

Phytogenic Blend Improves Intestinal Health and Reduces Obesity, Diabetes, Cholesterol and Cancers: A Path toward Customised Supplementation

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Trial and Bird Management
2.2. RNA Sequencing and Analysis
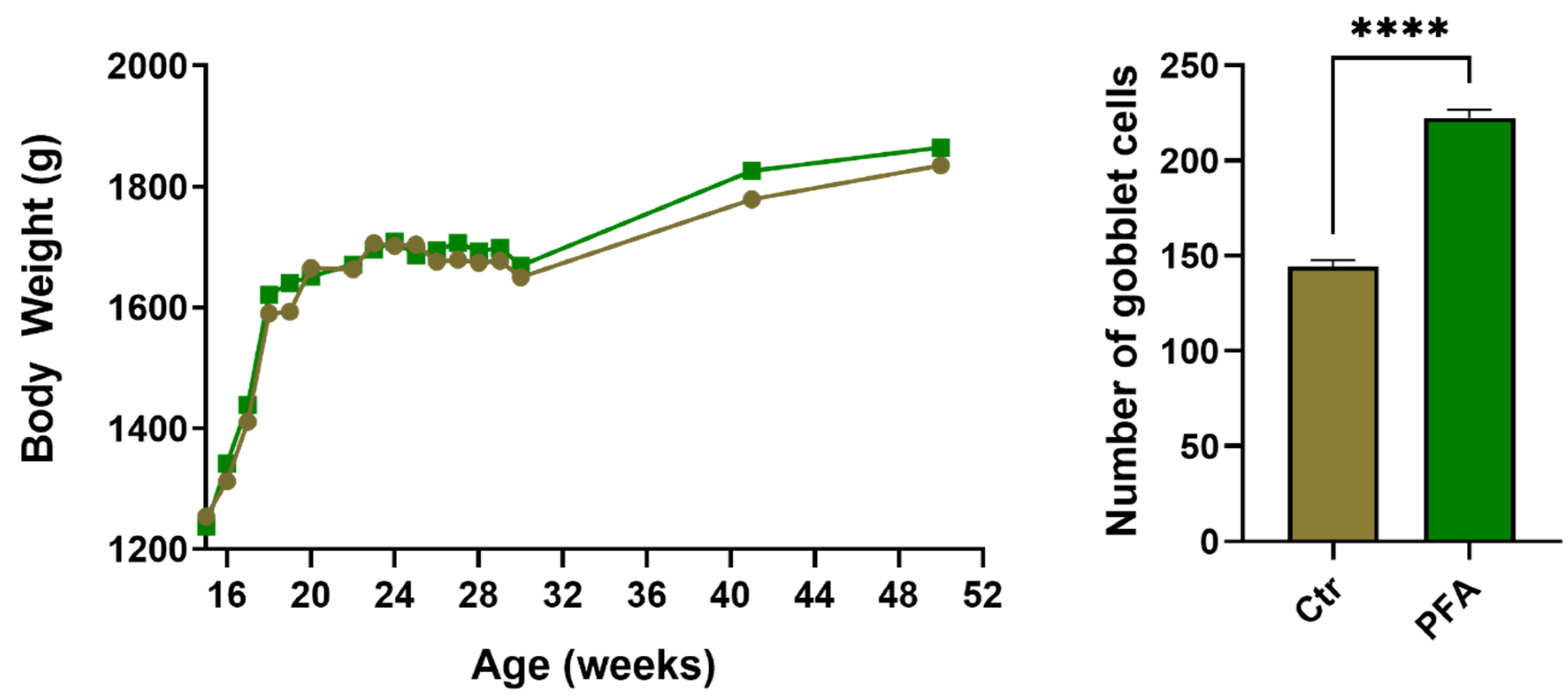
2.3. Histology
3. Results
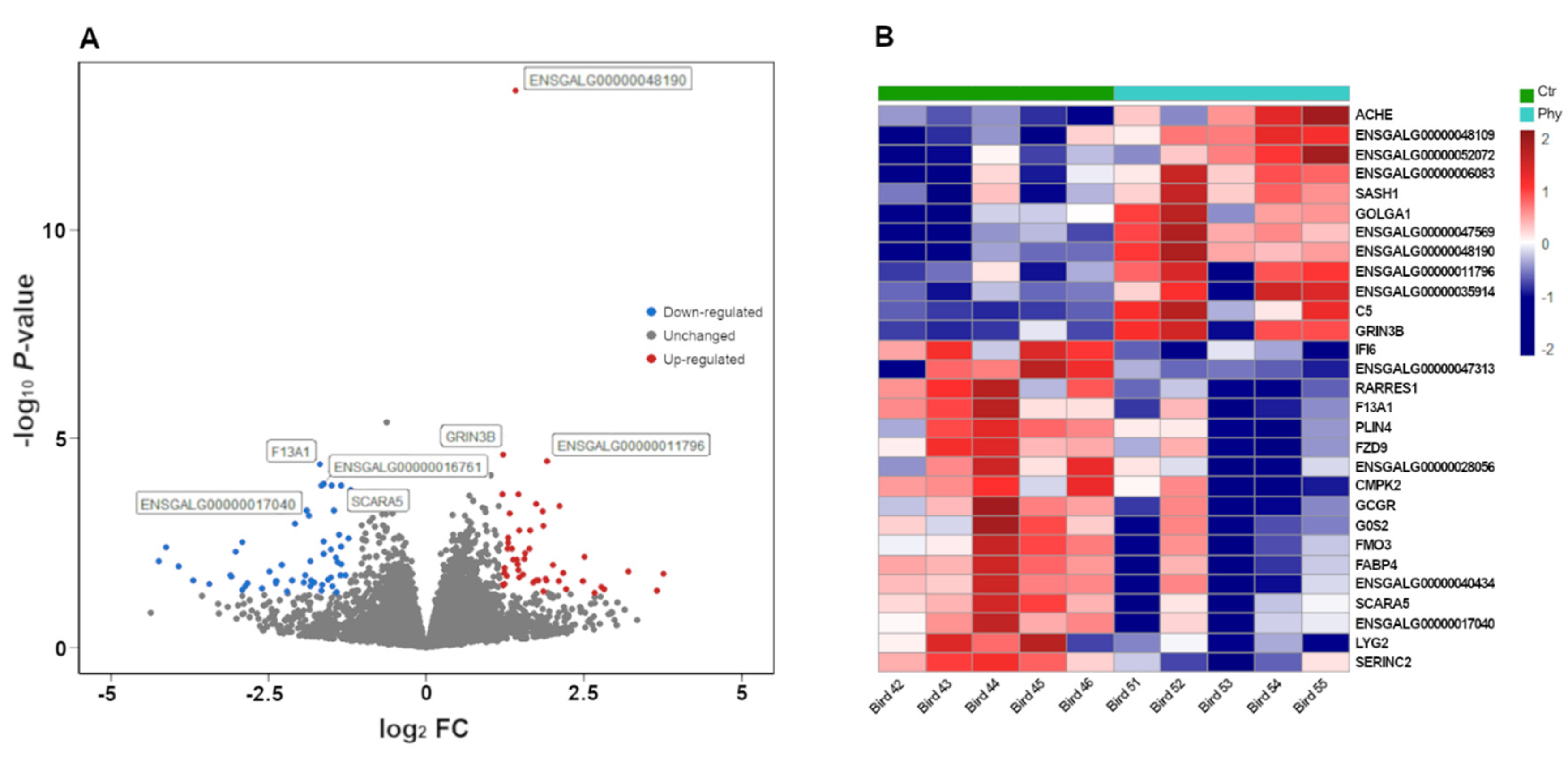
3.1. Sequencing Quality and Differential Analysis
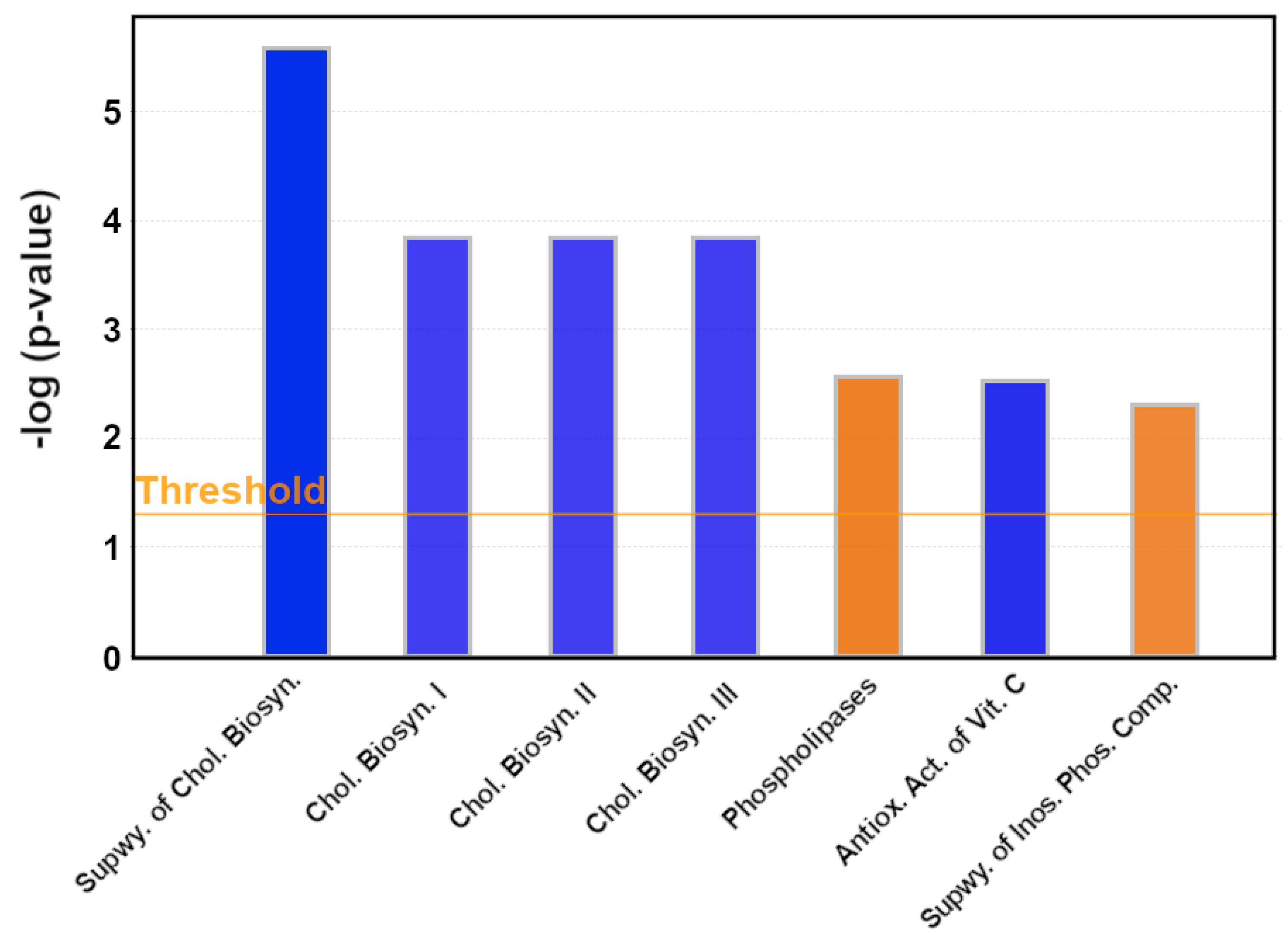
3.2. Pathway Analysis
3.3. Upstream Analysis
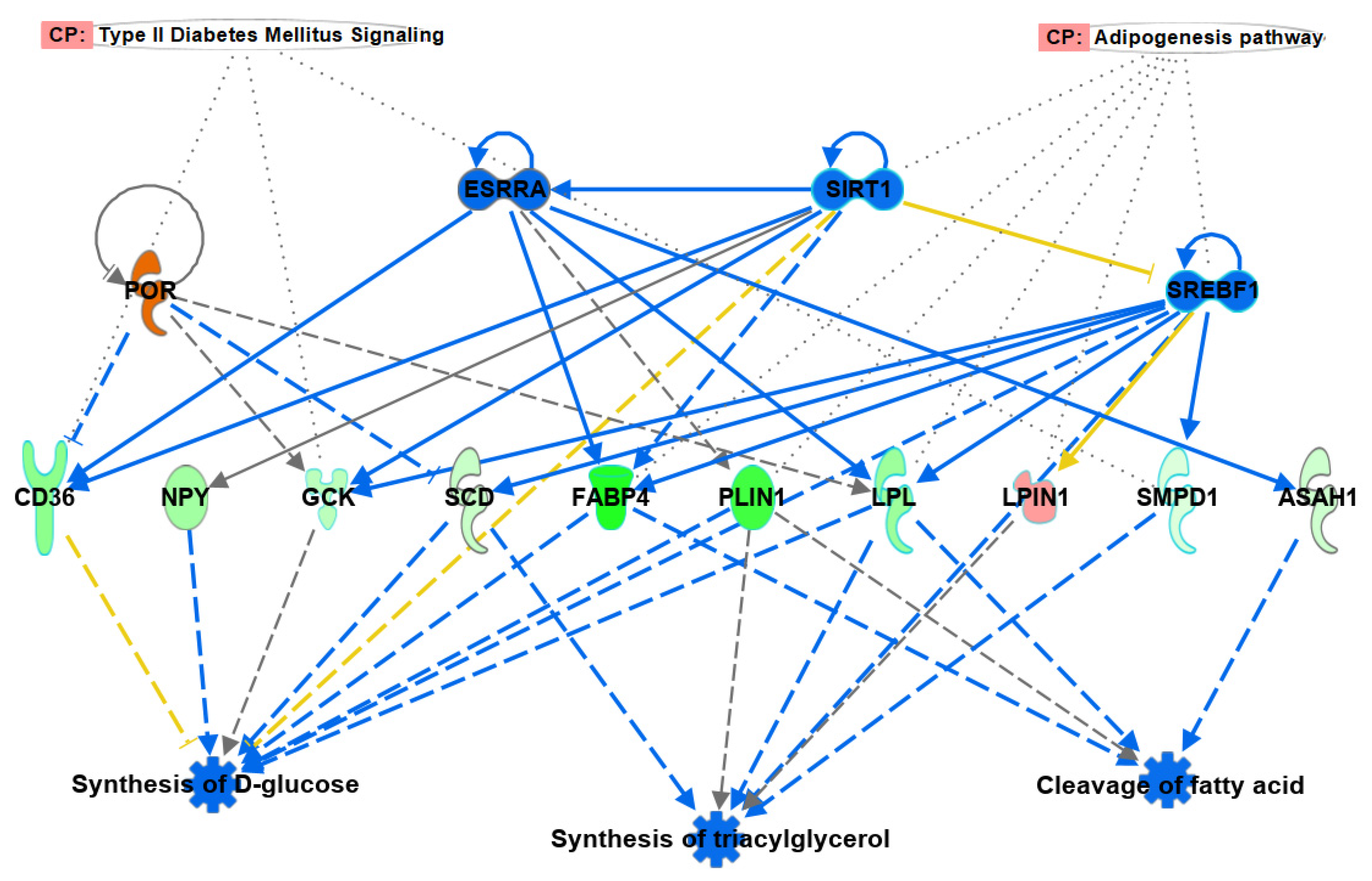
3.4. Regulatory Effects
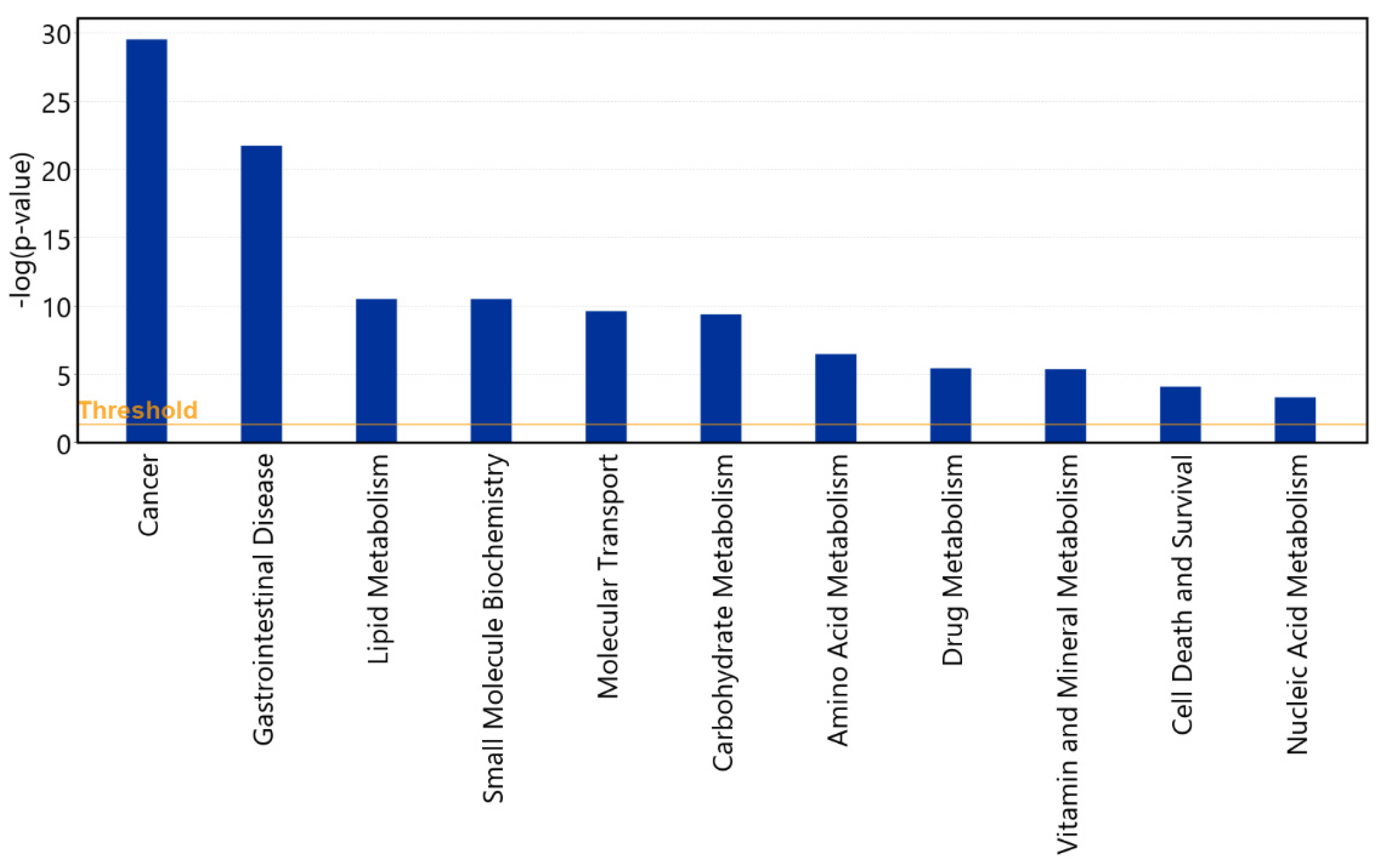
3.5. Diseases and Functions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ricke, S.C.; Dittoe, D.K.; Richardson, K.E. Formic acid as an antimicrobial for poultry production: A review. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Paiva, D.M.A. Necrotic enteritis: Applications for the poultry industry. J. Appl. Poult. Res. 2014, 23, 557–566. [Google Scholar] [CrossRef]
- Lee, J.Y.; Tsolis, R.M.; Baumler, A.J. The microbiome and gut homeostasis. Science 2022, 377, eabp9960. [Google Scholar] [CrossRef] [PubMed]
- Rinttilä, T.; Apajalahti, J. Intestinal microbiota and metabolites—Implications for broiler chicken health and performance. J. Appl. Poult. Res. 2013, 22, 647–658. [Google Scholar] [CrossRef]
- Yadav, S.; Jha, R. Strategies to modulate the intestinal microbiota and their effects on nutrient utilization, performance, and health of poultry. J. Anim. Sci. Biotechnol. 2019, 10, 2. [Google Scholar] [CrossRef] [Green Version]
- Oviedo-Rondón, E.O.; Hume, M.E.; Barbosa, N.A.; Sakomura, N.K.; Weber, G.; Wilson, J.W. Ileal and caecal microbial populations in broilers given specific essential oil blends and probiotics in two consecutive grow-outs. Avian Biol. Res. 2010, 3, 157–169. [Google Scholar] [CrossRef]
- Thompson, K.; Burkholder, K.; Patterson, J.; Applegate, T.J. Microbial ecology shifts in the ileum of broilers during feed withdrawal and dietary manipulations. Poult. Sci. 2008, 87, 1624–1632. [Google Scholar] [CrossRef]
