
The Antimicrobial Activity of Omiganan Alone and In Combination against Candida Isolated from Vulvovaginal Candidiasis and Bloodstream Infections

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Peptide Synthesis
2.3. Antimicrobial Activity
3. Results
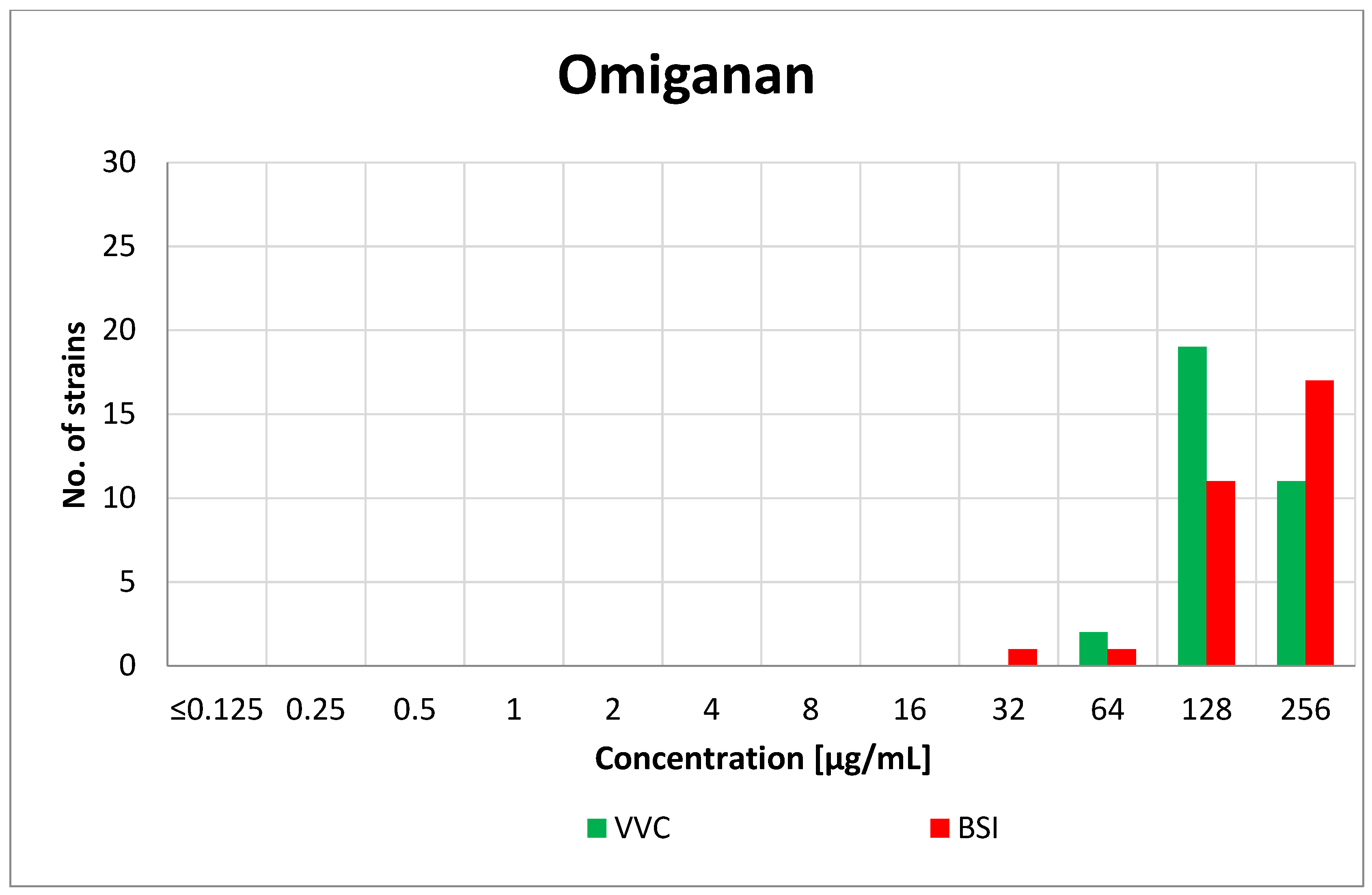
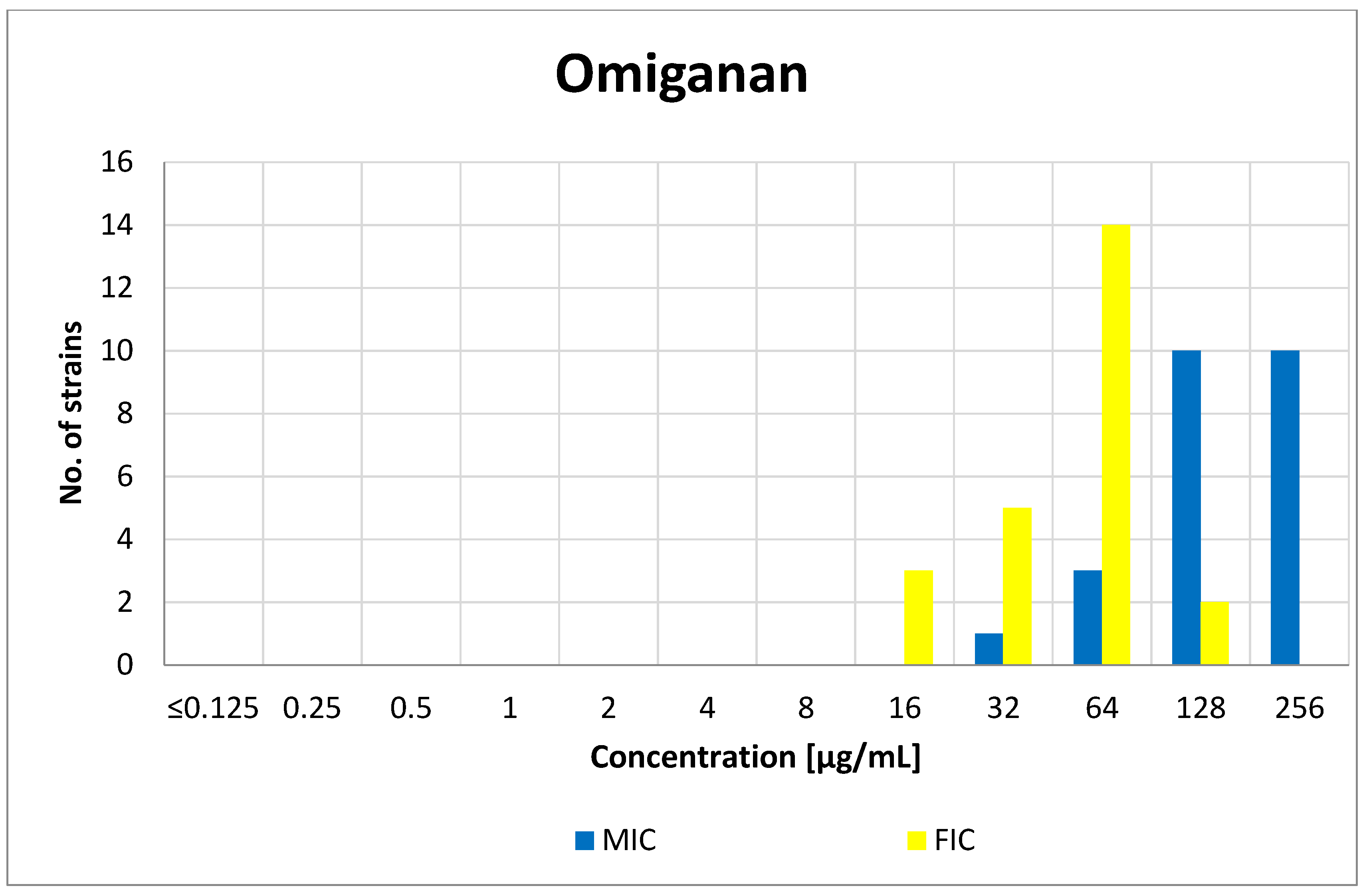
3.1. Minimum Inhibitory Concentration (MIC)
3.2. Minimal Biofilm Eradication Concentration (MBEC)
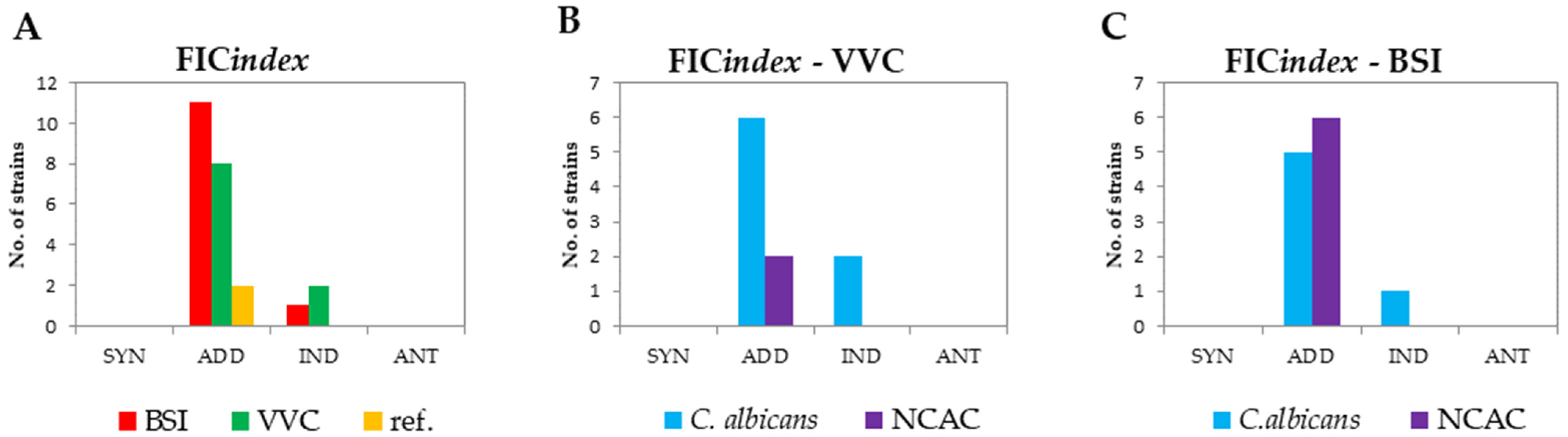
3.3. FIC Index
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naglik, J.R.; Challacombe, S.J.; Hube, B. Candida albicans Secreted Aspartyl Proteinases in Virulence and Pathogenesis. Microbiol. Mol. Biol. Rev. 2003, 67, 400–428. [Google Scholar] [CrossRef] [Green Version]
- Staniszewska, M.; Bondaryk, M.; Piłat, J.; Siennicka, K.; Magda, U.; Kurzatkowski, W. Virulence factors of Candida albicans. Przegl Epidemiol. 2012, 66, 629–633. [Google Scholar]
- Sidrim, J.; Júlio, C.; Rocha, M.F.G. Micologia Médica à Luz de Autores Conteponrâneos; Guanabara Koogan: Rio de Janeiro, Brazil, 2012. [Google Scholar]
- Achkar, J.M.; Fries, B.C. Candida infections of the genitourinary tract. Clin. Microbiol. Rev. 2010, 23, 253–273. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.R.; Klink, K.; Cohrssen, A. Evaluation of Vaginal Complaints. J. Am. Med. Assoc. 2004, 291, 1368–1379. [Google Scholar] [CrossRef]
- Sobel, J.D. Vulvovaginal candidosis. Lancet 2007, 369, 1961–1971. [Google Scholar] [CrossRef]
- Denning, D.W.; Kneale, M.; Sobel, J.D.; Rautemaa-Richardson, R. Global burden of recurrent vulvovaginal candidiasis: A systematic review. Lancet Infect. Dis. 2018, 18, e339–e347. [Google Scholar] [CrossRef]
- Sobel, J.D.; Sobel, R. Current treatment options for vulvovaginal candidiasis caused by azole-resistant Candida species. Expert Opin. Pharmacother. 2018, 19, 971–977. [Google Scholar] [CrossRef]
- Patel, D.A.; Gillespie, B.; Sobel, J.D.; Leaman, D.; Nyirjesy, P.; Weitz, M.V.; Foxman, B. Risk factors for recurrent vulvovaginal candidiasis in women receiving maintenance antifungal therapy: Results of a prospective cohort study. Am. J. Obstet. Gynecol. 2004, 190, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Ferreira, C.; Alves, C.T.; Henriques, M.; Azeredo, J.; Silva, S. Vulvovaginal candidiasis: Epidemiology, microbiology and risk factors. Crit. Rev. Microbiol. 