

Genomic Diversity of NDM-Producing Klebsiella Species from Brazil, 2013–2022

 ,
,  and
and
Abstract
:1. Introduction
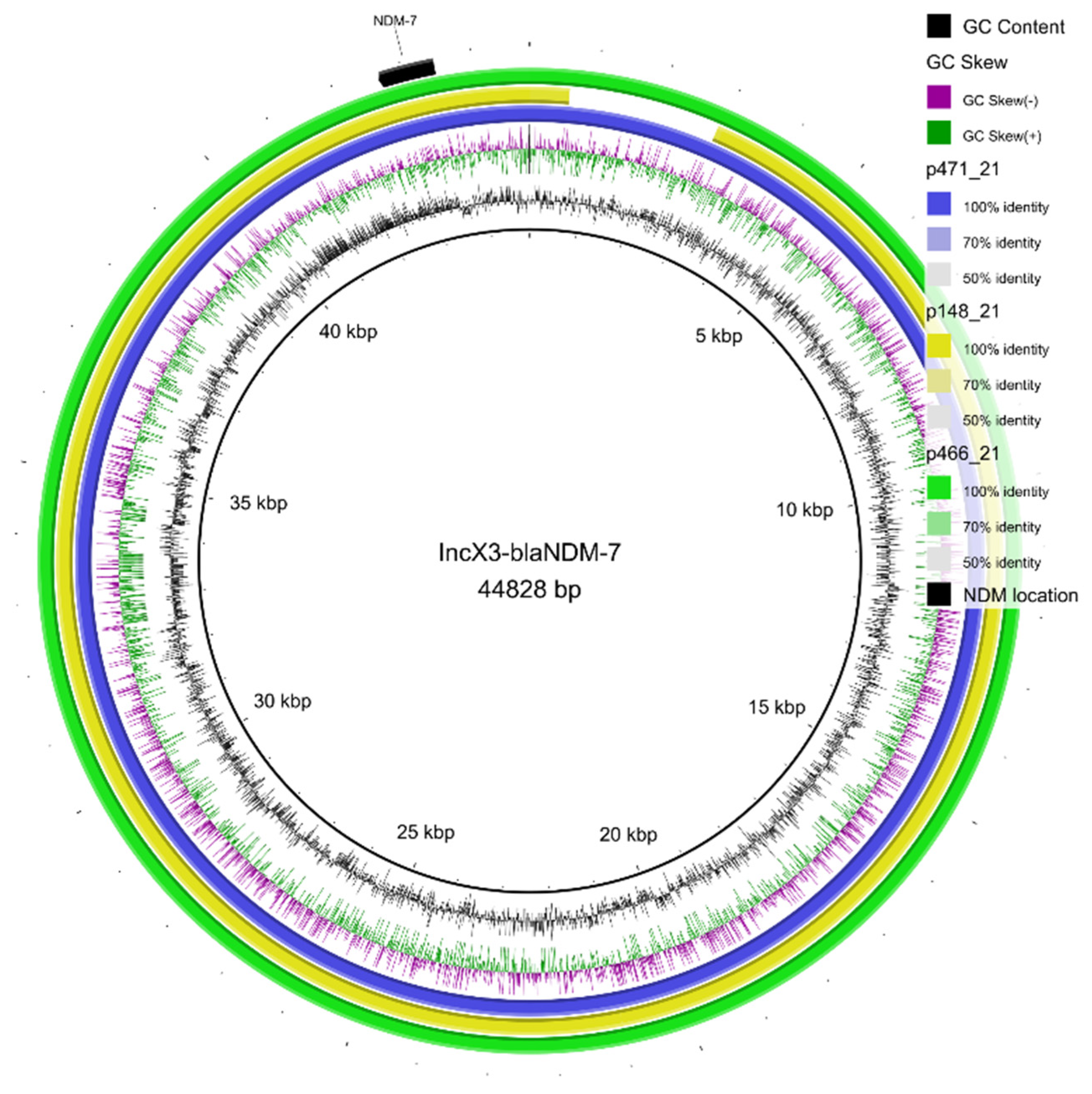
2. Results
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alsan, M.; Schoemaker, L.; Eggleston, K.; Kammili, N.; Kolli, P.; Bhattacharya, J. Out-of-Pocket Health Expenditures and Antimicrobial Resistance in Low-Income and Middle-Income Countries: An Economic Analysis. Lancet Infect. Dis. 2015, 15, 1203–1210. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F. The Challenges of Antimicrobial Resistance in Brazil. Clin. Infect. Dis. 2011, 52, 1138–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortaleza, C.M.C.B.; Padoveze, M.C.; Kiffer, C.R.V.; Barth, A.L.; Carneiro, I.C.D.R.S.; Giamberardino, H.I.G.; Rodrigues, J.L.N.; Santos Filho, L.; de Mello, M.J.G.; Pereira, M.S.; et al. Multi-State Survey of Healthcare-Associated Infections in Acute Care Hospitals in Brazil. J. Hosp. Infec.t 2017, 96, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, J.L.M.; Gales, A.C. Antimicrobial Resistance in Enterobacteriaceae in Brazil: Focus on β-Lactams and Polymyxins. Braz. J. Microbiol. 2016, 47, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a New Antibiotic Resistance Mechanism in India, Pakistan, and the UK: A Molecular, Biological, and Epidemiological Study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Hansen, G.T. Continuous Evolution: Perspective on the Epidemiology of Carbapenemase Resistance Among Enterobacterales and Other Gram-Negative Bacteria. Infect. Dis. Ther. 2021, 10, 75–92. [Google Scholar] [CrossRef]
- Carvalho-Assef, A.P.; Pereira, P.S.; Albano, R.M.; Berião, G.C.; Chagas, T.P.; Timm, L.N.; Da Silva, R.C.; Falci, D.R.; Asensi, M.D. Isolation of NDM-Producing Providencia Rettgeri in Brazil. J. Antimicrob. Chemother. 2013, 68, 2956–2957. [Google Scholar] [CrossRef] [Green Version]
- Camargo, C.H. Current Status of NDM-Producing Enterobacterales in Brazil: A Narrative Review. Braz. J. Microbiol. 2022, 53, 1339–1344. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, I.R.; Aires, C.A.M.; Conceição-Neto, O.C.; De Oliveira Santos, I.C.; Ferreira Pereira, N.; Moreno Senna, J.P.; Carvalho-Assef, A.P.D.A.; Asensi, M.D.; Rocha-De-Souza, C.M. Distribution of Clinical NDM-1-Producing Gram-Negative Bacteria in Brazil. Microb. Drug Resist. 2019, 25, 394–399. [Google Scholar] [CrossRef]
