Cecum Microbiota
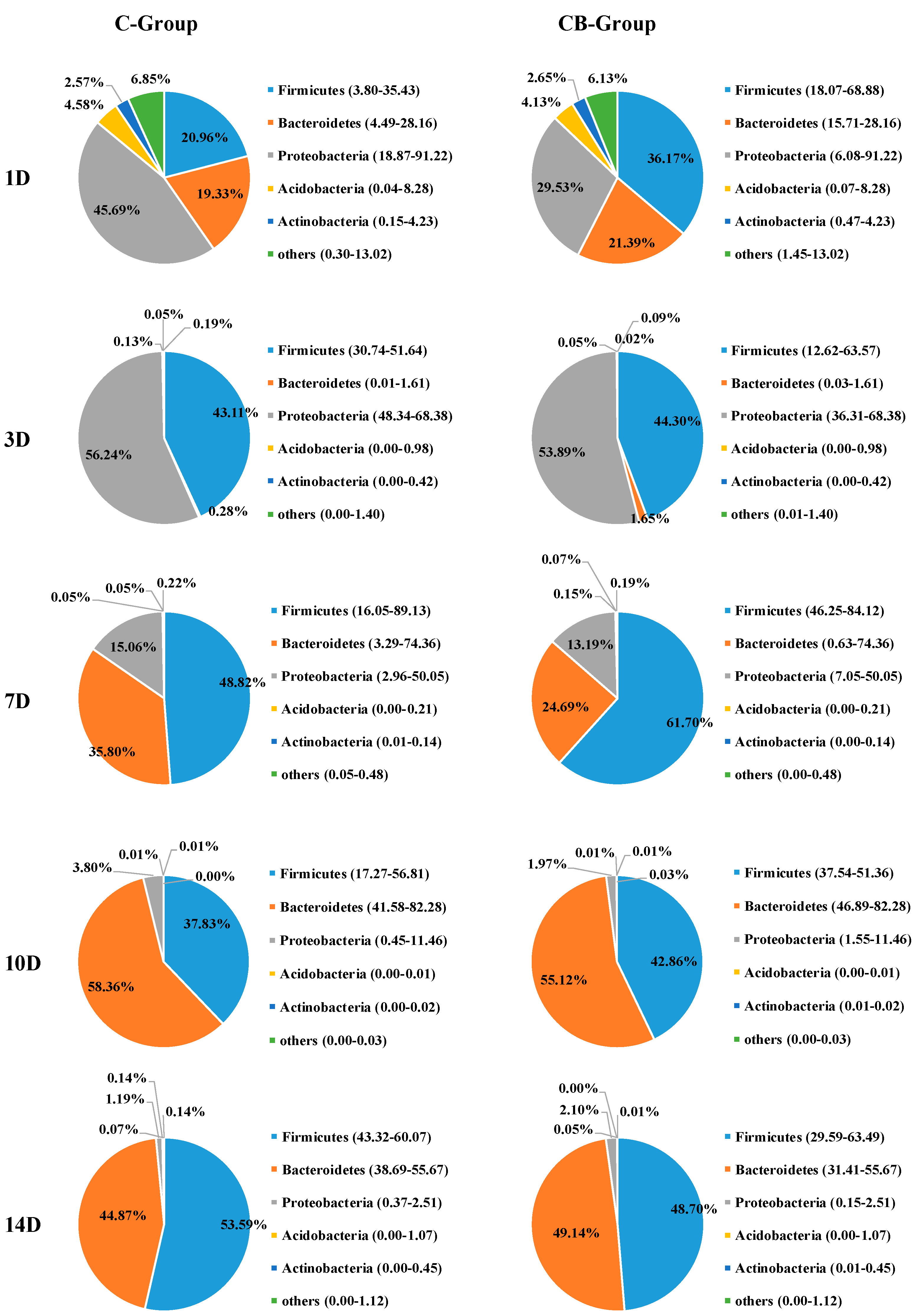
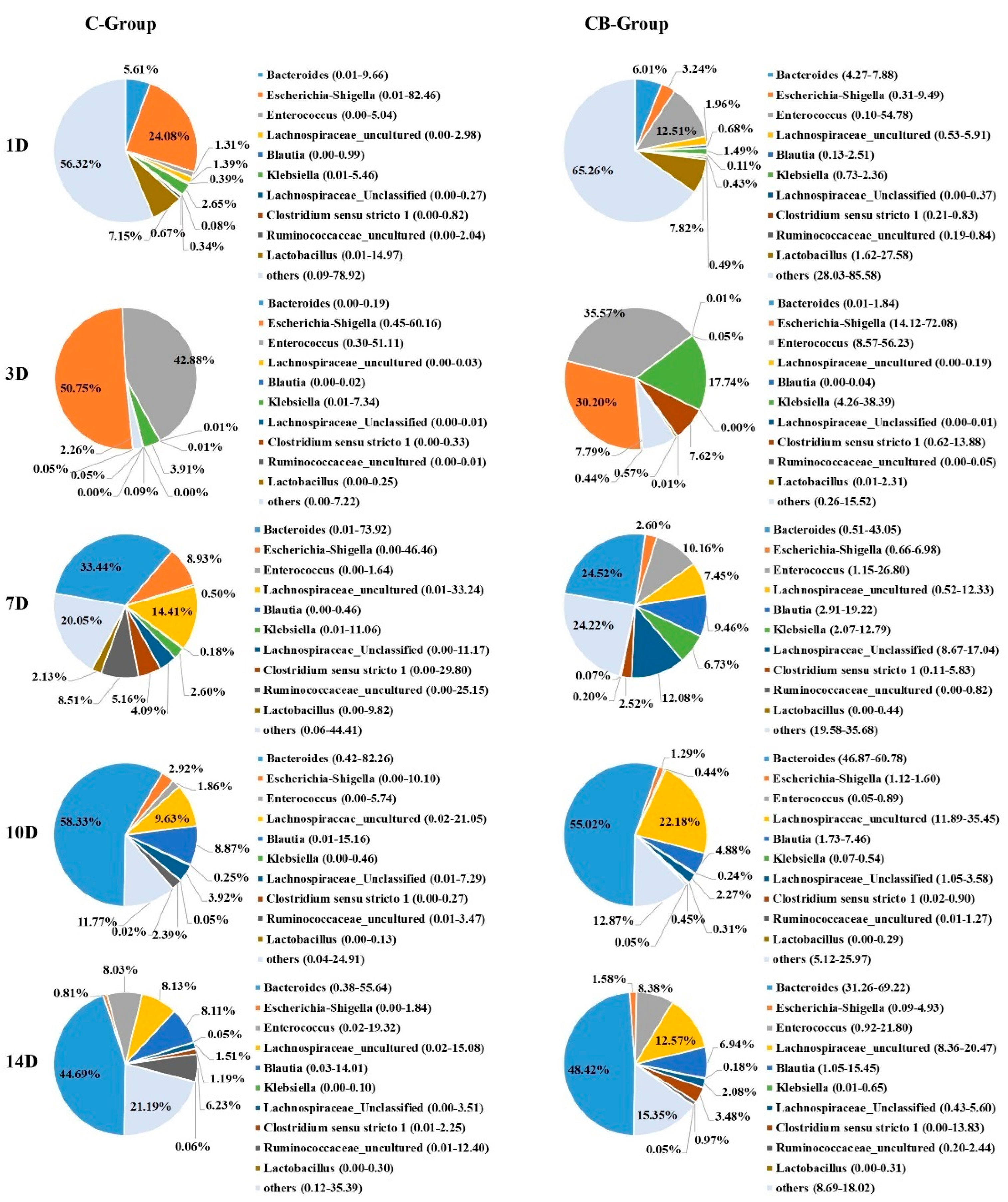
2.3. Cecum Microbiota Composition
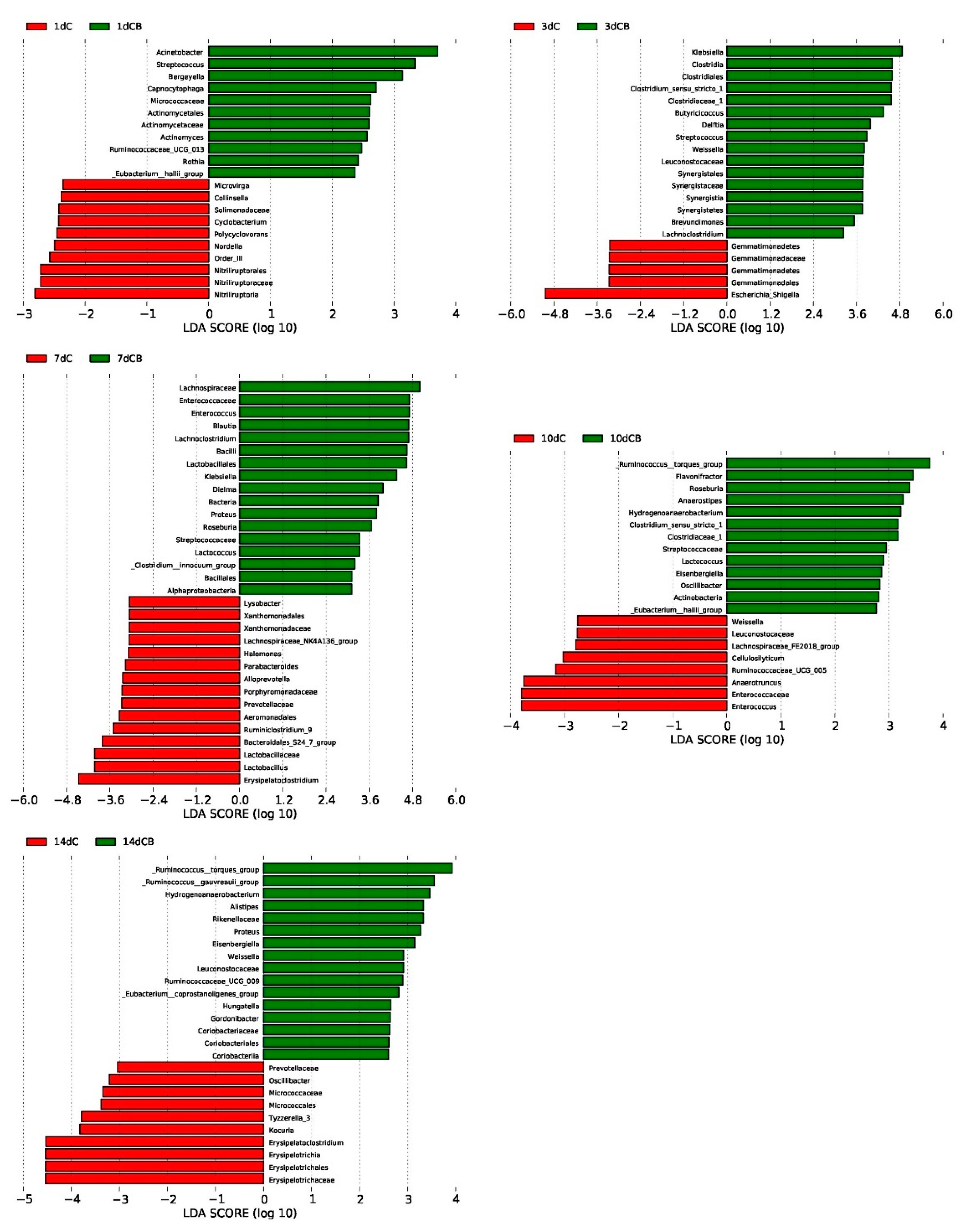
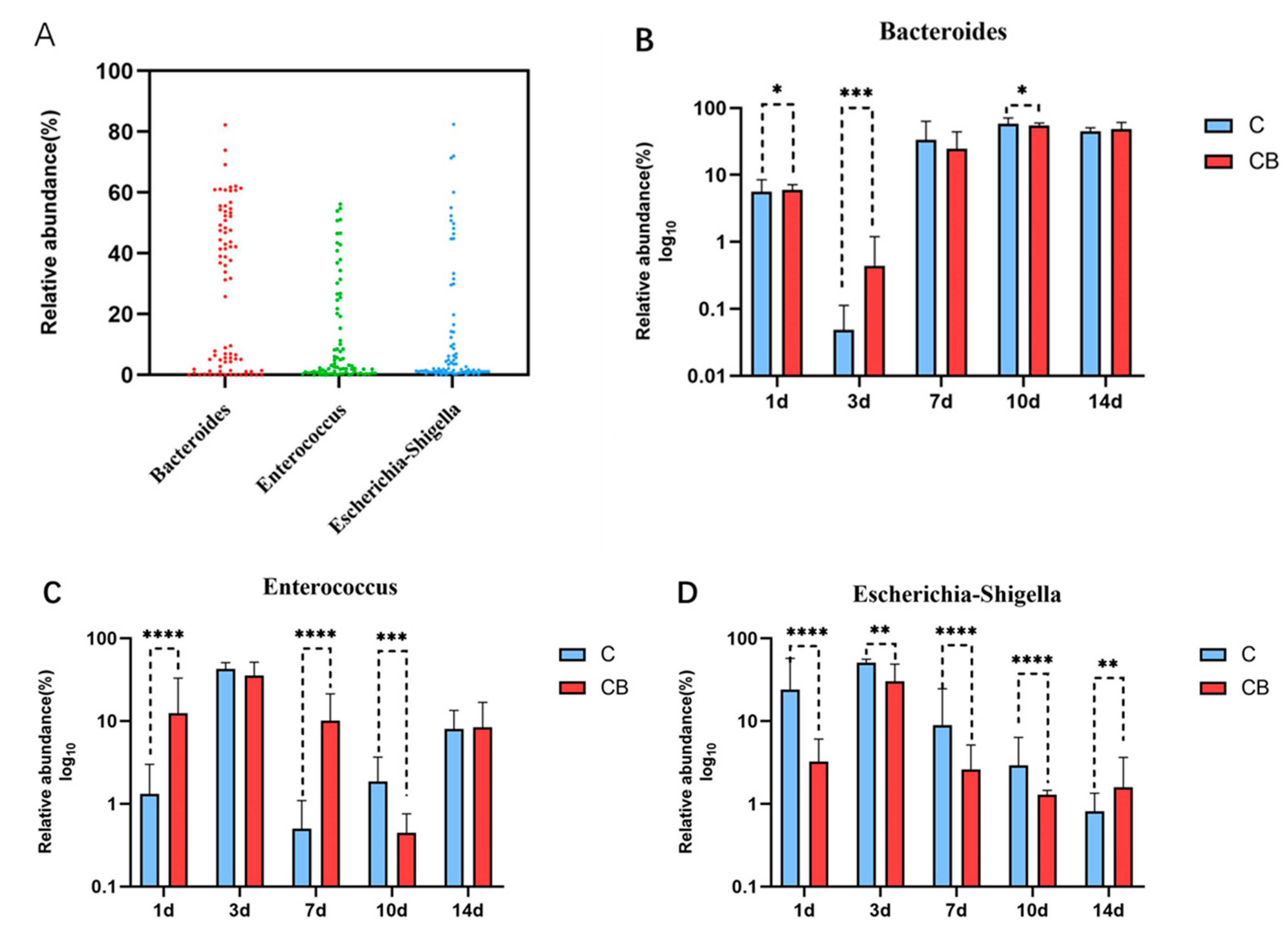
2.4. Core Microbial Genera in the Cecum Contents of the Ducks
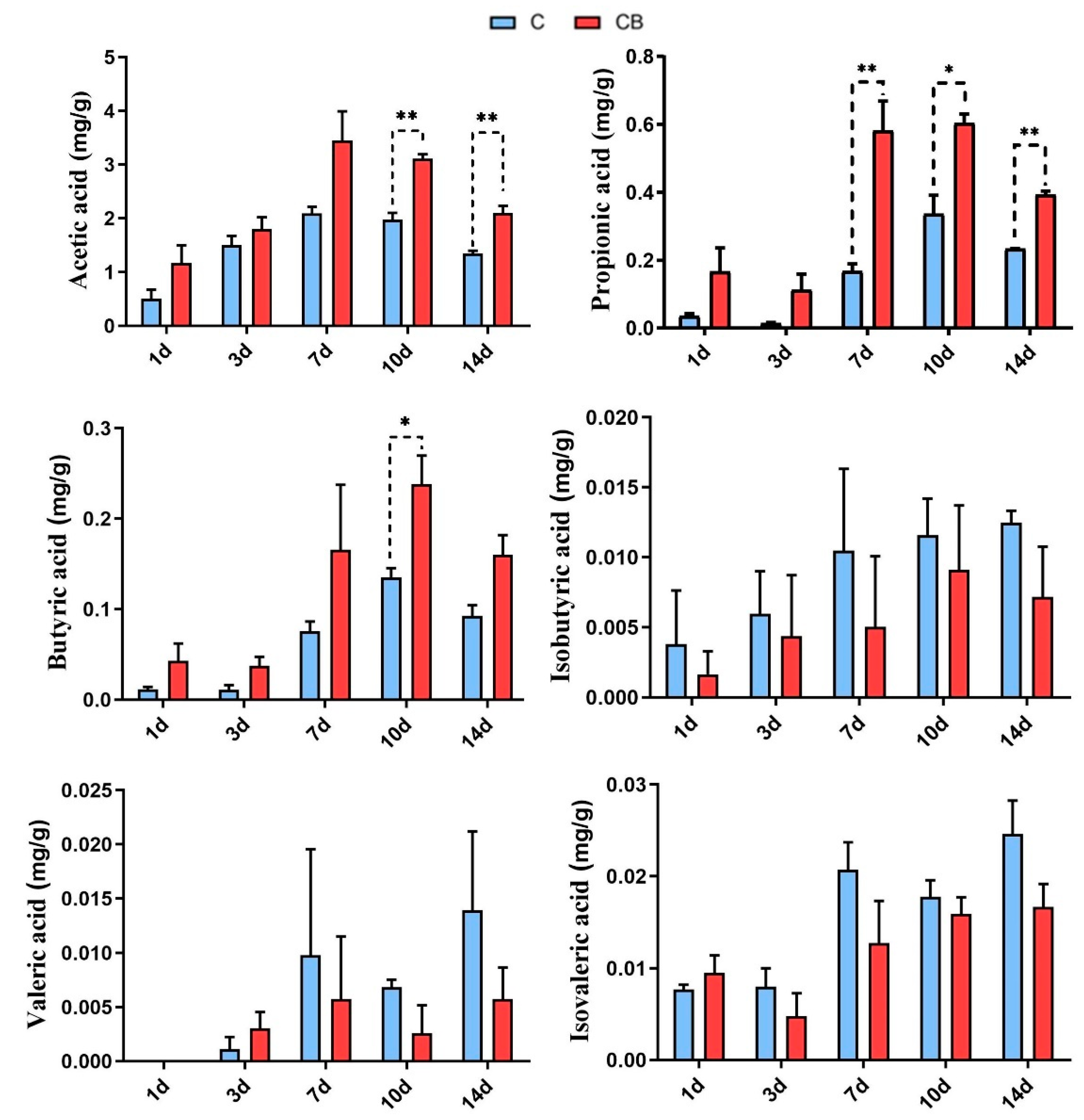
2.5. Short-Chain Fatty Acid Levels in Cecum
3. Discussion
4. Materials and Methods
4.1. Animal Experimental Design
4.2. Sample Collection
4.3. DNA Extraction and Purification
4.4. 16S rRNA Gene Sequencing and Data Processing
4.5. Short-Chain Fatty Acids Analysis
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.; Li, Y. Dietary supplementation with Clostridium butyricum modulates serum lipid metabolism, meat quality, and the amino acid and fatty acid composition of Peking ducks. Poult Sci. 2018, 97, 3218–3229. [Google Scholar] [CrossRef]
- Bacanlı, M.; Başaran, N. Importance of antibiotic residues in animal food. Food Chem Toxicol. 2019, 125, 462–466. [Google Scholar] [CrossRef]
- Oelschlaeger, T.A. Mechanisms of probiotic actions—A review. Int. J. Med Microbiol. 2010, 300, 57–62. [Google Scholar] [CrossRef]
- Jha, R.; Das, R. Probiotics (Direct-Fed Microbials) in Poultry Nutrition and Their Effects on Nutrient Utilization, Growth and Laying Performance, and Gut Health: A Systematic Review. Animals 2020, 10, 1863. [Google Scholar] [CrossRef]
