
New Insight on Antibiotic Resistance and Virulence of Escherichia coli from Municipal and Animal Wastewater


Abstract
:1. Introduction
2. Results
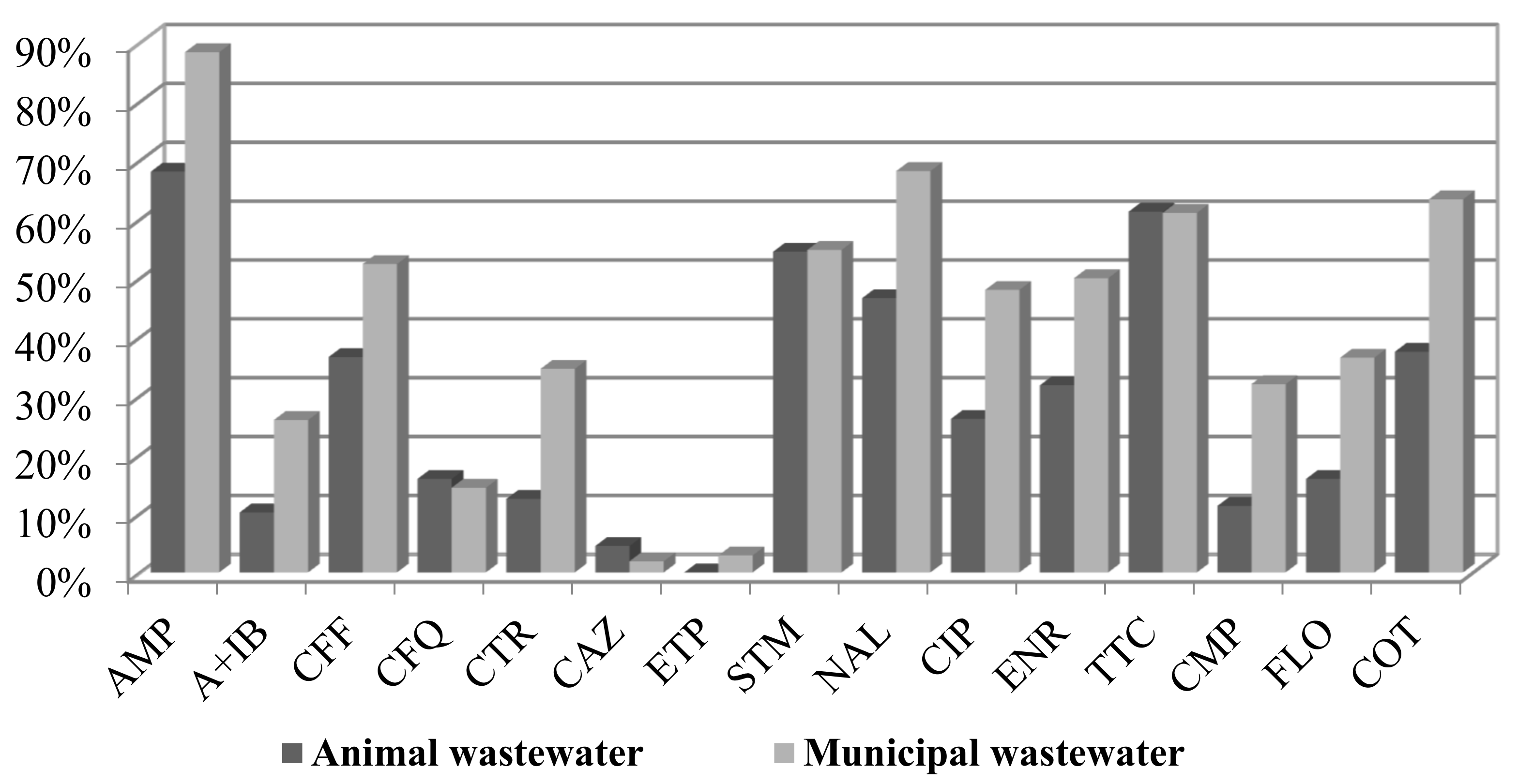
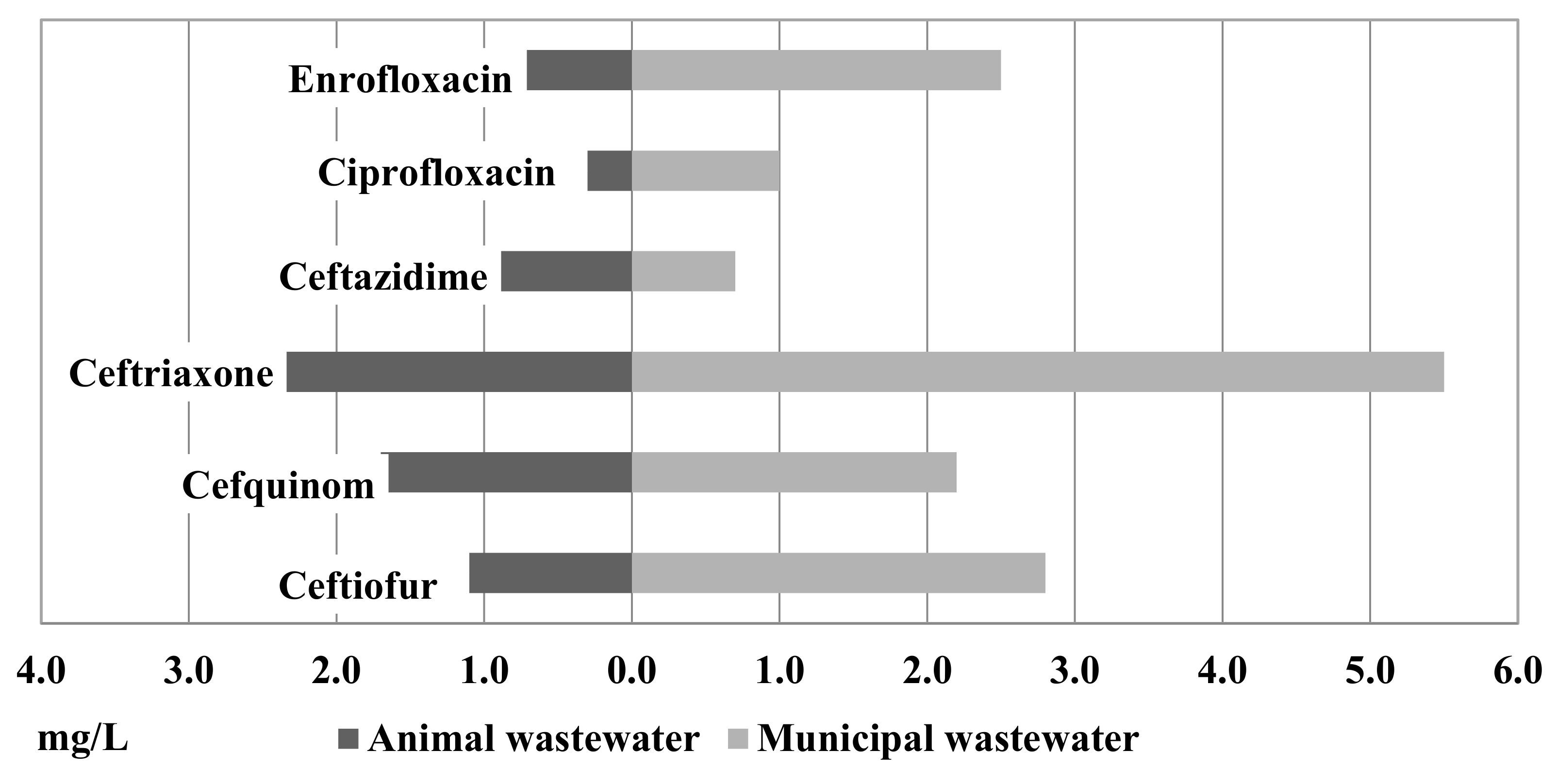
2.1. Antimicrobial Susceptibility Profiles
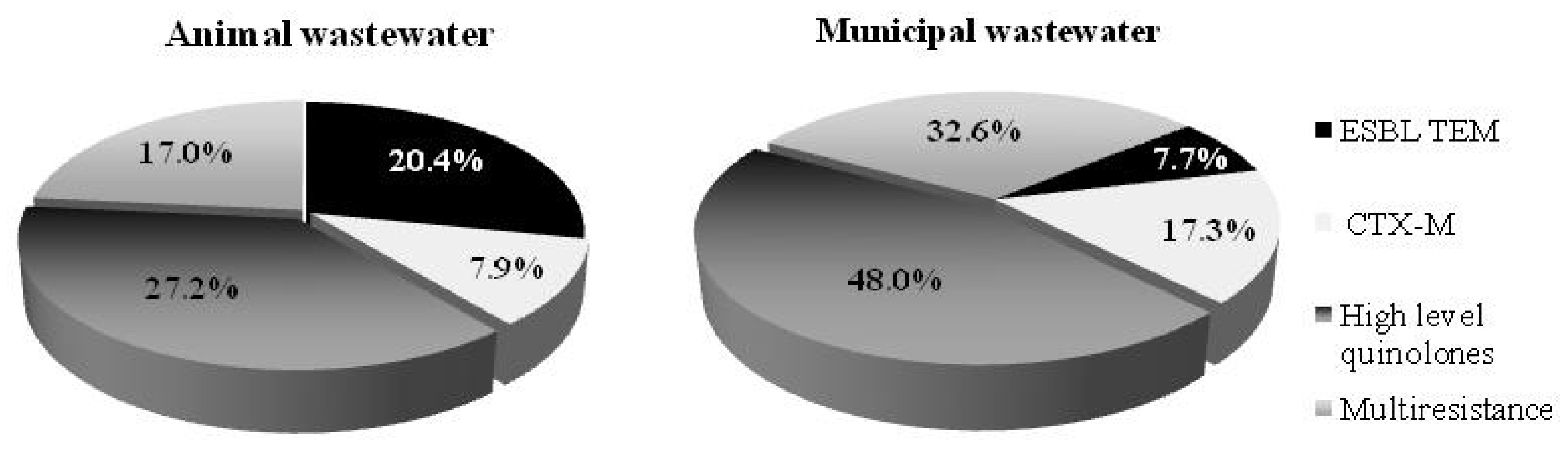
2.2. Genotyping Resistance, Mobilon, and Virulence Factors Detection
3. Discussion
4. Materials and Methods
4.1. Characteristics of the Places of Sampling
4.2. Identification of E. coli Strains and Antibiotic Susceptibility Detection
4.3. Detection of Antibiotic Resistance Genes in E. coli Isolates

{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence from 5′ to 3′ | Product bp | Anealing Temperature | References |
|---|---|---|---|---|
| Mobile Elements | ||||
| Int1 | GGGTCAAGGATCTGGATTTCG ACATGCGTGTAAATCATCGTCG | 483 | 62 °C | [39] |
| Tn3 | CACGAATGAGGGCCGACAGGA ACCCACTCGTGCACCCAACTG | 4000 | 58 °C | [40] |
| Genes of Resistance | ||||
| CIT | TGGCCAGAACTGACAGGCAAA TTTCTCCTGAACGTGGCTGGC | 462 | 61 °C | [37] |
| IMP | GAAGGCGTTTATGTTCATAC GTATGTTTCAAGAGTGATGC | 587 | 51 °C | [38] |
| qnrA | ATTTCTCACGCCAGGATTTG GATCGGCAAAGGTTAGGTCA | 516 | 53 °C | [42] |
| qnrB | GATCGTGAAAGCCAGAAAGG ACGATGCCTGGTAGTTGTCC | 469 | 53 °C | [42] |
| qnrS | ACGACATTCGTCAACTGCAA TAAATTGGCACCCTGTAGGC | 417 | 53 °C | [42] |
| aac(6’)IbCr | GATCTCATATCGTCGAGTGGTGG GAACCATGTACACGGCTGGAC | 435 | 58 °C | [42] |
| CTX-M1 | AAAAATCACTGCGCCAGTTC AGCTTATTCATCGCCACGTT | 415 | 52 °C | [36] |
| CTX-M 2 | CGACGCTACCCCTGCTATT CCAGCGTCAGATTTTTCAGG | 552 | 52 °C | [36] |
| CTX-M9 | CAAAGAGAGTGCAACGGATG ATTGGAAAGCGTTCATCACC | 205 | 52 °C | [36] |
4.4. Detection of Virulence Factors and Phylogenetic Group Assignment
