
Environmental Surveillance and Characterization of Antibiotic Resistant Staphylococcus aureus at Coastal Beaches and Rivers on the Island of Hawaiʻi


Abstract
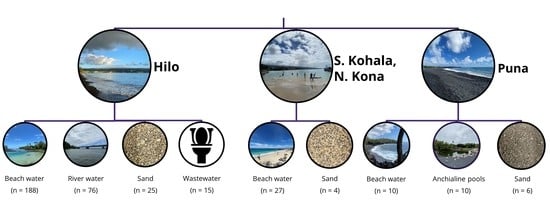
:
1. Introduction
2. Results
2.1. Prevalence of S. aureus
2.2. Genetic Characterization of MRSA and MSSA Isolates
2.3. Antibiotic Resistance Genes
2.4. Toxin, Exoenzyme, Host Immunity Genes
2.5. Plasmids
3. Discussion
4. Materials and Methods
4.1. Study Locations and Sampling
4.2. Isolation and Quantification of Staphylococci
4.3. WGS Assembly and Analysis
4.4. Prevalence of S. aureus
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monecke, S.; Gavier-Widén, D.; Hotzel, H.; Peters, M.; Guenther, S.; Lazaris, A.; Loncaric, I.; Müller, E.; Reissig, A.; Ruppelt-Lorz, A.; et al. Diversity of Staphylococcus aureus isolates in European wildlife. PLoS ONE 2016, 11, e0168433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challagundla, L.; Luo, X.; Tickler, I.A.; Didelot, X.; Coleman, D.C.; Shore, A.C.; Coombs, G.W.; Sordelli, D.O.; Brown, E.L.; Skov, R.; et al. Range expansion and the origin of USA300 North American epidemic methicillin-resistant Staphylococcus aureus. mBio 2018, 9, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Levin-Edens, E.; Meschke, J.S.; Roberts, M.C. Quantification of methicillin-resistant Staphylococcus aureus strains in marine and freshwater samples by the most-probable-number method. Appl. Environ. Microbiol. 2011, 77, 3541–3543. [Google Scholar] [CrossRef] [Green Version]
- Thapaliya, D.; Hellwig, E.J.; Kadariya, J.; Grenier, D.; Jefferson, A.J.; Dalman, M.; Kennedy, K.; DiPerna, M.; Orihill, A.; Taha, M.; et al. Prevalence and characterization of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus on public recreational beaches in Northeast Ohio. GeoHealth 2017, 1, 320–332. [Google Scholar] [CrossRef] [Green Version]
- Plano, L.R.W.; Shibata, T.; Garza, A.C.; Kish, J.; Fleisher, J.M.; Sinigalliano, C.D.; Gidley, M.L.; Withum, K.; Elmir, S.M.; Hower, S.; et al. Human-associated methicillin-resistant Staphylococcus aureus from a subtropical recreational marine beach. Microb. Ecol. 2013, 65, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Schulz, J.; Friese, A.; Klees, S.; Tenhagen, B.A.; Fetsch, A.; Rösler, U.; Hartung, J. Longitudinal study of the contamination of air and of soil surfaces in the vicinity of pig barns by livestock-associated methicillin-resistant Staphylococcus aureus. Appl. Environ. Microbiol. 2012, 78, 5666–5671. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, K.D.; McNay, M.; Cao, Y.; Ebentier, D.; Madison, M.; Griffith, J.F. A multi-beach study of Staphylococcus aureus, MRSA, and enterococci in seawater and beach sand. Water Res. 2012, 46, 4195–4207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plano, L.R.; Garza, A.C.; Shibata, T.; Elmir, S.M.; Kish, J.; Sinigalliano, C.D.; Gidley, M.L.; Miller, G.; Withum, K.; Fleming, L.E.; et al. Shedding of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus from adult and pediatric bathers in marine waters. BMC Microbiol. 2011, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Esiobu, N.; Green, M.; Echeverry, A.; Bonilla, T.D.; Stinson, C.M.; Hartz, A.; Rogerson, A.; McCorquodale, D.S. High numbers of Staphylococcus aureus at three bathing beaches in South Florida. Int. J. Environ. Health Res. 2013, 23, 46–57. [Google Scholar] [CrossRef]
