
Antimicrobial Compounds from Microorganisms

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Bacterial Sources of Antimicrobials

{kind=link}
{kind=link}
| Bacteriocin | Producer of Bacteriocin | Susceptible Microorganisms | Reference(s) |
|---|---|---|---|
| Nisin A | Lactococcus lactic subsp. lactis | E. faecalis ssp. Liquefaciens, Streptococcus equinus, Staphylococcus epidermidis (S. epidermidis), S. aureus, Streptococcus uberis (S. uberis), Streptococcus dysgalactiae (S. dysgalactiae), Streptococcus agalactiae (S. agalactiae), Streptococcus suis (S. suis) Mycobacterium avium subsp. Paratuberculosis | [48,49,50] |
| Nisin ANisin V | L. lactis NZ9700L. lactis NZ9800nisA:M21V | Listeria monocytogenes | [51] |
| Pediocin A | Pediococcus pentosaceus FBB61 | Clostridium perfringens | [52] |
| Enterocin M | Enterococcus faecium AL41 | Campylobacter spp., Clostridium spp. | [53] |
| Enterocin CLE34 | Enterococcus faecium CLE34 | Salmonella pullorum | [26,54] |
| Enterocin E-760 | Enterococcus durans, Enterococcus faecium, Enterococcus hirae | Salmonella enterica serovar Enteritidis, S. enterica serovar Choleraesuis, S. enterica serovar Typhimurium, S. enterica serovar Gallinarum, E. coli O157:H7, Yersinia enterocolitica, S. aureus, Campylobacter jejuni | [55] |
| Lacticin 3147 | Lactococcus lactis DPC3147. | S. dysgalactiae, S. agalactiae, S. aureus, S. uberis, Mycobacterium avium subsp. paratuberculosis | [50,56] |
| Macedocin ST91KM | Streptococcus gallolyticus subsp.macedonicus ST91KM | S. agalactiae, S. dysgalactiae, S. uberis, S. aureus | [57] |
3. Bacterial Sources of Antifungal Compounds
4. Fungal Sources of Antimicrobials
| Microorganism | Compounds | Antimicrobial Activity | Reference(s) |
|---|---|---|---|
| Hormonema spp. | Enfumafungin | Candida spp. and Aspergillus spp. | [92] |
| F. calocera | Favolon | Candida tenuis and Mucor plumbeus | [95] |
| C. comatus | Coprinuslactone | P. aeruginosa | [96] |
| Sanghuangporus spp. | Microporenic acid A | S. aureus and C. albicans | [97] |
| Aspergillus terreus | Rubrolide S | Influenza A virus (H1N1) | [98] |
| Cladosporium sphaerospermum 2005-01-E3 | Cladosin C | Influenza A H1N1 | [99] |
| Penicillium sp. IMB17-046 | Trypilepyrazinol and β-hydroxyergosta-8,14,24 (28)-trien-7-one | HIV and HCV | [101] |
5. Antimicrobial Peptides
6. Antiviral Peptides
7. Other Microbial Sources of Antimicrobial Compounds
| Microbial Sources | Compound(s) | Susceptible Organism(s) | Reference(s) |
|---|---|---|---|
| Synoicum pulmonaria | Synoxazolidinones A and B | MRSA | [126] |
| Serratia proteomaculan | Serraticin A | Escherichia coli and Salmonella enterica | [131] |
| Haliclona simulans | Subtilomycin | Bacillus cereus, Bacillus megaterium, Clostridium sporogenes, Listeria monocytogenes, Listeria innocua, Staphylococcus aureus, MRSA, and Vancomycin-resistant Staphylococcus aureus | [122,125] |
| Ochrolechia spp. | PAMC26625 | Gram-positives: Staphylococcus aureus, Bacillus subtilis, Micrococcus luteus; Gram-negatives, Pseudomonas aeruginosa, Escherichia coli, and Enterobacter cloacae | [133] |
| Tegella cf. spitzbergensis | Eusynstylamides | Pseudomonas aeruginosa, Staphylococcus aureus, Escherichia coli, and Corynebacterium glutamicum | [119,121] |
| Nocardioides spp. | Strain A-1 | Staphylococcus aureus and Xanthomonas oryzae | [136] |
8. Tools and Techniques Used for Antimicrobial Drug Discovery from Microorganisms
- -
- Diffusion methods have several types, including agar disk diffusion, antimicrobial gradient, agar well diffusion, agar plug diffusion, cross streak, and poisoned food methods. The agar disk diffusion method is a routine microbial susceptibility test that was developed in 1940 [139]. It is conducted to test for certain fastidious bacterial pathogens, such as Streptococci, Haemophilus influenza, Neisseria gonorrhea, Nisseria meningitidis, and Haemophilus parainfluenza [140]. In this test, a desired concentration of the test compound is placed on the surface of agar-containing microbes. Antimicrobial agents in the test compound diffuse into the agar and inhibit the proliferation of susceptible microbes. The diameter of inhibition growth zones is then measured [141]. Currently, this method is used to test for non-dermatophyte filamentous fungi using the antifungal disk diffusion approach [142]. Although agar disk diffusion cannot accurately determine the minimal inhibition concentration (MIC), it is simple and less expensive to practice [141].
- -
- The antimicrobial gradient method (Etest) involves a combination of dilution and diffusion methods to determine the MIC value of antibiotics, antifungals, and antimycobacterials. This method can also be used to determine the combined effect of two drugs [141,143]. Other diffusion methods, as mentioned, are agar plug diffusion [144], cross streak [145], and poisoned food [146,147] methods [141].
- -
- The dilution method is suitable for determining MIC values of fastidious or non-fastidious bacteria, yeast, and filamentous fungi [141]. Either broth or agar dilution can be used depending on the test being performed. In testing the action of antifungal drug agents, combinations against Candida sp. Aspergillus, Fusarium, and dermatophytes, agar dilutions are mostly used [148,149,150].
- -
- The time-kill test [151], ATP bioluminescence assay [152,153,154,155], and flow-cytofluorometric method [156] are all techniques used to screen and determine the susceptibility of microbes to antimicrobial compounds [141]. ATP bioluminescence has been used to estimate the amount of ATP present in different cell types [152]. The luciferin–luciferase bioluminescent assay method is mostly preferred due to its high sensitivity [152]. In this method, MgATP2+ changes luciferin into a state that can be catalytically oxidized by the luciferase in high quantum yield chemiluminescent reaction [152]. There is a relationship between light intensity and ATP concentration under the right conditions [152]. Cellular ATP can be measured when free ATP released from broken down cell is made to react with the luciferin–luciferase resulting in light emission [152]. The amount of light emitted is measured by a luminometer [141].
- -
- The time-kill test on the other hand, is suitable for evaluating bactericidal and fungicidal activity [141]. It provides information about the relationship between the antimicrobial agent and the microbial strain depending on the time taken for the action to occur and the concentration of the antimicrobial agent [141].
- -
- The flow cytofluorometric method exposes antimicrobial resistance and predicts the effect of the tested molecule on cell damage and viability of the tested microbe [141] using a flow cytometer [157]. In performing this procedure, the cells damaged by antimicrobial agents are dyed with an appropriate stain [141]. A known DNA stain is propidium iodide (PI) [141]. The quantity of damaged cells can be used to determine the antimicrobial activity of the test compound [141].
