
Effects of Verapamil and Two Bisbenzylisoquinolines, Curine and Guattegaumerine Extracted from Isolona hexaloba, on the Inhibition of ABC Transporters from Pseudomonas aeruginosa

 , and
, and
Abstract
:1. Introduction
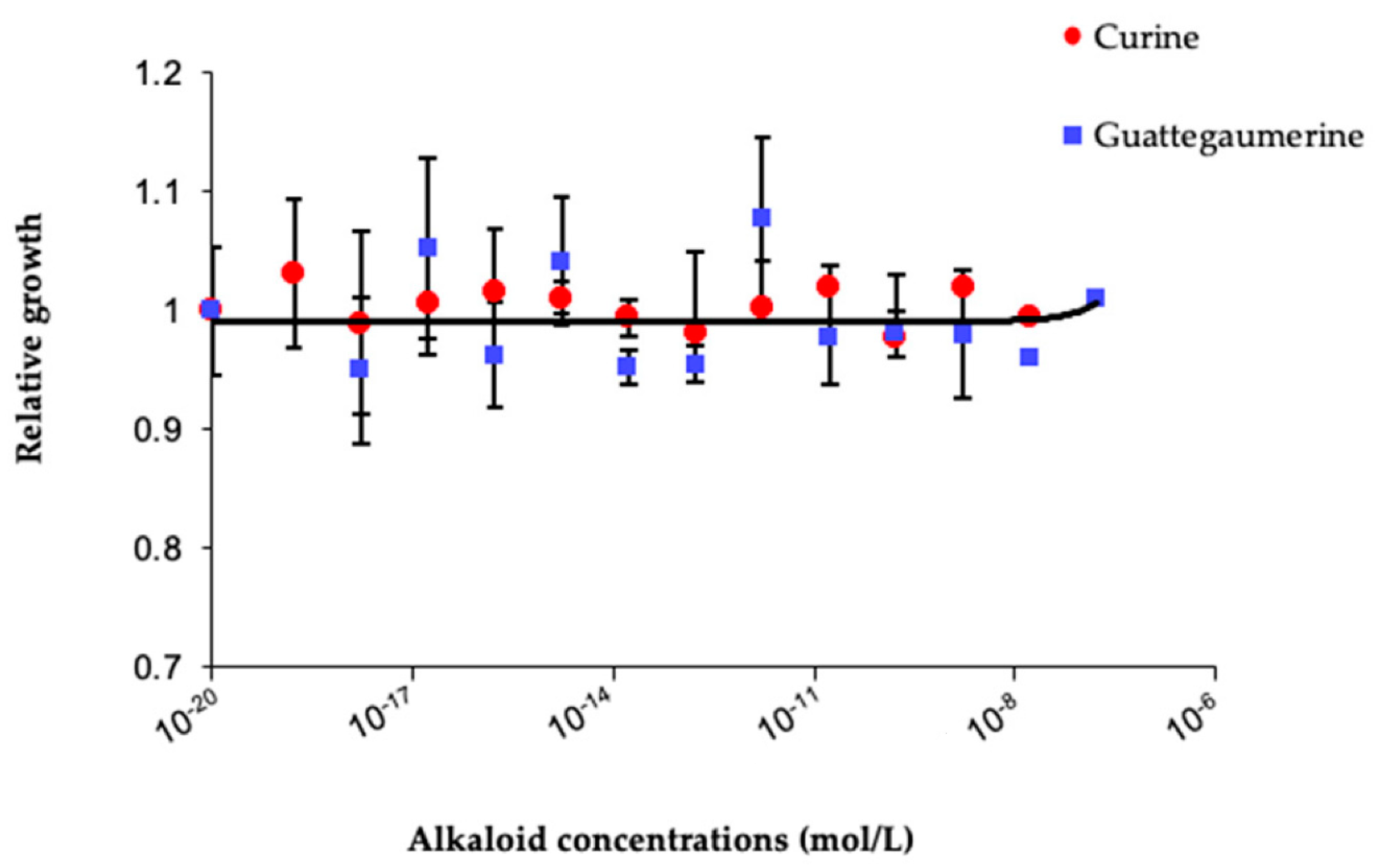
2. Results
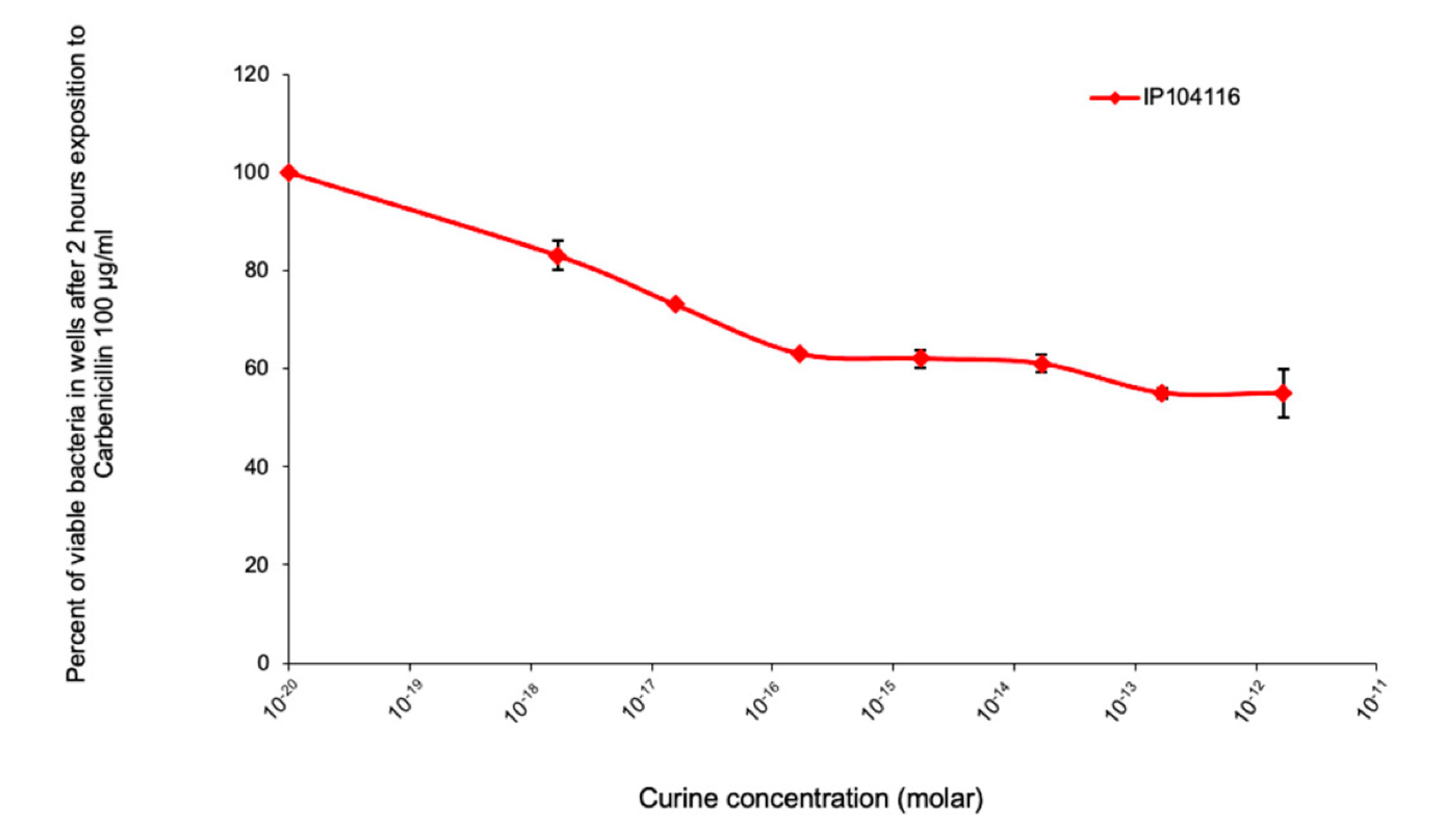
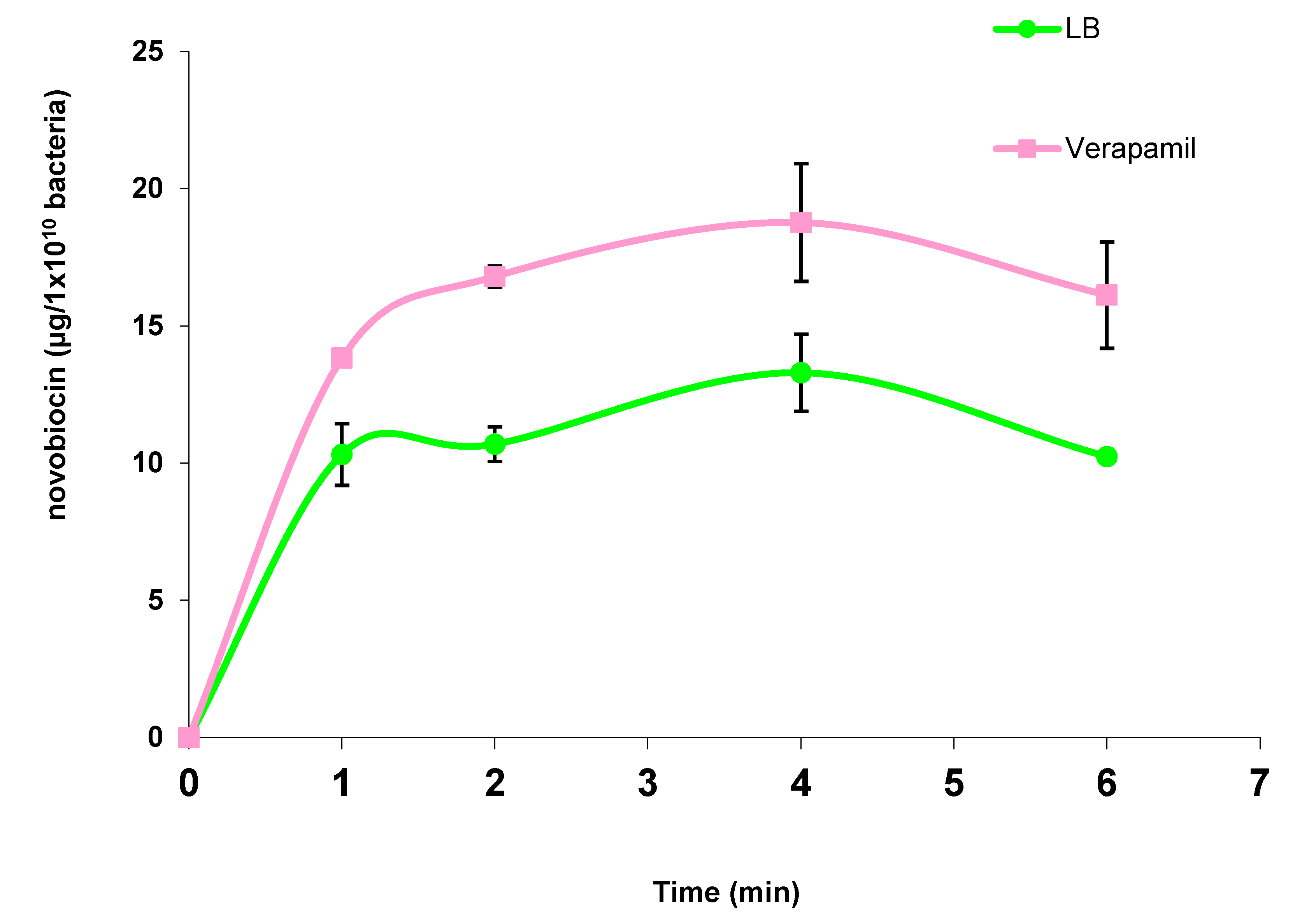
2.1. Evaluation of the Response of Pseudomonas Aeruginosa to Antibiotic Exposure in the Presence of Curine, Guattegaumerine, and Verapamil
- -
- Carbenicillin on periplasmic penicillin-binding proteins to block peptidoglycan synthesis;
- -
- Novobiocin on gyrase B-subunit by binding to the ATP-binding site to block DNA synthesis and transcription;
- -
- Erythromycin on the 50S ribosomal sub-unit to block protein synthesis.