- Kokoszyński, D.; Wilkanowska, A. Comparison of some meat quality and liver characteristics in Muscovy and mule ducks. Arch. Anim. Breed. 2020, 63, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Lyu, W.; Liu, X. Cecal Microbiota Modulates Fat Deposition in Muscovy Ducks. Front. Vet. Sci. 2021, 8, 609348. [Google Scholar] [CrossRef]
- Cassir, N.; Benamar, S. Clostridium butyricum: From beneficial to a new emerging pathogen. Clin. Microbiol. Infect. 2016, 22, 37–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Jia, Y. RNA-Seq transcriptome analysis of breast muscle in Pekin ducks supplemented with the dietary probiotic Clostridium butyricum. BMC Genom. 2018, 19, 844. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Cao, G. Effects of Clostridium butyricum and Enterococcus faecalis on growth performance, immune function, intestinal morphology, volatile fatty acids, and intestinal flora in a piglet model. Food Funct. 2019, 10, 7844–7854. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Tao, F. Positive effects of a Clostridium butyricum-based compound probiotic on growth performance, immune responses, intestinal morphology, hypothalamic neurotransmitters, and colonic microbiota in weaned piglets. Food Funct. 2019, 10, 2926–2934. [Google Scholar] [CrossRef] [PubMed]
- Molnár, A.; Such, N. Effects of Wheat Bran and Clostridium butyricum Supplementation on Cecal Microbiota, Short-Chain Fatty Acid Concentration, pH and Histomorphometry in Broiler Chickens. Animals 2020, 10, 2230. [Google Scholar] [CrossRef]