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- O’Neill, J. Antimicrobial Resistance:Tackling a crisis for the health and wealth of nations. Rev. Antimicrob. Resist. 2014. [Google Scholar]
- Laws, M.; Shaaban, A.; Rahman, K.M. Antibiotic resistance breakers: Current approaches and future directions. FEMS Microbiol. Rev. 2019, 43, 490–516. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Elissawy, A.M.; Dehkordi, E.S.; Mehdinezhad, N.; Ashour, M.L.; Pour, P.M. Cytotoxic Alkaloids Derived from Marine Sponges: A Comprehensive Review. Biomolecules 2021, 11, 258. [Google Scholar] [CrossRef]
- El-Demerdash, A.; Tammam, M.A.; Atanasov, A.G.; Hooper, J.N.A.; Al-Mourabit, A.; Kijjoa, A. Chemistry and Biological Activities of the Marine Sponges of the Genera Mycale (Arenochalina), Biemna and Clathria. Mar. Drugs 2018, 16, 214. [Google Scholar] [CrossRef] [Green Version]
- Karpinski, T.M. Marine Macrolides with Antibacterial and/or Antifungal Activity. Mar. Drugs 2019, 17, 241. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Meng, W.; Cao, C.; Wang, J.; Shan, W.; Wang, Q. Antibacterial and antifungal compounds from marine fungi. Mar. Drugs 2015, 13, 3479–3513. [Google Scholar] [CrossRef]
- Swift, C.L.; Louie, K.B.; Bowen, B.P.; Olson, H.M.; Purvine, S.O.; Salamov, A.; Mondo, S.J.; Solomon, K.V.; Wright, A.T.; Northen, T.R.; et al. Anaerobic gut fungi are an untapped reservoir of natural products. Proc. Natl. Acad. Sci. USA 2021, 118, e2019855118. [Google Scholar] [CrossRef]
- Maghembe, R.; Damian, D.; Makaranga, A.; Nyandoro, S.S.; Lyantagaye, S.L.; Kusari, S.; Hatti-Kaul, R. Omics for Bioprospecting and Drug Discovery from Bacteria and Microalgae. Antibiot 2020, 9, 229. [Google Scholar] [CrossRef]
- Moir, D.T.; Shaw, K.J.; Hare, R.S.; Vovis, G.F. Genomics and Antimicrobial Drug Discovery. Antimicrob. Agents Chemother. 1999, 43, 439. [Google Scholar] [CrossRef] [Green Version]
- Schnappinger, D. Genetic Approaches to Facilitate Antibacterial Drug Development. Cold Spring Harb. Perspect. Med. 2015, 5, a021139. [Google Scholar] [CrossRef] [Green Version]
- Tortorella, E.; Tedesco, P.; Espositom, F.P.; January, G.G.; Fani, R.; Jaspars, M.; de Pascale, D. Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives. Mar. Drugs 2018, 16, 355. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An emerging source for drug discovery. J. Antibiot. 2011, 64, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Rojas, V.; Rivas, L.; Cardenas, C.; Guzman, F. Cyanobacteria and Eukaryotic Microalgae as Emerging Sources of Antibacterial Peptides. Molecules 2020, 25, 5804. [Google Scholar] [CrossRef]
- Alsenani, F.; Tupally, K.R.; Chua, E.T.; Eltanahy, E.; Alsufyani, H.; Parekh, H.S.; Schenk, P.M. Evaluation of microalgae and cyanobacteria as potential sources of antimicrobial compounds. Saudi. Pharm. J. 2020, 28, 1834–1841. [Google Scholar] [CrossRef]
- Santovito, E.; Greco, D.; Marquis, V.; Raspoet, R.; D’Ascanio, V.; Logrieco, A.F.; Avantaggiato, G. Antimicrobial Activity of Yeast Cell Wall Products Against Clostridium perfringens. Foodborne Pathog. Dis. 2019, 16, 638–647. [Google Scholar] [CrossRef]
- Hatoum, R.; Labrie, S.; Fliss, I. Antimicrobial and probiotic properties of yeasts: From fundamental to novel applications. Front. Microbiol. 2012, 3, 421. [Google Scholar] [CrossRef] [Green Version]
- Mitcheltree, M.J.; Pisipati, A.; Syroegin, E.A.; Silvestre, K.J.; Klepacki, D.; Mason, J.D.; Terwilliger, D.W.; Testolin, G.; Pote, A.R.; Wu, K.J.Y.; et al. A synthetic antibiotic class overcoming bacterial multidrug resistance. Nature 2021, 599, 507–512. [Google Scholar] [CrossRef]
- Tenea, G.N.; Yépez, L. Bioactive Compounds of Lactic Acid Bacteria. Case Study: Evaluation of Antimicrobial Activity of Bacteriocin-producing Lactobacilli Isolated from Native Ecological Niches of Ecuador. Probiotics Prebiotics Hum. Nutr. Health 2016, xxx, 149–167. [Google Scholar]
- Todorov, S.D.; Dicks, L.M.T. Bacteriocin production by Lactobacillus pentosus ST712BZ isolated from boza. Braz. J. Microbiol. 2007, 38, 166–172. [Google Scholar] [CrossRef]
- Oscáriz, J.C.; Pisabarro, A.G. Classification and mode of action of membrane-active bacteriocins produced by Gram-positive bacteria. Int. Microbiol. 2001, 4, 13–19. [Google Scholar] [CrossRef]
- Klaenhammer, T.R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 39–85. [Google Scholar] [CrossRef]
- Güllüce, M.; Karadayı, M.; Barış, Ö. Bacteriocins: Promising natural antimicrobials. Local Environ. 2013, 3, 6. [Google Scholar]
- Cotter, P.D.; Hill, C.; Ross, R.P. Food microbiology: Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777–788. [Google Scholar] [CrossRef]
- Drider, D.; Fimland, G.; Héchard, Y.; McMullen, L.M.; Prévost, H. The Continuing Story of Class IIa Bacteriocins. Microbiol. Mol. Biol. Rev. 2006, 70, 564–582. [Google Scholar] [CrossRef] [Green Version]
- Hernández-González, J.C.; Martínez-Tapia, A.; Lazcano-Hernández, G.; García-Pérez, B.E.; Castrejón-Jiménez, N.S. Bacteriocins from lactic acid bacteria. A powerful alternative as antimicrobials, probiotics, and immunomodulators in veterinary medicine. Animals 2021, 11, 979. [Google Scholar] [CrossRef]
- Parada, J.L.; Caron, C.R.; Medeiros, A.B.P.; Soccol, C.R. Bacteriocins from lactic acid bacteria: Purification, properties and use as biopreservatives. Braz. Arch. Biol. Technol. 2007, 50, 521–542. [Google Scholar] [CrossRef] [Green Version]
- Bauer, R.; Dicks, L.M.T. Mode of action of lipid II-targeting lantibiotics. Int. J. Food Microbiol. 2005, 101, 201–216. [Google Scholar] [CrossRef]
- Paiva, A.D.; Breukink, E.; Mantovani, H.C. Role of lipid II and membrane thickness in the mechanism of action of the lantibiotic bovicin HC5. Antimicrob. Agents Chemother. 2011, 55, 5284–5293. [Google Scholar] [CrossRef] [Green Version]
- Nissen-Meyer, J.; Oppegård, C.; Rogne, P.; Haugen, H.S.; Kristiansen, P.E. Structure and Mode-of-Action of the Two-Peptide (Class-IIb) Bacteriocins. Probiotics Antimicrob. Proteins 2010, 2, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Circular and Leaderless Bacteriocins: Biosynthesis, Mode of Action, Applications, and Prospects. Front Microbiol. 2018, 9, 2085. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, R.S.; Pearson, L.; Kennedy, R.C. Mode of Action of a Lysostaphin-Like Bacteriolytic Agent Produced by Streptococcus zooepidemicus 4881. Appl. Environ. Microbiol. 1996, 62, 4536–4541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Wang, X.; Zhang, X.; Wu, H.; Zou, Y.; Li, P.; Sun, C.; Xu, W. Class III bacteriocin Helveticin-M causes sublethal damage on target cells through impairment of cell wall and membrane. J. Ind. Microbiol. Biotechnol. 2018, 45, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Swe, P.M.; Cook, G.M.; Tagg, J.R.; Jack, R.W. Mode of Action of Dysgalacticin: A Large Heat-Labile Bacteriocin. J. Antimicrob. Chemother. 2009, 63, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Meade, E.; Slattery, M.A.; Garvey, M. Bacteriocins, Potent Antimicrobial Peptides and the Fight against Multi Drug Resistant Species: Resistance Is Futile? Antibiotics 2020, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Mclller, E.; Radler, F. Caseicin, a Bacteriocin from Lactobacillus casei. Folia Microbiol. 1993, 38, 441–446. [Google Scholar] [CrossRef]