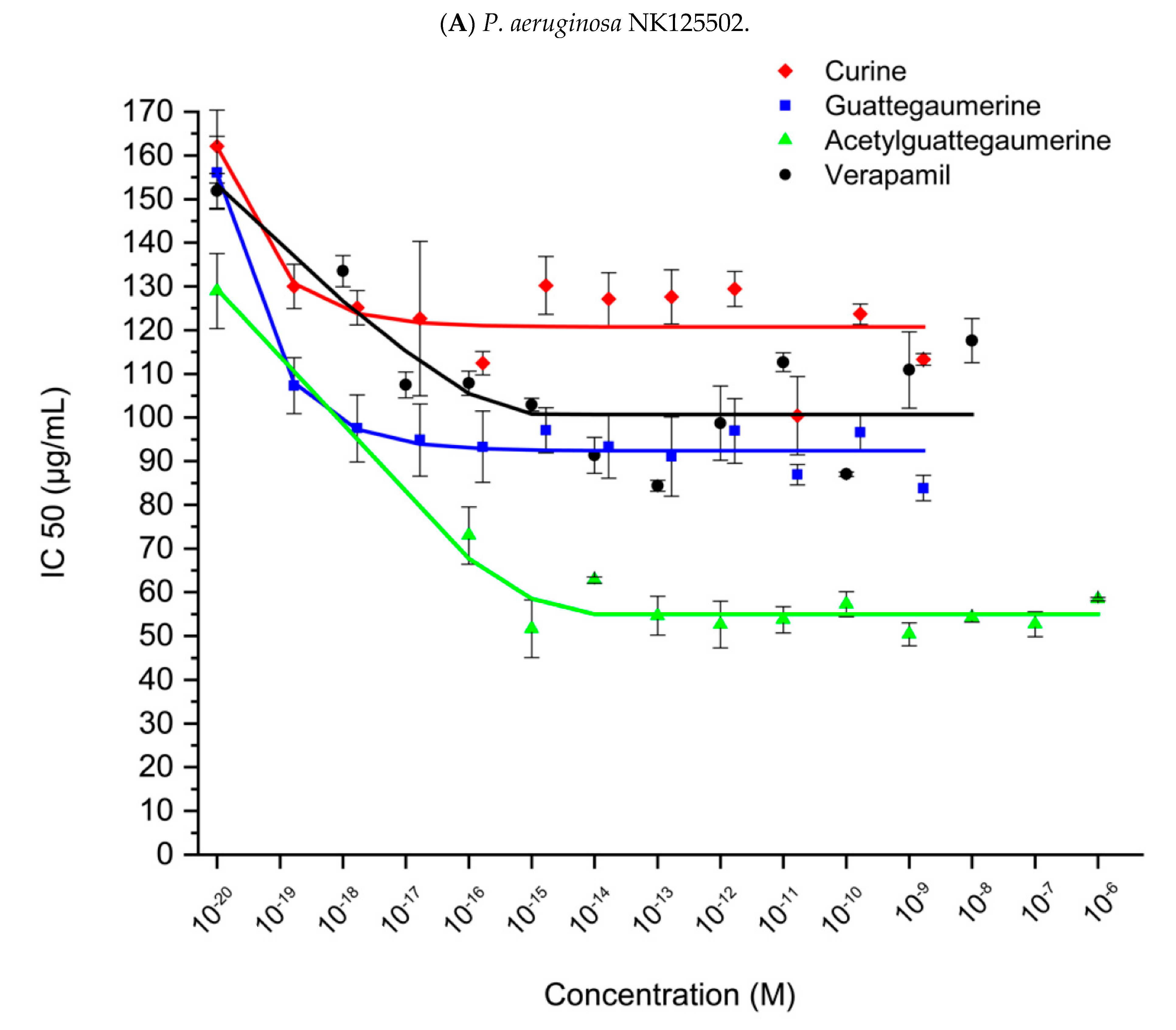
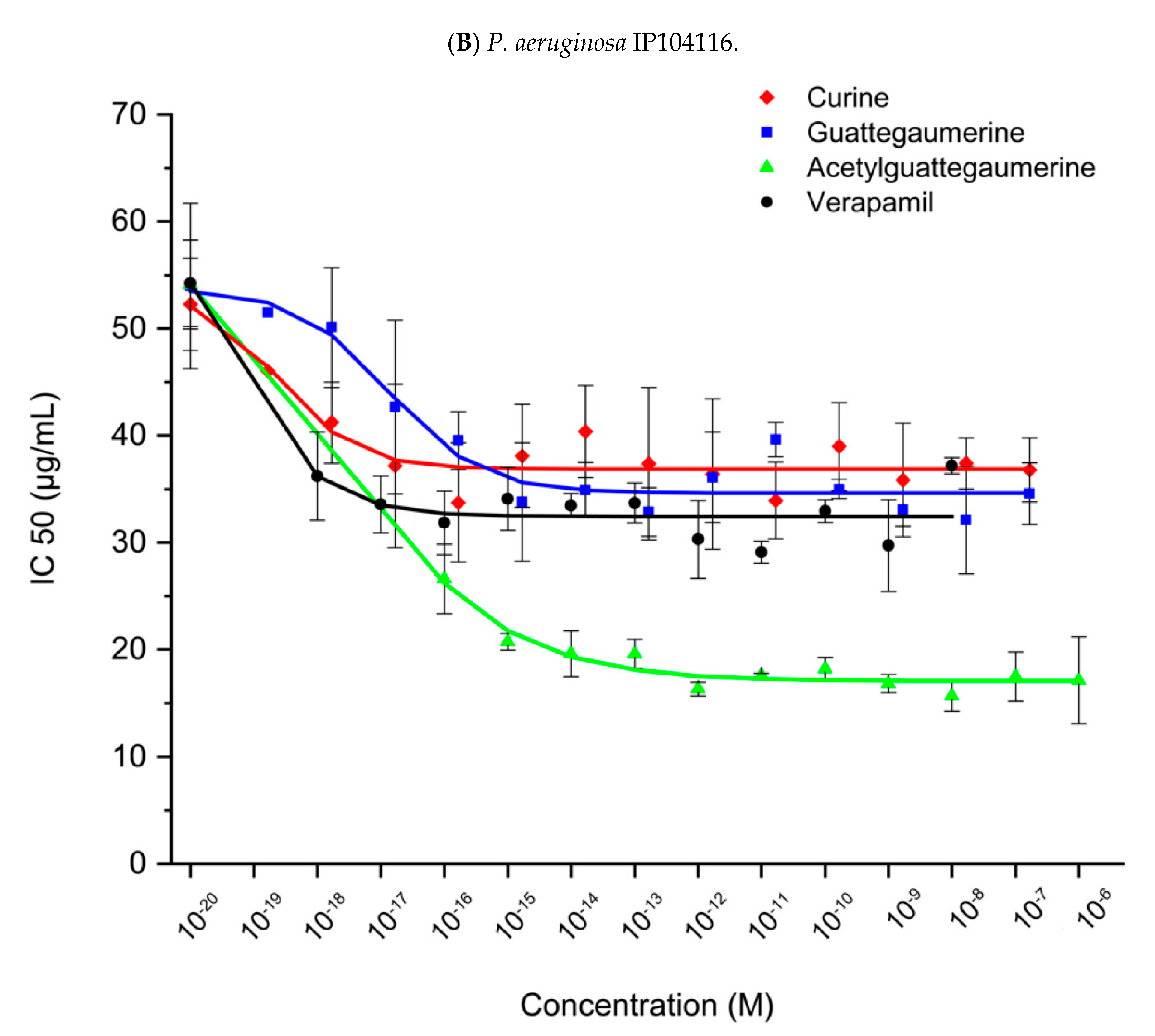
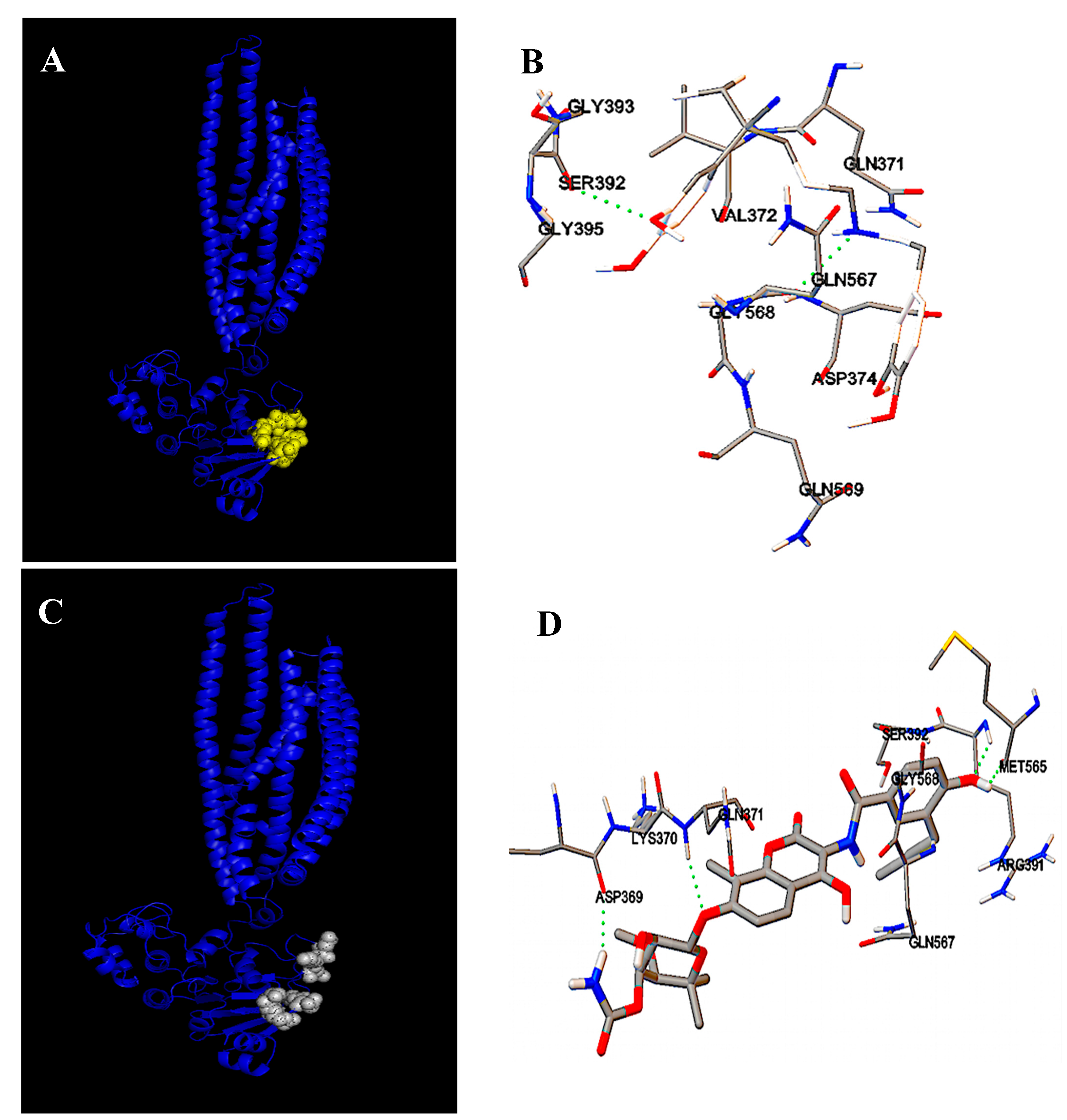
2.2. Identification of Carbenicillin and Novobiocin Transport Proteins in Pseudomonas Aeruginosa PAO1
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Chemicals
4.3. Alkaloidic Extraction and Isolation of the Bisbenzylisoquinolines
4.4. Triacetylguattegaumerine (3) Preparation
4.5. Molecular Structure Determination
4.6. Bacterial Strains
4.7. Sensitivity to Antibiotics
4.8. Kinetics of Novobiocin Accumulation
4.9. Molecular Modelization
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Govan, J.R.; Deretic, V. Microbial pathogenesis in cystic fibrosis; mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microbiol. Rev. 1996, 60, 539–574. [Google Scholar] [CrossRef] [PubMed]
- Mesaros, N.; Nordmann, P.; Plésiat, P.; Roussel-Delvallez, M.; Van Eldere, J.; Glupczynski, Y.; Van Laethem, Y.; Jacobs, F.; Lebecque, P.; Malfroot, A.; et al. Pseudomonas aeruginosa: Resistance and therapeutic options at the turn of the new millennium. Clin. Microbiol. Infect. 2007, 13, 560–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neu, H.C. The role of Pseudomonas aeruginosa in infections. J. Antimicrob. Chemother. 1983, 11, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacconelli, E.; Magrini, N.; Kahlmeter, G.; Singh, N. Global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. World Health Organ. 2017, 27, 318–327. [Google Scholar]
- Nasser, M.; Palwe, S.; Bhargava, R.N.; Feuilloley, M.G.J.; Kharat, A.S. Retrospective Analysis on Antimicrobial Resistance Trends and Prevalence of β-lactamases in Escherichia coli and ESKAPE Pathogens Isolated from Arabian Patients during 2000–2020. Microorganisms 2020, 8, 1626. [Google Scholar] [CrossRef]
- Bellido, F.; Martin, N.L.; Siehnel, R.J.; Hancock, R.E.W. Reevaluation using intact cells of the exclusion limit and role of porin OprF in Pseudomonas aeruginosa outer membrane permeability. J. Bacteriol. 1992, 174, 5196–5203. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, S.; Bouffartigues, E.; Bodilis, J.; Maillot, O.; Lesouhaitier, O.; Feuilloley, M.G.J.; Orange, N.; Dufour, A.; Cornelis, P. Structure, function and regulation of Pseudomonas aeruginosa porins. FEMS Microbiol. Rev. 2017, 41, 698–722. [Google Scholar] [CrossRef]
- Nikaido, H. Multidrug efflux pumps of gram-negative bacteria. J. Bacteriol. 1996, 178, 5853–5859. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Z.; Hiroshi, N.; Keith, P. Role of MexA-MexB-OprM in antibiotic efflux in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1995, 28, 630–635. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Z.; Barré, N.; Poole, K. Influence of the MexA-MexB-OprM multidrug efflux system on expression of the MexC-MexD-OprJ and MexE-MexF-OprN multidrug efflux systems in Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2000, 46, 885–893. [Google Scholar] [CrossRef]