- Shi, D.; Bai, L.; Qu, Q.; Zhou, S.; Yang, M.; Guo, S.; Li, Q.; Liu, C. Impact of gut microbiota structure in heat-stressed broilers. Poult. Sci. 2019, 98, 2405–2413. [Google Scholar] [CrossRef]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and environmental factors affecting the intestinal microbiota in chickens. Front. Microbiol. 2018, 9, 235. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Pielsticker, C.; Gerzova, L.; Rychlik, I.; Rautenschlein, S. The influence of age on Campylobacter jejuni infection in chicken. Dev. Comp. Immunol. 2016, 62, 58–71. [Google Scholar] [CrossRef]
- Camp, J.G.; Frank, C.L.; Lickwar, C.R.; Guturu, H.; Rube, T.; Wenger, A.M.; Chen, J.; Bejerano, G.; Crawford, G.E.; Rawls, J.F. Microbiota modulate transcription in the intestinal epithelium without remodeling the accessible chromatin landscape. Genome Res. 2014, 24, 1504–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowat, A.M.; Agace, W.W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 2014, 14, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Bastos, M.S.; Del Vesco, A.P.; Santana, T.P.; Santos, T.S.; de Oliveira Junior, G.M.; Fernandes, R.P.M.; Barbosa, L.T.; Gasparino, E. The role of cinnamon as a modulator of the expression of genes related to antioxidant activity and lipid metabolism of laying quails. PLoS ONE 2017, 12, e0189619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajagai, Y.S.; Petranyi, F.; Horyanto, D.; Batacan, R., Jr.; Lobo, E.; Ren, X.; Whitton, M.M.; Yu, S.J.; Kayal, A.; Stanley, D. Ileum transcriptional response to prolonged supplementation with phytogenic product containing menthol, carvacrol and carvone. Heliyon 2022, 8, e09131. [Google Scholar] [CrossRef]
- Bajagai, Y.S.; Steel, J.C.; Radovanovic, A.; Stanley, D. Prolonged continual consumption of oregano herb interferes with the action of steroid hormones and several drugs, and effects signaling across the brain-gut axis. Food Funct. 2021, 12, 726–738. [Google Scholar] [CrossRef]
- Sabino, M.; Capomaccio, S.; Cappelli, K.; Verini-Supplizi, A.; Bomba, L.; Ajmone-Marsan, P.; Cobellis, G.; Olivieri, O.; Pieramati, C.; Trabalza-Marinucci, M. Oregano dietary supplementation modifies the liver transcriptome profile in broilers: RNASeq analysis. Res. Vet. Sci. 2018, 117, 85–91. [Google Scholar] [CrossRef]
- Sampaio, L.A.; Pina, L.T.S.; Serafini, M.R.; Tavares, D.D.S.; Guimaraes, A.G. Antitumor effects of carvacrol and thymol: A systematic review. Front. Pharmacol. 2021, 12, 702487. [Google Scholar] [CrossRef]
