
Control of Early Blight Fungus (Alternaria alternata) in Tomato by Boric and Phenylboronic Acid

 ,
, 
 and
and 
Abstract
:
1. Introduction
2. Results
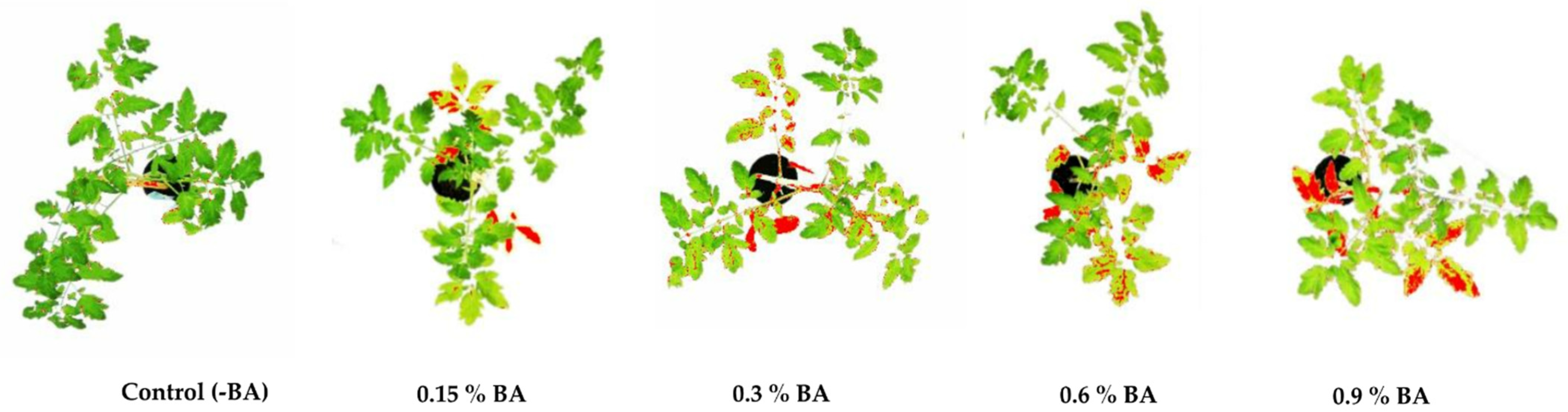
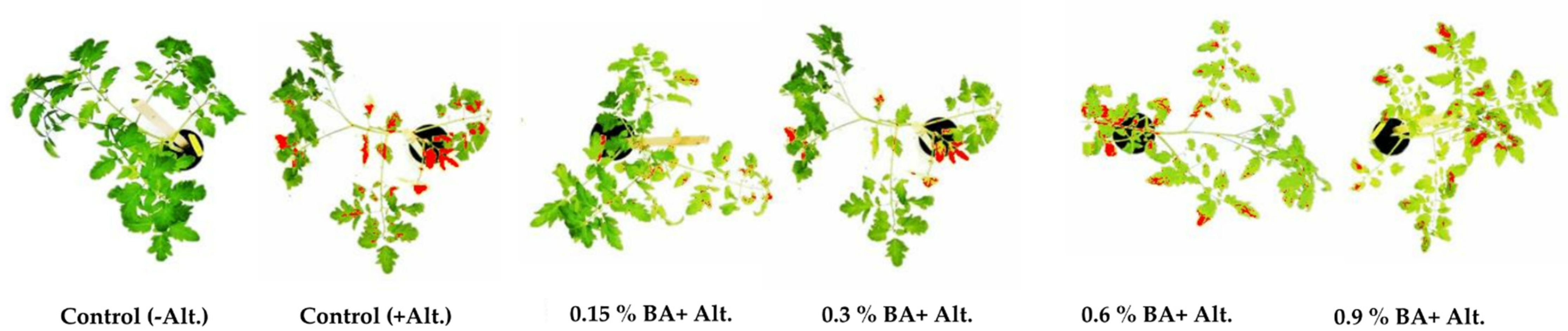
2.1. BA Effect on Tomato Plants
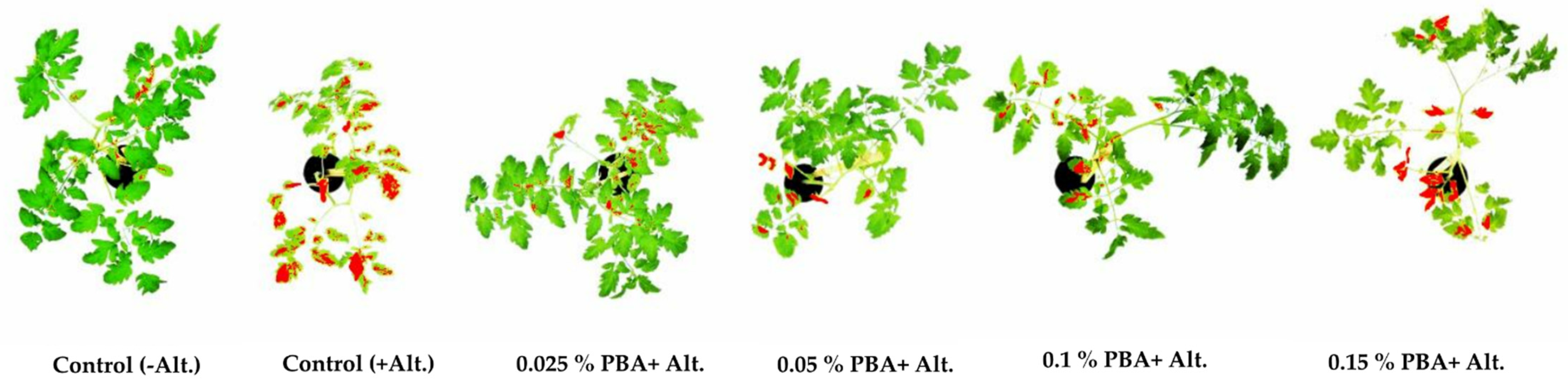
2.2. PBA Effect on Tomato Plants
2.3. Antifungal Effect of BA on Alternaria alternata
2.4. Antifungal Effect of PBA on Alternaria alternata
3. Discussion
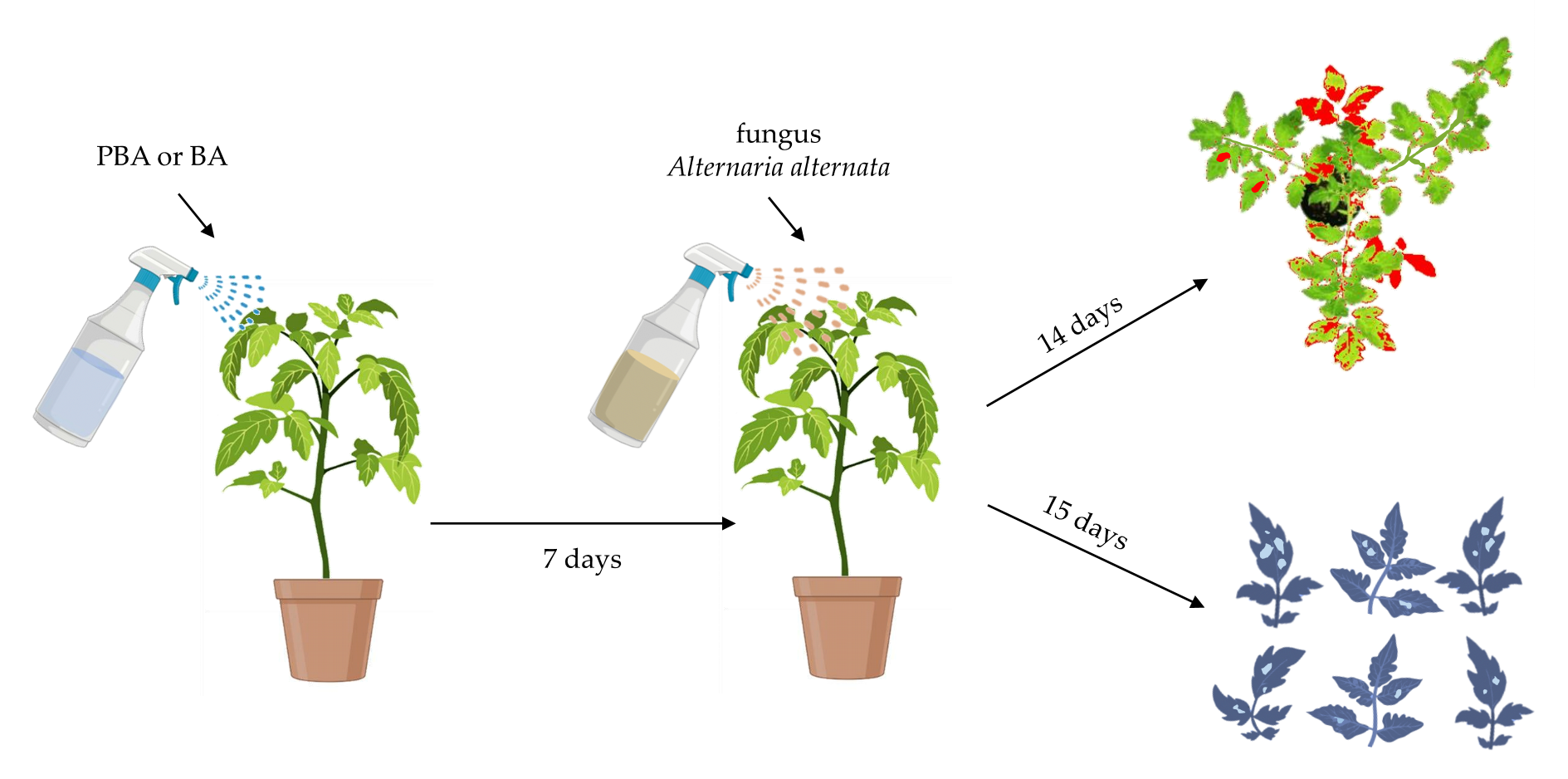
4. Materials and Methods
4.1. Preparation of BA and PBA Solutions
4.2. Preparation of Suspension of A. alternata Spores
4.3. Implementation of the Experiment In Vivo
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Michaelis, A.; Becker, B. Ueber Monophenylborchlorid und die Valenz des Bors. Ber. Dtsch. Chem. Ges. 1880, 13, 58–61. [Google Scholar] [CrossRef] [Green Version]
- Lopalco, A.; Lopedota, A.A.; Laquintana, V.; Denora, N.; Stella, V.J. Boric Acid, a Lewis Acid With Unique and Unusual Properties: Formulation Implications. J. Pharm. Sci. 2020, 109, 2375–2386. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.H.; Tankard, A.R. The determination of boric acid in cider, fruits, etc. Analyst 1904, 29, 301–304. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Laks, P.E.; Pruner, M.S. A preliminary report on the wood preservative properties of phenylboronic acid. For. Prod. J. 1994, 44, 46–48. [Google Scholar]
- Freeman, A.; Village, Y.; Segal, R.; Dror, Y. Methods and Compositions for Treating Fungal Infections. Available online: https://patentimages.storage.googleapis.com/02/6b/2f/a0a7dd363f4ba6/US20040220146A1.pdf (accessed on 23 July 2021).
- Yalinkilic, M.K.; Yoshimura, T.; Takahashi, M. Enhancement of the biological resistance of wood by phenylboronic acid treatment. J. Wood Sci. 1998, 44, 152–157. [Google Scholar] [CrossRef]
- Marasović, M.; Ivanković, S.; Stojković, R.; Đermić, D.; Galić, B.; Miloš, M. In vitro and in vivo antitumour effects of phenylboronic acid against mouse mammary adenocarcinoma 4T1 and squamous carcinoma SCCVII cells. J. Enzym. Inhib. Med. Chem. 2017, 32, 1299–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, A.; Sato, N.; Miyahara, Y. Label free carbohydrate detection by using phenylboronic acid gate-modified field effect transistor. Curr. Appl. Phys. 2009, 9, e214–e217. [Google Scholar] [CrossRef]
