

Ciprofloxacin Concentrations 1/1000th the MIC Can Select for Antimicrobial Resistance in N. gonorrhoeae—Important Implications for Maximum Residue Limits in Food
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Determination of Minimum Selection Concentrations (MSCs)
2.1.1. Bacterial Strains and Growth Conditions
2.1.2. MSC Determination
2.1.3. MSCdenovo
2.1.4. MSCselect
- Growth rate. The intrinsic growth rate of the population, r, is the growth rate that would occur if there were no restrictions imposed on growth. Growthcurver uses the nonlinear least-squares Levenberg–Marquardt algorithm to determine r.
- Area under the curve. Growthcurver calculates the area under the logistic curve (AUC). This integrates information from the carrying capacity, growth rate and the population size at time 0.
2.1.5. Whole-Genome Sequencing
2.2. Ecological Association between Quinolone Use and Ciprofloxacin MICs
2.2.1. Quinolone Use for Animal Food Production Data
2.2.2. Quinolone Consumption in Humans
2.2.3. N. gonorrhoeae Ciprofloxacin Resistance Data
2.2.4. Statistical Analysis
3. Results
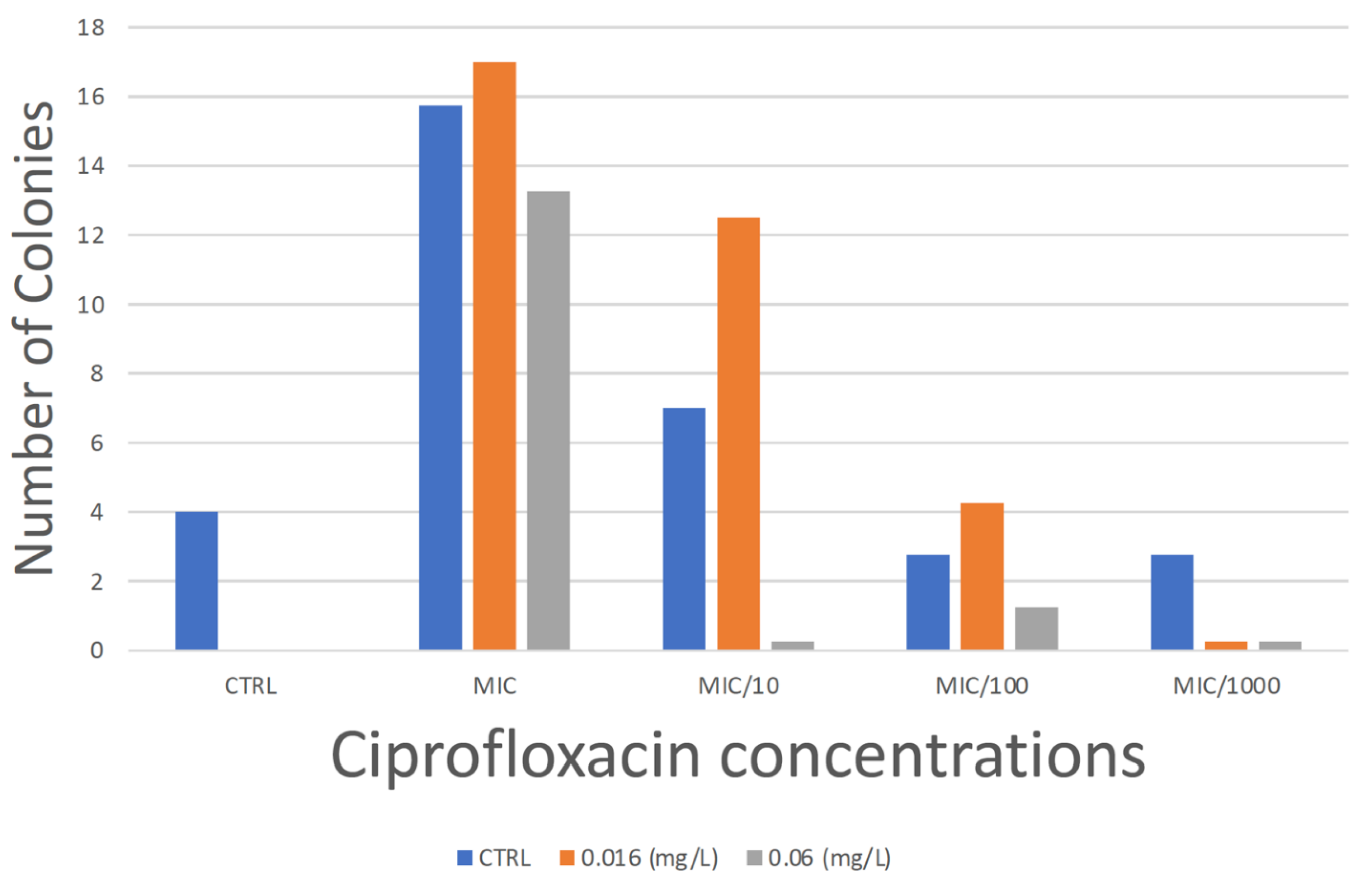
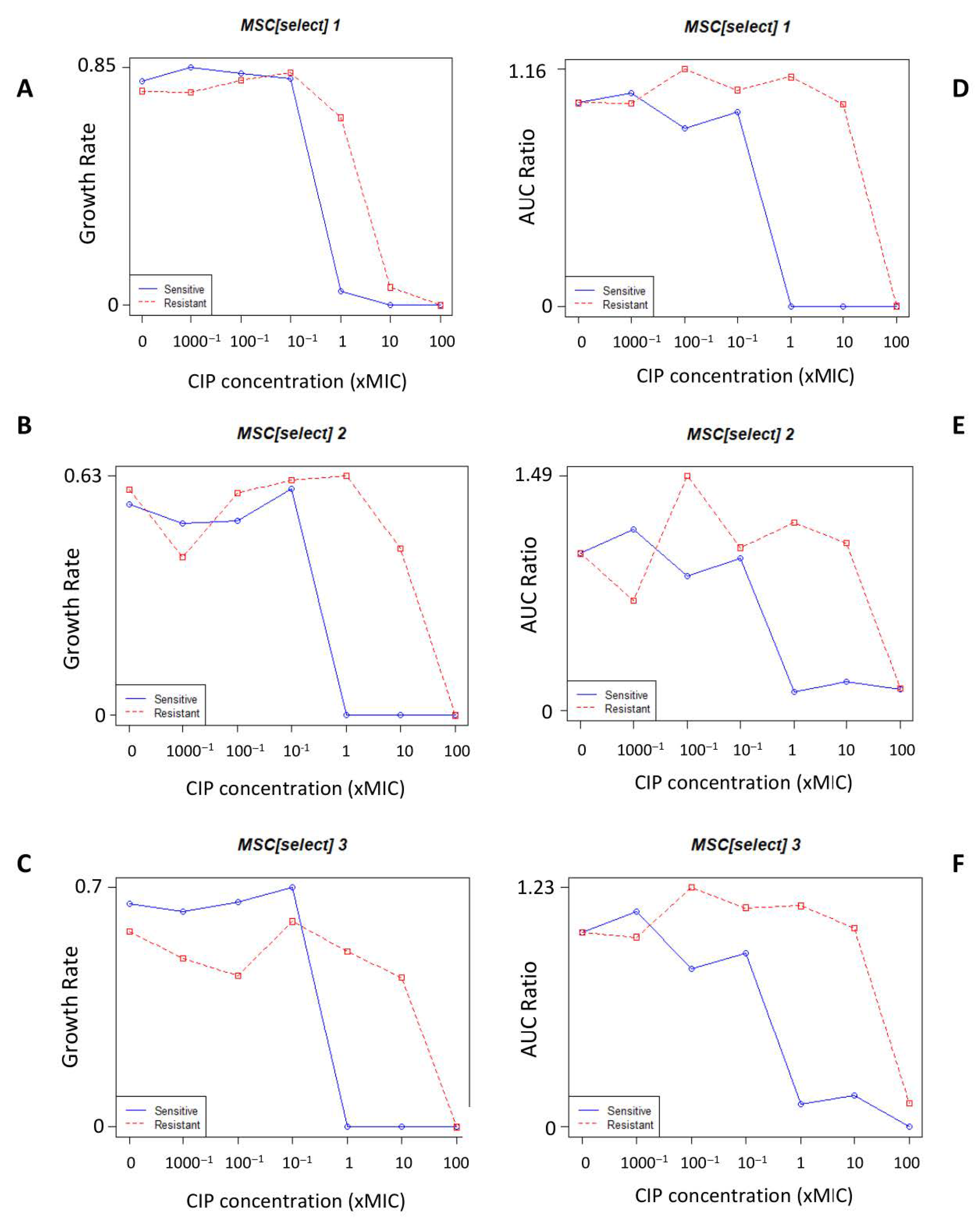
3.1. Minimum Selection Concentration
3.1.1. MSCdenovo
3.1.2. MSCselect
3.1.3. Ecological Association between Quinolone Consumption and Ciprofloxacin Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bash, M.C.; McKnew, D.L.; Tapsall, J.W. Antibiotic Resistance in Neisseria. In Antimicrobial Drug Resistance; Springer: Berlin/Heidelberg, Germany, 2009; pp. 763–782. [Google Scholar]
- Kenyon, C.; Buyze, J.; Wi, T. Antimicrobial consumption and susceptibility of Neisseria gonorrhoeae: A global ecological analysis. Front. Med. 2018, 5, 329. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C.; Laumen, J.; Van Dijck, C.; De Baetselier, I.; Abdelatti, S.; Manoharan-Basil, S.S.; Unemo, M. Gonorrhoea treatment combined with population-level general cephalosporin and quinolone consumption may select for Neisseria gonorrhoeae antimicrobial resistance at the levels of NG-MAST genogroup: An ecological study in Europe. J. Glob. Antimicrob. Resist. 2020, 23, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Spratt, B.G.; Bowler, L.D.; Zhang, Q.Y.; Zhou, J.; Smith, J.M. Role of interspecies transfer of chromosomal genes in the evolution of penicillin resistance in pathogenic and commensal Neisseria species. J. Mol. Evol. 1992, 34, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Potts, C.C.; Retchless, A.C.; McNamara, L.A.; Marasini, D.; Reese, N.; Swint, S.; Hu, F.; Sharma, S.; Blain, A.E.; Lonsway, D.; et al. Acquisition of ciprofloxacin resistance among an expanding clade of β-lactamase positive, serogroup Y Neisseria meningitidis in the United States. Clin. Infect. Dis. 2021, 73, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhang, C.; Zhang, X.; Chen, M. Meningococcal quinolone resistance originated from several commensal Neisseria species. Antimicrob. Agents Chemother. 