- Hocquet, D.; Roussel-Delvallez, M.; Cavallo, J.D.; Plesiat, P. MexAB-OprM and MexXY-overproducing mutants are very prevalent among clinical strains of Pseudomonas aeruginosa with reduced susceptibility to tricarcillin. Antimicrob. Agents Chemother. 2007, 51, 1582–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakhtah, H.; Koyama, L.; Zhang, Y.; Morales, D.K.; Fields, B.L.; Price-whelan, A.; Hogan, D.A.; Shepard, K.; Dietrich, L.E.P. The Pseudomonas aeruginosa efflux pump MexGHI-OpmD transports a natural phenazine that controls gene expression and biofilm development. Proc. Natl. Acad. Sci. USA 2016, 113, E3538–E3547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.A.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial multidrug efflux pumps: Much more than antibiotic resistance determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, M.; Anju, C.P.; Biswas, L.; Anil Kumar, V.; Gopi Mohan, C. Antibiotic resistance in Pseudomonas aeruginosa and alternative therapeutic options. Int. J. Med. Microbiol. 2016, 306, 48–58. [Google Scholar] [CrossRef]
- Sergent, J.-A.; Mathouet, H.; Hulen, C.; Lameiras, P.; Marc Feuilloley, M.; Elomri, A.; Lomri, N. Effects of two natural bisbenzylisoquinolines, curine and guattegaumerine, extracted from Isolona hexaloba on rhodamine efflux by abcb1b from rat glycocholic acid resistant hepatocarcinoma cells. Molecules 2022, 27, 3030. [Google Scholar] [CrossRef]
- Fournet, A.; Ferreira, M.E.; de Arias, A.R.; Schinini, A.; Nakayama, H.; Torres, S.; Sanabria, R.; Guinaudeau, H.; Bruneton, J. The effect of bisbenzylisoquinoline alkaloids on Trypanosoma cruzi infections in mice. Int. J. Antimicrob. Agents 1997, 8, 163–170. [Google Scholar] [CrossRef]
- Marshall, S.J.; Russell, P.F.; Wright, C.W.; Anderson, M.M.; Phillipson, J.D.; Kirby, G.C.; Warhust, P.L.; Schiff, J.R. In vitro antiplasmodial, antiamoebic, and cytotoxic activities of a series of bisbenzylisoquinoline alkaloids. Antimicrob. Agents Chemother. 1994, 38, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Dias, C.; Barbosa-Filho, V.S.; Lemos, S.J.M.; Côrtes, S.F. Mechanisms involved in vasodilatator effect of curine in rat resistance arteries. Planta Med. 2002, 68, 1049–1051. [Google Scholar] [CrossRef]
- Dantas, B.B.; Faheina-Martins, G.V.; Coulidiati, T.H.; Barbosa-Bomfim, C.C.; da Silva Dias, C.; Barbosa-Filho, J.M.; Machado Araujo, D.M. Effects of curine in HL-60 leukemic cells: Cell cycle arrest and apoptosis induction. J. Nat. Med. 2015, 69, 218–223. [Google Scholar] [CrossRef]