- Bollenbach, T. Antimicrobial interactions: Mechanisms and implications for drug discovery and resistance evolution. Curr. Opin. Microbiol. 2015, 27, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Cunha, E.; Trovão, T.; Pinheiro, A.; Nunes, T.; Santos, R.; Moreira, J.; Braz, B.S.; Tavares, L.; Veiga, A.S.; Oliveira, M. Potential of Two Delivery Systems for Nisin Topical Application to Dental Plaque Biofilms in Dogs. BMC Vet. Res. 2018, 14, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Howeir, T.H.; Fioreliinp, J.P. The Effect of a Mouthrinse Based on Nisin, a Bacteriocin, on Developing Plaque and Gingivitis in Beagle Dogs. J. Clin. Periodontol. 1993, 20, 335–339. [Google Scholar] [CrossRef]
- Herzog, M.; Tiso, T.; Blank, L.M.; Winter, R. Interaction of rhamnolipids with model biomembranes of varying complexity. Biochim. Biophys. Acta-Biomembr. 2020, 1862, 183431. [Google Scholar] [CrossRef]
- Jiang, J.; Zu, Y.; Li, X.; Meng, Q.; Long, X. Recent progress towards industrial rhamnolipids fermentation: Process optimization and foam control. Bioresour. Technol. 2020, 298, 122394. [Google Scholar] [CrossRef] [PubMed]
- Thakur, P.; Saini, N.K.; Thakur, V.K.; Gupta, V.K.; Saini, R.V.; Saini, A.K. Rhamnolipid the Glycolipid Biosurfactant: Emerging trends and promising strategies in the field of biotechnology and biomedicine. Microb. Cell Factories 2021, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Christova, N.; Tuleva, B.; Kril, A.; Georgieva, M.; Konstantinov, S.; Terziyski, I.; Nikolova, B.; Stoineva, I. Chemical Structure and In Vitro Antitumor Activity of Rhamnolipids from Pseudomonas aeruginosa BN10. Appl. Biochem. Biotechnol. 2013, 170, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Borah, S.N.; Goswami, D.; Sarma, H.K.; Cameotra, S.S.; Deka, S. Rhamnolipid biosurfactant against Fusarium verticillioides to control stalk and ear rot disease of maize. Front. Microbiol. 2016, 7, 1505. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Black, W.; Sawyer, T. Application of Environment-Friendly Rhamnolipids against Transmission of Enveloped Viruses Like SARS-CoV2. Viruses 2021, 13, 322. [Google Scholar] [CrossRef]
- Remichkova, M.; Galabova, D.; Roeva, I.; Karpenko, E.; Shulga, A.; Galabov, A.S. Anti-herpesvirus activities of Pseudomonas sp. S-17 rhamnolipid and its complex with alginate. Zeitschrift Fur Naturforsch Sect. C J. Biosci. 2008, 63, 75–81. [Google Scholar] [CrossRef]
- Giugliano, R.; Buonocore, C.; Zannella, C.; Chianese, A.; Esposito, F.P.; Tedesco, P.; De Filippis, A.; Galdiero, M.; Franci, G.; de Pascale, D. Antiviral Activity of the Rhamnolipids Mixture from the Antarctic Bacterium Pseudomonas gessardii M15 against Herpes Simplex Viruses and Coronaviruses. Pharmaceutics 2021, 13, 2121. [Google Scholar] [CrossRef]
- Broadbent, J.R.; Chou, Y.C.; Gillies, K.; Kondo, J.K. Nisin Inhibits Several Gram-Positive, Mastitis-Causing Pathogens. J. Dairy Sci. 1989, 72, 3342–3345. [Google Scholar] [CrossRef]
- Lebel, G.; Piché, F.; Frenette, M.; Gottschalk, M.; Grenier, D. Antimicrobial activity of nisin against the swine pathogen Streptococcus suis and its synergistic interaction with antibiotics. Peptides 2013, 50, 19–23. [Google Scholar] [CrossRef]
- Carroll, J.; Draper, L.A.; O’Connor, P.M.; Coffey, A.; Hill, C.; Ross, R.P.; Cotter, P.D.; O’Mahony, J. Comparison of the activities of the lantibiotics nisin and lacticin 3147 against clinically significant mycobacteria. Int. J. Antimicrob. Agents 2010, 36, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Campion, A.; Casey, P.G.; Field, D.; Cotter, P.D.; Hill, C.; Ross, R.P. In vivo activity of Nisin A and Nisin v against Listeria monocytogenes in mice. BMC Microbiol. 2013, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Grilli, E.; Messina, M.; Catelli, E.; Morlacchini, M.; Piva, A. Pediocin a improves growth performance of broilers challenged with Clostridium perfringens. Poult. Sci. 2009, 88, 2152–2158. [Google Scholar] [CrossRef] [PubMed]
- Lauková, A.; Styková, E.; Kubašová, I.; Gancarčíková, S.; Plachá, I.; Mudroňová, D.; Kandričáková, A.; Miltko, R.; Belzecki, G.; Valocký, I. Enterocin M and its Beneficial Effects in Horses—A Pilot Experiment. Probiotics Antimicrob. Proteins 2018, 10, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cui, Y.; Wang, W.; Xu, J.; Xu, L. Production of two bacteriocins in various growth conditions produced by gram-positive bacteria isolated from chicken cecum. Can. J. Microbiol. 2012, 58, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Line, J.E.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, V.; Mitsevich, I.P.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S.; Siragusa, G.R.; et al. Isolation and Purification of Enterocin E-760 with Broad Antimicrobial Activity against Gram-Positive and Gram-Negative Bacteria. Antimicrob. Agents Chemother. 2008, 52, 1094–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, M.P.; Flynn, J.; Hill, C.; Ross, R.P.; Meaney, W.J. The natural food grade inhibitor, lacticin 3147, reduced the incidence of mastitis after experimental challenge with Streptococcus dysgalactiae in nonlactating dairy cows. J. Dairy Sci. 1999, 82, 2625–2631. [Google Scholar] [CrossRef]
- Pieterse, R.; Todorov, S.D.; Dicks, L.M.T. Mode of action and in vitro susceptibility of mastitis pathogens to macedocin ST91KM and preparation of a teat seal containing the bacteriocin. Braz. J. Microbiol. 2010, 41, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Tomita, K.; Nishio, M.; Saitoh, K.; Yamamoto, H.; Hoshino, Y.; Ohkuma, H.; Konishi, M.; Miyaki, T.; Oki, T. Pradimicins A, B and C: New antifungal antibiotics. I. Taxonomy, production, isolation and physico-chemical properties. J. Antibiot. 1990, 43, 755–762. [Google Scholar] [CrossRef] [Green Version]
- Walsh, T.J.; Giri, N. Pradimicins: A novel class of broad-spectrum antifungal compounds. Eur. J. Clin. Microbiol. Infect. Dis. 1997, 16, 93–97. [Google Scholar] [CrossRef]
- Kerr, J.R. Bacterial inhibition of fungal growth and pathogenicity. Microb. Ecol. Health Dis. 1999, 11, 129–142. [Google Scholar]