- Huang, L.-L.; Jin, Y.-J.; Zhao, D.; Yu, C.; Hao, J.; Xie, H.-Y. A fast and biocompatible living virus labeling method based on sialic acid-phenylboronic acid recognition system. Anal. Bioanal. Chem. 2014, 406, 2687–2693. [Google Scholar] [CrossRef]
- De Seta, F.; Schmidt, M.; Vu, B.; Essmann, M.; Larsen, B. Antifungal mechanisms supporting boric acid therapy of Candida vaginitis. J. Antimicrob. Chemother. 2008, 63, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Kohno, J.; Kawahata, T.; Otake, T.; Morimoto, M.; Mori, H.; Ueba, N.; Nishio, M.; Kinumaki, A.; Komatsubara, S.; Kawashima, K. Boromycin, an Anti-HIV Antibiotic. Biosci. Biotechnol. Biochem. 1996, 60, 1036–1037. [Google Scholar] [CrossRef]
- Adamczyk-Woźniak, A.; Komarovska-Porokhnyavets, O.; Misterkiewicz, B.; Novikov, V.P.; Sporzyński, A. Biological activity of selected boronic acids and their derivatives. Appl. Organomet. Chem. 2012, 26, 390–393. [Google Scholar] [CrossRef]
- Wieczorek, D.; Lipok, J.; Borys, K.M.; Adamczyk-Woźniak, A.; Sporzyński, A. Investigation of fungicidal activity of 3-piperazine-bis(benzoxaborole) and its boronic acid analogue. Appl. Organomet. Chem. 2014, 28, 347–350. [Google Scholar] [CrossRef]
- Bonnen, A.; Hammerschmidt, R. Cutinolytic enzymes from Colletotrichum lagenarium. Physiol. Mol. Plant Pathol. 1989, 35, 463–474. [Google Scholar] [CrossRef]
- Moghaddam, G.A.; Rezayatmand, Z.; Esfahani, M.N.; Khozaei, M. Genetic defense analysis of tomatoes in response to early blight disease, Alternaria alternata. Plant Physiol. Biochem. 2019, 142, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Hussein, G.S. The Use of PCR to Detect Alternaria alternata Fungi which Isolated from Tomato Fruit. Biochem. Cell. Arch. 2019, 19, 4203–4206. [Google Scholar] [CrossRef]
- Yang, L.-N.; He, M.-H.; Ouyang, H.-B.; Zhu, W.; Pan, Z.-C.; Sui, Q.-J.; Shang, L.-P.; Zhan, J. Cross-resistance of the pathogenic fungus Alternaria alternata to fungicides with different modes of action. BMC Microbiol. 2019, 19, 205. [Google Scholar] [CrossRef] [Green Version]