- Liao, X.; Wu, R. Effects of Clostridium butyricum on antioxidant properties, meat quality and fatty acid composition of broiler birds. Lipids Health Dis. 2015, 14, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Zhang, Y. Effect of dietary Clostridium butyricum on growth, intestine health status and resistance to ammonia stress in Pacific white shrimp Litopenaeus vannamei. Fish Shellfish. Immunol. 2017, 65, 25–33. [Google Scholar] [CrossRef]
- Kanai, T.; Mikami, Y.; Hayashi, A. A breakthrough in probiotics: Clostridium butyricum regulates gut homeostasis and anti-inflammatory response in inflammatory bowel disease. J. Gastroenterol. 2015, 50, 928–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, D.R.; Briggs, W. Cecal microbiome composition and metabolic function in probiotic treated broilers. PLoS ONE 2020, 15, e0225921. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, J. Clostridium butyricum Ameliorates Salmonella Enteritis Induced Inflammation by Enhancing and Improving Immunity of the Intestinal Epithelial Barrier at the Intestinal Mucosal Level. Front. Microbiol. 2020, 11, 299. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Q.; Wu, X. Early-Life Intervention Using Fecal Microbiota Combined with Probiotics Promotes Gut Microbiota Maturation, Regulates Immune System Development, and Alleviates Weaning Stress in Piglets. Int. J. Mol. Sci. 2020, 21, 503. [Google Scholar] [CrossRef] [Green Version]