- Hashemipour, H.; Kermanshahi, H.; Golian, A.; Veldkamp, T. Effect of thymol and carvacrol feed supplementation on performance, antioxidant enzyme activities, fatty acid composition, digestive enzyme activities, and immune response in broiler chickens. Poult. Sci. 2013, 92, 2059–2069. [Google Scholar] [CrossRef]
- Barboza, J.N.; da Silva Maia Bezerra Filho, C.; Silva, R.O.; Medeiros, J.V.R.; de Sousa, D.P. An overview on the anti-inflammatory potential and antioxidant profile of eugenol. Oxid. Med. Cell. Longev. 2018, 2018, 3957262. [Google Scholar] [CrossRef]
- Muanda, F.; Kone, D.; Dicko, A.; Soulimani, R.; Younos, C. Phytochemical composition and antioxidant capacity of three malian medicinal plant parts. Evid. Based Complement. Alternat. Med. 2011, 2011, 674320. [Google Scholar] [CrossRef]
- Yanishlieva, N.V.; Marinova, E.M.; Gordon, M.H.; Raneva, V.G. Antioxidant activity and mechanism of action of thymol and carvacrol in two lipid systems. Food Chem. 1999, 64, 59–66. [Google Scholar] [CrossRef]
- Omonijo, F.A.; Liu, S.; Hui, Q.; Zhang, H.; Lahaye, L.; Bodin, J.C.; Gong, J.; Nyachoti, M.; Yang, C. Thymol improves barrier function and attenuates inflammatory responses in porcine intestinal epithelial cells during Lipopolysaccharide (LPS)-induced inflammation. J. Agric. Food Chem. 2019, 67, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Youdim, K.A.; Deans, S.G. Effect of thyme oil and thymol dietary supplementation on the antioxidant status and fatty acid composition of the ageing rat brain. Br. J. Nutr. 2000, 83, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinus, R.D.; Sheoran, S.N.; Maan, N.; Tewatia, B. Potential benefits of herbal supplements in poultry feed: A review. Pharm. Innov. J. 2018, 7, 651–656. [Google Scholar]
- Flees, J.J.; Ganguly, B.; Dridi, S. Phytogenic feed additives improve broiler feed efficiency via modulation of intermediary lipid and protein metabolism-related signaling pathways. Poult. Sci. 2021, 100, 100963. [Google Scholar] [CrossRef]
- Wati, T.; Ghosh, T.K.; Syed, B.; Haldar, S. Comparative efficacy of a phytogenic feed additive and an antibiotic growth promoter on production performance, caecal microbial population and humoral immune response of broiler chickens inoculated with enteric pathogens. Anim. Nutr. 2015, 1, 213–219. [Google Scholar] [CrossRef]