- Safavi, M.; Bostanshirin, N.; Hajikhani, B.; Yaslianifard, S.; van Belkum, A.; Goudarzi, M.; Hashemi, A.; Darban-Sarokhalil, D.; Dadashi, M. Global Genotype Distribution of Human Clinical Isolates of New Delhi Metallo-β-Lactamase-Producing Klebsiella Pneumoniae; A Systematic Review. J. Glob. Antimicrob. Resist. 2020, 23, 420–429. [Google Scholar] [CrossRef]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef] [Green Version]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, J.; Inoue, F.M.; Lobo, A.P.T.; Ibanes, A.S.; Tufik, S.; Kiffer, C.R.V. A Major Monoclonal Hospital Outbreak of NDM-1-Producing Klebsiella Pneumoniae ST340 and the First Report of ST2570 in Brazil. Infect. Control Hosp. Epidemiol. 2019, 40, 492–494. [Google Scholar] [CrossRef] [Green Version]
- Quezada-Aguiluz, M.; Opazo-Capurro, A.; Lincopan, N.; Esposito, F.; Fuga, B.; Mella-Montecino, S.; Riedel, G.; Lima, C.A.; Bello-Toledo, H.; Cifuentes, M.; et al. Novel Megaplasmid Driving NDM-1-Mediated Carbapenem Resistance in Klebsiella Pneumoniae ST1588 in South America. Antibiotics 2022, 11, 1207. [Google Scholar] [CrossRef]
- Poirel, L.; Dortet, L.; Bernabeu, S.; Nordmann, P. Genetic Features of BlaNDM-1-Positive Enterobacteriaceae. Antimicrob. Agents Chemother. 2011, 55, 5403–5407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozales, F.P.; Magagnin, C.M.; Campos, J.C.; Pagano, M.; Nunes, L.S.; Pancotto, L.R.; Sampaio, J.L.M.; Zavascki, A.P.; Barth, A.L. Characterization of Transformants Obtained from NDM-1-Producing Enterobacteriaceae in Brazil. Infect. Control Hosp. Epidemiol. 2017, 38, 634–636. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F.; Girardello, R.; Cury, A.P.; Di Gioia, T.S.R.; Almeida, J.N.D.; Duarte, A.J.D.S.; Rossi, F.; Girardello, R.; Cury, A.P.; Di Gioia, T.S.R.; et al. Emergence of Colistin Resistance in the Largest University Hospital Complex of São Paulo, Brazil, over Five Years. Braz. J. Infect. Dis. 2017, 21, 98–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhanel, G.G.; Lawrence, C.K.; Adam, H.; Schweizer, F.; Zelenitsky, S.; Zhanel, M.; Lagacé-Wiens, P.R.S.; Walkty, A.; Denisuik, A.; Golden, A.; et al. Imipenem–Relebactam and Meropenem–Vaborbactam: Two Novel Carbapenem-β-Lactamase Inhibitor Combinations. Drugs 2018, 78, 65–98. [Google Scholar] [CrossRef]
- Papadimitriou-Olivgeris, M.; Bartzavali, C.; Lambropoulou, A.; Solomou, A.; Tsiata, E.; Anastassiou, E.D.; Fligou, F.; Marangos, M.; Spiliopoulou, I.; Christofidou, M. Reversal of Carbapenemase-Producing Klebsiella Pneumoniae Epidemiology from BlaKPC-to BlaVIM-Harbouring Isolates in a Greek ICU after Introduction of Ceftazidime/Avibactam. J. Antimicrob. Chemother. 2019, 74, 2051–2054. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for Detection of Acquired Carbapenemase Genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barrett, T.J. Standardization of Pulsed-Field Gel Electrophoresis Protocols for the Subtyping of Escherichia Coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Nekrutenko, A.; Grüning, B.A.; Blankenberg, D.; Goecks, J.; Schatz, M.C.; Ostrovsky, A.E.; Mahmoud, A.; Lonie, A.J.; Syme, A.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2022 Update. Nucleic Acids Res. 2022, 50, W345–W351. [Google Scholar] [CrossRef]