- Levin-Edens, E.; Soge, O.O.; No, D.; Stiffarm, A.; Meschke, J.S.; Roberts, M.C. Methicillin-resistant Staphylococcus aureus from Northwest Marine and fresh water recreational beaches. FEMS Microbiol. Ecol. 2012, 79, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Soge, O.O.; Meschke, J.S.; No, D.B.; Roberts, M.C. Characterization of methicillin-resistant Staphylococcus aureus and methicillin-resistant coagulase-negative Staphylococcus spp. isolated from US West coast public marine beaches. J. Antimicrob. Chemother. 2009, 64, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, L.R.; Haack, S.K.; Johnson, H.E.; Brennan, A.K.; Isaacs, N.M.; Spencer, C. Staphylococcus aureus and methicillin-resistant S. aureus (MRSA) at ambient freshwater beaches. J. Water Health 2015, 13, 680–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estivariz, C.F.; Park, S.Y.; Hageman, J.C.; Dvorin, J.; Melish, M.M.; Arpon, R.; Coon, P.; Slavish, S.; Kim, M.; McDougal, L.K.; et al. Emergence of community-associated methicillin resistant Staphylococcus aureus in Hawaii, 2001–2003. J. Infect. 2007, 54, 349–357. [Google Scholar] [CrossRef]
- Viau, E.J.; Goodwin, K.D.; Yamahara, K.M.; Layton, B.A.; Sassoubre, L.M.; Burns, S.L.; Tong, H.-I.; Wong, S.H.C.; Lu, Y.; Boehm, A.B. Bacterial pathogens in Hawaiian coastal streams—Associations with fecal indicators, land cover, and water quality. Water Res. 2011, 45, 3279–3290. [Google Scholar] [CrossRef] [PubMed]
- Economy, L.M.; Wiegner, T.N.; Strauch, A.M.; Awaya, J.D.; Gerken, T. Rainfall and streamflow effects on estuarine Staphylococcus aureus and fecal indicator bacteria concentrations. J. Environ. Qual. 2019, 48, 1711–1721. [Google Scholar] [CrossRef]
- Fowler, T. Development of Methods Using CHROMagarTM Media to Determine the Prevalence of Staphylococcus aureus and Methicillin-Resistant S. aureus (MRSA) in Hawaiian Marine Recreational Waters. Master’s Thesis, University of Hawaiʻi at Mānoa, Honolulu, HI, USA, 2005. [Google Scholar]
- Klein, E.Y.; Mojica, N.; Jiang, W.; Cosgrove, S.E.; Septimus, E.; Morgan, D.J.; Laxminarayan, R. Trends in methicillin-resistant Staphylococcus aureus hospitalizations in the United States, 2010–2014. Clin. Infect. Dis. 2017, 65, 1921–1923. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, W.R.; Schlosser, J.; Chinn, R.Y.; Tweeten, S.; Jackson, M. National prevalence of methicillin-resistant Staphylococcus aureus in inpatients at US health care facilities, 2006. Am. J. Infect. Control 2007, 35, 631–637. [Google Scholar] [CrossRef]
- Jarvis, W.R.; Jarvis, A.A.; Chinn, R.Y. National prevalence of methicillin-resistant Staphylococcus aureus in inpatients at United States health care facilities, 2010. Am. J. Infect. Control 2012, 40, 194–200. [Google Scholar] [CrossRef]
- Li, F.; Park, S.Y.; Ayers, T.L.; Miller, F.D.; MacFadden, R.; Nakata, M.; Lee, M.C.; Effler, P.V. Methicillin-resistant Staphylococcus aureus, Hawaii, 2000–2002. Emerg. Infect. Dis. 2005, 11, 1205–1210. [Google Scholar] [CrossRef]
- Len, K.A.; Bergert, L.; Patel, S.; Melish, M.; Kimata, C.; Erdem, G. Community-acquired Staphylococcus aureus pneumonia among hospitalized children in Hawaii. Pediatr. Pulmonol. 2010, 45, 898–905. [Google Scholar] [CrossRef]
- Li, F.; Miller, F.D.; Effler, P.V. Epidemiology of methicillin-resistant Staphylococcus aureus among incarcerated population in Hawaiʻi, 2000–2005. Hawaii Med. J. 2010, 69, 99–102. [Google Scholar] [PubMed]
- Li, F.; Arnsberger, P.; Miller, F.D. Profile of methicillin-resistant Staphylococcus aureus among nursing home residents in Hawaiʻi. Hawaii Med. J. 2010, 69, 126–129. [Google Scholar]