- Murugesan, G.R.; Syed, B.; Haldar, S.; Pender, C. Phytogenic feed additives as an alternative to antibiotic growth promoters in broiler chickens. Front. Vet. Sci. 2015, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.J.; Bajagai, Y.S.; Petranyi, F.; Stanley, D. Phytogen improves performance during Spotty Liver Disease by impeding bacterial metabolism and pathogenicity. Appl. Environ. Microbiol. 2022, 88, e00758-22. [Google Scholar] [CrossRef]
- Willson, N.L.; Van, T.T.H.; Bhattarai, S.P.; Courtice, J.M.; McIntyre, J.R.; Prasai, T.P.; Moore, R.J.; Walsh, K.; Stanley, D. Feed supplementation with biochar may reduce poultry pathogens, including Campylobacter hepaticus, the causative agent of Spotty Liver Disease. PLoS ONE 2019, 14, e0214471. [Google Scholar] [CrossRef]
- Connerton, P.L.; Richards, P.J.; Lafontaine, G.M.; O’Kane, P.M.; Ghaffar, N.; Cummings, N.J.; Smith, D.L.; Fish, N.M.; Connerton, I.F. The effect of the timing of exposure to Campylobacter jejuni on the gut microbiome and inflammatory responses of broiler chickens. Microbiome 2018, 6, 88. [Google Scholar] [CrossRef]
- Schokker, D.; Veninga, G.; Vastenhouw, S.A.; Bossers, A.; de Bree, F.M.; Kaal-Lansbergen, L.M.T.E.; Rebel, J.M.J.; Smits, M.A. Early life microbial colonization of the gut and intestinal development differ between genetically divergent broiler lines. BMC Genomics 2015, 16, 418. [Google Scholar] [CrossRef] [PubMed]
- Bajagai, Y.S.; Radovanovic, A.; Steel, J.C.; Stanley, D. The effects of continual consumption of Origanum vulgare on liver transcriptomics. Animals 2021, 11, 398. [Google Scholar] [CrossRef] [PubMed]
- Baenke, F.; Peck, B.; Miess, H.; Schulze, A. Hooked on fat: The role of lipid synthesis in cancer metabolism and tumour development. Dis. Model. Mech. 2013, 6, 1353–1363. [Google Scholar] [CrossRef] [Green Version]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Bian, X.; Liu, R.; Meng, Y.; Xing, D.; Xu, D.; Lu, Z. Lipid metabolism and cancer. J. Exp. Med. 2021, 218, e20201606. [Google Scholar] [CrossRef] [PubMed]
- Bengoechea-Alonso, M.T.; Ericsson, J. SREBP in signal transduction: Cholesterol metabolism and beyond. Curr. Opin. Cell. Biol. 2007, 19, 215–222. [Google Scholar] [CrossRef]