2019, 64, e01494-19. [Google Scholar] [CrossRef]
- Kenyon, C.; Manoharan-Basil, S.S.; Van Dijck, C. Gonococcal resistance can be viewed productively as part of a syndemic of antimicrobial resistance: An ecological analysis of 30 European countries. Antimicrob. Resist. Infect. Control 2020, 9, 97. [Google Scholar] [CrossRef]
- Gu, B.; Cao, Y.; Pan, S.; Zhuang, L.; Yu, R.; Peng, Z.; Qian, H.; Wei, Y.; Zhao, L.; Liu, G.; et al. Comparison of the prevalence and changing resistance to nalidixic acid and ciprofloxacin of Shigella between Europe–America and Asia–Africa from 1998 to 2009. Int. J. Antimicrob. Agents 2012, 40, 9–17. [Google Scholar] [CrossRef]
- Lu, P.-L.; Liu, Y.-C.; Toh, H.-S.; Lee, Y.-L.; Liu, Y.-M.; Ho, C.-M.; Huang, C.C.; Liu, C.E.; Ko, W.C.; Wang, J.H.; et al. Epidemiology and antimicrobial susceptibility profiles of Gram-negative bacteria causing urinary tract infections in the Asia-Pacific region: 2009–2010 results from the Study for Monitoring Antimicrobial Resistance Trends (SMART). Int. J. Antimicrob. Agents 2012, 40, S37–S43. [Google Scholar] [CrossRef]
- Song, J.H. Antimicrobial resistance control in Asia. Monitoring, surveillance and national plans. AMR Control 2015, 41–45. [Google Scholar]
- Yam, E.L.Y.; Hsu, L.Y.; Yap, E.P.; Yeo, T.W.; Lee, V.; Schlundt, J.; Lwin, M.O.; Limmathurotsakul, D.; Jit, M.; Dedon, P.; et al. Antimicrobial Resistance in the Asia Pacific region: A meeting report. Antimicrob. Resist. Infect. Control 2019, 8, 202. [Google Scholar] [CrossRef] [Green Version]
- Fiore, M.A.; Raisman, J.C.; Wong, N.R.H.; Hudson, A.O.; Wadsworth, C.B. Exploration of the Neisseria Resistome Reveals Resistance Mechanisms in Commensals That May Be Acquired by N. gonorrhoeae through Horizontal Gene Transfer. Antibiotics 2020, 9, 656. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.F.; Chen, M.L. Prevalence, sequence type, and quinolone resistance of Neisseria lactamica carried in children younger than 15 years in Shanghai, China. J. Infect. 2020, 80, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Liao, M.M.; Gu, W.M.; Bell, K.; Wu, L.; Eng, N.F.; Zhang, C.G.; Chen, Y.; Jolly, A.M.; Dillon, J.A.R. Antimicrobial susceptibility and molecular determinants of quinolone resistance in Neisseria gonorrhoeae isolates from Shanghai. J. Antimicrob. Chemother. 2006, 58, 868–872. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2018/2019. EFSA J. 2021, 19, e06490. [Google Scholar]