- Ramezani, Y.; Taheri, P.; Mamarabadi, M. Identification of Alternaria spp. associated with tomato early blight in Iran and investigating some of their virulence factors. J. Plant Pathol. 2019, 101, 647–659. [Google Scholar] [CrossRef]
- Chaerani, R.; Voorrips, R.E. Tomato early blight (Alternaria solani): The pathogen, genetics, and breeding for resistance. J. Gen. Plant Pathol. 2006, 72, 335–347. [Google Scholar] [CrossRef]
- Bessadat, N.; Berruyer, R.; Hamon, B.; Kihal, M.; Henni, D.E.; Simoneau, P.; Bataille-Simoneau, N.; Benichou, S. Alternaria species associated with early blight epidemics on tomato and other Solanaceae crops in northwestern Algeria. Eur. J. Plant Pathol. 2016, 148, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.; Zehra, A.; Dubey, M.K.; Aamir, M.; Gupta, V.K.; Upadhyay, R.S. Comparative Evaluation of Biochemical Changes in Tomato (Lycopersicon esculentum Mill.) Infected by Alternaria alternata and Its Toxic Metabolites (TeA, AOH, and AME). Front. Plant Sci. 2016, 7, 1408. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.C. Diseases of Vegetable Crops, 1st ed.; MacGraw-Hill: New York, NY, USA, 1952; pp. 190–191. [Google Scholar]
- Rogerson, C.T.; Sherf, A.E.; Macnab, A.A. Vegetable Diseases and Their Control. Brittonia 1987, 39, 378. [Google Scholar] [CrossRef]
- Wang, Y.; Bao, Y.; Shen, D.; Feng, W.; Yu, T.; Zhang, J.; Zheng, X.D. Biocontrol of Alternaria alternata on cherry tomato fruit by use of marine yeast Rhodosporidium paludigenum Fell & Tallman. Int. J. Food Microbiol. 2008, 123, 234–239. [Google Scholar] [CrossRef]
- El-Gazzar, N.; Ismail, A.M. The potential use of Titanium, Silver and Selenium nanoparticles in controlling leaf blight of tomato caused by Alternaria alternata. Biocatal. Agric. Biotechnol. 2020, 27, 101708. [Google Scholar] [CrossRef]
- Dinh, A.; Naeem, A.; Sagervanshi, A.; Mühling, K. One-Time Foliar Application and Continuous Resupply via Roots Equally Improved the Growth and Physiological Response of B-Deficient Oilseed Rape. Plants 2021, 10, 866. [Google Scholar] [CrossRef]
- Du, W.; Pan, Z.-Y.; Hussain, S.B.; Han, Z.-X.; Peng, S.-A.; Liu, Y.-Z. Foliar Supplied Boron Can Be Transported to Roots as a Boron-Sucrose Complex via Phloem in Citrus Trees. Front. Plant Sci. 2020, 11, 250. [Google Scholar] [CrossRef]