- Boumehira, A.Z.; El-Enshasy, H.A.; Hacene, H.; Elsayed, E.A.; Aziz, R.; Park, E.Y. Recent progress on the development of antibiotics from the genus Micromonospora. Biotechnol. Bioprocess Eng. 2016, 21, 199–223. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, R.E.; Giacobbe, R.A.; Monaghan, R.L. L-671,329, a new antifungal agent. I. Fermentation and isolation. J. Antibiot. 1989, 42, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, N.; Kondo, Y.; Koyama, M.; Omoto, S.; Iwata, M.; Tsuruoka, T.; Inouye, S. Studies on a new nucleotide antibiotic, Dapiramicin. J. Antibiot. 1983, 37, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fushimi, S.; Furihata, K.; Seto, H. Studies of new phosphate ester antifungal antibiotics phoslactomycins. II. Structure elucidation of phoslactomycins A to F. J. Antibiot. 1989, 42, 1026–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanafi, M.; Shibata, K.; Ueki, M.; Taniguchi, M. UK-2A, B, C and D, novel antifungal antibiotics from Streptomyces sp. 517-02: II. Structural elucidation. J. Antibiot. 1996, 49, 1226–1231. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Kanaya, I.; Takahashi, Y.; Shinose, M.; Tanaka, H.; Omura, S.; Phthoxazolin, A. Phthoxazolin A, a specific inhibitor of cellulose biosynthesis from microbial origin. I. Discovery, taxonomy of producing microorganism, fermentation, and biological activity. J. Antibiot. 1993, 46, 1208–1213. [Google Scholar] [CrossRef]
- Mitsuhashi, S.; Inoue, K. In vitro antibacterial activity of azithromycin, a new macrolide antibiotic. Jpn. J. Chemother. 1995, 43, 1–7. [Google Scholar]
- Kotake, C.; Yamasaki, T.; Moriyama, T.; Shinoda, M.; Komiyama, N.; Furumai, T.; Konishi, M.; Oki, T. Butyrolactols a and bΔ, new antifungal antibiotics taxonomy, isolation, physico-chemical properties, structure and biological activity. J. Antibiot. 1992, 45, 1442–1450. [Google Scholar] [CrossRef] [Green Version]
- Ohkuma, H.; Naruse, N.; Nishiyama, Y.; Tsuno, T.; Hoshino, Y.; Sawada, Y.; Konishi, M. Streptomyces roseiscleroticus Taxonomyof the Producing Organism. J. Antibiot. 1992, 45, 1239–1249. [Google Scholar] [CrossRef]
- Uramoto, M.; Uzawa, J.; Suzuki, S.; Isono, K.; Liehr, J.G.; McCloskey, J.A. Isolation and structure of polyoxin N. Nucleic Acids Res. 1978, 1, s327–s332. [Google Scholar] [CrossRef]
- Hochlowski, J.E.; Mullally, M.M.; Brill, G.M.; Whittern, D.N.; Buko, A.M.; Hill, P.; McAlpine, J.B. Dunaimycins, a new complex of spiroketal 24-membered macrolides with immunosuppressive activity: II. isolation and elucidation of structures. J. Antibiot. 1991, 44, 1318–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunlap, C.A.; Bowman, M.J.; Rooney, A.P. Iturinic lipopeptide diversity in the bacillus subtilis species group-important antifungals for plant disease biocontrol applications. Front. Microbiol. 2019, 10, 1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebbadi, M.; Gálvez, A.; Maqueda, M.; Martínez-Bueno, M.; Valdivia, E. Fungicin M4: A narrow spectrum peptide antibiotic from Bacillus licheniformis M-4. J. Appl. Bacteriol. 1994, 77, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, A.; Maqueda, M.; Martínez-Bueno, M.; Lebbadi, M.; Valdivia, E. Isolation and physico-chemical characterization of an antifungal and antibacterial peptide produced by Bacillus licheniformis A12. Appl. Microbiol. Biotechnol. 1993, 39, 438–442. [Google Scholar] [CrossRef]
- Chernin, L.; Brandis, A.; Ismailov, Z.; Chet, I. Pyrrolnitrin production by an Enterobacter agglomerans strain with a broad spectrum of antagonistic activity towards fungal and bacterial phytopathogens. Curr. Microbiol. 1996, 32, 208–212. [Google Scholar] [CrossRef]
- Greiner, M.; Winkelmann, G. Fermentation and isolation of herbicolin A, a peptide antibiotic produced by Erwinia herbicola strain A 111. Appl. Microbiol. Biotechnol. 1991, 34, 565–569. [Google Scholar] [CrossRef]
- Winkelmann, G.; Lupp, R.; Jung, G. Herbicolins—New peptide antibiotics from erwinia herbicola. J. Antibiot. 1980, 33, 353–358. [Google Scholar] [CrossRef]
- Shoji, J.; Hinoo, H.; Sakazaki, R.; Kato, T.; Hattori, T.; Matsumoto, K.; Tawara, K.; Kikuchi, J.; Terui, Y. Isolation of CB-25-I, an antifungal antibiotic, from serratia plymuthica. J. Antibiot. 1989, 42, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Serino, L.; Reimmann, C.; Visca, P.; Beyeler, M.; Chiesa, V.D.; Haas, D. Biosynthesis of pyochelin and dihydroaeruginoic acid requires the iron-regulated pchDCBA operon in Pseudomonas aeruginosa. J. Bacteriol. 1997, 179, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Kerr, J.R.; Taylor, G.W.; Rutman, A.; Høiby, N.; Cole, P.J.; Wilson, R. Pseudomonas aeruginosa pyocyanin and 1-hydroxyphenazine inhibit fungal growth. J. Clin. Pathol. 1999, 52, 385–387. [Google Scholar] [CrossRef] [Green Version]
- Vincent, M.N.; Harrison, L.A.; Brackin, J.M.; Kovacevich, P.A.; Mukerji, P.; Weller, D.M.; Pierson, E.A. Genetic analysis of the antifungal activity of a soilborne Pseudomonas aureofaciens strain. Appl. Environ. Microbiol. 1991, 57, 2928–2934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, L.; Teplow, D.B.; Rinaldi, M.; Strobel, G. Pseudomycins, a family of novel peptides from Pseudomonas syringae possessing broad-spectrum antifungal activity. J. Gen. Microbiol. 1991, 137, 2857–2865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, M.; Park, H.J.; Ishizuka, S.; Omata, K.; Hirama, M. Chemistry and Antimicrobial Activity of Caryoynencins Analogs. J. Med. Chem. 1995, 38, 5015–5022. [Google Scholar] [CrossRef] [PubMed]
- Barker, W.R.; Callaghan, C.; Hill, L.; Noble, D.; Acred, P.; Harper, P.B.; Sowa, M.A.; Fletton, R.A. G1549, a new cyclic hydroxamic acid antibiotic, isolated from culture broth of pseudomonas alcaligenes. J. Antibiot. 1979, 32, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.; Suh, J.W.; Kim, S.; Hyun, B.; Kim, C.; Lee, C. hoon Cepacidine A, A novel antifungal antibiotic produced by pseudomonas cepacia. II. Physico-chemical properties and structure elucidation. J. Antibiot. 1994, 47, 1406–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, M.; Nakazawa, T. Characterization of Hemolytic and Antifungal Substance, Cepalycin, from Pseudomonas cepacia. Microbiol. Immunol. 1994, 38, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bisacchi, G.S.; Parker, W.L.; Hockstein, D.R.; Koster, W.H.; Rathnum, M.L.; Unger, S.E. Xylocandin: A new complex of antifungal peptides. II. Structural studies and chemical modifications. J. Antibiot. 1987, 40, 1520–1529. [Google Scholar] [CrossRef] [Green Version]
- Saalim, M.; Villegas-Moreno, J.; Clark, B.R. Bacterial Alkyl-4-quinolones: Discovery, Structural Diversity and Biological Properties. Molecules 2020, 25, 5689. [Google Scholar] [CrossRef]
- Knappe, T.A.; Linne, U.; Zirah, S.; Rebuffat, S.; Xie, X.; Marahiel, M.A. Isolation and structural characterization of capistruin, a lasso peptide predicted from the genome sequence of Burkholderia thailandensis E264. J. Am. Chem. Soc. 2008, 130, 11446–11454. [Google Scholar] [CrossRef]
- Watanabe, T.; Izaki, K.; Takahashi, H. New polyenic antibiotics active against gram-positive and -negative bacteria: I. Isolation and purification of antibiotics produced by gl uconobacter sp. W-315. J. Antibiot. 1982, 35, 1141–1147. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; Deshmukh, S.K.; Reddy, M.S.; Prasad, R.; Goel, M. Endolichenic fungi: A hidden source of bioactive metabolites. S. Afr. J. Bot. 2020, 134, 163–186. [Google Scholar] [CrossRef]