- Argimón, S.; David, S.; Underwood, A.; Abrudan, M.; Wheeler, N.E.; Kekre, M.; Abudahab, K.; Yeats, C.A.; Goater, R.; Taylor, B.; et al. Rapid Genomic Characterization and Global Surveillance of Klebsiella Using Pathogenwatch. Clin. Infect. Dis. 2021, 73, S325–S335. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A Reference Database for Bacterial Virulence Factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef] [Green Version]
- Argimón, S.; Abudahab, K.; Goater, R.J.E.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.J.; Holden, M.T.G.; Yeats, C.A.; Grundmann, H.; et al. Microreact: Visualizing and Sharing Data for Genomic Epidemiology and Phylogeography. Microb. Genom. 2016, 2, e000093. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations Using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A Genome Comparison Visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Antimicrobial Category | Antimicrobial Agent | Susceptible | Susceptible, Increased Exposure | Resistant | |||
|---|---|---|---|---|---|---|---|
| n | % | n | % | n | % | ||
| Aminoglycosides | Amikacin | 76 | 56.3 | 0 | 0.0 | 59 | 43.7 |
| Gentamicin | 48 | 35.6 | 0 | 0.0 | 87 | 64.4 | |
| Tobramycin | 16 | 11.9 | 0 | 0.0 | 119 | 88.1 | |
| Quinolones | Ciprofloxacin | 9 | 6.7 | 6 | 4.4 | 120 | 88.9 |
| Levofloxacin | 14 | 10.4 | 11 | 8.1 | 110 | 81.5 | |
| Folate pathway inhibitors | Trimethoprim- sulfamethoxazole | 21 | 15.6 | 2 | 1.5 | 112 | 83.0 |
| Penicillin | Ampicillin | 0 | 0.0 | 0 | 0.0 | 135 | 100.0 |
| Cephalosporins | Cefepime | 4 | 3.0 | 0 | 0.0 | 131 | 97.0 |
| Cefotaxime | 3 | 2.2 | 0 | 0.0 | 132 | 97.8 | |
| Ceftazidime | 4 | 3.0 | 0 | 0.0 | 131 | 97.0 | |
| Carbapenems | Ertapenem | 4 | 3.0 | 0 | 0.0 | 131 | 97.0 |
| Imipenem | 5 | 3.7 | 23 | 17.0 | 107 | 79.3 | |
| Meropenem | 5 | 3.7 | 10 | 7.4 | 120 | 88.9 | |
| Cephamycins | Cefoxitin | 2 | 1.5 | 0 | 0.0 | 133 | 98.5 |
| β-lactams + β-lactamase inhibitors | Amoxicillin-clavulanic acid | 4 | 3.0 | 0 | 0.0 | 131 | 97.0 |
| Ampicillin-sulbactam | 3 | 2.2 | 0 | 0.0 | 132 | 97.8 | |
| Piperacillin-tazobactam | 3 | 2.2 | 1 | 0.7 | 131 | 97.0 | |
| Monobactam | Aztreonam | 11 | 8.1 | 2 | 1.5 | 122 | 90.4 |
| Phenicols | Chloramphenicol | 54 | 40.0 | 0 | 0.0 | 81 | 60.0 |