- Gómez-Soto, D.; Galván, S.; Rosales, E.; Bienert, P.; Abreu, I.; Bonilla, I.; Bolaños, L.; Reguera, M. Insights into the role of phytohormones regulating pAtNIP5;1 activity and boron transport in Arabidopsis thaliana. Plant Sci. 2019, 287, 110198. [Google Scholar] [CrossRef]
- Ustin, S.L.; A Gitelson, A.; Jacquemoud, S.; Schaepman, E.M.; Asner, G.P.; Gamon, A.J.; Zarco-Tejada, P. Retrieval of foliar information about plant pigment systems from high resolution spectroscopy. Remote Sens. Environ. 2009, 113, S67–S77. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A.; Chivkunova, O.B.; Merzlyak, M.N. Nondestructive estimation of anthocyanins and chlorophylls in anthocyanic leaves. Am. J. Bot. 2009, 96, 1861–1868. [Google Scholar] [CrossRef]
- Feild, T.S.; Lee, D.W.; Holbrook, N.M. Why Leaves Turn Red in Autumn. The Role of Anthocyanins in Senescing Leaves of Red-Osier Dogwood. Plant Physiol. 2001, 127, 566–574. [Google Scholar] [CrossRef]
- Wrolstad, R.E. Anthocyanin Pigments-Bioactivity and Coloring Properties. J. Food Sci. 2006, 69, C419–C425. [Google Scholar] [CrossRef]
- Lev-Yadun, S.; Gould, K.S. Role of Anthocyanins in Plant Defence. In Anthocyanins; Springer: New York, NY, USA, 2008; pp. 22–28. [Google Scholar] [CrossRef]
- Kiferle, C.; Fantini, E.; Bassolino, L.; Povero, G.; Spelt, C.; Buti, S.; Giuliano, G.; Quattrocchio, F.M.; Koes, R.; Perata, P.; et al. Tomato R2R3-MYB Proteins SlANT1 and SlAN2: Same Protein Activity, Different Roles. PLoS ONE 2015, 10, e0136365. [Google Scholar] [CrossRef] [Green Version]
- Sivankalyani, V.; Feygenberg, O.; Diskin, S.; Wright, B.; Alkan, N. Increased anthocyanin and flavonoids in mango fruit peel are associated with cold and pathogen resistance. Postharvest Biol. Technol. 2016, 111, 132–139. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Gould, K.S. Nature’s Swiss Army Knife: The Diverse Protective Roles of Anthocyanins in Leaves. J. Biomed. Biotechnol. 2004, 2004, 314–320. [Google Scholar] [CrossRef] [Green Version]