2016, 42, 905–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amouri, I.; Sellami, H.; Borji, N.; Abbes, S.; Sellami, A.; Cheikhrouhou, F.; Maazoun, L.; Khaled, S.; Khrouf, S.; Boujelben, Y.; et al. Epidemiological survey of vulvovaginal candidosis in Sfax, Tunisia. Mycoses 2011, 54, e499–e505. [Google Scholar] [CrossRef]
- Brandolt, T.M.; Klafke, G.B.; Gonçalves, C.V.; Bitencourt, L.R.; de Martinez, A.M.B.; Mendes, J.F.; Meireles, M.C.A.; Xavier, M.O. Prevalence of Candida spp. in cervical-vaginal samples and the in vitro susceptibility of isolates. Braz. J. Microbiol. 2017, 48, 145–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura-Vasconcelos, S.S.; Fiorini, A.; Zanni, P.D.; de Souza Bonfim-Mendonça, P.; Godoy, J.R.; Almeida-Apolonio, A.A.; Consolaro, M.E.L.; Svidzinski, T.I.E. Emergence of Candida glabrata in vulvovaginal candidiasis should be attributed to selective pressure or virulence ability? Arch. Gynecol. Obstet. 2017, 296, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Sobel, J.D.; Kapernick, P.S.; Zervos, M.; Reed, B.D.; Hooton, T.; Soper, D.; Nyirjesy, P.; Heine, M.W.; Willems, J.; Panzer, H.; et al. Treatment of complicated Candida vaginitis: Comparison of single and sequential doses of fluconazole. Am. J. Obstet. Gynecol. 2001, 185, 363–369. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Liu, J.H.; Liu, F.D.; Xia, Y.H.; Wang, J.; Liu, X.; Zhang, Z.Q.; Zhu, N.; Ying, Y.; Huang, X.T. Vulvovaginal candidiasis: Species distribution, fluconazole resistance and drug efflux pump gene overexpression. Mycoses 2014, 57, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Puumala, E.; Robbins, N.; Cowen, L.E. Antifungal Drug Resistance: Molecular Mechanisms in Candida albicans and beyond. Chem. Rev. 2021, 121, 3390–3411. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Sobel, J.D.; White, T.C. A combination fluorescence assay demonstrates increased efflux pump activity as a resistance mechanism in azole-resistant vaginal Candida albicans isolates. Antimicrob. Agents Chemother. 2016, 60, 5858–5866. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and mechanisms of antifungal resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef]