| Polymyxins | Polymyxin B 1 | 120 | 88.9 | 0 | 0.0 | 15 | 11.1 |
| Strain Identification | J53 | 146_19_trans | 148_21_trans | 229_19_trans | 466_21_trans | 471_21_trans | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Parental Species | E. coli | K. pneumoniae | K. pneumoniae | K. pneumoniae | K. pneumoniae | K. quasipneumoniae | |||||
| NDM-Type | Recipient Strain | NDM-1 | NDM-7 | NDM-1 | NDM-7 | NDM-7 | |||||
| Antimicrobial Agent | MIC | MIC | X Fold * | MIC | X Fold | MIC | X Fold | MIC | X Fold | MIC | X Fold |
| Imipenem | 0.125 | 32.0 | 256 | >32.0 | >256 | 12.0 | 96 | >32.0 | >256 | 4.0 | 32 |
| Meropenem | 0.016 | 4.0 | 250 | 6.0 | 375 | 3.0 | 188 | >32.0 | >2000 | 1.0 | 63 |
| Ampicillin | 2.0 | >256 | >128 | >256 | >128 | >256 | >128 | >256 | >128 | >256 | >128 |
| Cefotaxime | 0.015 | >256 | >17,067 | >256 | >17,067 | 256.0 | 17,067 | >256 | >17,067 | >256 | >17,067 |
| Cefepime | 0.32 | 192.0 | 600 | 32.0 | 100 | 8.0 | 25 | 24.0 | 75 | 16.0 | 50 |
| Ceftriaxone | 0.015 | >32.0 | >2133 | >32.0 | >2133 | >32.0 | >2133 | >32.0 | >2133 | >32 | >2133 |
| Ticarcillin-clavulanic acid | 1.5 | >256 | >171 | >256 | >171 | >256 | >171 | >256 | >171 | >256 | >171 |
| Cephalotin | 3.0 | >256 | >85 | >256 | >85 | >256 | >85 | >256 | >85 | >256 | >85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camargo, C.H.; Yamada, A.Y.; Souza, A.R.d.; Reis, A.D.; Santos, M.B.N.; Assis, D.B.d.; Carvalho, E.d.; Takagi, E.H.; Cunha, M.P.V.; Tiba-Casas, M.R. Genomic Diversity of NDM-Producing Klebsiella Species from Brazil, 2013–2022. Antibiotics 2022, 11, 1395. https://doi.org/10.3390/antibiotics11101395
Camargo CH, Yamada AY, Souza ARd, Reis AD, Santos MBN, Assis DBd, Carvalho Ed, Takagi EH, Cunha MPV, Tiba-Casas MR. Genomic Diversity of NDM-Producing Klebsiella Species from Brazil, 2013–2022. Antibiotics. 2022; 11(10):1395. https://doi.org/10.3390/antibiotics11101395
Chicago/Turabian StyleCamargo, Carlos Henrique, Amanda Yaeko Yamada, Andreia Rodrigues de Souza, Alex Domingos Reis, Marlon Benedito Nascimento Santos, Denise Brandão de Assis, Eneas de Carvalho, Elizabeth Harummyy Takagi, Marcos Paulo Vieira Cunha, and Monique Ribeiro Tiba-Casas. 2022. "Genomic Diversity of NDM-Producing Klebsiella Species from Brazil, 2013–2022" Antibiotics 11, no. 10: 1395. https://doi.org/10.3390/antibiotics11101395
APA StyleCamargo, C. H., Yamada, A. Y., Souza, A. R. d., Reis, A. D., Santos, M. B. N., Assis, D. B. d., Carvalho, E. d., Takagi, E. H., Cunha, M. P. V., & Tiba-Casas, M. R. (2022). Genomic Diversity of NDM-Producing Klebsiella Species from Brazil, 2013–2022. Antibiotics, 11(10), 1395. https://doi.org/10.3390/antibiotics11101395