



Antimicrobial Efficacy against Antibiotic-Tolerant Staphylococcus aureus Depends on the Mechanism of Antibiotic Tolerance


Abstract
:1. Introduction
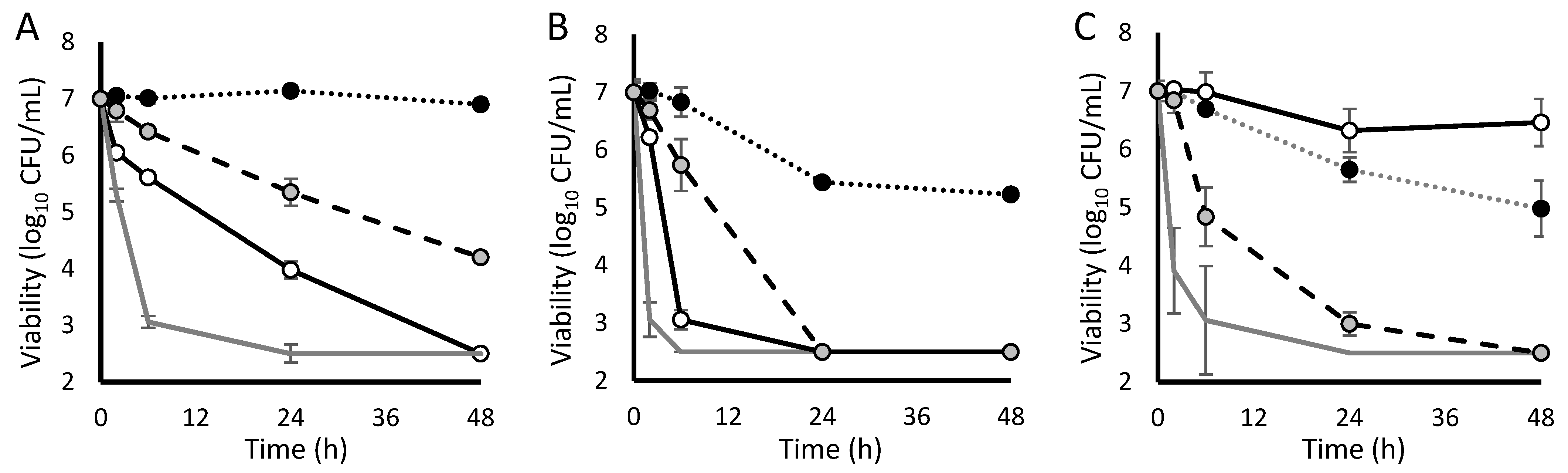
2. Results
3. Discussion
4. Materials and Methods
4.1. Strain Characterization, Cultivation Conditions and Antibiotic Selection
4.2. Nutritional Tolerance
4.3. Competitive Tolerance
4.4. Oxidative Tolerance
4.5. Humoral Tolerance
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kourtis, A.P.; Hatfield, K.; Baggs, J.; Mu, Y.; See, I.; Epson, E.; Nadle, J.; Kainer, M.A.; Dumyati, G.; Petit, S.; et al. Vital Signs: Epidemiology and Recent Trends in Methicillin-Resistant and in Methicillin-Susceptible Staphylococcus Aureus Bloodstream Infections—United States. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 214–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.-D.; Chen, Y.-Y.; Chen, T.-L.; Liu, C.-Y. Risk Factors and Mortality in Patients with Nosocomial Staphylococcus Aureus Bacteremia. Am. J. Infect. Control 2008, 36, 118–122. [Google Scholar] [CrossRef] [PubMed]
- van Hal, S.J.; Jensen, S.O.; Vaska, V.L.; Espedido, B.A.; Paterson, D.L.; Gosbell, I.B. Predictors of Mortality in Staphylococcus Aureus Bacteremia. Clin. Microbiol. Rev. 2012, 25, 362–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing between Resistance, Tolerance and Persistence to Antibiotic Treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef]
- Westblade, L.F.; Errington, J.; Dörr, T. Antibiotic Tolerance. PLoS Pathog. 2020, 16, e1008892. [Google Scholar] [CrossRef]
- Kuehl, R.; Morata, L.; Meylan, S.; Mensa, J.; Soriano, A. When Antibiotics Fail: A Clinical and Microbiological Perspective on Antibiotic Tolerance and Persistence of Staphylococcus Aureus. J. Antimicrob. Chemother. 2020, 75, 1071–1086. [Google Scholar] [CrossRef]
- Wiuff, C.; Zappala, R.M.; Regoes, R.R.; Garner, K.N.; Baquero, F.; Levin, B.R. Phenotypic Tolerance: Antibiotic Enrichment of Noninherited Resistance in Bacterial Populations. Antimicrob. Agents Chemother. 2005, 49, 1483–1494. [Google Scholar] [CrossRef] [Green Version]
- Britt, N.S.; Patel, N.; Shireman, T.I.; El Atrouni, W.I.; Horvat, R.T.; Steed, M.E. Relationship between Vancomycin Tolerance and Clinical Outcomes in Staphylococcus Aureus Bacteraemia. J. Antimicrob. Chemother. 2017, 72, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Bayer, A.S.; Cheung, A.; Lu, L.; Abdelhady, W.; Donegan, N.P.; Hong, J.-I.; Yeaman, M.R.; Xiong, Y.Q. The Stringent Response Contributes to Persistent Methicillin-Resistant Staphylococcus Aureus Endovascular Infection through the Purine Biosynthetic Pathway. J. Infect. Dis. 2020, 222, 1188–1198. [Google Scholar] [CrossRef]
- Bryson, D.; Hettle, A.G.; Boraston, A.B.; Hobbs, J.K. Clinical Mutations That Partially Activate the Stringent Response Confer Multidrug Tolerance in Staphylococcus Aureus. Antimicrob. Agents Chemother. 2020, 64, e02103-19. [Google Scholar] [CrossRef]