- Charoenca, N.; Fujioka, R.S. Assessment of Staphylococcus bacteria in Hawaii’s marine recreational waters. Water Sci. Technol. 1993, 27, 283–289. [Google Scholar] [CrossRef]
- Cheung, W.H.S.; Chang, K.C.K.; Hung, R.P.S.; Kleevens, J.W.L. Health effects of beach water pollution in Hong Kong. Epidemiol. Infect. 1990, 105, 139–162. [Google Scholar] [CrossRef]
- Seifred, S.E.; Tice, A.D.; Eischen, M. Diversity of community associated strains of methicillin-resistant Staphylococcus aureus in Hawaii. J. Infect. Dis. 2007, 195, 305–307. [Google Scholar] [CrossRef] [PubMed]
- Charoenca, N.; Fujioka, R.S. Association of staphylococcal skin infections and swimming. Water Sci. Technol. 1995, 31, 11–17. [Google Scholar] [CrossRef]
- Strauß, L.; Stegger, M.; Akpaka, P.E.; Alabi, A.; Breurec, S.; Coombs, G.; Egyir, B.; Larsen, A.R.; Laurent, F.; Monecke, S.; et al. Origin, evolution, and global transmission of community-acquired Staphylococcus aureus ST8. Proc. Natl. Acad. Sci. USA 2017, 114, E10596–E10604. [Google Scholar] [CrossRef] [Green Version]
- Bothwell, N.E.; Shvidler, J.; Cable, B.B. Acute rise in methicillin-resistant Staphylococcus aureus infections in a coastal community. Otolaryngol. Neck Surg. 2007, 137, 942–946. [Google Scholar] [CrossRef]
- Early, G.J.; Seifried, S.E. Risk Factors for Community-associated Staphylococcus aureus skin infection in children of Maui. Hawaiʻi J. Med. Public Health 2012, 71, 218–223. [Google Scholar]
- Jay, J.; Rugh, M.; Burdick, B.; Diaz, M.; Hammett, M.; Merlos, F.; Molina, N.; Santana, J.; Smith, C. Prevalence of MRSA Colonization in Surfers Following Exposure in Select Southern California Coastal Waters; Institute of the Environment and Sustainability: Los Angeles, CA, USA, 2019. [Google Scholar]
- Butaye, P.; Argudín, M.A.; Smith, T.C. Livestock-associated MRSA and its current evolution. Curr. Clin. Microbiol. Rep. 2016, 3, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Chroboczek, T.; Boisset, S.; Rasigade, J.-P.; Tristan, A.; Bes, M.; Meugnier, H.; Vandenesch, F.; Etienne, J.; Laurent, F. Clonal complex 398 methicillin susceptible Staphylococcus aureus: A frequent unspecialized human pathogen with specific phenotypic and genotypic characteristics. PLoS ONE 2013, 8, e68462. [Google Scholar] [CrossRef]
- Jackson, K.A.; Gokhale, R.H.; Nadle, J.; Ray, S.M.; Dumyati, G.; Schaffner, W.; Ham, D.C.; Magill, S.S.; Lynfield, R.; See, I. Public health importance of invasive methicillin-sensitive Staphylococcus aureus Infections: Surveillance in 8 US Counties, 2016. Clin. Infect. Dis. 2020, 1021–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüger, H.; Ji, X.; Wang, Y.; Feßler, A.T.; Wang, Y.; Wu, C.; Schwarz, S. Identification of Tn553, a novel Tn554-related transposon that carries a complete blaZ-blaR1-blaI β-lactamase operon in Staphylococcus aureus. J. Antimicrob. Chemother. 2021, dkab210. [Google Scholar] [CrossRef]
- Gómez-Sanz, E.; Kadlec, K.; Feßler, A.T.; Zarazaga, M.; Torres, C.; Schwarz, S. Novel erm(T)-carrying multiresistance plasmids from porcine and human isolates of methicillin-resistant Staphylococcus aureus ST398 that also harbor cadmium and copper resistance determinants. Antimicrob. Agents Chemother. 2013, 57, 3275–3282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawaiʻi State Department of Health. Report to the Twenty-Ninth Legislature State of Hawaiʻi 2018 Regular Session Relating to Cesspools and Prioritization for Replacement; Hawaiʻi State Department of Health: Honolulu, HI, USA, 2017.