- Muloi, D.; Ward, M.J.; Pedersen, A.B.; Fevre, E.M.; Woolhouse, M.E.; van Bunnik, B.A. Are food animals responsible for transfer of antimicrobial-resistant Escherichia coli or their resistance determinants to human populations? A systematic review. Foodborne Pathog. Dis. 2018, 15, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Ben, Y.; Fu, C.; Hu, M.; Liu, L.; Wong, M.H.; Zheng, C. Human health risk assessment of antibiotic resistance associated with antibiotic residues in the environment: A review. Environ. Res. 2019, 169, 483–493. [Google Scholar] [CrossRef]
- Cantas, L.; Shah, S.Q.; Cavaco, L.M.; Manaia, C.M.; Walsh, F.; Popowska, M.; Garelick, H.; Bürgmann, H.; Sørum, H. A brief multi-disciplinary review on antimicrobial resistance in medicine and its linkage to the global environmental microbiota. Front. Microbiol. 2013, 4, 96. [Google Scholar] [CrossRef] [Green Version]
- Răpuntean, S.; FiŃ, N.; Chirilă, F.; Nadăş, G.; Cosmina, C.; Călina, D.; Drig, C. Physical factors action on Neisseria spp. strains isolated from animals. Bull. UASVM 2008, 65, 1. [Google Scholar]
- Sneath, P.; Barrett, S. A new species of Neisseria from the dental plaque of the domestic cow, Neisseria dentiae sp. nov. Lett. Appl. Microbiol. 1996, 23, 355–358. [Google Scholar] [CrossRef]
- Kayalvizhi, N.; Anthony, T.; Gunasekaran, P. Characterization of the Predominant Bacteria Associated with Sheep and Goat Skin. J. Am. Leather Chem. Assoc. 2008, 103, 182–187. [Google Scholar]
- Liu, G.; Tang, C.M.; Exley, R.M. Non-pathogenic Neisseria: Members of an abundant, multi-habitat, diverse genus. Microbiology 2015, 161, 1297–1312. [Google Scholar] [CrossRef] [PubMed]
- Gullberg, E.; Albrecht, L.M.; Karlsson, C.; Sandegren, L.; Andersson, D.I. Selection of a multidrug resistance plasmid by sublethal levels of antibiotics and heavy metals. mBio 2014, 5, e01918-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanton, I.C.; Murray, A.K.; Zhang, L.; Snape, J.; Gaze, W.H. Evolution of antibiotic resistance at low antibiotic concentrations including selection below the minimal selective concentration. Commun. Biol. 2020, 3, 467. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Fong, A.; Becket, E.; Yuan, J.; Tamae, C.; Medrano, L.; Maiz, M.; Wahba, C.; Lee, C.; Lee, K.; et al. Selective advantage of resistant strains at trace levels of antibiotics: A simple and ultrasensitive color test for detection of antibiotics and genotoxic agents. Antimicrob. Agents Chemother. 