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mackulak, T.; Nagyova, K.; Faberova, M.; Grabic, R.; Koba, O.; Gal, M.; Birosova, L. Utilization of Fenton-like reaction for antibiotics and resistantbacteria elimination in different parts of WWTP. Environ. Toxicol. Pharmacol. 2015, 40, 492–497. [Google Scholar] [CrossRef]
- Mao, D.; Yu, S.; Rysz, M.; Luo, Y.; Yang, F.; Li, F.; Hou, J.; Mu, Q.; Alvarez, P.J.J. Prevalence and proliferation of antibiotic resistance genes in two municipal wastewater treatment plants. Water Res. 2015, 85, 458–466. [Google Scholar] [CrossRef]
- Numberger, D.; Ganzert, L.; Zoccarato, L.; Muhldorfer, K.; Sauer, S.; Grossart, H.P.; Greenwood, A.D. Characterization of bacterial communities in wastewater with enhanced taxonomic resolution by full-length 16S rRNA sequencing. Sci. Rep. 2019, 9, 9673. [Google Scholar] [CrossRef] [Green Version]
- Baquero, F.; Lanza, V.F.; Cantón, R.; Coque, T.M. Public health evolutionary biology of antimicrobial resistance: Priorities for intervention. Evol. Appl. 2014, 8, 223–239. [Google Scholar] [CrossRef]
- De Kraker, M.E.A.; Stewardson, A.J.; Harbarth, S. Will 10 Million People Die a Year due to Antimicrobial Resistance by 2050? PLoS Med. 2016, 13, e1002184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabate, J.M.; Jouet, P.; Harnois, F.; Mechler, C.; Msika, S.; Grossin, M.; Coffin, B. High prevalence of small intestinal bacterial overgrowth in patients with morbid obesity: A contributor to severe hepatic steatosis. Obes. Surg. 2008, 18, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drugdova, Z.; Kmet, V. Prevalence of β-lactam and fluoroquinolone resistance, and virulence factors in Escherichia coli isolated from chickens in Slovakia. Biologia 2013, 68, 11–17. [Google Scholar] [CrossRef]
- Kmet, V.; Kmetova, M. High level of quinolone resistance in Escherichia coli from healthy chicken broilers. Folia Microbiol. 2010, 55, 79–82. [Google Scholar] [CrossRef]