- Global Volcanism Program. Report on Kilauea (United States). Bull. Glob. Volcanism Netw. 2019, 44. [Google Scholar] [CrossRef]
- Myers, D.N.; Stoeckel, D.M.; Bushon, R.N.; Francy, D.S.; Brady, A.M.G. Chapter A7. Section 7.1. Fecal Indicator Bacteria; Techniques of Water-Resources Investigations; U.S. Geological Survey: Reston, VA, USA, 2014; p. 73. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A.; Prjibelsky, A.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling genomes and mini-metagenomes from highly chimeric reads. In Proceedings of the Research in Computational Molecular Biology, Beijing, China, 7–10 April 2013; pp. 158–170. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| District | Source | # Stations | # Quanti-Tray 2000® Samples | # MRSA Isolates | # MSSA Isolates | # S. aureus Isolates | # Quanti-Tray 2000® Samples Prevalence (%) | ||
|---|---|---|---|---|---|---|---|---|---|
| MRSA | MSSA | S. aureus | |||||||
| Hilo | Beaches | 14 | 188 | 2 | 17 | 19 | 2 (1.1) | 11 (5.9) | 13 (6.9) |
| Rivers/Stream | 7 | 76 | 0 | 3 | 3 | 0 (0.0) | 3 (3.9) | 3 (3.9) | |
| Sand | 5 * | 25 | 3 | 3 | 6 | 3 (12.0) | 2 (8.0) | 3 (12.0) | |
| Wastewater | N/A | 15 | 0 | 0 | 0 | 0 (0.0) | 0 (0.0) | 0 (0.0) | |
| S. Kohala, N. Kona | Beaches | 9 | 27 | 0 | 1 | 1 | 0 (0.0) | 1 (3.7) | 1 (3.7) |
| Sand | 4 * | 4 | 0 | 0 | 0 | 0 (0.0) | 0 (0.0) | 0 (0.0) | |
| Puna | Beaches | 2 | 10 | 3 | 0 | 3 | 2 (20.0) | 0 (0.0) | 2 (20.0) |
| Anchialine Pools | 4 | 10 | 1 | 3 | 4 | 1 (10.0) | 3 (30.0) | 4 (40.0) | |
| Sand | 2 * | 6 | 0 | 0 | 0 | 0 (0.0) | 0 (0.0) | 0 (0.0) | |
| Summary | |||||||||
| Hilo, S. Kohala, N. Kona, Puna | Beaches | 25 | 225 | 5 | 18 | 23 d | 4 (1.8) | 12 (5.3) | 16 (7.1) |
| Rivers/Stream | 7 | 76 | 0 | 3 | 3 | 0 (0.0) | 3 (3.9) | 3 (3.9) | |
| Anchialine Pools | 4 | 10 | 1 | 3 | 4 | 1 (10.0) | 3 (30.0) | 4 (40.0) | |
| Sand | 11 * | 35 | 3 | 3 | 6 e | 3 (8.6) | 2 (5.7) | 3 (8.6) | |
| Wastewater | N/A | 15 | 0 | 0 | 0 | 0 (0.0) | 0 (0.0) | 0 (0.0) | |
| Total | 36 | 361 | 9 a | 27 b | 36 c | 8 (2.2) | 20 (5.5) | 26 (7.2) | |
| Station | District | Source | Date | Isolate # | MRSA/MSSA | ST b | CC c | Antimicrobial Resistance d | Virulence Gene Profile e | Plasmids | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Toxin | Exoenzyme | Host Immunity | ||||||||||
| Lehia Beach Park (station#1) | Hilo | sand | 12/12/2020 | 108 | MRSA a | 8 f | 8 | aph(3’)-III, blaZ, erm(C), erm(T), mecA, mph(C), msr(A) | hlgA, hlgB, hlgC, lukD, lukE, lukF-PV, lukS-PV, sek, seq | aur, splA, splB, splE | ACME, sak, scn | rep10, rep19 |
| Richardson’s Beach Park (station #3) | Hilo | beach water | 09/04/2020 | 60 | MRSA a | 8 | 8 | aph(3’)-III, ant(6)-Ia, blaZ, erm(C), mecA, mph(C), msr(A) | hlgA, hlgB, hlgC, lukD, lukE, lukF-PV, lukS-PV, sek, seq | aur, splA, splB, splE | ACME, sak, scn | rep10, rep19 |
| Reed’s Bay (station#11) | Hilo | sand | 12/14/2020 | 105 | MRSA a | 8 | 8 | aph(3’)-III, blaZ, erm(C), mecA, mph(C), msr(A), tet(K) | hlgA, hlgB, hlgC, lukD, lukE, lukF-PV, lukS-PV, sek, seq | aur, splA, splB, splE | ACME, sak, scn | rep7a, rep7c, rep10, rep19 |