- de Vries, S.; von Dahlen, J.K.; Schnake, A.; Ginschel, S.; Schulz, B.; Rose, E.L. Broad-spectrum inhibition of Phytophthora infestans by fungal endophytes. FEMS Microbiol. Ecol. 2018, 94, fiy037. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, C. Changes in pigment accumulation as indicators of physical stress in Eucalyptus pauciflora. Field Stud. Ecol. 2019, 2. Available online: https://studentjournals.anu.edu.au/index.php/fse/article/view/220 (accessed on 3 May 2021).
- Meng, L.; Mestdagh, H.; Ameye, M.; Audenaert, K.; Höfte, M.; Van Labeke, M.-C. Phenotypic Variation of Botrytis cinerea Isolates Is Influenced by Spectral Light Quality. Front. Plant Sci. 2020, 11, 1233. [Google Scholar] [CrossRef]
- Romero-Pérez, A.; Ameye, M.; Audenaert, K.; Van Damme, E.J.M. Overexpression of F-Box Nictaba Promotes Defense and Anthocyanin Accumulation in Arabidopsis thaliana After Pseudomonas syringae Infection. Front. Plant Sci. 2021, 12, 692606. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, Q.; Bu, Y.; Luo, R.; Hao, S.; Zhang, J.; Tian, J.; Yao, Y. Flavonoid Accumulation Plays an Important Role in the Rust Resistance of Malus Plant Leaves. Front. Plant Sci. 2017, 8, 1286. [Google Scholar] [CrossRef] [Green Version]
- Štambuk, P.; Šikuten, I.; Preiner, D.; Nimac, A.; Lazarević, B.; Marković, Z.; Maletić, E.; Kontić, J.; Tomaz, I. Screening of Croatian Native Grapevine Varieties for Susceptibility to Plasmopara viticola Using Leaf Disc Bioassay, Chlorophyll Fluorescence, and Multispectral Imaging. Plants 2021, 10, 661. [Google Scholar] [CrossRef]
- Pye, M.F.; Ehakuno, F.; Macdonald, J.D.; Bostock, R.M. Induced resistance in tomato by SAR activators during predisposing salinity stress. Front. Plant Sci. 2013, 4, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uppalapati, S.R.; Ishiga, Y.; Wangdi, T.; Urbanczyk-Wochniak, E.; Ishiga, T.; Mysore, K.; Bender, C.L. Pathogenicity of Pseudomonas syringae pv. tomato on Tomato Seedlings: Phenotypic and Gene Expression Analyses of the Virulence Function of Coronatine. Mol. Plant Microbe Interact. 2008, 21, 383–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Shennawy, M.Z.; Abd El, A.M. Evaluation of Some Antioxidants against Tomato Early Blight. Available online: https://alexja.journals.ekb.eg/article_81831_25c37f21d6b2adc1640330edd2083fdc.pdf (accessed on 19 July 2021).