- Shao, W.; Espenshade, P.J. Expanding roles for SREBP in metabolism. Cell. Metab. 2012, 16, 414–419. [Google Scholar] [CrossRef] [Green Version]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell. Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Garcia-Barros, M.; Coant, N.; Truman, J.P.; Snider, A.J.; Hannun, Y.A. Sphingolipids in colon cancer. Biochim. Biophys. Acta 2014, 1841, 773–782. [Google Scholar] [CrossRef] [Green Version]
- Nagahashi, M.; Tsuchida, J.; Moro, K.; Hasegawa, M.; Tatsuda, K.; Woelfel, I.A.; Takabe, K.; Wakai, T. High levels of sphingolipids in human breast cancer. J. Surg. Res. 2016, 204, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Gills, J.J.; Lopiccolo, J.; Dennis, P.A. Nelfinavir, a new anti-cancer drug with pleiotropic effects and many paths to autophagy. Autophagy 2008, 4, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Bruning, A.; Friese, K.; Burges, A.; Mylonas, I. Tamoxifen enhances the cytotoxic effects of nelfinavir in breast cancer cells. Breast Cancer Res. 2010, 12, R45. [Google Scholar] [CrossRef] [Green Version]
- Jensen, K.; Bikas, A.; Patel, A.; Kushchayeva, Y.; Costello, J.; McDaniel, D.; Burman, K.; Vasko, V. Nelfinavir inhibits proliferation and induces DNA damage in thyroid cancer cells. Endocr. Relat. Cancer 2017, 24, 147–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okubo, K.; Isono, M.; Asano, T.; Sato, A. Panobinostat and nelfinavir inhibit renal cancer growth by inducing endoplasmic reticulum stress. Anticancer Res. 2018, 38, 5615–5626. [Google Scholar] [CrossRef]
- Shirzad, M.; Taghdisi, M.H.; Dehdari, T.; Abolghasemi, J. Oral health education program among pre-school children: An application of health-promoting schools approach. Health Promot. Perspect. 2016, 6, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Wawruszak, A.; Borkiewicz, L.; Okon, E.; Kukula-Koch, W.; Afshan, S.; Halasa, M. Vorinostat (SAHA) and breast cancer: An overview. Cancers 2021, 13, 4700. [Google Scholar] [CrossRef] [PubMed]
- Okubo, K.; Isono, M.; Miyai, K.; Asano, T.; Sato, A. Fluvastatin potentiates anticancer activity of vorinostat in renal cancer cells. Cancer Sci. 2020, 111, 112–126. [Google Scholar] [CrossRef] [Green Version]