2011, 55, 1204–1210. [Google Scholar] [CrossRef] [PubMed]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilbäck, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Qiu, W.; Li, Y.; Liu, L. Antibiotic residues in poultry food in Fujian Province of China. Food Addit. Contam. Part B Surveill. 2020, 13, 177–184. [Google Scholar] [CrossRef]
- Huang, L.; Mo, Y.; Wu, Z.; Rad, S.; Song, X.; Zeng, H.; Bashir, S.; Kang, B.; Chen, Z. Occurrence, distribution, and health risk assessment of quinolone antibiotics in water, sediment, and fish species of Qingshitan reservoir, South China. Sci. Rep. 2020, 10, 15777. [Google Scholar] [CrossRef]
- Zheng, N.; Wang, J.; Han, R.; Xu, X.; Zhen, Y.; Qu, X.; Sun, P.; Li, S.; Yu, Z. Occurrence of several main antibiotic residues in raw milk in 10 provinces of China. Food Addit. Contam. Part B 2013, 6, 84–89. [Google Scholar] [CrossRef]
- Wang, Q.-J.; Mo, C.-H.; Li, Y.-W.; Gao, P.; Tai, Y.-P.; Zhang, Y.; Ruan, Z.L.; Xu, J.W. Determination of four fluoroquinolone antibiotics in tap water in Guangzhou and Macao. Environ. Pollut. 2010, 158, 2350–2358. [Google Scholar]
- Ben, Y.; Hu, M.; Zhang, X.; Wu, S.; Wong, M.H.; Wang, M.; Andrews, C.B.; Zheng, C. Efficient detection and assessment of human exposure to trace antibiotic residues in drinking water. Water Res. 2020, 175, 115699. [Google Scholar] [CrossRef]
- Kraupner, N.; Ebmeyer, S.; Hutinel, M.; Fick, J.; Flach, C.F.; Larsson, D.G.J. Selective concentrations for trimethoprim resistance in aquatic environments. Environ. Int. 2020, 144, 106083. [Google Scholar] [CrossRef] [PubMed]
- Unemo, M.; Shafer, W.M. Antimicrobial resistance in Neisseria gonorrhoeae in the 21st century: Past, evolution, and future. Clin. Microbiol. Rev. 2014, 27, 587–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhoeven, E.; Abdellati, S.; Nys, P.; Laumen, J.; De Baetselier, I.; Crucitti, T.; Kenyon, C. Construction and optimization of a ‘NG Morbidostat’-An automated continuous-culture device for studying the pathways towards antibiotic resistance in Neisseria gonorrhoeae. F1000Research 2019, 8, 560. [Google Scholar] [CrossRef] [PubMed]
- Laumen, J.G.E.; Manoharan-Basil, S.S.; Verhoeven, E.; Abdellati, S.; De Baetselier, I.; Crucitti, T.; Xavier, B.B.; Chapelle, S.; Lammens, C.; Van Dijck, C.; et al. Molecular pathways to high-level azithromycin resistance in Neisseria gonorrhoeae. J. Antimicrob. Chemother. 2021, 76, 1752–1758. [Google Scholar] [CrossRef] [PubMed]