- Ogawa, D.; Nakajima, N.; Seo, S.; Mitsuhara, I.; Kamada, H.; Ohashi, Y. The phenylalanine pathway is the main route of salicylic acid biosynthesis in Tobacco mosaic virus-infected tobacco leaves. Plant Biotechnol. 2006, 23, 395–398. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Hwang, B.K. An important role of the pepper phenylalanine ammonia-lyase gene (PAL1) in salicylic acid-dependent signalling of the defence response to microbial pathogens. J. Exp. Bot. 2014, 65, 2295–2306. [Google Scholar] [CrossRef] [Green Version]
- Hao, Q.; Wang, W.; Han, X.; Wu, J.; Lyu, B.; Chen, F.; Caplan, A.; Li, C.; Wu, J.; Wang, W.; et al. Isochorismate-based salicylic acid biosynthesis confers basal resistance to Fusarium graminearum in barley. Mol. Plant Pathol. 2018, 19, 1995–2010. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Chen, S.; Wang, S.; Shan, W.; Wang, X.; Lin, Y.; Su, F.; Yang, Z.; Yu, X. Defensive Responses of Tea Plants (Camellia sinensis) Against Tea Green Leafhopper Attack: A Multi-Omics Study. Front. Plant Sci. 2020, 10, 1075. [Google Scholar] [CrossRef]
- Lefevere, H.; Bauters, L.; Gheysen, G. Salicylic Acid Biosynthesis in Plants. Front. Plant Sci. 2020, 11, 338. [Google Scholar] [CrossRef]
- Nawaz, M.; Ishaq, S.; Ishaq, H.; Khan, N.; Iqbal, N.; Ali, S.; Rizwan, M.; Alsahli, A.A.; Alyemeni, M.N. Salicylic Acid Improves Boron Toxicity Tolerance by Modulating the Physio-Biochemical Characteristics of Maize (Zea mays L.) at an Early Growth Stage. Agronomy 2020, 10, 2013. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I. Visible and near-infrared reflectance techniques for diagnosing plant physiological status. Trends Plant Sci. 1998, 3, 151–156. [Google Scholar] [CrossRef]
- Li, Y.; Yang, D.; Cui, J. Graphene oxide loaded with copper oxide nanoparticles as an antibacterial agent against Pseudomonas syringae pv. tomato. RSC Adv. 2017, 7, 38853–38860. [Google Scholar] [CrossRef] [Green Version]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Solovchenko, A.; Pogosyan, S.I. Application of Reflectance Spectroscopy for Analysis of Higher Plant Pigments. Russ. J. Plant Physiol. 2003, 50, 704–710. [Google Scholar] [CrossRef]
- Nashwa, S.; Abo-Elyousr, K. Evaluation of various plant extracts against the early blight disease of tomato plants under greenhouse and field conditions. Plant Prot. Sci. 2012, 48, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Meier, U. Growth Stages of Mono- and Dycotyledonous Plant. Available online: https://www.politicheagricole.it/flex/AppData/WebLive/Agrometeo/MIEPFY800/BBCHengl2001.pdf (accessed on 19 November 2021).
- Shamshiri, R.R.; Jones, J.W.; Thorp, K.; Ahmad, D.; Man, H.C.; Taheri, S. Review of optimum temperature, humidity, and vapour pressure deficit for microclimate evaluation and control in greenhouse cultivation of tomato: A review. Int. Agrophysics 2018, 32, 287–302. [Google Scholar] [CrossRef]
- Qadoos, M.; Kahn, M.I.; Suleman, M.; Khan, H.; Aqeel, M.; Rafiq, M. Comparison of Poison Food Technique and Drench Method for In Vitro Control of Alternaria sp., the Cause of Leaf Spot of Bitter Gourd. Available online: https://meritresearchjournals.org/asss/Content/2016/September/Qadoos%20et%20al.pdf (accessed on 14 December 2021).