| Honoliʻi Beach Park (station #20) | Hilo | beach water | 08/15/2020 | 59 | MRSA a | 8 | 8 | aph(3’)-III, ant(6)-Ia, blaZ, erm(C), mecA, mph(C), msr(A) | hlgA, hlgB, hlgC, lukD, lukE, lukF-PV, lukS-PV, sek, seq | aur, splA, splB, splE | ACME, sak, scn | rep7c, rep10, rep19 |
| Honoliʻi Beach Park (station #20) | Hilo | sand | 12/14/2020 | 109 | MRSA a | 8 g | 8 | aph(3’)-III, ant(6)-Ia, blaZ, erm(C), mecA, mph(C), msr(A), qacG, tet(K) | hlgA, hlgB, hlgC, lukD, lukE, lukF-PV, lukS-PV, seg, sem, sen, sek, seq | aur, splA, splB, splE | ACME, sak, scn | rep7a, rep10, rep19 |
| Kehena (station#31) | Hilo | beach water | 08/20/2020 | 61 | MRSA a | 8 | 8 | aph(3’)-III, ant(6)-Ia, erm(C), mecA, mph(C), msr(A) | hlgA, hlgB, hlgC, lukD, lukE, lukF-PV, lukS-PV, sek, seq | aur, splA, splB, splE | ACME, sak, scn | rep7c, rep10, rep19 |
| Pohoiki Beach (station #32) | Puna | beach water | 08/20/2020 | 51 | MRSA a | 8 h | 8 | aph(3’)-III, ant(6)-Ia, blaZ, erm(C), mecA, mph(C), msr(A) | hlgA, hlgB, hlgC, lukD, lukE, lukF-PV, lukS-PV, sek, seq | aur, splA, splB, splE | ACME, sak, scn | rep10, rep19 |
| Pohoiki Beach (station #32) | Puna | beach water | 08/20/2020 | 88 | MRSA a | 8 h | 8 | aph(3’)-III, ant(6)-Ia, blaZ, erm(C), mecA, mph(C), msr(A), qacD | hlgA, hlgB, hlgC, lukD, lukE, lukF-PV, lukS-PV, sek, seq | aur, splA, splB, splE | ACME, sak, scn | rep10, rep19 |
| Pohoiki Harbor (station #33) | Puna | anchialine pool | 12/16/2020 | 89 | MRSA a | 8 | 8 | aph(3’)-III, ant(6)-Ia, blaZ, erm(C), lnu(A), mecA, mph(C), msr(A), qacD | hlgA, hlgB, hlgC, lukD, lukE, lukF-PV, lukS-PV, sek, seq | aur, splA, splB, splE | ACME, sak, scn | rep10, rep19, rep21 |
| Richardson’s Beach Park (station #2) | Hilo | beach water | 11/14/2020 | 47 | MSSA | 72 i | 8 | blaZ | hlgA, hlgB, hlgC, lukD, lukE, sec, seg, sei, sel, sem, sen, seo, seu | aur, splA, splB, splE | sak, scn | - |
| Richardson’s Beach Park (station #2) | Hilo | beach water | 11/14/2020 | 81 | MSSA | 72 i | 8 | blaZ, lnu(A) | hlgA, hlgB, hlgC, lukD, lukE, sec, seg, sei, sel, sem, sen, seo, seu | aur, splA, splB, splE | sak, scn | rep7c |
| Honoliʻi Beach Park (station #2) | Hilo | sand | 12/14/2020 | 106 | MSSA | 72 g | 8 | blaZ | hlgA, hlgB, hlgC, lukD, lukE, sec, seg, sei, sel, sem, sen, seo, seu | aur, splA, splB, splE | sak, scn | rep7c |
| Richardson’s Beach Park (station #2) | Hilo | beach water | 08/21/2020 | 79 | MSSA | 1181 j | 8 | aph(3’)-III, ant(6)-Ia, blaZ, tet(K) | hlgA, hlgB, hlgC, lukD, lukE, sep | aur, splA | sak, scn | rep7a |
| Lehia Beach Park (station #1) | Hilo | sand | 12/14/2020 | 111 | MSSA | 5 f | 5 | blaZ, erm(T), mph(C), tet(K) | hlgA, hlgB, hlgC, lukD, lukE, seg, sei, sem, sen, seo, seu | aur, splA, splB | sak, scn | rep7a, rep13 |
| Lalakea (station #6) | Hilo | beach water | 08/21/2020 | 38 | MSSA | 5 | 5 | - | hlgA, hlgB, hlgC, lukD, lukE, seg, sei, sem, sen, seo, seu | aur, splA | sak, scn | - |