- Gregova, G.; Kmetova, M.; Kmet, V.; Venglovsky, J.; Feher, A. Antibiotic resistance of Escherichia coli isolated from a poultry slaughterhouse. Ann. Agric. Environ. Med. 2012, 19, 75–77. [Google Scholar] [PubMed]
- Dodd, M.C. Potential impacts of disinfection processes on elimination and deactivation of antibiotic resistance genes during water and wastewater treatment. J. Environ. Monit. 2012, 12, 3047–3310. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Johnson, N.; Cizmas, L.; Mcdonald, T.J.; Kim, H. A review of the influence of treatment strategies on antibiotic resistant bacteria and antibiotic resistance genes. Chemosphere 2016, 150, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.B.; Guo, M.T.; Yang, J. Fate of Antibiotic Resistant Bacteria and Genes during Wastewater Chlorination: Implication for Antibiotic Resistance Control. PLoS ONE 2015, 10, e0119403. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhuang, Y.; Geng, J.; Ren, H.; Zhang, Y.; Ding, L.; Xu, K. Inactivation of antibiotic resistance genes in municipal wastewater effluent by chlorination and sequential UV/chlorination disinfection. Sci. Total Environ. 2015, 512–513, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xu, Y.; Dong, X.; Shen, C. Removal of Intracellular and Extracellular Antibiotic Resistance Genes in Municipal Wastewater Effluent by Electrocoagulation. Environ. Eng. Sci. 2020, 37, 783–789. [Google Scholar] [CrossRef]
- Korzeniewska, E.; Korzeniewska, A.; Harnisz, M. Antibiotic resistant Escherichia coli in hospital and municipal sewage and their emission to the environment. Ecotoxicol. Environ. Saf. 2013, 91, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Czekalski, N.; Berthold, T.; Caucci, S.; Egli, A.; Buergmann, H. Increased levels of multiresistant bacteria and resistance genes after wastewater treatment and their dissemination into Lake Geneva, Switzerland. Front. Microbiol. 