- Svenningsen, S.L.; Kongstad, M.; Stenum, T.S.; Muñoz-Gómez, A.J.; Sørensen, M.A. Transfer RNA Is Highly Unstable during Early Amino Acid Starvation in Escherichia coli. Nucleic Acids Res. 2017, 45, 793–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittmar, K.A.; Sørensen, M.A.; Elf, J.; Ehrenberg, M.; Pan, T. Selective Charging of TRNA Isoacceptors Induced by Amino-Acid Starvation. EMBO Rep. 2005, 6, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, T.; Kästle, B.; Gratani, F.L.; Goerke, C.; Wolz, C. Two Small (p)PpGpp Synthases in Staphylococcus Aureus Mediate Tolerance against Cell Envelope Stress Conditions. J. Bacteriol. 2014, 196, 894–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakama, T.; Nureki, O.; Yokoyama, S. Structural Basis for the Recognition of Isoleucyl-Adenylate and an Antibiotic, Mupirocin, by Isoleucyl-TRNA Synthetase. J. Biol. Chem. 2001, 276, 47387–47393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limoli, D.H.; Hoffman, L.R. Help, Hinder, Hide and Harm: What Can We Learn from the Interactions between Pseudomonas Aeruginosa and Staphylococcus Aureus during Respiratory Infections? Thorax 2019, 74, 684–692. [Google Scholar] [CrossRef] [Green Version]
- Filkins, L.M.; Graber, J.A.; Olson, D.G.; Dolben, E.L.; Lynd, L.R.; Bhuju, S.; O’Toole, G.A. Coculture of Staphylococcus Aureus with Pseudomonas Aeruginosa Drives S. Aureus towards Fermentative Metabolism and Reduced Viability in a Cystic Fibrosis Model. J. Bacteriol. 2015, 197, 2252–2264. [Google Scholar] [CrossRef] [Green Version]
- Radlinski, L.C.; Rowe, S.E.; Brzozowski, R.; Wilkinson, A.D.; Huang, R.; Eswara, P.; Conlon, B.P. Chemical Induction of Aminoglycoside Uptake Overcomes Antibiotic Tolerance and Resistance in Staphylococcus Aureus. Cell Chem. Biol. 2019, 26, 1355–1364.e4. [Google Scholar] [CrossRef]
- Radi, R. Peroxynitrite, a Stealthy Biological Oxidant. J. Biol. Chem. 2013, 288, 26464–26472. [Google Scholar] [CrossRef] [Green Version]
- Beam, J.E.; Wagner, N.J.; Shook, J.C.; Bahnson, E.S.M.; Fowler, V.G.; Rowe, S.E.; Conlon, B.P. Macrophage-Produced Peroxynitrite Induces Antibiotic Tolerance and Supersedes Intrinsic Mechanisms of Persister Formation. Infect. Immun. 2021, 89, e0028621. [Google Scholar] [CrossRef]
- Muzamal, U.; Gomez, D.; Kapadia, F.; Golemi-Kotra, D. Diversity of Two-Component Systems: Insights into the Signal Transduction Mechanism by the Staphylococcus Aureus Two-Component System GraSR. F1000Research 2014, 3, 252. [Google Scholar] [CrossRef]
- Falord, M.; Karimova, G.; Hiron, A.; Msadek, T. GraXSR Proteins Interact with the VraFG ABC Transporter to Form a Five-Component System Required for Cationic Antimicrobial Peptide Sensing and Resistance in Staphylococcus Aureus. Antimicrob. Agents Chemother. 2012, 56, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falord, M.; Mäder, U.; Hiron, A.; Débarbouillé, M.; Msadek, T. Investigation of the Staphylococcus Aureus GraSR Regulon Reveals Novel Links to Virulence, Stress Response and Cell Wall Signal Transduction Pathways. PLoS ONE 2011, 6, e21323. [Google Scholar] [CrossRef] [PubMed]
- Ledger, E.V.K.; Mesnage, S.; Edwards, A.M. Human Serum Triggers Antibiotic Tolerance in Staphylococcus Aureus. Nat. Commun. 2022, 13, 2041. [Google Scholar] [CrossRef] [PubMed]