| Kealoha Beach Park (station #8) | Hilo | beach water | 10/31/2020 | 35 | MSSA | 5 | 5 | blaZ | hlgA, hlgB, hlgC, lukD, lukE, sed, seg, sei, sej, sem, sen, seo, ser, seu | aur, splA, splB | sak, scn | rep20 |
| Kealoha Beach Park (station #8) | Hilo | beach water | 07/03/2020 | 70 | MSSA | 5 k | 5 | lnu(A) | hlgA, hlgB, hlgC, lukD, lukE, seg, sei, sem, sen, seo, seu | aur, splA, splB | sak, scn | - |
| Onekahakaha Beach Park (station #9) | Hilo | beach water | 08/21/2020 | 30 | MSSA | 5 l | 5 | blaZ | hlgA, hlgB, hlgC, lukD, lukE, seg, sei, sem, sen, seo, seu | aur, splA, splB | sak, scn | rep16 |
| Onekahakaha Beach Park (station #9) | Hilo | beach water | 08/21/2020 | 69 | MSSA | 5 l | 5 | - | hlgA, hlgB, hlgC, lukD, lukE, seg, sei, sem, sen, seo, seu | aur, splA, splB | sak, scn | - |
| Wailuku River–Upper (station #19) | Hilo | river water | 10/31/2020 | 14 | MSSA | 5 | 5 | blaZ | hlgA, hlgB, hlgC, lukD, lukE, sed, seg, sei, sej, sem, sen, seo, ser, seu | aur, splA, splB | sak, scn | rep20 |
| Richardson’s Beach Park (station #3) | Hilo | beach water | 10/31/2020 | 46 | MSSA | 5 | 5 | aph(3’)-III, ant(6)-Ia, blaZ, mph(C), msr(A) | hlgA, hlgB, hlgC, lukD, lukE, seg, sei, sem, sen, seo, seu | aur, splA, splB | sak, scn | rep19 |
| Pohoiki (station #34) | Puna | anchialine pool | 09/18/2020 | 54 | MSSA | 6 | 5 | blaZ | hlgA, hlgB, hlgC, lukD, lukE, sea, seb, sek, seq | aur, splA, splB, splE | sak, scn | rep16 |
| Kealoha Beach Park (station#8) | Hilo | beach water | 07/03/2020 | 32 | MSSA | 518 k | 5 | - | hlgA, hlgB, hlgC, lukD, lukE, seg, sei, sem, sen, seo, seu | aur, splA, splB | sak, scn | - |
| Honoliʻi Beach Park (station #20) | Hilo | sand | 12/14/2020 | 107 | MSSA | 518 g | 5 | blaZ, erm(T), mph(C), tet(K) | hlgA, hlgB, hlgC, lukD, lukE, seg, sei, sem, sen, seo, seu | aur, splA, splB | sak, scn | - |
| Lehia Beach Park (station #1) | Hilo | beach water | 10/31/2020 | 50 | MSSA | 398 m | 398 | blaZ, erm(T) | hlgA, hlgB, hlgC | aur | scn | rep13 |
| Lehia Beach Park (station #1) | Hilo | beach water | 10/31/2020 | 84 | MSSA | 398 m | 398 | blaZ, erm(T) | hlgA, hlgB, hlgC | aur | scn | rep13 |
| Kailua Pier (station #28) | N. Kona | beach water | 10/17/2020 | 58 | MSSA | 398 | 398 | blaZ, erm(T) | hlgA, hlgB, hlgC | aur | scn | rep13 |
| PohoikiBeach (station #35) | Puna | anchialine pool | 12/16/2020 | 55 | MSSA | 398 | 398 | erm(T) | hlgA, hlgB, hlgC | aur | scn | rep13 |
| Richardson’s Beach Park (station #3) | Hilo | beach water | 08/21/2020 | 45 | MSSA | 15 j | 15 | blaZ | hlgA, hlgB, hlgC, lukD, lukE | aur, splA, splB, splE | scn | rep16 |
| PohoikiBeach (station #34) | Puna | anchialine pool | 12/16/2020 | 56 | MSSA | 15 | 15 | blaZ | hlgA, hlgB, hlgC, lukD, lukE | aur, splA, splB, splE | scn | - |
| Waiʻōlena Beach Park (station #5) | Hilo | beach water | 07/31/2020 | 39 | MSSA | 508 | 45 | blaZ | hlgA, hlgB, hlgC, sec, seg, sei, sel, sem, sen, seo, seu, tst | aur | sak, scn | - |
| Wailuku River Estuary (station #17) | Hilo | river water | 12/14/2020 | 16 | MSSA | 508 | 45 | blaZ | hlgA, hlgB, hlgC, sec, seg, sei, sel, sem, sen, seo, seu, tst | aur | sak, scn | repUS5 |