- Ribeiro-Filho, J.; Piuvezam, M.R.; Bozza, P.T. Anti-allergic properties of curine, a bisbenzylisoquinoline alkaloid. Molecules 2015, 20, 4695–4707. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro-Filho, J.; Carvalho Leite, F.; Surrage Calheiros, A.; de Brito Carneiro, A.; Alves Azeredo, J.; Fernandes de Assis, E.; da Silva Dias, C.; Piuvezam, M.R.; Bozzal, P.T. Curine inhibits macrophage activation and neutrophil recruitment in a mouse model of Lipopoly-saccharide-induced inflammation. Toxins 2019, 11, 705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lijie, W.; Yan, S. Efflux mechanism and pathway of verapamil pumping by human P-glycoprotein. Arch. Biochem. Biophys. 2020, 696, 108675. [Google Scholar] [CrossRef]
- Leclercq, J.; Quetin, J.; De Pauw-Gillet, M.-C.; Bassleer, R.; Angenot, L. Antimitotic and cytotoxic activities of guattegaumerine, a bisbenzylisoquinoline alkaloid. Planta Med. 1987, 53, 116–117. [Google Scholar] [CrossRef] [PubMed]
- Lü, Q.; Xu, X.L.; He, Z.; Huang, X.J.; Guo, L.J.; Wang, H.X. Guattegaumerine protects primary cultured cortical neurons against oxidative stress injury induced by Hydrogen Peroxide concomitant with serum deprivation. Cell Mol. Neurobiol. 2009, 29, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Mathouet, H.; Cazoulat, I.; Oulyadi, H.; Seguin, E.; Daïch, A.; Elomri, A. Cazolobine, a new sesquiterpene from Isolona hexaloba (Annonaceae). Z. Naturforsch. B 2004, 59, 1118–1120. [Google Scholar] [CrossRef]
- Mathouet, H.; Elomri, A.; Lameiras, P.; Daïch, A.; Vérité, P. An alkaloid, two conjugate sesquiterpenes and phenylpropanoid from Pachypodanthium confine Engl. and Diels. Phytochemistry 2007, 68, 1813–1818. [Google Scholar] [CrossRef] [PubMed]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Substrate specificities of MexAB-OprM, MexCD-OprJ, and MexXY-OprM efflux pumps in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3322–3327. [Google Scholar] [CrossRef] [Green Version]
- Litman, T.; Skovsgoard, T.; Stein, W.D. Pumping of drugs by P-glycoprotein: A two-step process. J. Pharmacol. Exp. Ther. 2003, 307, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Van Veen, H.W.; Callaghan, R.; Soceneantu, L.; Sardini, A.; Konings, W.N.; Higgins, C.F. A bacterial antibiotic-resistant gene that complement the human multidrug-resistance P-glycoprotein gene. Nature 1998, 391, 291–295. [Google Scholar] [CrossRef]
- Orelle, C.; Dalmas, O.; Gros, P.; Di Pietro, A.; Jault, J.-M. The conserved glutamate residue adjacent to the Walker-B motif is the catalytic base for ATP hydrolysis in the ATP-binding cassette transporter BmrA. J. Biol. Chem. 2003, 278, 47002–47008. [Google Scholar] [CrossRef] [Green Version]