- Nagrale, D.T.; Gaikwad, A.P.; Sharma, L. Morphological and cultural characterization of Alternaria alternata (Fr.) Keissler blight of gerbera (Gerbera jamesonii H. Bolus ex J.D. Hook). J. Appl. Nat. Sci. 2013, 5, 171–178. [Google Scholar] [CrossRef]
- Zheng, H.H.; Zhao, J.; Wang, T.Y.; Wu, X.H. Characterization of Alternaria species associated with potato foliar diseases in China. Plant Pathol. 2015, 64, 425–433. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Fallik, E.; Bashan, Y.; Okon, Y.; Cahaner, A.; Kedar, N. Inheritance and sources of resistance to bacterial speck of tomato caused by Pseudomonas syringae pv. tomato. Ann. Appl. Biol. 1983, 102, 365–371. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Laflamme, B.; Middleton, M.; Lo, T.; Desveaux, D.; Guttman, D.S. Image-Based Quantification of Plant Immunity and Disease. Mol. Plant Microbe Interact. 2016, 29, 919–924. [Google Scholar] [CrossRef] [Green Version]
- Tukey, J.W. Comparing Individual Means in the Analysis of Variance. Available online: http://webspace.ship.edu/pgmarr/Geo441/Readings/Tukey%201949%20-%20Comparing%20Individual%20Means%20in%20the%20Analysis%20of%20Variance.pdf (accessed on 27 February 2021).
- IBM Corp. IBM SPSS Statistics for Windows. Available online: https://hadoop.apache.org/ (accessed on 11 May 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BA Volume Concentration (%) | 0.0 | 0.15 | 0.3 | 0.6 | 0.9 |
|---|---|---|---|---|---|
| Mean value of physiological lesion area * ± SD | 0 a ± 0 | 9.5 b ± 1 | 22.3 c ± 2.1 | 34.4 d ± 1.4 | 44.4 e ± 2.2 |
| BA Concentration (%) | ARI Mean Value of Lesion Area ± SD | CHI Mean Value of Lesion Area ± SD | HUE Mean Value of Lesion Area ± SD | NIR Mean Value of Lesion Area ± SD |
|---|---|---|---|---|
| 0 | 2.46 ± 0.7 | 1.88 ± 0.4 | 99.3 ± 4.7 | 18,329.5 ± 1112.2 |
| 0.15 | 2.77 ± 0.9 | 1.96 ± 0.5 | 100.7 ± 8.7 | 18,203.5 ± 491.8 |
| 0.3 | 2.49 ± 1 | 1.81 ± 0.5 | 98.3 ± 8.4 | 18,832.1 ± 1112.5 |
| 0.6 | 2.27 ± 0.9 | 1.68 ± 0.6 | 95.3 ± 12.1 | 18,173.5 ± 937.8 |
| 0.9 | 2.11 ± 1 | 1.67 ± 0.5 | 95.9 ± 8.5 | 17,485.8 ± 1029.0 |
| PBA Concentration (%) | 0.0 | 0.025 | 0.05 | 0.1 | 0.15 |
|---|---|---|---|---|---|
| Mean value of physiological lesion area * ± SD | 0 a ± 0 | 6.7 b ± 0.7 | 24.7 c ± 2.3 | 31 d ± 3.3 | 29.2 cd ± 2.3 |
| PBA Concentration (%) | ARI Mean Value of Lesion Area ± SD | CHI Mean Value of Lesion Area ± SD | HUE Mean Value of Lesion Area ± SD | NIR Mean Value of Lesion Area ± SD |
|---|---|---|---|---|
| 0 | 2.46 ± 0.4 | 1.88 ± 0.9 | 99.3 ± 4.7 | 18,329.5 ± 1112.2 |
| 0.025 | 2.34 ± 0.5 | 1.91 ± 0.7 | 100.3 ± 7.7 | 18,201.7 ± 440.6 |
| 0.05 | 2.71 ± 0.5 | 2.03 ± 0.8 | 102.1 ± 5.8 | 18,351.3 ± 507.1 |
| 0.1 | 2.48 ± 0.5 | 1.90 ± 0.9 | 99.6 ± 7.4 | 18,302.7 ± 757.1 |