- Sprouffske, K.; Wagner, A. Growthcurver: An R package for obtaining interpretable metrics from microbial growth curves. BMC Bioinform. 2016, 17, 172. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Van Boeckel, T.P.; Glennon, E.E.; Chen, D.; Gilbert, M.; Robinson, T.P.; Grenfell, B.T.; Levin, S.A.; Bonhoeffer, S.; Laxminarayan, R. Reducing antimicrobial use in food animals. Science 2017, 357, 1350–1352. [Google Scholar] [CrossRef] [Green Version]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef] [Green Version]
- Wi, T.; Lahra, M.M.; Ndowa, F.; Bala, M.; Dillon, J.R.; Ramon-Pardo, P.; Eremin, S.R.; Bolan, G.; Unemo, M. Antimicrobial resistance in Neisseria gonorrhoeae: Global surveillance and a call for international collaborative action. PLoS Med. 2017, 14, e1002344. [Google Scholar] [CrossRef] [Green Version]
- Ji, K.; Kho, Y.; Park, C.; Paek, D.; Ryu, P.; Paek, D.; Kim, M.; Kim, P.; Choi, K. Influence of water and food consumption on inadvertent antibiotics intake among general population. Environ. Res. 2010, 110, 641–649. [Google Scholar] [CrossRef]
- Wang, H.; Tang, C.; Yang, J.; Wang, N.; Jiang, F.; Xia, Q.; He, G.; Chen, Y.; Jiang, Q. Predictors of urinary antibiotics in children of Shanghai and health risk assessment. Environ. Int. 2018, 121, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Zhu, F.; Zheng, D.Y.; Gao, M.M.; Guo, B.F.; Zhang, N.; Meng, Y.; Wu, G.L.; Zhou, Y.L.; Huo, X. Detection of antibiotics in the urine of children and pregnant women in Jiangsu, China. Environ. Res. 2021, 196, 110945. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Duan, Y.J.; Wang, S.P.; Wang, L.T.; Hou, Z.L.; Cui, Y.X.; Hou, J.; Das, R.; Mao, D.Q.; Luo, Y. Occurrence and distribution of clinical and veterinary antibiotics in the faeces of a Chinese population. J. Hazard. Mater. 2020, 383, 121129. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.; Kho, Y.L.; Park, Y.; Choi, K. Influence of a five-day vegetarian diet on urinary levels of antibiotics and phthalate metabolites: A pilot study with “Temple Stay” participants. Environ. Res. 2010, 110, 375–382. [Google Scholar] [CrossRef]
- ECDC; EFSA; EMA. ECDC/EFSA/EMA second joint report on the integrated analysis of the consumption of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from humans and food-producing animals: Joint Interagency Antimicrobial Consumption and Resistance Analysis (JIACRA) Report. EFSA J. 2017, 15, e04872. [Google Scholar] [CrossRef]
- Muaz, K.; Riaz, M.; Akthar, S.; Park, S.; Ismail, A. Antibiotic Residues in Chicken Meat: Global Prevalence, Threats, and Decontamination Strategies: A Review. J. Food Prot. 2018, 81, 619–627. [Google Scholar] [CrossRef]