2012, 3, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, C.M.; Zhao, C.W.; Debroy, C. Antimicrobial resistance of Escherichia coli O157 isolated from humans, cattle, swine, and food. Appl. Environ. Microbiol. 2002, 68, 576–581. [Google Scholar] [CrossRef] [Green Version]
- Guardabassi, L.; Kruse, H. Guide to Antimicrobial Use in Animals. In Principles of Prudent and Trational Use of Antimicrobials in Animals; Guardabassi, L., Jensen, L.B., Kruse, H., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2008; pp. 1–13. ISBN 978-1-4051-5079-8. [Google Scholar]
- European Medicines Agency; European Surveillance of Veterinary Antimicrobial Consumption (ESVAC). Sales of Veterinary Antimicrobial Agents in 31 European Union Countries in 2018; Tenth ESVAC Report; EMA: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Van Doorslaer, X.; Dewulf, J.; Van Langenhove, H.; Demeestere, K. Fluoroquinolone antibiotics: An emerging class of environmental micropollutants. Sci. Total Environ. 2014, 500–501, 250–269. [Google Scholar] [CrossRef]
- Kaplan, E.; Ofek, M.; Jurkevitch, E.; Cytryn, E. Characterization of fluoroquinolone resistance and qnr diversity in Enterobacteriaceae from municipal biosolids. Front. Microbiol. 2013, 4, 144. [Google Scholar] [CrossRef] [Green Version]
- Kümmerer, K. The presence of pharmaceuticals in the environment due to human use—Present knowledge and future challenges. J. Environ. Manag. 2009, 90, 2354–2366. [Google Scholar] [CrossRef]
- Oslinska, A.; Harnisz, M.; Korzeniewska, E. Prevalence of plasmid-mediated multidrug resistance determinants in fluoroquinolone-resistant bacteria isolated from sewage and surface water. Environ. Sci. Pollut. Res. 2016, 23, 10818–10831. [Google Scholar] [CrossRef] [Green Version]