| Honoliʻi Beach Park (station #20) | Hilo | beach water | 09/04/2020 | 13 | MSSA | 97 n | 97 | - | hlgA, hlgB, hlgC, lukD, lukE | aur, splA, splB | sak, scn | - |
| Honoliʻi Beach Park (station #20) | Hilo | beach water | 09/04/2020 | 66 | MSSA | 97 n | 97 | - | hlgA, hlgB, hlgC, lukD, lukE | aur, splA, splB | sak, scn | - |
| Richardson’s Beach Park (station #2) | Hilo | beach water | 10/31/2020 | 43 | MSSA | 1155 | 101 | blaZ | hlgA, hlgB, lukD, lukE | aur, splA, splB, splE | sak, scn | - |
| Wailuku River Estuary (station #17) | Hilo | river water | 08/15/2020 | 15 | MSSA | 3269 | 133 | - | hlgA, hlgB, hlgC, lukD, lukE | aur, splA, splB | - | - |
| Station | District | Source | Date | Isolate # | MRSA/MSSA | ST a | CC b | Antibiotic Resistance Genes (rep Family) | Virulence Genes (rep Family) | Plasmid | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Name | Accession # | ||||||||||
| Lehia Beach Park (station #1) | Hilo | sand | 12/12/2020 | 108 | MRSA | 8 | 8 | blaZ (rep19) erm(C) (rep10) | - | pWBG759 pDLK1 | GQ900401.1 GU562624.1 |
| Richardson’s Beach Park (station #3) | Hilo | beach water | 09/04/2020 | 60 | MRSA | 8 | 8 | blaZ (rep19) erm(C) (rep10) | ACME (rep7c) | TCH60 pDLK1 - | CP002111.1 GU562624.1 KF175393.1 |
| Reed’s Bay (station #11) | Hilo | sand | 12/14/2020 | 105 | MRSA | 8 | 8 | blaZ (rep19) erm(C) (rep10) mecA (rep7c) tet(K) (rep7a) | aur (rep7c) | pWBG759 pDLK1 - - | GQ900401.1 GU562624.1 BX571857.1 AB037671.1 |
| Honoliʻi Beach Park (station #20) | Hilo | beach water | 08/15/2020 | 59 | MRSA | 8 | 8 | blaZ (rep19) erm(C) (rep10) mecA (rep7c) | aur (rep7c) | TCH60 pDLK1 - | CP002111.1 GU562624.1 BX571857.1 |
| Honoliʻi Beach Park (station #20) | Hilo | sand | 12/14/2020 | 109 | MRSA | 8 | 8 | blaZ (rep19) erm(C) (rep10) tet(K) (rep7a) | - | TCH60 pDLK1 - | CP002111.1 GU562624.1 AB037671.1 |
| Kehena (station #31) | Hilo | beach water | 08/20/2020 | 61 | MRSA | 8 | 8 | erm(C) (rep10) mecA (rep7c) mph(C) (rep19) | aur (rep7c) | pDLK1 - pWBG759 | GU562624.1 BX571857.1 GQ900401.1 |
| Pohoiki Beach (station #32) | Puna | beach water | 08/20/2020 | 51 | MRSA | 8 | 8 | msr(A) (rep19) erm(C) (rep10) | - | TCH60 pDLK1 | CP002111.1 GU562624.1 |
| Pohoiki Beach (station #32) | Puna | beach water | 08/20/2020 | 88 | MRSA | 8 | 8 | blaZ (rep19) erm(C) (rep10) | - | pWBG759 pDLK1 | GQ900401.1 GU562624.1 |
| Pohoiki Harbor (station #33) | Puna | anchialine pool | 12/16/2020 | 89 | MRSA | 8 | 8 | blaZ (rep19) erm(C) (rep10) lnu(A) (rep21) | - | pWBG759 pKLD1 pKH21 | GQ900401.1 GU562624.1 EU350088.1 |
| Richardson’s Beach Park (station #2) | Hilo | beach water | 11/14/2020 | 81 | MSSA | 72 | 8 | - | aur (rep7c) | - | BX571857.1 |
| Honoliʻi Beach Park (station #2) | Hilo | sand | 12/14/2020 | 106 | MSSA | 72 | 8 | - | aur (rep7c) | - | BX571857.1 |
| Richardson’s Beach Park (station #2) | Hilo | beach water | 08/21/2020 | 79 | MSSA | 1181 | 8 | tet(K) (rep7a) | - | pS035-1 | AM990993.1 |
| Lehia Beach Park (station #1) | Hilo | sand | 12/14/2020 | 111 | MSSA | 5 | 5 | erm(T) (rep13) tet(K) (rep7a) | - | pLNU9 - | AM399082.1 AB037671.1 |