- Brandt, M.E. Candida and Candidiasis. Emerg. Infect. Dis. 2002, 8, 876. [Google Scholar] [CrossRef]
- Enoch, D.A.; Ludlam, H.A.; Brown, N.M. Invasive fungal infections: A review of epidemiology and management options. J. Med. Microbiol. 2006, 55, 809–818. [Google Scholar] [CrossRef]
- Schelenz, S. Management of candidiasis in the intensive care unit. J. Antimicrob. Chemother. 2008, 61, 31–34. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, S.N.; Blijlevens, N.M.A.; Mahfouz, T.H.; Anaissie, E.J. Infections in patients with hematological cancer: Recent developments. Hematol. Am. Soc. Hematol. Educ. Program. 2003, 438–472. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowska, M.; Sienkiewicz, M.; Kwiatkowski, P.; Dąbrowski, M. Diagnosis and treatment of invasive Candida infections—A review article. Ann. Univ. Mariae Curie Sklodowska Sect. C Biol. 2019, 73, 47. [Google Scholar] [CrossRef]
- Garczewska, B.; Kamińska, W.; Dzierzanowska, D. Phenotype and genotype characterization of Candida albicans strains isolated from patients hospitalized at the Children’s Memorial Health Institute. Med. Dosw. Mikrobiol. 2008, 60, 231–241. [Google Scholar]
- Barbara Modrzewska, P.K.D. Invasion of host cells by fungi progresses in several phases. Polish Parasitol. Soc. 2013, 59, 57–66. [Google Scholar]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garipov, M.R.; Sabirova, A.E.; Pavelyev, R.S.; Shtyrlin, N.V.; Lisovskaya, S.A.; Bondar, O.V.; Laikov, A.V.; Romanova, J.G.; Bogachev, M.I.; Kayumov, A.R.; et al. Targeting pathogenic fungi, bacteria and fungal-bacterial biofilms by newly synthesized quaternary ammonium derivative of pyridoxine and terbinafine with dual action profile. Bioorg. Chem. 2020, 104. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Ozhegov, G.D.; Sabirova, A.E.; Novikova, V.V.; Lisovskaya, S.A.; Khabibrakhmanova, A.M.; Kurbangalieva, A.R.; Bogachev, M.I.; Kayumov, A.R. Increasing Susceptibility of Drug-Resistant Candida albicans to Fluconazole and Terbinafine by 2(5H)-Furanone Derivative. Molecules 2020, 25, 642. [Google Scholar] [CrossRef] [Green Version]
- Peschel, A.; Vincent Collins, L. Staphylococcal resistance to antimicrobial peptides of mammalian and bacterial origin. Peptides 2001, 22, 1651–1659. [Google Scholar] [CrossRef]