- Su, H.C.; Hu, X.J.; Xu, Y.; Xu, W.J.; Huang, X.S.; Wen, G.L.; Yang, K.; Li, Z.; Cao, Y. Persistence and spatial variation of antibiotic resistance genes and bacterial populations change in reared shrimp in south china. Environ. Int. 2018, 119, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Savin, M.; Bierbaum, G.; Kreyenschmidt, J.; Schmithausen, R.M.; Sib, E.; Schmoger, S.; Käsbohrer, A.; Hammerl, J.A. Clinically Relevant Escherichia coli Isolates from Process Waters and Wastewater of Poultry and Pig Slaughterhouses in Germany. Microorganisms 2021, 9, 698. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Fakhr, M.K.; Nolan, L.K. Comparison of Escherichia coli isolates implicated in human urinary tract infection and avian colibacillosis. Microbiology 2005, 151, 2097–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewers, R.M.; Scharlemann, J.P.W.; Balmford, A.; Green, R.E. Do increases in agricultural yield spare land for nature? Glob. Chang. Biol. 2009, 15, 1716–1726. [Google Scholar] [CrossRef]
- Adegoke, A.A.; Madu, C.E.; Aiyegoro, O.A.; Stenström, T.A.; Okoh, A.I. Antibiogram and beta-lactamase genes among cefotaxime resistant E. coli from wastewater treatment plant. Antimicrob. Resist. Infect. Cont. 2020, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Cornejova, T.; Venglovsky, J.; Gregova, G.; Kmetova, M.; Kmet, V. Extended spectrum beta-lactamases in Escherichia coli from municipal wastewater. Ann. Agric. Environ. Med. 2015, 22, 447–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregova, G.; Kmet, V. Antibiotic resistance and virulence of Escherichia coli strains isolated from animal rendering plant. Sci. Rep. 2020, 10, 17108. [Google Scholar] [CrossRef] [PubMed]
- CLSI Document VET01-S2 Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2013; pp. 1–168.
- EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance, Version 2.0; EUCAST: Växjö, Sweden, 2017; 1–43.