- Pelaez, F.; Cabello, A.; Platas, G.; Díez, M.T.; Del Val, A.G.; Basilio, A.; Martán, I.; Vicente, F.; Bills, G.F.; Giacobbe, R.A.; et al. The discovery of enfumafungin, a novel antifungal compound produced by an endophytic Hormonema species biological activity and taxonomy of the producing organisms. Syst. Appl. Microbiol. 2000, 23, 333–343. [Google Scholar] [CrossRef]
- Kuhnert, E.; Li, Y.; Lan, N.; Yue, Q.; Chen, L.; Cox, R.J.; An, Z.; Yokoyama, K.; Bills, G.F. Enfumafungin synthase represents a novel lineage of fungal triterpene cyclases. Environ. Microbiol. 2018, 20, 3325–3342. [Google Scholar] [CrossRef] [PubMed]
- Sauter, H.; Steglich, W.; Anke, T. Strobilurins: Evolution of a new class of active substances. Angew. Chem.-Int. Ed. 1999, 38, 1328–1349. [Google Scholar] [CrossRef]
- Chepkirui, C.; Richter, C.; Matasyoh, J.C.; Stadler, M. Monochlorinated calocerins A–D and 9-oxostrobilurin derivatives from the basidiomycete Favolaschia calocera. Phytochemistry 2016, 132, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Chepkirui, C.; Cheng, T.; Matasyoh, J.; Decock, C.; Stadler, M. An unprecedented spiro [furan-2,1’-indene]-3-one derivative and other nematicidal and antimicrobial metabolites from Sanghuangporus sp. (Hymenochaetaceae, Basidiomycota) collected in Kenya. Phytochem. Lett. 2018, 25, 141–146. [Google Scholar] [CrossRef]
- Zhu, T.; Chen, Z.; Liu, P.; Wang, Y.; Xin, Z.; Zhu, W. New rubrolides from the marine-derived fungus Aspergillus terreus OUCMDZ-1925. J. Antibiot. 2014, 67, 315–318. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Sun, X.; Yu, G.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. Cladosins A–E, hybrid polyketides from a deep-sea-derived fungus, Cladosporium sphaerospermum. J. Nat. Prod. 2014, 77, 270–275. [Google Scholar] [CrossRef]
- Peng, J.; Zhang, X.; Du, L.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. Sorbicatechols A and B, antiviral sorbicillinoids from the marine-derived fungus Penicillium chrysogenum PJX-17. J. Nat. Prod. 2014, 77, 424–428. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Hao, X.; Li, S.; Jia, J.; Guan, Y.; Peng, Z.; Bi, H.; Xiao, C.; Cen, S.; et al. Broad-Spectrum Antiviral Natural Products from the Marine-Derived Penicillium sp. IMB17-046. Molecules 2019, 24, 2821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Fang, W.; Tan, S.; Lin, X.; Xun, T.; Yang, B.; Liu, S.; Liu, Y. Aspernigrins with anti-HIV-1 activities from the marine-derived fungus Aspergillus niger SCSIO Jcsw6F30. Bioorg. Med. Chem. Lett. 2016, 26, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Raveh, A.; Delekta, P.C.; Dobry, C.J.; Peng, W.; Schultz, P.J.; Blakely, P.K.; Tai, A.W.; Matainaho, T.; Irani, D.N.; Sherman, D.H.; et al. Discovery of potent broad spectrum antivirals derived from marine actinobacteria. PLoS ONE 2013, 8, e82318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Dou, X.; Song, J.; Lyu, Y.; Zhu, X.; Xu, L.; Li, W.; Shan, A. Antimicrobial peptides: Promising alternatives in the post feeding antibiotic era. Med. Res. Rev. 2019, 39, 831–859. [Google Scholar] [CrossRef]
- Narayana, J.L.; Chen, J.Y. Antimicrobial peptides: Possible anti-infective agents. Peptides 2015, 72, 88–94. [Google Scholar] [CrossRef]
- Kang, X.; Dong, F.; Shi, C.; Liu, S.; Sun, J.; Chen, J.; Li, H.; Xu, H.; Lao, X.; Zheng, H. DRAMP 2.0, an updated data repository of antimicrobial peptides. Sci. Data 2019, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- da Cunha, N.B.; Cobacho, N.B.; Viana, J.F.C.; Lima, L.A.; Sampaio, K.B.O.; Dohms, S.S.M.; Ferreira, A.C.R.; de la Fuente-Núñez, C.; Costa, F.F.; Franco, O.L.; et al. The next generation of antimicrobial peptides (AMPs) as molecular therapeutic tools for the treatment of diseases with social and economic impacts. Drug Discov. Today 2017, 22, 234–248. [Google Scholar] [CrossRef]
- Huerta-Cantillo, J.; Navarro-García, F. Properties and design of antimicrobial peptides as potential tools againstpathogens and malignant cells. Investig. En Discapac. 2016, 5, 96–115. [Google Scholar]
- Li, C.; Zhu, C.; Ren, B.; Yin, X.; Shim, S.H.; Gao, Y.; Zhu, J.; Zhao, P.; Liu, C.; Yu, R.; et al. Two optimized antimicrobial peptides with therapeutic potential for clinical antibiotic-resistant Staphylococcus aureus. Eur. J. Med. Chem. 2019, 183, 111686. [Google Scholar] [CrossRef]
- Kosikowska, P.; Lesner, A. Antimicrobial peptides (AMPs) as drug candidates: A patent review (2003–2015). Expert Opin. Ther. Pat. 2016, 26, 689–702. [Google Scholar] [CrossRef]
- Boas, L.C.P.V.; Campos,, M.L.; Berlanda, R.L.A.; de Carvalho Neves, N.; Franco, O.L. Antiviral peptides as promising therapeutic drugs. Cell. Mol. Life Sci. 2019, 76, 3525–3542. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582–779. [Google Scholar] [CrossRef] [PubMed]
- Yasin, B.; Pang, M.; Turner, J.S.; Cho, Y.; Dinh, N.N.; Waring, A.J.; Lehrer, R.I.; Wagar, E.A. Evaluation of the inactivation of infectious herpes simplex virus by host-defense peptides. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Carriel-Gomes, M.C.; Kratz, J.M.; Barracco, M.A.; Bachére, E.; Barardi, C.R.M.; Simões, C.M.O. In vitro antiviral activity of antimicrobial peptides against herpes simplex virus 1, adenovirus, and rotavirus. Mem. Inst. Oswaldo Cruz 2007, 102, 469–472. [Google Scholar] [CrossRef]
- Kobori, T.; Iwamoto, S.; Takeyasu, K.; Ohtani, T. Biopolymers Volume 85/Number 4295. Biopolymers 2007, 85, 392–406. [Google Scholar]
- Xia, S.; Liu, M.; Wang, C.; Xu, W.; Lan, Q.; Feng, S.; Qi, F.; Bao, L.; Du, L.; Liu, S.; et al. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res. 2020, 30, 343–355. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Song, F.; Xiao, X.; Huang, P.; Li, L.; Monte, A.; Abdel-Mageed, W.M.; Wang, J.; Guo, H.; He, W.; et al. Abyssomicins from the South China Sea deep-sea sediment Verrucosispora sp.: Natural thioether Michael addition adducts as antitubercular prodrugs. Angew. Chem. Int. Ed. Engl. 2013, 52, 1231–1234. [Google Scholar] [CrossRef] [Green Version]
- Tadesse, M.; Tabudravu, J.N.; Jaspars, M.; Strom, M.B.; Hansen, E.; Andersen, J.H.; Kristiansen, P.E.; Haug, T. The antibacterial ent-eusynstyelamide B and eusynstyelamides D, E, and F from the Arctic bryozoan Tegella cf. spitzbergensis. J. Nat. Prod. 2011, 74, 837–841. [Google Scholar] [CrossRef]
- Kennedy, J.; Codling, C.E.; Jones, B.V.; Dobson, A.D.; Marchesi, J.R. Diversity of microbes associated with the marine sponge, Haliclona simulans, isolated from Irish waters and identification of polyketide synthase genes from the sponge metagenome. Environ. Microbiol. 2008, 10, 1888–1902. [Google Scholar] [CrossRef]