- Sanaei, M.; Kavoosi, F. Effect of vorinostat on INK4 family and HDACs 1, 2, and 3 in pancreatic cancer and hepatocellular carcinoma. Res. Pharm. Sci. 2021, 16, 260–268. [Google Scholar] [CrossRef]
- Beer, T.M.; Myrthue, A. Calcitriol in cancer treatment: From the lab to the clinic. Mol. Cancer Ther. 2004, 3, 373–381. [Google Scholar] [CrossRef]
- Paolini, M.; Abdel-Rahman, S.Z.; Sapone, A.; Pedulli, G.F.; Perocco, P.; Cantelli-Forti, G.; Legator, M.S. Beta-carotene: A cancer chemopreventive agent or a co-carcinogen? Mutat. Res. 2003, 543, 195–200. [Google Scholar] [CrossRef]
- Chen, Q.H.; Wu, B.K.; Pan, D.; Sang, L.X.; Chang, B. Beta-carotene and its protective effect on gastric cancer. World J. Clin. Cases 2021, 9, 6591–6607. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.E.; Kwon, M.; Kim, Y.S.; Kim, Y.; Chung, M.G.; Heo, S.C.; Kim, Y. Beta-carotene regulates cancer stemness in colon cancer in vivo and in vitro. Nutr. Res. Pract. 2022, 16, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Dulinska, J.; Gil, D.; Zagajewski, J.; Hartwich, J.; Bodzioch, M.; Dembinska-Kiec, A.; Langmann, T.; Schmitz, G.; Laidler, P. Different effect of beta-carotene on proliferation of prostate cancer cells. Biochim. Biophys. Acta 2005, 1740, 189–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; He, L.; Wu, Y.; Zhang, Y. Carvacrol affects breast cancer cells through TRPM7 mediated cell cycle regulation. Life Sci. 2021, 266, 118894. [Google Scholar] [CrossRef]
- Ahmad, A.; Ansari, I.A. Carvacrol exhibits chemopreventive potential against cervical cancer cells via caspase-dependent apoptosis and abrogation of cell cycle progression. Anticancer Agents Med. Chem. 2021, 21, 2224–2235. [Google Scholar] [CrossRef]
- Vimala, K.; Kannan, S. Phyto-drug conjugated nanomaterials enhance apoptotic activity in cancer. Adv. Protein Chem. Struct. Biol. 2021, 125, 275–305. [Google Scholar] [CrossRef]
- Brandt, C.; McFie, P.J.; Stone, S.J. Biochemical characterization of human acyl coenzyme A: 2-monoacylglycerol acyltransferase-3 (MGAT3). Biochem. Biophys. Res. Commun. 2016, 475, 264–270. [Google Scholar] [CrossRef]
- Ning, T.; Zou, Y.; Yang, M.; Lu, Q.; Chen, M.; Liu, W.; Zhao, S.; Sun, Y.; Shi, J.; Ma, Q.; et al. Genetic interaction of DGAT2 and FAAH in the development of human obesity. Endocrine 2017, 56, 366–378. [Google Scholar] [CrossRef]