| Kealoha Beach Park (station #8) | Hilo | beach water | 10/31/2020 | 35 | MSSA | 5 | 5 | blaZ (rep20) | sed (rep20) | SAP074A | GQ900426.1 |
| Onekahakaha Beach Park (station #9) | Hilo | beach water | 08/21/2020 | 30 | MSSA | 5 | 5 | blaZ (rep16) | - | pSAS | BX571858.1 |
| Wailuku River–Upper (station #19) | Hilo | river water | 10/31/2020 | 14 | MSSA | 5 | 5 | blaZ (rep20) | sej (rep20) | SAP074A | GQ900426.1 |
| Richardson’s Beach Park (station #3) | Hilo | beach water | 10/31/2020 | 46 | MSSA | 5 | 5 | mph(C) (rep19) | - | pSJH901 | CP000704.1 |
| Pohoiki (station #34) | Puna | anchialine pool | 09/18/2020 | 54 | MSSA | 6 | 5 | blaZ (rep16) | - | pSaa6159 | CP002115.1 |
| Lehia Beach Park (station #1) | Hilo | beach water | 10/31/2020 | 50 | MSSA | 398 | 398 | erm(T) (rep13) | - | pC194 | V01277.1 |
| Lehia Beach Park (station #1) | Hilo | beach water | 10/31/2020 | 84 | MSSA | 398 | 398 | erm(T) (rep13) | - | pC194 | V01277.1 |
| Kailua Pier (station #28) | N. Kona | beach water | 10/17/2020 | 58 | MSSA | 398 | 398 | erm(T) (rep13) | - | pC194 | V01277.1 |
| Pohoiki Beach (station #35) | Puna | anchialine pool | 12/16/2020 | 55 | MSSA | 398 | 398 | erm(T) (rep13) | - | pC194 | V01277.1 |
| Richardson’s Beach Park (station #3) | Hilo | beach water | 08/21/2020 | 45 | MSSA | 15 | 15 | blaZ (rep16) | - | pSaa6159 | CP002115.1 |
| WailukuRiver Estuary (station #17) | Hilo | river water | 12/14/2020 | 16 | MSSA | 508 | 45 | blaZ (repUS5) | - | unnamed | NC_003265.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerken, T.J.; Roberts, M.C.; Dykema, P.; Melly, G.; Lucas, D.; De Los Santos, V.; Gonzalez, J.; Butaye, P.; Wiegner, T.N. Environmental Surveillance and Characterization of Antibiotic Resistant Staphylococcus aureus at Coastal Beaches and Rivers on the Island of Hawaiʻi. Antibiotics 2021, 10, 980. https://doi.org/10.3390/antibiotics10080980
Gerken TJ, Roberts MC, Dykema P, Melly G, Lucas D, De Los Santos V, Gonzalez J, Butaye P, Wiegner TN. Environmental Surveillance and Characterization of Antibiotic Resistant Staphylococcus aureus at Coastal Beaches and Rivers on the Island of Hawaiʻi. Antibiotics. 2021; 10(8):980. https://doi.org/10.3390/antibiotics10080980
Chicago/Turabian StyleGerken, Tyler J., Marilyn C. Roberts, Philip Dykema, Geoff Melly, Darren Lucas, Vanessa De Los Santos, Joenice Gonzalez, Patrick Butaye, and Tracy N. Wiegner. 2021. "Environmental Surveillance and Characterization of Antibiotic Resistant Staphylococcus aureus at Coastal Beaches and Rivers on the Island of Hawaiʻi" Antibiotics 10, no. 8: 980. https://doi.org/10.3390/antibiotics10080980
APA StyleGerken, T. J., Roberts, M. C., Dykema, P., Melly, G., Lucas, D., De Los Santos, V., Gonzalez, J., Butaye, P., & Wiegner, T. N. (2021). Environmental Surveillance and Characterization of Antibiotic Resistant Staphylococcus aureus at Coastal Beaches and Rivers on the Island of Hawaiʻi. Antibiotics, 10(8), 980. https://doi.org/10.3390/antibiotics10080980