- Tapiolas, D.M.; Bowden, B.F.; Abou-Mansour, E.; Willis, R.H.; Doyle, J.R.; Muirhead, A.N.; Liptrot, C.; Llewellyn, L.E.; Wolff, C.W.; Wright, A.D.; et al. Eusynstyelamides A, B, and C, nNOS inhibitors, from the ascidian Eusynstyela latericius. J. Nat. Prod. 2009, 72, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- McAuliffe, O.; Ross, R.P.; Hill, C. Lantibiotics: Structure, biosynthesis and mode of action. FEMS Microbiol. Rev. 2001, 25, 285–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freire, J.M.; Dias, S.A.; Flores, L.; Veiga, A.S.; Castanho, M.A. Mining viral proteins for antimicrobial and cell-penetrating drug delivery peptides. Bioinformatics 2015, 31, 2252–2256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukunaga, Y.; Kurahashi, M.; Tanaka, K.; Yanagi, K.; Yokota, A.; Harayama, S. Pseudovibrio ascidiaceicola sp. nov., isolated from ascidians (sea squirts). Int. J. Syst. Evol. Microbiol. 2006, 56, 343–347. [Google Scholar] [CrossRef]
- Phelan, R.W.; Barret, M.; Cotter, P.D.; O’Connor, P.M.; Chen, R.; Morrissey, J.P.; Dobson, A.D.; O’Gara, F.; Barbosa, T.M. Subtilomycin: A new lantibiotic from Bacillus subtilis strain MMA7 isolated from the marine sponge Haliclona simulans. Mar. Drugs 2013, 11, 1878–1898. [Google Scholar] [CrossRef] [Green Version]
- Tadesse, M.; Strom, M.B.; Svenson, J.; Jaspars, M.; Milne, B.F.; Torfoss, V.; Andersen, J.H.; Hansen, E.; Stensvag, K.; Haug, T. Synoxazolidinones A and B: Novel bioactive alkaloids from the ascidian Synoicum pulmonaria. Org. Lett. 2010, 12, 4752–4755. [Google Scholar] [CrossRef]
- Blond, A.; Cheminant, M.; Destoumieux-Garzon, D.; Segalas-Milazzo, I.; Peduzzi, J.; Goulard, C.; Rebuffat, S. Thermolysin-linearized microcin J25 retains the structured core of the native macrocyclic peptide and displays antimicrobial activity. Eur. J. Biochem. 2002, 269, 6212–6222. [Google Scholar] [CrossRef]
- Shieh, W.Y.; Lin, Y.T.; Jean, W.D. Pseudovibrio denitrificans gen. nov., sp. nov., a marine, facultatively anaerobic, fermentative bacterium capable of denitrification. Int. J. Syst. Evol. Microbiol. 2004, 54, 2307–2312. [Google Scholar] [CrossRef] [Green Version]
- Graca, A.P.; Bondoso, J.; Gaspar, H.; Xavier, J.R.; Monteiro, M.C.; de la Cruz, M.; Oves-Costales, D.; Vicente, F.; Lage, O.M. Antimicrobial activity of heterotrophic bacterial communities from the marine sponge Erylus discophorus (Astrophorida, Geodiidae). PLoS ONE 2013, 8, e78992. [Google Scholar] [CrossRef] [Green Version]
- Flemer, B.; Kennedy, J.; Margassery, L.M.; Morrissey, J.P.; O’Gara, F.; Dobson, A.D. Diversity and antimicrobial activities of microbes from two Irish marine sponges, Suberites carnosus and Leucosolenia sp. J. Appl. Microbiol. 2012, 112, 289–301. [Google Scholar] [CrossRef]
- Sanchez, L.A.; Hedstrom, M.; Delgado, M.A.; Delgado, O.D. Production, purification and characterization of serraticin A, a novel cold-active antimicrobial produced by Serratia proteamaculans 136. J. Appl. Microbiol. 2010, 109, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.Q.; Zhang, S.Y.; Wang, N.; Li, Z.L.; Hua, H.M.; Hu, J.C.; Wang, S.J. New spirotetronate antibiotics, lobophorins H and I, from a South China Sea-derived Streptomyces sp. 12A35. Mar. Drugs 2013, 11, 3891–3901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.K.; Park, H.; Oh, T.J. Antibacterial and antioxidant capacity of polar microorganisms isolated from Arctic lichen Ochrolechia sp. Pol. J. Microbiol. 2014, 63, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Lo Giudice, A.; Bruni, V.; Michaud, L. Characterization of Antarctic psychrotrophic bacteria with antibacterial activities against terrestrial microorganisms. J. Basic Microbiol. 2007, 47, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Papaleo, M.C.; Fondi, M.; Maida, I.; Perrin, E.; Lo Giudice, A.; Michaud, L.; Mangano, S.; Bartolucci, G.; Romoli, R.; Fani, R. Sponge-associated microbial Antarctic communities exhibiting antimicrobial activity against Burkholderia cepacia complex bacteria. Biotechnol. Adv. 2012, 30, 272–293. [Google Scholar] [CrossRef]
- Gesheva, V.; Vasileva-Tonkova, E. Production of enzymes and antimicrobial compounds by halophilic Antarctic Nocardioides sp. grown on different carbon sources. World J. Microbiol. Biotechnol. 2012, 28, 2069–2076. [Google Scholar] [CrossRef] [Green Version]
- Hay, M.; Thomas, D.W.; Craighead, J.L.; Economides, C.; Rosenthal, J. Clinical development success rates for investigational drugs. Nat. Biotechnol. 2014, 32, 40–51. [Google Scholar] [CrossRef]
- DiMasi, J.A.; Grabowski, H.G.; Hansen, R.W. Innovation in the pharmaceutical industry: New estimates of R&D costs. J. Health Econ. 2016, 47, 20–33. [Google Scholar]
- Trussell, P.C.; Baird, E.A. A rapid method for the assay of penicillin. Can. J. Res. 1947, 25, 5–8. [Google Scholar] [CrossRef]
- Rex, J.; Ghannoum, M.A.; Alexander, D.B.; Andes, D.; Brown, D.A.; Diekema, D.J.; Espinel-Ingroff, A.; Fowler, C.L.; Johnson, E.J.; Knapp, C.C.; et al. Method for Antifungal Disk Diffusion Susceptibility Testing of Nondermatophyte Filamentous Fungi; Approved Guideline; CLSI Document M51-A; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2010; Volume 30, pp. 1–29. [Google Scholar]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, M.P.; Patel, J.B.; Bobenchik, A.M.; Campeau, S.; Cullen, S.K.; Gallas, M.F.; Gold, H.; Humphries, R.M.; Kirn, T.J.; Lewis, J.S.; et al. Performance Standards for Antimicrobial Disk Susceptibility Tests: Approved Standard, 29th ed.; Clinical and Laboratory Standards Institute Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019; Volume 32. [Google Scholar]
- White, R.L.; Burgess, D.S.; Manduru, M.; Bosso, J.A. Comparison of three different in vitro methods of detecting synergy: Time-kill, checkerboard, and E test. Antimicrob. Agents Chemother. 1996, 40, 1914–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Esquilín, A.E.; Roane, T.M. Antifungal activities of actinomycete strains associated with high-altitude sagebrush rhizosphere. J. Ind. Microbiol. Biotechnol. 2005, 32, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Sci, W.J. A Comparison of Two Methods Used for Measuring the Antagonistic Activity of Bacillus Species. Culture 2008, 5, 161–171. [Google Scholar]
- Ali-Shtayeh, M.S.; Abu Ghdeib, S.I. Antifungal activity of plant extracts against dermatophytes. Mycoses 1999, 42, 665–672. [Google Scholar] [CrossRef]
- Kumar, S.N.; Nambisan, B.; Sundaresan, A.; Mohandas, C.; Anto, R.J. Isolation and identification of antimicrobial secondary metabolites from Bacillus cereus associated with a rhabditid entomopathogenic nematode. Ann. Microbiol. 2014, 64, 209–218. [Google Scholar] [CrossRef]
- Menon, T.; Umamaheswari, K.; Kumarasamy, N.; Solomon, S.; Thyagarajan, S.P. Efficacy of fluconazole and itraconazole in the treatment of oral candidiasis in HIV patients. Acta Trop. 2001, 80, 151–154. [Google Scholar] [CrossRef]
- Imhof, A.; Balajee, S.A.; Marr, K.A. New Methods To Assess Susceptibilities of Aspergillus Isolates to Caspofungin. J. Clin. Microbiol. 2003, 41, 5683–5688. [Google Scholar]
- Mock, M.; Monod, M.; Baudraz-Rosselet, F.; Panizzon, R.G. Tinea capitis dermatophytes: Susceptibility to antifungal drugs tested in vitro and in vivo. Dermatology 1998, 197, 361–367. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Sheehan, D.J.; Rex, J.H. Determination of Fungicidal Activities against Yeasts and Molds: Lessons Learned from Bactericidal Testing and the Need for Standardization. Clin. Microbiol. Rev. 2004, 17, 268–280. [Google Scholar] [CrossRef] [Green Version]