- Mao, H.; Yin, Z.; Wang, M.; Zhang, W.; Raza, S.H.A.; Althobaiti, F.; Qi, L.; Wang, J. Expression of DGAT2 gene and its associations with intramuscular fat content and breast muscle fiber characteristics in domestic pigeons (Columba livia). Front. Vet. Sci. 2022, 9, 847363. [Google Scholar] [CrossRef]
- Mao, H.G.; Dong, X.Y.; Cao, H.Y.; Xu, N.Y.; Yin, Z.Z. Association of DGAT2 gene polymorphisms with carcass and meat quality traits in domestic pigeons (Columba livia). Br. Poult. Sci. 2018, 59, 149–153. [Google Scholar] [CrossRef]
- Zang, L.; Wang, Y.; Sun, B.; Zhang, X.; Yang, C.; Kang, L.; Zhao, Z.; Jiang, Y. Identification of a 13 bp indel polymorphism in the 3’-UTR of DGAT2 gene associated with backfat thickness and lean percentage in pigs. Gene 2016, 576, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.F.; Choi, S.H.; Li, Q.; Wang, Y.; Sun, B.; Tang, L.; Wang, E.Z.; Hua, H.; Li, X.Z. Overexpression of DGAT2 stimulates lipid droplet formation and triacylglycerol accumulation in bovine satellite cells. Animals 2022, 12, 1847. [Google Scholar] [CrossRef] [PubMed]
- Birchenough, G.M.; Johansson, M.E.; Gustafsson, J.K.; Bergstrom, J.H.; Hansson, G.C. New developments in goblet cell mucus secretion and function. Mucosal. Immunol. 2015, 8, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.E.; Hansson, G.C. Is the intestinal goblet cell a major immune cell? Cell Host Microbe 2014, 15, 251–252. [Google Scholar] [CrossRef] [Green Version]
- Van der Sluis, M.; De Koning, B.A.; De Bruijn, A.C.; Velcich, A.; Meijerink, J.P.; Van Goudoever, J.B.; Buller, H.A.; Dekker, J.; Van Seuningen, I.; Renes, I.B.; et al. Muc2-deficient mice spontaneously develop colitis, indicating that MUC2 is critical for colonic protection. Gastroenterology 2006, 131, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Saif, L.J. Goblet cell depletion in small intestinal villous and crypt epithelium of conventional nursing and weaned pigs infected with porcine epidemic diarrhea virus. Res. Vet. Sci. 2017, 110, 12–15. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Description (IPK Database) |
|---|---|
| Vorinostat | Anticancer drug |
| Salirasib | Anticancer drug |
| Tanespimycin | Anticancer drug |
| Arsenic trioxide | Anticancer drug |