- Zasloff, M. Reconstructing one of nature’s designs. Trends Pharmacol. Sci. 2000, 21, 236–238. [Google Scholar] [CrossRef]
- Sader, H.S.; Fedler, K.A.; Rennie, R.P.; Stevens, S.; Jones, R.N. Omiganan pentahydrochloride (MBI 226), a topical 12-amino-acid cationic peptide: Spectrum of antimicrobial activity and measurements of bactericidal activity. Antimicrob. Agents Chemother. 2004, 48, 3112–3118. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.B.; Seo, J. Antimicrobial peptides under clinical investigation. Pept. Sci. 2019, 111, e24122. [Google Scholar] [CrossRef]
- Melo, M.N.; Dugourd, D.; Castanho, M.A.R.B. Omiganan Pentahydrochloride in the Front Line of Clinical Applications of Antimicrobial Peptides. Recent Pat. Antiinfect. Drug Discov. 2006, 1, 201–207. [Google Scholar] [CrossRef]
- Neubauer, D.; Jaśkiewicz, M.; Migoń, D.; Bauer, M.; Sikora, K.; Sikorska, E.; Kamysz, E.; Kamysz, W. Retro analog concept: Comparative study on physico-chemical and biological properties of selected antimicrobial peptides. Amino Acids 2017, 49, 1755–1771. [Google Scholar] [CrossRef] [Green Version]
- Rijsbergen, M.; Rijneveld, R.; Todd, M.; Feiss, G.L.; Kouwenhoven, S.T.P.; Quint, K.D.; Alewijk, D.C.J.G.; Koning, M.N.C.; Klaassen, E.S.; Burggraaf, J.; et al. Results of phase 2 trials exploring the safety and efficacy of omiganan in patients with human papillomavirus-induced genital lesions. Br. J. Clin. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Ng, S.M.S.; Yap, Y.Y.A.; Cheong, J.W.D.; Ng, F.M.; Lau, Q.Y.; Barkham, T.; Teo, J.W.P.; Hill, J.; Chia, C.S.B. Antifungal peptides: A potential new class of antifungals for treating vulvovaginal candidiasis caused by fluconazole-resistant Candida albicans. J. Pept. Sci. 2017, 23, 215–221. [Google Scholar] [CrossRef]
- Rubinchik, E.; Dugourd, D.; Algara, T.; Pasetka, C.; Friedland, H.D. Antimicrobial and antifungal activities of a novel cationic antimicrobial peptide, omiganan, in experimental skin colonisation models. Int. J. Antimicrob. Agents 2009, 34, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Czechowicz, P.; Jaśkiewicz, M.; Neubauer, D.; Gościniak, G.; Kamysz, W. Anticandidal Activity of Omiganan and Its Retro Analog Alone and in Combination with Fluconazole. Probiotics Antimicrob. Proteins 2021, 13, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standards-Second Edition, in CLSI Document M27-2A 2002; CLSI: Wayne, PA, USA, 2002; Volume 22, ISBN 1562384694. [Google Scholar]