- Hagihara, M.; Kuroki, Y. Clostridium butyricum Modulates the Microbiome to Protect Intestinal Barrier Function in Mice with Antibiotic-Induced Dysbiosis. iScience 2020, 23, 100772. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xiao, Y. Transcriptome Analysis Reveals the Genes Involved in Growth and Metabolism in Muscovy Ducks. BioMed Res. Int. 2021, 2021, 6648435. [Google Scholar]
- Zhang, L.; Zhang, L. Effects of dietary supplementation of probiotic, Clostridium butyricum, on growth performance, immune response, intestinal barrier function, and digestive enzyme activity in broiler chickens challenged with Escherichia coli K88. J. Anim. Sci. Biotechnol. 2016, 7, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Peng, X.Y. The Effect of Clostridium butyricum on Gut Microbiota, Immune Response and Intestinal Barrier Function During the Development of Necrotic Enteritis in Chickens. Front Microbiol. 2019, 10, 2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, J.J.; Bobeck, E.A. Eggshell and environmental bacteria contribute to the intestinal microbiota of growing chickens. J. Anim. Sci. Biotechnol. 2020, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tsai, T. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Ji, H. Changes in the diversity and composition of gut microbiota of weaned piglets after oral administration of Lactobacillus or an antibiotic. Appl. Microbiol. Biotechnol. 2016, 100, 10081–10093. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, G. Effects of rhamnolipids on growth performance and intestinal health parameters in Linnan yellow broilers. Poult. Sci. 2021, 100, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Yang, H. Early Intervention with Cecal Fermentation Broth Regulates the Colonization and Development of Gut Microbiota in Broiler Chickens. Front. Microbiol. 2019, 10, 1422. [Google Scholar] [CrossRef]
- Yang, H.; Lyu, W. Biogeography of microbiome and short-chain fatty acids in the gastrointestinal tract of duck. Poult. Sci. 2020, 99, 4016–4027. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, K. Comparison of the Intestinal Microbial Community in Ducks Reared Differently through High-Throughput Sequencing. BioMed Res. Int. 2019, 2019, 9015054. [Google Scholar] [CrossRef]
- Zhao, L.L.; Yin, H.C. Application of high-throughput sequencing for microbial diversity detection in feces of specific-pathogen-free ducks. Poult. Sci. 2018, 97, 2278–2286. [Google Scholar] [CrossRef]
- Zhu, C.; Song, W. Analysis of microbial diversity and composition in small intestine during different development times in ducks. Poult. Sci. 2020, 99, 1096–1106. [Google Scholar] [CrossRef]