- Gattringer, R.; Niks, M.; Ostertag, R.; Schwarz, K.; Medvedovic, H.; Graninger, W.; Georgopoulos, A. Evaluation of MIDITECH automated colorimetric MIC reading for antimicrobial susceptibility testing. J. Antimicrob. Chemother. 2002, 49, 651–659. [Google Scholar] [CrossRef]
- Livermore, D.M.; Winstanley, T.G.; Shannon, K.P. Interpretative reading: Recognizing the unusual and inferring resistance mechanisms from resistance phenotypes. J. Antimicrob. Chemother. 2001, 48, 87–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum (β)-lactamases. J. Antimicrob. Chemother. 2006, 57, 154–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Perez, F.J.; Hanson, N.D. Detection of Plasmid-Mediated AmpC β-Lactamase Genes in Clinical Isolates by Using Multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fallah, F.; Borhan, R.S.; Hashemi, A. Detection of bla(IMP) and bla(VIM) metallo-β-lactamases genes among Pseudomonas aeruginosa strains. Int. J. Burn. Trauma. 2013, 3, 122–124. [Google Scholar]
- Mazel, D.; Dychingo, B.; Webb, V.A.; Davies, J. Antibiotic resistance in the ECOR collection: Integrons and identification of a novel aad gene. Antimicrob. Agents Chemother. 2000, 44, 1568–1574. [Google Scholar] [CrossRef] [Green Version]
- Weill, F.W.; Demartin, M.; Fabre, L.; Grimot, P.A.D. Extended-spectrum-β-lactamase (TEM-52)-producing strains of Salmonella enterica of various serotypes isolated in France. J. Clin. Microbiol. 2004, 42, 3359–3362. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, W.; Pan, W.; Yin, J.; Pan, Z.; Gao, S.; Jiao, X. Prevalence of qnr, aac (6’)-Ib-cr, qepA, and oqxAB in Escherichia coli isolates from humans, animals, and the environment. Antimicrob. Agents Chemother. 2012, 56, 3423–3427. [Google Scholar] [CrossRef] [Green Version]
- Robicsek, A.; Strahilevitz, J.; Sahm, D.F.; Jacoby, G.A.; Hooper, D.C. Qnr Prevalence in Ceftazidime-Resistant Enterobacteriaceae Isolates from the United States. Antimicrob. Agents Chemother. 2006, 50, 2872–2874. [Google Scholar] [CrossRef] [Green Version]
- Iranpour, D.; Hassanpour, M.; Ansari, H.; Tajbakhsh, S.; Khamisipour, G.; Najafi, A. Phylogenetic Groups of Escherichia coli Strains from Patients with Urinary Tract Infection in Iran Based on the New Clermont Phylotyping Method. Biomed Res. Int. 2015, 2015, 846219. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Foley, S.L.; Home, S.M.; Giddings, C.W.; Robinson, M.; Nolan, L.K. Iss from a virulent avian Escherichia coli. Avian Dis. 2000, 44, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Dozois, C.M.; Dho-Moulin, M.; Brée, A.; Fairbrother, J.M.; Desautels, C.; Curtiss, R. Relationship between the Tsh Autotransporter and Pathogenicity of Avian Escherichia coli and Localization and Analysis of the tsh Genetic Region. ASM J. Infect. Immun. 2000, 68, 4145–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Bouguenec, C.; Archambaud, M.; Labigne, A. Rapid and specifc detection of the pap, afa, and sfa adhesin-encoding operons in uropathogenic Escherichia coli strains by polymerase chain reaction. J. Clin. Microb. 1992, 30, 1189–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattoir, V.; Poirel, L.; Nordmann, P. Plasmid-mediated quinolone resistance pump QepA2 in an Escherichia coli isolate from France. Antimicrob. Agents Chemother. 2008, 52, 3801–3804. [Google Scholar] [CrossRef] [PubMed] [Green Version]