- Krom, B.P.; Cohen, J.B.; McElhaney-Feser, G.; Busscher, H.J.; Van Der Mei, H.C.; Cihlar, R.L. Conditions for optimal Candida biofilm development in microtiter plates. Methods Mol. Biol. 2009, 499, 55–62. [Google Scholar] [CrossRef]
- Nilius, A.M. Antimicrobial Pharmacodynamics in Theory and Clinical Practice; Informa Healthcare USA: New York, NY, USA, 2002. [Google Scholar]
- Jaśkiewicz, M.; Neubauer, D.; Kazor, K.; Bartoszewska, S.; Kamysz, W. Antimicrobial activity of selected antimicrobial peptides against planktonic culture and biofilm of Acinetobacter baumannii. Probiotics Antimicrob. Proteins 2019, 11, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, M.J.; Edwards, J.R.; Culver, D.H.; Gaynes, R.P. Nosocomial Infections in Combined Medical-Surgical Intensive Care Units in the United States. Infect. Control. Hosp. Epidemiol. 2000, 21, 510–515. [Google Scholar] [CrossRef]
- Fridkin, S.K.; Jarvis, W.R. Epidemiology of nosocomial fungal infections. Clin. Microbiol. Rev. 1996, 9, 499–511. [Google Scholar] [CrossRef]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Cerdeira, C.; Gregorio, M.C.; Molares-Vila, A.; López-Barcenas, A.; Fabbrocini, G.; Bardhi, B.; Sinani, A.; Sánchez-Blanco, E.; Arenas-Guzmán, R.; Hernandez-Castro, R. Biofilms and vulvovaginal candidiasis. Colloids Surf. B Biointerfaces 2019, 174, 110–125. [Google Scholar] [CrossRef]
- Chow, J.K.; Golan, Y.; Ruthazer, R.; Karchmer, A.W.; Carmeli, Y.; Lichtenberg, D.; Chawla, V.; Young, J.; Hadley, S. Factors associated with candidemia caused by non-albicans Candida species versus Candida albicans in the intensive care unit. Clin. Infect. Dis. 2008, 46, 1206–1213. [Google Scholar] [CrossRef] [Green Version]
- Ostrosky-Zeichner, L.; Pappas, P.G. Invasive candidiasis in the intensive care unit. Crit. Care Med. 2006, 34, 857–863. [Google Scholar] [CrossRef]