- Crouch, S.P.M.; Kozlowski, R.; Slater, K.J.; Fletcher, J. Use of ATP as a measure of cell proliferation and cell toxicity. J. Immunol. Methods 1993, 160, 81–88. [Google Scholar] [CrossRef]
- Paloque, L.; Vidal, N.; Casanova, M.; Dumètre, A.; Verhaeghe, P.; Parzy, D.; Azas, N. A new, rapid and sensitive bioluminescence assay for drug screening on Leishmania. J. Microbiol. Methods 2013, 95, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Finger, S.; Wiegand, C.; Buschmann, H.J.; Hipler, U.C. Antibacterial properties of cyclodextrin-antiseptics-complexes determined by microplate laser nephelometry and ATP bioluminescence assay. Int. J. Pharm. 2013, 452, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Galiger, C.; Brock, M.; Jouvion, G.; Savers, A.; Parlato, M.; Ibrahim-Granet, O. Assessment of efficacy of antifungals against Aspergillus fumigatus: Value of real-time bioluminescence imaging. Antimicrob. Agents Chemother. 2013, 57, 3046–3059. [Google Scholar] [CrossRef] [Green Version]
- Peyron, F.; Favel, A.; Guiraud-Dauriac, H.; El Mzibri, M.; Chastin, C.; Duménil, G.; Regli, P. Evaluation of a flow cytofluorometric method for rapid determination of amphotericin B susceptibility of yeast isolates. Antimicrob. Agents Chemother. 1997, 41, 1537–1540. [Google Scholar] [CrossRef] [Green Version]
- Paparella, A.; Taccogna, L.; Aguzzi, I.; Chaves-López, C.; Serio, A.; Marsilio, F.; Suzzi, G. Flow cytometric assessment of the antimicrobial activity of essential oils against Listeria monocytogenes. Food Control 2008, 19, 1174–1182. [Google Scholar] [CrossRef]
- Landaburu, L.U.; Berenstein, A.J.; Videla, S.; Maru, P.; Shanmugam, D.; Chernomoretz, A.; Agüero, F. TDR Targets 6: Driving drug discovery for human pathogens through intensive chemogenomic data integration. Nucleic Acids Res. 2020, 48, D992. [Google Scholar]
- Bumol, T.F.; Watanabe, A.M. Genetic Information, Genomic Technologies, and the Future of Drug Discovery. JAMA 2001, 285, 551–555. [Google Scholar] [CrossRef] [Green Version]
- Carroll, P.M.; Dougherty, B.; Ross-Macdonald, P.; Browman, K.; FitzGerald, K. Model systems in drug discovery: Chemical genetics meets genomics. Pharmacol. Ther. 2003, 99, 183–220. [Google Scholar] [CrossRef]
- Chavali, A.K.; D’Auria, K.M.; Hewlett, E.L.; Pearson, R.D.; Papin, J.A. A metabolic network approach for the identification and prioritization of antimicrobial drug targets. Trends Microbiol. 2012, 20, 113. [Google Scholar] [CrossRef] [Green Version]
- Uddin, R.; Saeed, K.; Khan, W.; Azam, S.S.; Wadood, A. Metabolic pathway analysis approach: Identification of novel therapeutic target against methicillin resistant Staphylococcus aureus. Gene 2015, 556, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Mahmud, A.; Iqbal, A.; Hoque, S.F.; Hasan, M. Subtractive genomics approach towards the identification of novel therapeutic targets against human Bartonella bacilliformis. Inform. Med. Unlocked 2020, 20, 100385. [Google Scholar] [CrossRef]
- Shahid, F.; Shehroz, M.; Zaheer, T.; Ali, A. Subtractive Genomics Approaches: Towards Anti-Bacterial Drug Discovery. Front. Anti-Infect. Drug Discov. 2020, 8, 144–158. [Google Scholar]
- Zhan, P.; Pannecouque, C.; De Clercq, E.; Liu, X. Anti-HIV Drug Discovery and Development: Current Innovations and Future Trends. J. Med. Chem. 2016, 59, 2849–2878. [Google Scholar] [CrossRef]
- Hackbarth, C.J.; Chen, D.Z.; Lewis, J.G.; Clark, K.; Mangold, J.B.; Cramer, J.A.; Margolis, P.S.; Wang, W.; Koehn, J.; Wu, C.; et al. N-alkyl urea hydroxamic acids as a new class of peptide deformylase inhibitors with antibacterial activity. Antimicrob. Agents Chemother. 2002, 46, 2752–2764. [Google Scholar] [CrossRef] [Green Version]
- Naor, N.; Gadot, O.; Meir, M.; Barkan, D. Peptide Deformylase (def) is essential in Mycobacterium smegmatis, but the essentiality is compensated by inactivation of methionine formylation. BMC Microbiol. 2019, 19, 232. [Google Scholar] [CrossRef]
- Teo, J.W.; Thayalan, P.; Beer, D.; Yap, A.S.; Nanjundappa, M.; Ngew, X.; Duraiswamy, J.; Liung, S.; Dartois, V.; Schreiber, M.; et al. Peptide deformylase inhibitors as potent antimycobacterial agents. Antimicrob. Agents Chemother. 2006, 50, 3665–3673. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Hackbarth, C.; Ni, Z.J.; Wu, C.; Wang, W.; Jain, R.; He, Y.; Bracken, K.; Weidmann, B.; Patel, D.V.; et al. Peptide deformylase inhibitors as antibacterial agents: Identification of VRC3375, a proline-3-alkylsuccinyl hydroxamate derivative, by using an integrated combinatorial and medicinal chemistry approach. Antimicrob. Agents Chemother. 2004, 48, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, N.; Albert, M.; Awrey, D.; Bardouniotis, E.; Berman, J.; Clarke, T.; Dorsey, M.; Hafkin, B.; Ramnauth, J.; Romanov, V.; et al. Mode of action, in vitro activity, and in vivo efficacy of AFN-1252, a selective antistaphylococcal FabI inhibitor. Antimicrob. Agents Chemother. 2012, 56, 5865–5874. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.J.; White, S.W.; Rock, C.O. Inhibitors of fatty acid synthesis as antimicrobial chemotherapeutics. Appl. Microbiol. Biotechnol. 2002, 58, 695–703. [Google Scholar]
- Wohlleben, W.; Mast, Y.; Stegmann, E.; Ziemert, N. Antibiotic Drug Discovery. Microb. Biotechnol. 2016, 9, 541–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanelli, U.; Pappalardo, M.; Chine, V.; Gismondi, P.; Neglia, C.; Argentiero, A.; Calderaro, A.; Prati, A.; Esposito, S. Role of Artificial Intelligence in Fighting Antimicrobial Resistance in Pediatrics. Antibiotics 2020, 9, 767. [Google Scholar] [CrossRef]
- Vamathevan, J.; Clark, D.; Czodrowski, P.; Dunham, I.; Ferran, E.; Lee, G.; Li, B.; Madabhushi, A.; Shah, P.; Spitzer, M.; et al. Applications of machine learning in drug discovery and development. Nat. Rev. Drug Discov. 2019, 18, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.J.; Lim, C.H.; Foo, S.C.; Tan, H.S. The role of artificial intelligence in the battle against antimicrobial-resistant bacteria. Curr. Genet. 2021, 67, 421–429. [Google Scholar] [CrossRef]
- Stokes, J.M.; Yang, K.; Swanson, K.; Jin, W.; Cubillos-Ruiz, A.; Donghia, N.M.; MacNair, C.R.; French, S.; Carfrae, L.A.; Bloom-Ackermann, Z.; et al. A Deep Learning Approach to Antibiotic Discovery. Cell 2020, 181, 475–483. [Google Scholar] [CrossRef]
- Gravato-Nobre, M.J.; Hodgkin, J. Caenorhabditis elegans as a model for innate immunity to pathogens. Cell. Microbiol. 2005, 7, 741–751. [Google Scholar] [CrossRef]
- Rasheed, S.; Fries, F.; Muller, R.; Herrmann, J. Zebrafish: An Attractive Model to Study Staphylococcus aureus Infection and Its Use as a Drug Discovery Tool. Pharmaceuticals 2021, 14, 594. [Google Scholar] [CrossRef]