- Wexler, A.G.; Goodman, A.L. An insider’s perspective: Bacteroides as a window into the microbiome. Nat. Microbiol. 2017, 2, 17026. [Google Scholar] [CrossRef] [Green Version]
- Rizzatti, G.; Lopetuso, L.R. Proteobacteria: A Common Factor in Human Diseases. BioMed Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.; Crouse, A. Enterobacteria and host resistance to infection. Mamm. Genome. 2018, 29, 558–576. [Google Scholar] [CrossRef]
- Rodrigues, D.R.; Winson, E. Intestinal Pioneer Colonizers as Drivers of Ileal Microbial Composition and Diversity of Broiler Chickens. Front. Microbiol. 2019, 10, 2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Wang, X. Change of intestinal microbiota in cerebral ischemic stroke patients. BMC Microbiol. 2019, 19, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamage, H.; Tetu, S.G. Fiber Supplements Derived from Sugarcane Stem, Wheat Dextrin and Psyllium Husk Have Different In Vitro Effects on the Human Gut Microbiota. Front. Microbiol. 2018, 9, 1618. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawley, B.; Tannock, G.W. Analysis of 16S rRNA Gene Amplicon Sequences Using the QIIME Software Package. Methods Mol Biol. 2017, 1537, 153–163. [Google Scholar]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Contents |
|---|---|
| Ingredients (air-dried basis, %) | |
| Corn | 54.50 |
| Soybean meal | 21.00 |
| Wheat middlings | 10.00 |
| DDGS | 4.50 |