| 0.15 | 2.68 ± 0.5 | 1.95 ± 1.0 | 100.3 ± 7.0 | 18,158.3 ± 643.6 |
| BA Concentration (%) | 0.0 | 0.15 | 0.3 | 0.6 | 0.9 |
|---|---|---|---|---|---|
| Mean value of symptoms area * ± SD | 29.8 d ± 0.9 | 5 a ± 1.1 | 9.5 b ± 0.5 | 24 c ± 1.8 | 27.7 d ± 1.4 |
| BA Concentration (%) | ARI Mean Values of Symptoms * ± SD | CHI Mean Values of Symptoms * ± SD | HUE Mean Values of Symptoms * ± SD | NIR Mean Values of Symptoms * ± SD |
|---|---|---|---|---|
| 0 | 2.46 a ± 0.4 | 1.88 a ± 0.9 | 99.3 a ± 4.7 | 18,329.5 a ± 1112.2 |
| 0 + Alt. | 1 a ± 0.5 | 0.9 a ± 0.4 | 80.2 a ± 10.3 | 16,404.0 a ± 1199 |
| 0.15 + Alt. | 2.1 b ± 0.5 | 1.7 b ± 0.3 | 96.7 b ± 4.9 | 17,250.6 a ± 618.8 |
| 0.3 + Alt. | 1.7 ab ± 0.4 | 1.5 ab ± 0.3 | 93.6 b ± 4.1 | 17,414.1 a ± 410.7 |
| 0.6 + Alt. | 1.7 ab ± 0.7 | 1.4 ab ± 0.4 | 91.3 ab ± 9.5 | 17,160.6 a ± 723.7 |
| 0.9 + Alt. | 2.1 b ± 0.6 | 1.7 b ± 0.3 | 96.4 b ± 6.3 | 17,336.6 a ± 426.4 |
| PBA Concentration (%) | 0 | 0.025 | 0.05 | 0.1 | 0.15 |
|---|---|---|---|---|---|
| Mean value of symptoms area * ± SD | 29.8 e ± 0.9 | 2.8 a ± 0.2 | 5.5 b ± 0.5 | 9.6 c ± 0.6 | 17.5 d ± 1.7 |
| PBA Concentration (%) | ARI Mean Value of Symptoms * ± SD | CHI Mean Value of Symptoms * ± SD | HUE Mean Value of Symptoms * ± SD | NIR Mean Value of Symptoms * ± SD |
|---|---|---|---|---|
| 0 | 2.46 a ± 0.4 | 1.88 a ± 0.9 | 99.3 a ± 4.7 | 18,329.5 a ± 1112.2 |
| 0 + Alt. | 1.0 a ± 0.5 | 0.9 a ± 0.4 | 80.2 a ± 10.3 | 16,404.0 a ± 1199.0 |
| 0.025 + Alt. | 2.2 b ± 0.6 | 1.8 b ± 0.3 | 98.7 b ± 4.4 | 17,504.8 ab ± 237.1 |
| 0.05 + Alt. | 2.0 b ± 0.6 | 1.7 b ± 0.3 | 96.6 b ± 6.2 | 17,832.1 b ± 390.4 |
| 0.1 + Alt. | 2.2 b ± 0.6 | 1.8 b ± 0.3 | 99.0 b ± 5.1 | 17,450.0 ab ± 763.6 |
| 0.15 + Alt. | 2.4 b ± 0.6 | 1.8 b ± 0.3 | 99.4 b ± 4.9 | 17,376.0 ab ± 719.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinko, K.; Ivanković, S.; Lazarević, B.; Đermić, E.; Đermić, D. Control of Early Blight Fungus (Alternaria alternata) in Tomato by Boric and Phenylboronic Acid. Antibiotics 2022, 11, 320. https://doi.org/10.3390/antibiotics11030320
Martinko K, Ivanković S, Lazarević B, Đermić E, Đermić D. Control of Early Blight Fungus (Alternaria alternata) in Tomato by Boric and Phenylboronic Acid. Antibiotics. 2022; 11(3):320. https://doi.org/10.3390/antibiotics11030320
Chicago/Turabian StyleMartinko, Katarina, Siniša Ivanković, Boris Lazarević, Edyta Đermić, and Damir Đermić. 2022. "Control of Early Blight Fungus (Alternaria alternata) in Tomato by Boric and Phenylboronic Acid" Antibiotics 11, no. 3: 320. https://doi.org/10.3390/antibiotics11030320
APA StyleMartinko, K., Ivanković, S., Lazarević, B., Đermić, E., & Đermić, D. (2022). Control of Early Blight Fungus (Alternaria alternata) in Tomato by Boric and Phenylboronic Acid. Antibiotics, 11(3), 320. https://doi.org/10.3390/antibiotics11030320