- Antoine, T.E.; Jones, K.S.; Dale, R.M.; Shukla, D.; Tiwari, V. Zebrafish: Modeling for herpes simplex virus infections. Zebrafish 2014, 11, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Burgos, J.S.; Ripoll-Gomez, J.; Alfaro, J.M.; Sastre, I.; Valdivieso, F. Zebrafish as a new model for herpes simplex virus type 1 infection. Zebrafish 2008, 5, 323–333. [Google Scholar] [CrossRef]
- Ugur, B.; Chen, K.; Bellen, H.J. Drosophila tools and assays for the study of human diseases. Dis. Models Mech. 2016, 9, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Chamilos, G.; Samonis, G.; Kontoyiannis, D.P. Drosophila melanogaster as a model host for the study of microbial pathogenicity and the discovery of novel antimicrobial compounds. Curr. Pharm. Des. 2011, 17, 1246–1253. [Google Scholar] [CrossRef]
- Champion, O.L.; Wagley, S.; Titball, R.W. Galleria mellonella as a model host for microbiological and toxin research. Virulence 2016, 7, 840–845. [Google Scholar] [CrossRef] [Green Version]
- Lebreton, F.; Le Bras, F.; Reffuveille, F.; Ladjouzi, R.; Giard, J.C.; Leclercq, R.; Cattoir, V. Galleria mellonella as a model for studying Enterococcus faecium host persistence. J. Mol. Microbiol. Biotechnol. 2011, 21, 191–196. [Google Scholar] [CrossRef]
- Norville, I.H.; Hartley, M.G.; Martinez, E.; Cantet, F.; Bonazzi, M.; Atkins, T.P. Galleria mellonella as an alternative model of Coxiella burnetii infection. Microbiology 2014, 160, 1175–1181. [Google Scholar] [CrossRef] [Green Version]
- Hamamoto, H.; Tonoike, A.; Narushima, K.; Horie, R.; Sekimizu, K. Silkworm as a model animal to evaluate drug candidate toxicity and metabolism. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 149, 334–339. [Google Scholar] [CrossRef]
- Giunti, S.; Andersen, N.; Rayes, D.; De Rosa, M.J. Drug discovery: Insights from the invertebrate Caenorhabditis elegans. Pharmacol. Res. Perspect. 2021, 9, e00721. [Google Scholar] [CrossRef]
- Artal-Sanz, M.; de Jong, L.; Tavernarakis, N. Caenorhabditis elegans: A versatile platform for drug discovery. Biotechnol. J. 2006, 1, 1405–1418. [Google Scholar] [CrossRef]
- Uccelletti, D.; Zanni, E.; Marcellini, L.; Palleschi, C.; Barra, D.; Mangoni, M.L. Anti-Pseudomonas activity of frog skin antimicrobial peptides in a Caenorhabditis elegans infection model: A plausible mode of action in vitro and in vivo. Antimicrob. Agents Chemother. 2010, 54, 3853–3860. [Google Scholar] [CrossRef] [Green Version]
- Swem, L.R.; Swem, D.L.; O’Loughlin, C.T.; Gatmaitan, R.; Zhao, B.; Ulrich, S.M.; Bassler, B.L. A quorum-sensing antagonist targets both membrane-bound and cytoplasmic receptors and controls bacterial pathogenicity. Mol. Cell 2009, 35, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Takaki, K.; Cosma, C.L.; Troll, M.A.; Ramakrishnan, L. An in vivo platform for rapid high-throughput antitubercular drug discovery. Cell Rep. 2012, 2, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Malkani, N.; Schmid, J.A. Some secrets of fluorescent proteins: Distinct bleaching in various mounting fluids and photoactivation of cyan fluorescent proteins at YFP-excitation. PLoS ONE 2011, 6, e18586. [Google Scholar] [CrossRef]
- Parker, G.J.; Law, T.L.; Lenoch, F.J.; Bolger, R.E. Development of high throughput screening assays using fluorescence polarization: Nuclear receptor-ligand-binding and kinase/phosphatase assays. J. Biomol. Screen. 2000, 5, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Bazin, H.; Préaudat, M.; Trinquet, E.; Mathis, G. Homogeneous time resolved fluorescence resonance energy transfer using rare earth cryptates as a tool for probing molecular interactions in biology. Spectrochim. Acta-Part A Mol. Biomol. Spectrosc. 2001, 57, 2197–2211. [Google Scholar] [CrossRef]
- Hajare, A.A.; Salunkhe, S.S.; Sachin, M.S.; Gorde, S.S.; Nadaf, J.S.; Pishawikar, A.S. Review on: High-throughput screening is an approach to drug discovery. Review On: High-Throughput Screening Is An Approach To Drug. Am. J. Pharmtech Res. 2014, 4, 113–129. [Google Scholar]
- Sinha, R.; Shukla, P. Antimicrobial Peptides: Recent Insights on Biotechnological Interventions and Future Perspectives. Protein Pept. Lett. 2019, 26, 79–87. [Google Scholar] [CrossRef]


| Microorganism | Chemical Compound | Molecular Class | Antimicrobial Activity | Reference |
|---|---|---|---|---|
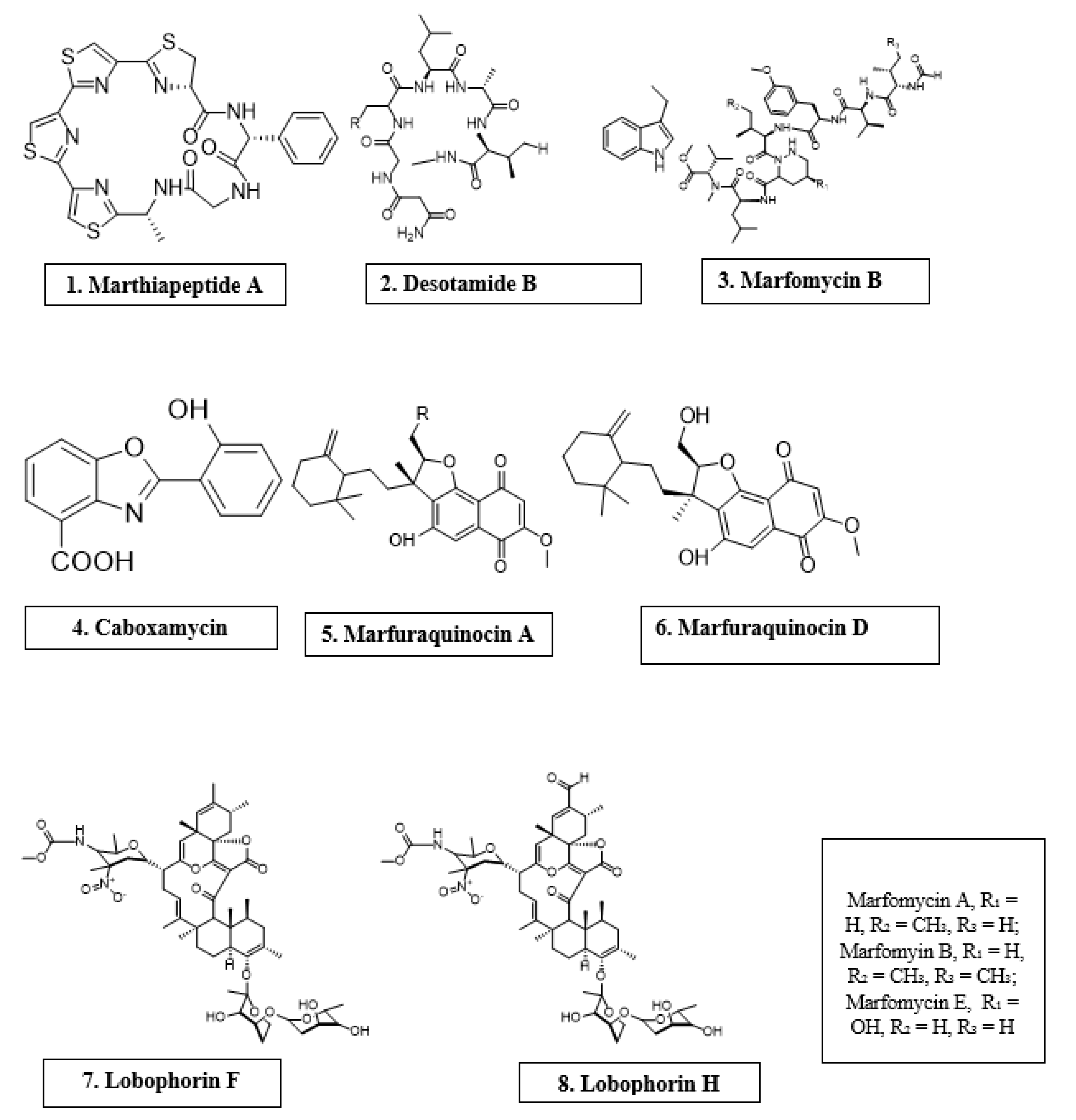
| Marinactinospora thermotolerans | Marthiapeptide A | Cyclic peptide | S. aureus, M. luteus, B. subtillis, B. thuringiensis | [12] |
| Streptomyces scopuliridis | Desotamide B | Cyclic peptide | S. aureus, S. aureus | [12] |
| Streptomyces drozdowiczii | Marfomycins A, B, E | Cyclic peptide | M. luteus | |
| Verrucosispora spp. | Abyssomicin C | Spirotetronate polyketides | Methicillin-resistant Staphylococcus aureus, Vancomycin-resistant Staphylococcus aureus | [12] |
| Streptomyces spp. | Lobophorin F | Spirotetronate polyketides | S. aureus, E. feacalis | [12] |
| Streptomyces spp. | Lobophorin H | Spirotetronate polyketides | B. subtilis | [12] |
| Streptomyces sp. | Caboxamycin | Alkaloid | S. epidermis, S. lentus, B. subtillis | [12] |
| Streptomyces niveus | Marfuraquinocin A, D | Sesquiterpene derivative | S. aureus, Methicillin-resistant Staphylococcus aureus | [12] |
| Microorganism | Compound(s) | Susceptible Organism(s) | Reference |
|---|---|---|---|
| A. hibisca | Pradimicins A, B, C | Candida spp. and Aspergillus spp. | [58] |
| Actinoplanes spp. | Purpuromycin | T. mentagrophytes | [60] |
| Micromonospora species ATCC 53803 | Spartanamycin B | C. albicans, A. cladosporium, and Cryptococcus spp. | [61] |
| M. neiheumicin | Neihumicin | S. cerevisae | [61] |
| Micromonospora species SCC 1792 | Sch 37137 | Dermatophytes and Candida spp. | [62] |
| B. subtilis | Iturin A and related peptides | Phytopathogens | [60,72] |
| Micromonospora species SF-1917 | Dapiramicins A and B | R. solania | [63] |
| B. cereus | Azoxybacilin, Bacereutin, Cispentacin, and Mycocerein | Aspergillus spp., Saccharomyces spp, and C. albicans | [60] |
| B. lichenformis | Fungicin M-4 | Microsporum canis, Mucor spp., and Sporothrix spp. | [73,74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaning Danquah, C.; Minkah, P.A.B.; Osei Duah Junior, I.; Amankwah, K.B.; Somuah, S.O. Antimicrobial Compounds from Microorganisms. Antibiotics 2022, 11, 285. https://doi.org/10.3390/antibiotics11030285
Amaning Danquah C, Minkah PAB, Osei Duah Junior I, Amankwah KB, Somuah SO. Antimicrobial Compounds from Microorganisms. Antibiotics. 2022; 11(3):285. https://doi.org/10.3390/antibiotics11030285
Chicago/Turabian StyleAmaning Danquah, Cynthia, Prince Amankwah Baffour Minkah, Isaiah Osei Duah Junior, Kofi Bonsu Amankwah, and Samuel Owusu Somuah. 2022. "Antimicrobial Compounds from Microorganisms" Antibiotics 11, no. 3: 285. https://doi.org/10.3390/antibiotics11030285
APA StyleAmaning Danquah, C., Minkah, P. A. B., Osei Duah Junior, I., Amankwah, K. B., & Somuah, S. O. (2022). Antimicrobial Compounds from Microorganisms. Antibiotics, 11(3), 285. https://doi.org/10.3390/antibiotics11030285