- Jammoul, A.; El Darra, N. Evaluation of Antibiotics Residues in Chicken Meat Samples in Lebanon. Antibiotics 2019, 8, 69. [Google Scholar] [CrossRef] [Green Version]
- Quintanilla, P.; Doménech, E.V.A.; Escriche, I.; Beltrán, M.C.; Molina, M.P. Food Safety Margin Assessment of Antibiotics: Pasteurized Goat’s Milk and Fresh Cheese. J. Food Prot. 2019, 82, 1553–1559. [Google Scholar] [CrossRef]
- Vanbaelen, T.; Van Dijck, C.; Laumen, J.; Gonzalez, N.; De Baetselier, I.; Manoharan-Basil, S.S.; De Block, T.; Kenyon, C. Global epidemiology of antimicrobial resistance in commensal Neisseria: A systematic review. Int. J. Med. Microbiol. 2022, 312, 151551. [Google Scholar] [CrossRef]
- Klümper, U.; Recker, M.; Zhang, L.; Yin, X.; Zhang, T.; Buckling, A.; Gaze, W.H. Selection for antimicrobial resistance is reduced when embedded in a natural microbial community. ISME J. 2019, 13, 2927–2937. [Google Scholar] [CrossRef]
- Murray, A.K.; Stanton, I.; Gaze, W.H.; Snape, J. Dawning of a new ERA: Environmental Risk Assessment of antibiotics and their potential to select for antimicrobial resistance. Water Res. 2021, 200, 117233. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.Y.S.; Hatzis, C.L.; Lau, A.; Williamson, D.A.; Chow, E.P.F.; Fairley, C.K.; Hocking, J.S. Treatment efficacy for pharyngeal Neisseria gonorrhoeae: A systematic review and meta-analysis of randomized controlled trials. J. Antimicrob. Chemother. 2020, 75, 3109–3119. [Google Scholar] [CrossRef]
- de Block, T.; Laumen, J.G.E.; Van Dijck, C.; Abdellati, S.; De Baetselier, I.; Manoharan-Basil, S.S.; Van den Bossche, D.; Kenyon, C. WGS of commensal Neisseria reveals acquisition of a new ribosomal protection protein (MsrD) as a possible explanation for high level azithromycin resistance in Belgium. Pathogens 2021, 10, 384. [Google Scholar] [CrossRef]
- Aneesh, T.P.; Jose, A.; Chandran, L.; Zachariah, S.M. Pharmacogenomics: The right drug to the right person. J. Clin. Med. Res. 2009, 1, 191–194. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, K.; Ma, M.; So, M. A Natural Mouse Model for Neisseria Persistent Colonization. Methods Mol. Biol. 2019, 1997, 403–412. [Google Scholar] [CrossRef]
- Ma, M.; Powell, D.A.; Weyand, N.J.; Rhodes, K.A.; Rendon, M.A.; Frelinger, J.A.; So, M. A Natural Mouse Model for Neisseria Colonization. Infect. Immun. 2018, 86, e00839-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO/WHO. Codex Alimentarius Commission (CAC): Maximum Residue Limits. Available online: http://www.fao.org/fao-who-codexalimentarius/codex-texts/maximum-residue-limits/en/ (accessed on 1 February 2021).