| Sirolimus | Anticancer drug; Immunosuppressant |
| Dimethyl sulfoxide | Anti-inflammatory agent |
| Nelfinavir | Antiviral drug |
| Calcitriol | Calcium regulator |
| Genistein | Angiogenesis inhibitor |
| Isoquercitrin | Anti-cachexia drug |
| Streptozocin | Anticancer drug |
| Methylprednisolone | Anti-inflammatory and immunosuppressive medication. |
| Norephedrine | Direct agonist of adrenergic receptors |
| 17-alpha-ethinylestradiol | Estrogen used widely in birth control pills |
| Clozapine | Psychiatric medication, antipsychotic (schizophrenia) |
| Rosuvastatin | Statin medication used to prevent cardiovascular disease and treat abnormal lipids |
| Rosiglitazone | The antidiabetic drug, works as an insulin sensitiser |
| Pitavastatin | For high blood cholesterol |
| Clofibrate | For high blood cholesterol |
| Cerivastatin | For high blood cholesterol |
| Ezetimibe | For high blood cholesterol |
| Lovastatin | For high blood cholesterol |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.J.; Bajagai, Y.S.; Petranyi, F.; de las Heras-Saldana, S.; Van, T.T.H.; Stanley, D. Phytogenic Blend Improves Intestinal Health and Reduces Obesity, Diabetes, Cholesterol and Cancers: A Path toward Customised Supplementation. Antibiotics 2022, 11, 1428. https://doi.org/10.3390/antibiotics11101428
Yu SJ, Bajagai YS, Petranyi F, de las Heras-Saldana S, Van TTH, Stanley D. Phytogenic Blend Improves Intestinal Health and Reduces Obesity, Diabetes, Cholesterol and Cancers: A Path toward Customised Supplementation. Antibiotics. 2022; 11(10):1428. https://doi.org/10.3390/antibiotics11101428
Chicago/Turabian StyleYu, Sung J., Yadav S. Bajagai, Friedrich Petranyi, Sara de las Heras-Saldana, Thi Thu Hao Van, and Dragana Stanley. 2022. "Phytogenic Blend Improves Intestinal Health and Reduces Obesity, Diabetes, Cholesterol and Cancers: A Path toward Customised Supplementation" Antibiotics 11, no. 10: 1428. https://doi.org/10.3390/antibiotics11101428
APA StyleYu, S. J., Bajagai, Y. S., Petranyi, F., de las Heras-Saldana, S., Van, T. T. H., & Stanley, D. (2022). Phytogenic Blend Improves Intestinal Health and Reduces Obesity, Diabetes, Cholesterol and Cancers: A Path toward Customised Supplementation. Antibiotics, 11(10), 1428. https://doi.org/10.3390/antibiotics11101428