- Friberg, C.; Haaber, J.K.; Vestergaard, M.; Fait, A.; Perrot, V.; Levin, B.R.; Ingmer, H. Human Antimicrobial Peptide, LL-37, Induces Non-Inheritable Reduced Susceptibility to Vancomycin in Staphylococcus Aureus. Sci. Rep. 2020, 10, 13121. [Google Scholar] [CrossRef]
- Horsburgh, M.J.; Aish, J.L.; White, I.J.; Shaw, L.; Lithgow, J.K.; Foster, S.J. SigmaB Modulates Virulence Determinant Expression and Stress Resistance: Characterization of a Functional RsbU Strain Derived from Staphylococcus Aureus 8325-4. J. Bacteriol. 2002, 184, 5457–5467. [Google Scholar] [CrossRef] [Green Version]
- Otero, L.H.; Rojas-Altuve, A.; Llarrull, L.I.; Carrasco-López, C.; Kumarasiri, M.; Lastochkin, E.; Fishovitz, J.; Dawley, M.; Hesek, D.; Lee, M.; et al. How Allosteric Control of Staphylococcus Aureus Penicillin Binding Protein 2a Enables Methicillin Resistance and Physiological Function. Proc. Natl. Acad. Sci. USA 2013, 110, 16808–16813. [Google Scholar] [CrossRef] [Green Version]
- M100-ED29 2019; Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Susceptibility Testing. CLSI Document: Wayne, PA, USA, 2019; Self-Published.
- Reiss, S.; Pané-Farré, J.; Fuchs, S.; François, P.; Liebeke, M.; Schrenzel, J.; Lindequist, U.; Lalk, M.; Wolz, C.; Hecker, M.; et al. Global Analysis of the Staphylococcus Aureus Response to Mupirocin. Antimicrob. Agents Chemother. 2012, 56, 787–804. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Antibiotic | No Induction | Nutritional Tolerance | Competitive Tolerance | Oxidative Tolerance | Humoral Tolerance |
|---|---|---|---|---|---|
| Ceftaroline | 11.1 ± 0.19 | † | 11.1 ± 1.05 | 47.6 ± 0.34 * | † |
| Daptomycin | 1.5 ± 0.25 | † | 4.4 ± 0.09 * | 17.7 ± 0.95 * | 12.9 ± 0.72 * |
| Gentamicin | 1.3 ± 0.17 | 23.4 ± 0.02 * | † | 21.9 ± 1.95 * | † |
| Levofloxacin | 1.7 ± 0.13 | † | 4.9 ± 0.56 * | 31.8 ±3.00 * | † |
| Oritavancin | 1.2 ± 0.08 | 5.3 ± 0.41 * | 1.2 ± 0.02 | 1.4 ± 0.12 | † |
| Vancomycin | 25.8 ± 6.12 | † | † | 45.2 ± 1.39 * | † |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meredith, E.M.; Harven, L.T.; Berti, A.D. Antimicrobial Efficacy against Antibiotic-Tolerant Staphylococcus aureus Depends on the Mechanism of Antibiotic Tolerance. Antibiotics 2022, 11, 1810. https://doi.org/10.3390/antibiotics11121810
Meredith EM, Harven LT, Berti AD. Antimicrobial Efficacy against Antibiotic-Tolerant Staphylococcus aureus Depends on the Mechanism of Antibiotic Tolerance. Antibiotics. 2022; 11(12):1810. https://doi.org/10.3390/antibiotics11121810
Chicago/Turabian StyleMeredith, Emily M., Lauren T. Harven, and Andrew D. Berti. 2022. "Antimicrobial Efficacy against Antibiotic-Tolerant Staphylococcus aureus Depends on the Mechanism of Antibiotic Tolerance" Antibiotics 11, no. 12: 1810. https://doi.org/10.3390/antibiotics11121810
APA StyleMeredith, E. M., Harven, L. T., & Berti, A. D. (2022). Antimicrobial Efficacy against Antibiotic-Tolerant Staphylococcus aureus Depends on the Mechanism of Antibiotic Tolerance. Antibiotics, 11(12), 1810. https://doi.org/10.3390/antibiotics11121810