- Parmeland, L.; Gazon, M.; Guerin, C.; Argaud, L.; Lehot, J.J.; Bastien, O.; Allaouchiche, B.; Michallet, M.; Picot, S.; Bienvenu, A.L.; et al. Candida albicans and non-Candida albicans fungemia in an institutional hospital during a decade. Med. Mycol. 2013, 51, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Prim. 2018, 4, 18026. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chander, J.; Singla, N.; Sidhu, S.K.; Gombar, S. Epidemiology of Candida blood stream infections: Experience of a tertiary care centre in North India. J. Infect. Dev. Ctries. 2013, 7, 670–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, B.M.; Yano, J.; Noverr, M.C.; Fidel, P.L. Candida Vaginitis: When Opportunism Knocks, the Host Responds. PLoS Pathog. 2014, 10, e1003965. [Google Scholar] [CrossRef] [Green Version]
- Robbins, N.; Uppuluri, P.; Nett, J.; Rajendran, R.; Ramage, G.; Lopez-Ribot, J.L.; Andes, D.; Cowen, L.E. Hsp90 Governs Dispersion and Drug Resistance of Fungal Biofilms. PLoS Pathog. 2011, 7, e1002257. [Google Scholar] [CrossRef]
- Ciszewski, M.; Czekaj, T. Grzybicze zakażenia szpitalne-narastające zagrożenie. Nowa Med. 2014, 2, 73–76. [Google Scholar]
- Bondaryk, M.; Kurzaątkowski, W.; Staniszewska, M. Antifungal agents commonly used in the superficial and mucosal candidiasis treatment: Mode of action and resistance development. Postep. Dermatol. Alergol. 2013, 30, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Reboli, A.C.; Rotstein, C.; Pappas, P.G.; Chapman, S.W.; Kett, D.H.; Kumar, D.; Betts, R.; Wible, M.; Goldstein, B.P.; Schranz, J.; et al. Anidulafungin versus Fluconazole for Invasive Candidiasis. N. Engl. J. Med. 2007, 356, 2472–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiolo, E.M.; Tafin, U.F.; Borens, O.; Trampuz, A. Activities of fluconazole, caspofungin, anidulafungin, and amphotericin b on planktonic and biofilm Candida species determined by microcalorimetry. Antimicrob. Agents Chemother. 2014, 58, 2709–2717. [Google Scholar] [CrossRef] [Green Version]
- Czechowicz, P.; Nowicka, J. Antimicrobial activity of lipopeptides. Postep. Mikrobiol. 2018, 57, 213–226. [Google Scholar] [CrossRef]