- European Comission. 37/2010. Commission Regulation (EU) No. 37/2010 of 22 December 2009 on pharmacologically active substances and their classification regarding maximum residue limits in foodstuffs of animal origin. Off. J. Eur. Union 2010, 15, 1–72. [Google Scholar]
- The European Agency for the Evaluation of Medicinal Products. Committee for Veterinary Medicinal Products: Enrofloxacin Summary Report (2); EMEA: London, UK, 1998. [Google Scholar]
- Mitchell, J.; Griffiths, M.; McEwen, S.; McNab, W.; Yee, A. Antimicrobial drug residues in milk and meat: Causes, concerns, prevalence, regulations, tests, and test performance. J. Food Prot. 1998, 61, 742–756. [Google Scholar] [CrossRef]
- Olesky, M.; Hobbs, M.; Nicholas, R.A. Identification and analysis of amino acid mutations in porin IB that mediate intermediate-level resistance to penicillin and tetracycline in Neisseria gonorrhoeae. Antimicrob. Agents Chemother. 2002, 46, 2811–2820. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Model 1 | Model 2 | Model 3 | |
|---|---|---|---|
| Quinolones Humans | 0.02 (−0.007–0.043) | - | 0.2 (0.001–0.046) * |
| Quinolones Food animals | - | 0.2 (0.02–0.38) * | 0.2 (0.07–0.40) ** |
| N | 30 | 34 | 30 |
| R2 | 0.07 | 0.14 | 0.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, N.; Abdellati, S.; De Baetselier, I.; Laumen, J.G.E.; Van Dijck, C.; Block, T.d.; Manoharan-Basil, S.S.; Kenyon, C. Ciprofloxacin Concentrations 1/1000th the MIC Can Select for Antimicrobial Resistance in N. gonorrhoeae—Important Implications for Maximum Residue Limits in Food. Antibiotics 2022, 11, 1430. https://doi.org/10.3390/antibiotics11101430
González N, Abdellati S, De Baetselier I, Laumen JGE, Van Dijck C, Block Td, Manoharan-Basil SS, Kenyon C. Ciprofloxacin Concentrations 1/1000th the MIC Can Select for Antimicrobial Resistance in N. gonorrhoeae—Important Implications for Maximum Residue Limits in Food. Antibiotics. 2022; 11(10):1430. https://doi.org/10.3390/antibiotics11101430
Chicago/Turabian StyleGonzález, Natalia, Saïd Abdellati, Irith De Baetselier, Jolein Gyonne Elise Laumen, Christophe Van Dijck, Tessa de Block, Sheeba Santhini Manoharan-Basil, and Chris Kenyon. 2022. "Ciprofloxacin Concentrations 1/1000th the MIC Can Select for Antimicrobial Resistance in N. gonorrhoeae—Important Implications for Maximum Residue Limits in Food" Antibiotics 11, no. 10: 1430. https://doi.org/10.3390/antibiotics11101430
APA StyleGonzález, N., Abdellati, S., De Baetselier, I., Laumen, J. G. E., Van Dijck, C., Block, T. d., Manoharan-Basil, S. S., & Kenyon, C. (2022). Ciprofloxacin Concentrations 1/1000th the MIC Can Select for Antimicrobial Resistance in N. gonorrhoeae—Important Implications for Maximum Residue Limits in Food. Antibiotics, 11(10), 1430. https://doi.org/10.3390/antibiotics11101430