- Uwate, M.; Ichise, Y.-K.; Shirai, A.; Omasa, T.; Nakae, T.; Maseda, H. Two routes of MexS-MexT-mediated regulation of MexEF-OprN and MexAB-OprM efflux pump expression in Pseudomonas aeruginosa. Microbiol. Immunol. 2013, 57, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Hulen, C.; Racine, P.-J.; Chevalier, S.; Feuilloley, M.; Lomri, N. Identification of the PA1113 gene product as an ABC transporter involved in the uptake of carbenicillin in Pseudomonas aeruginosa PAO1. Antibiotics 2020, 9, 596. [Google Scholar] [CrossRef] [PubMed]
- Ghanei, H.; Abeyrathme, P.D.; Lam, J.S. Biochemical characterization of MsbA from Pseudomonas aeruginosa. J. Biol. Chem. 2007, 282, 26939–26947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padayatti, P.S.; Lee, S.C.; Stanfield, R.L.; Wen, P.-C.; Tajkhorshid, E.; Wilson, I.A.; Zhang, Q. Structural Insights into the Lipid A Transport Pathway in MsbA. Structure 2019, 27, 1114–1123. [Google Scholar] [CrossRef]
- Reuter, G.; Janvilisri, T.; Venter, H.; Shahi, S.; Balakrishnan, L.; van Veen, H.W. The ATP binding cassette multidrug transporter LmrA and lipid transporter MsbA have overlapping substrate specificities. J. Biol. Chem. 2003, 278, 35193–35198. [Google Scholar] [CrossRef] [Green Version]
- Hocquemiller, R.; Cabalion, P.; Fournet, A.; Cave, A. Alcaloïdes des Annonacées XIIX: Alcaloïdes d’Isolona hexaloba Engler & Diels, I. zenkeri Engler and Diels et I. pilosa Diels. Planta Med. 1984, 50, 23–25. [Google Scholar]
- Mao, W.; Warren, M.S.; Black, D.S.; Satou, T.; Murata, T.; Nishino, T.; Gotoh, N.; Lomovskaya, O. On the mechanism of substrate specificity by resistance nodulation division (RND)-type multidrug resistance pumps: The large periplasmic loops of MexD from Pseudomonas aeruginosa are involved in substrate recognition. Mol. Microbiol. 2002, 46, 889–901. [Google Scholar] [CrossRef]
- Van Veen, H.W.; Venema, K.; Bolhuis, H.; Oussenko, I.; Kok, J.; Poolman, B.; Driessen, A.J.M.; Konings, W.M. Multidrug resistance mediated by a bacterial homolog of the human transporter MDR1. Proc. Natl. Acad. Sci. USA 1996, 93, 10668–10672. [Google Scholar] [CrossRef] [Green Version]
- Orelle, C.; Gubellini, F.; Durand, A.; Marco, S.; Levy, D.; Gros, P.; Di Pietro, A.; Jault, J.-M. Conformational change induced by ATP binding in the multidrug ATP-binding cassette transporter BmrA. Biochemistry 2008, 47, 2404–2412. [Google Scholar] [CrossRef]
- Woebking, B.; Reuter, G.; Shilling, R.A.; Velamakanni, S.; Shahi, S.; Venter, H.; Balakrishnan, L.; van Veen, H.W. Drug-lipid A interactions on the Escherichia coli ABC transporter MsbA. J. Bacteriol. 2005, 187, 6363–6369. [Google Scholar] [CrossRef] [Green Version]