- Tellapragada, C.; Eshwara, V.K.; Johar, R.; Shaw, T.; Malik, N.; Bhat, P.V.; Kamath, A.; Mukhopadhyay, C. Antifungal Susceptibility Patterns, In Vitro Production of Virulence Factors, and Evaluation of Diagnostic Modalities for the Speciation of Pathogenic Candida from Blood Stream Infections and Vulvovaginal Candidiasis. J. Pathog. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pahwa, N.; Kumar, R. Species distribution and drug susceptibility of Candida in clinical isolates from a tertiary care centre at Indore. Indian J. Med. Microbiol. 2014, 32, 44–48. [Google Scholar] [CrossRef]
- Tomaszewski, J. The statement of polish gynecological society experts on the etiology and treatment of recurrent vulvovaginal candidiasis. Ginekol. Pol. 2011, 82, 869–873. [Google Scholar]
- Sherry, L.; Kean, R.; McKloud, E.; O’Donnell, L.E.; Metcalfe, R.; Jones, B.L.; Ramage, G. Biofilms formed by isolates from recurrent vulvovaginal candidiasis patients are heterogeneous and insensitive to fluconazole. Antimicrob. Agents Chemother. 2017, 61, e01065-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venisse, N.; Grégoire, N.; Marliat, M.; Couet, W. Mechanism-based pharmacokinetic-pharmacodynamic models of in vitro fungistatic and fungicidal effects against Candida albicans. Antimicrob. Agents Chemother. 2008, 52, 937–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertas, A. Aktywność przeciwgrzybicza flukonazolu wobec klinicznych szczepów Candida albicans oraz innych Candida spp.—Przegląd badań in vitro przeprowadzonych w różnych ośrodkach medycznych the antifungal activity of fl uconazole against clinical of the researc. Ann. Acad. Med. Silesiensis 2011, 65, 29–37. [Google Scholar]
- Singh, K.; Rani, J. Sequential and Structural Aspects of Antifungal Peptides from Animals, Bacteria and Fungi Based on Bioinformatics Tools. Probiotics Antimicrob. Proteins 2016, 8, 85–101. [Google Scholar] [CrossRef]
- De Aguiar, F.L.L.; Cavalcante, C.; dos Santos Fontenelle, R.O.; Falcão, C.B.; Andreu, D.; Rádis-Baptista, G. The antiproliferative peptide Ctn[15–34] is active against multidrug-resistant yeasts Candida albicans and Cryptococcus neoformans. J. Appl. Microbiol. 2020, 128, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Dawgul, M.; Barańska-Rybak, W.; Bielińska, S.; Nowicki, R.; Kamysz, W. Wpływ peptydów przeciwdrobnoustrojowych na biofilm Candida The influence of antimicrobial peptides on Candida biofilm. Alerg. Astma Immunol. 2010, 15, 220–225. [Google Scholar]