| Animal Wastewater (n = 14) | |||
|---|---|---|---|
| Commensals (number strains) | PG * | Pathogens (number strains) | PG * |
| Cit, Int1, Tn3 (n = 1) | A | Cit, Int1, Tn3 (n = 1) | B2 |
| Cit, Int1 (n = 3) | A | Cit, Int1 (n = 1) | B2 |
| Int1 (n = 4) | A | Cit, Tn3 (n = 1) | D |
| Tn3 (n = 2) | A | Tn3 (n = 1) | B2 |
| Municipal Wastewater (n = 13) | |||
| Commensals (number strains) | PG * | Pathogens (number strains) | PG * |
| CTX-M1, Int1, Tn3 (n = 1) | A | Int1, Tn3 (n = 1) | A |
| CTX-M1, Int1, (n = 2) | A | CTX-M1, Cit, Int1 (n = 1) | B2 |
| Cit, Int1 (n = 2) | A | CTX-M1, Int1, Tn3 (n = 1) | B2 |
| IMP, Int1 (n = 1) | A | CTX-M1, Tn3 (n = 1) | B2 |
| qnrS (n = 1) | A | CTX-M1 (n = 1) | D |
| CTX-M1, Cit, qnrS, Int1, Tn3 (n = 1) | nd | ||
| Primer | Sequence from 5′ to 3′ | Product bp | Anealing Temperature | References |
|---|---|---|---|---|
| Phylogenetic Group | ||||
| ChuA | GACGAACCA ACGGTCAGGAT TGCCGCCAGTACC AAAGACA | 279 | 55 °C | [7] |
| YjaA | TGAAGTGTCAGGAGACGCTG ATGGAGAATGCGTTCCTCAAC | 211 | 55 °C | [7] |
| TspE4C2 | GAGTAATGTCGGGGCATTCA CGCGCCAACAAAGTATTACG | 152 | 55 °C | [7] |
| arpA | AACGCTATTCGCCAGCTTGC TCTCCCCATACCGTACGCTA | 400 | 59 °C | [43] |
| trpA | AGTTTTATGCCCAGTGCGAG TCTGCGCCGGTCACGCCC | 219 | 59 °C | [43] |
| Virulence Factors | ||||
| iutA | GGCTGGACATGGGAACTGG CGTCGGGAACGGGTAGAATCG | 300 | 63 °C | [44] |
| iss | GTGGCGAAAACTAGTAAAACAGC CGCCTCGGGGTGGATAA | 760 | 61 °C | [45] |
| cvaC | CACACACAAACGGGAGCTGTT CTTCCCGCAGCATAGTTCCAT | 680 | 63 °C | [44] |
| kpsII | GCGCATTTGCTGATACTGTTG CATCCAGACGATAAGCATGAGCA | 272 | 63 °C | [44] |
| tsh | GGTGGTGCACTGGAGTGG AGTCCAGCGTGATAGTGG | 640 | 55 °C | [46] |
| papC | GACGGCTGTACTGCAGGGTGTGGCG ATATCCTTTCTGCAGGGATGCAATA | 328 | 61 °C | [47] |
| oqxA | GACAGCGTCGCACAGAATG GGAGACGAGGTTGGTATGGA | 339 | 62 °C | [41] |
| oqx B | CGAAGAAAGACCTCCCTACCC CGCCGCCAATGAGATACA | 240 | 62 °C | [41] |
| qepA | CTGCAGGTACTGCGTCATG CGTGTTGCTGGAGTTCTTC | 403 | 60 °C | [48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gregova, G.; Kmet, V.; Szaboova, T. New Insight on Antibiotic Resistance and Virulence of Escherichia coli from Municipal and Animal Wastewater. Antibiotics 2021, 10, 1111. https://doi.org/10.3390/antibiotics10091111
Gregova G, Kmet V, Szaboova T. New Insight on Antibiotic Resistance and Virulence of Escherichia coli from Municipal and Animal Wastewater. Antibiotics. 2021; 10(9):1111. https://doi.org/10.3390/antibiotics10091111
Chicago/Turabian StyleGregova, Gabriela, Vladimir Kmet, and Tatiana Szaboova. 2021. "New Insight on Antibiotic Resistance and Virulence of Escherichia coli from Municipal and Animal Wastewater" Antibiotics 10, no. 9: 1111. https://doi.org/10.3390/antibiotics10091111
APA StyleGregova, G., Kmet, V., & Szaboova, T. (2021). New Insight on Antibiotic Resistance and Virulence of Escherichia coli from Municipal and Animal Wastewater. Antibiotics, 10(9), 1111. https://doi.org/10.3390/antibiotics10091111