- Smriti, Z.P.; Mchaourab, H.S. Mapping daunorubicin-binding sites in the ATP-binding cassette transporter MsbA using site-specific quenching by spin labels. J. Biol. Chem. 2009, 284, 13904–13913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doshi, R.; van Veen, H.W. Substrate binding stabilizes a pre-translocation intermediate in the ATP-binding cassette transport protein MsbA. J. Biol. Chem. 2013, 288, 21638–21647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, J.M.; Owens, T.W.; Mandler, M.D.; Simpson, B.W.; Lazarusa, M.B.; Shermana, D.J.; Davis, R.M.; Okuda, S.; Massefski, W.; Ruiz, N.; et al. The antibiotic novobiocin binds and activates the ATPase that powers lipopolysaccharide transport. J. Am. Chem. Soc. 2017, 139, 17221–17224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldas, J.; Porter, Q.N.; Bick, I.R.C.; Vernengo, M.J. Some observations on the mass-spectrometry of bisbenzylisoquinoline alkaloids. Tetrahedron Lett. 1966, 19, 2059–2068. [Google Scholar] [CrossRef]
- Dehaussy, H.; Tits, M.; Angenot, L. Guattegaumerine, new bisbenzylisoquinoline alkaloid from Guatteria gaumeri Greenm. Planta Med. 1983, 49, 25–27. [Google Scholar] [CrossRef]
- Källberg, M.; Wang, H.; Wang, S.; Peng, J.; Wang, Z.; Lu, H.; Xu, J. Template-based protein structure modeling using the RaptorX web server. Nat. Protoc. 2012, 7, 1511–1522. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A Programming Language for Software Integration and Development. J. Mol. Graphics Mod. 1999, 17, 57–61. [Google Scholar]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Autodock4 and AutoDockTools4: Automated docking with selective receptor flexiblity. J. Comput. Chem. 2009, 16, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Mahamoud, A.; Chevalier, J.; Alibert-Franco, S.; Kern, W.V.; Pagès, J.-M. Antibiotic efflux pumps in Gram-negative bacteria: The inhibitor response strategy. J. Antimicrob. Chemother. 2007, 59, 1223–1229. [Google Scholar] [CrossRef]
- Zechini, B.; Versace, I. Inhibitors of multidrug resistant efflux systems in bacteria. Recent Pat. Anti-Infect. Drug Discov. 2009, 4, 37–50. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC 50 (µg/mL) | |||
|---|---|---|---|
| Strain IP 104116 | Carbenicillin | Novobiocin | Erythromycin |
| Mean ± SD, (p-Value) | |||
| None Curine Guattegaumerine Triacetylguattegaumerine Verapamil | 58 ± 3.5 37 ± 1.5, (p = 0.038) 42 ± 1, (p = 0.01) 34 ± 1, (p = 0.02) 35 ± 3.5, (p = 0.052) | 289 ± 3.5 172.52 ± 4.94, (p = 0.14) 180.74 ± 7.28, (p = 0.16) 208.54 ± 2.43, (p = 0.09) 191.94 ± 2.26, (p = 0.19) | 40 ± 4 29.86 ± 1.74, (p = 0.18) 22.39 ± 1.47, (p = 0.22) 29.85 ± 2.48, (p = 0.19) 30.60 ± 0.49, (p = 0.17) |
| IC 50 (µg/mL) | |||
|---|---|---|---|
| Strain NK 125502 | Carbenicillin | Novobiocin | Erythromycin |