| Fish meal | 3.00 |
| Rapeseed meal | 3.00 |
| Soybean oil | 1.50 |
| Dicalcium phosphate | 0.75 |
| Methionine | 0.23 |
| Lysine | 0.32 |
| Salt | 0.20 |
| Premix 1 | 1.00 |
| Nutrient Content 2 | |
| Crude protein, % | 19.50 |
| Metabolizable energy, MJ/kg | 12.12 |
| Lysine, % | 0.95 |
| Methionine + cysteine, % | 0.68 |
| Ca, % | 0.86 |
| Total phosphorus, % | 0.40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, X.; Fu, Z.; Li, N.; Yang, H.; Wang, W.; Lyu, W. Modulation of the Intestinal Microbiota by the Early Intervention with Clostridium Butyricum in Muscovy Ducks. Antibiotics 2021, 10, 826. https://doi.org/10.3390/antibiotics10070826
Xiao X, Fu Z, Li N, Yang H, Wang W, Lyu W. Modulation of the Intestinal Microbiota by the Early Intervention with Clostridium Butyricum in Muscovy Ducks. Antibiotics. 2021; 10(7):826. https://doi.org/10.3390/antibiotics10070826
Chicago/Turabian StyleXiao, Xingning, Zixian Fu, Na Li, Hua Yang, Wen Wang, and Wentao Lyu. 2021. "Modulation of the Intestinal Microbiota by the Early Intervention with Clostridium Butyricum in Muscovy Ducks" Antibiotics 10, no. 7: 826. https://doi.org/10.3390/antibiotics10070826
APA StyleXiao, X., Fu, Z., Li, N., Yang, H., Wang, W., & Lyu, W. (2021). Modulation of the Intestinal Microbiota by the Early Intervention with Clostridium Butyricum in Muscovy Ducks. Antibiotics, 10(7), 826. https://doi.org/10.3390/antibiotics10070826