- Sun, W.; Zhang, L.; Lu, X.; Feng, L.; Sun, S. The synergistic antifungal effects of sodium phenylbutyrate combined with azoles against Candida albicans via the regulation of the ras–cAMP–PKA signalling pathway and virulence. Can. J. Microbiol. 2019, 65, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Maurya, I.K.; Thota, C.K.; Sharma, J.; Tupe, S.G.; Chaudhary, P.; Singh, M.K.; Thakur, I.S.; Deshpande, M.; Prasad, R.; Chauhan, V.S. Mechanism of action of novel synthetic dodecapeptides against Candida albicans. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 5193–5203. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FIC Index | Interpretation |
|---|---|
| ≤0.5 | Synergism (SYN) |
| >0.5 to 1.0 | Addition (ADD) |
| >1.0 to ≤2.0 | Indifference (IND) |
| >2.0 | Antagonism (ANT) |
| Peptide | GM | IC50 [42] | SI |
|---|---|---|---|
| Omiganan | 166.43 | 79.39 | 0.48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żyrek, D.; Wajda, A.; Czechowicz, P.; Nowicka, J.; Jaśkiewicz, M.; Neubauer, D.; Kamysz, W. The Antimicrobial Activity of Omiganan Alone and In Combination against Candida Isolated from Vulvovaginal Candidiasis and Bloodstream Infections. Antibiotics 2021, 10, 1001. https://doi.org/10.3390/antibiotics10081001
Żyrek D, Wajda A, Czechowicz P, Nowicka J, Jaśkiewicz M, Neubauer D, Kamysz W. The Antimicrobial Activity of Omiganan Alone and In Combination against Candida Isolated from Vulvovaginal Candidiasis and Bloodstream Infections. Antibiotics. 2021; 10(8):1001. https://doi.org/10.3390/antibiotics10081001
Chicago/Turabian StyleŻyrek, Dawid, Andrzej Wajda, Paulina Czechowicz, Joanna Nowicka, Maciej Jaśkiewicz, Damian Neubauer, and Wojciech Kamysz. 2021. "The Antimicrobial Activity of Omiganan Alone and In Combination against Candida Isolated from Vulvovaginal Candidiasis and Bloodstream Infections" Antibiotics 10, no. 8: 1001. https://doi.org/10.3390/antibiotics10081001
APA StyleŻyrek, D., Wajda, A., Czechowicz, P., Nowicka, J., Jaśkiewicz, M., Neubauer, D., & Kamysz, W. (2021). The Antimicrobial Activity of Omiganan Alone and In Combination against Candida Isolated from Vulvovaginal Candidiasis and Bloodstream Infections. Antibiotics, 10(8), 1001. https://doi.org/10.3390/antibiotics10081001