| Mean ± SD, p-Value | |||
| None Curine Guattegaumerine Triacetylguattegaumerine Verapamil | 154.61 ± 13.5 105.27 ± 9.8, (p = 0.004) 95.79 ± 5.36, (p = 0.122) 82.06 ± 6.8, (p = 0.075) 101.05 ± 5.48, (p = 0.063) | 24.77 ± 1.54 19.85 ± 0.93, (p = 0.13) 17.76 ± 2.18, (p = 0.12) 20.70 ± 0.91, (p = 0.16) 19.11 ± 0.90, (p = 0.11) | 38.95 ± 3.43 38.16 ± 1.23, (p = 0.01) 38.80 ± 0.55, (p = 0.03) 36.82 ± 1.77, (p = 0.05) 39.81 ± 1.81, (p = 0.03) |
| Carbenicillin IC 50 (µg/mL) | Novobiocin IC 50 (µg/mL) | ||||
|---|---|---|---|---|---|
| Strain | Gene | None | Verapamil | None | Verapamil |
| Mean ± SD | Mean ± SD, p-Value | Mean ± SD | Mean ± SD, p-Value | ||
| PAO1 Washigton | 29 ± 3.5 | 21 ± 2, (p = 0.05) | 193 ± 3 | 166 ± 2, (p = 0.002) | |
| PW2567 | PA0860 | nd | nd | 267 ± 12 | 231 ± 9, (p = 0.015) |
| PW8020 | PA4143 | 42 ± 2 | 23 ± 1, (p = 0.08) | 180 ± 11.5 | 158.5 ± 6.5, (p= 0.002) |
| PW6408 | PA3228 | 55 ± 4 | 39 ± 4, (p = 0.09) | 249 ± 8 | 210 ± 5.5, (p = 0.09) |
| PW3010 | PA1113 | 54 ± 2 | 54 ± 1.5, (p = 0.01) | 146.5 ± 3.5 | 127 ± 3.5, (p = 0.10) |
| Percentage of Identity | |||
|---|---|---|---|
| LmrA (Lactococcus lactis) | MsbA (E. Coli) | MsbA (P. aeruginosa) | |
| LmrA (Lactococcus lactis) | 100 | 75.18 | 77.33 |
| MsbA (E.Coli) | 75.18 | 100 | 76 |
| MsbA (P. aeruginosa) | 77.33 | 76 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hulen, C.; Racine, P.-J.; Feuilloley, M.; Elomri, A.; Lomri, N.-E. Effects of Verapamil and Two Bisbenzylisoquinolines, Curine and Guattegaumerine Extracted from Isolona hexaloba, on the Inhibition of ABC Transporters from Pseudomonas aeruginosa. Antibiotics 2022, 11, 700. https://doi.org/10.3390/antibiotics11050700
Hulen C, Racine P-J, Feuilloley M, Elomri A, Lomri N-E. Effects of Verapamil and Two Bisbenzylisoquinolines, Curine and Guattegaumerine Extracted from Isolona hexaloba, on the Inhibition of ABC Transporters from Pseudomonas aeruginosa. Antibiotics. 2022; 11(5):700. https://doi.org/10.3390/antibiotics11050700
Chicago/Turabian StyleHulen, Christian, Pierre-Jean Racine, Marc Feuilloley, Abdelhakim Elomri, and Nour-Eddine Lomri. 2022. "Effects of Verapamil and Two Bisbenzylisoquinolines, Curine and Guattegaumerine Extracted from Isolona hexaloba, on the Inhibition of ABC Transporters from Pseudomonas aeruginosa" Antibiotics 11, no. 5: 700. https://doi.org/10.3390/antibiotics11050700
APA StyleHulen, C., Racine, P. -J., Feuilloley, M., Elomri, A., & Lomri, N. -E. (2022). Effects of Verapamil and Two Bisbenzylisoquinolines, Curine and Guattegaumerine Extracted from Isolona hexaloba, on the Inhibition of ABC Transporters from Pseudomonas aeruginosa. Antibiotics, 11(5), 700. https://doi.org/10.3390/antibiotics11050700