
Ethyl Acetate Fraction of Bixa orellana and Its Component Ellagic Acid Exert Antibacterial and Anti-Inflammatory Properties against Mycobacterium abscessus subsp. massiliense

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Identification of Phytochemical Compounds–Profile of Fraction Groups
2.2. In Silico Analysis of the Biological Activities of ElAc and Prediction of Its Toxic Effects, Including Hepatotoxicity
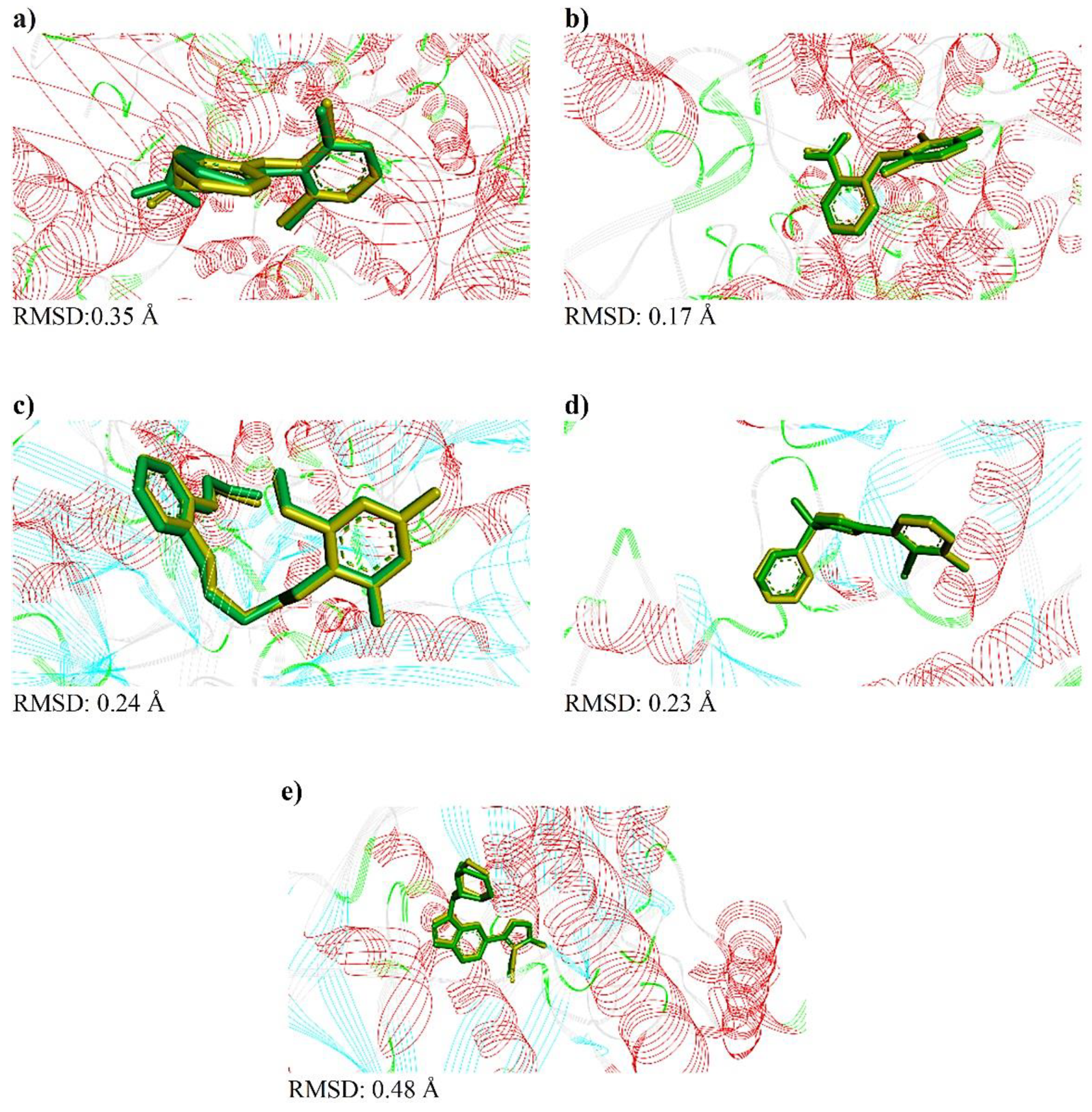
2.3. Molecular Docking
2.4. Antibacterial Activity of BoEA and ElAc
2.5. Time–Kill Assay
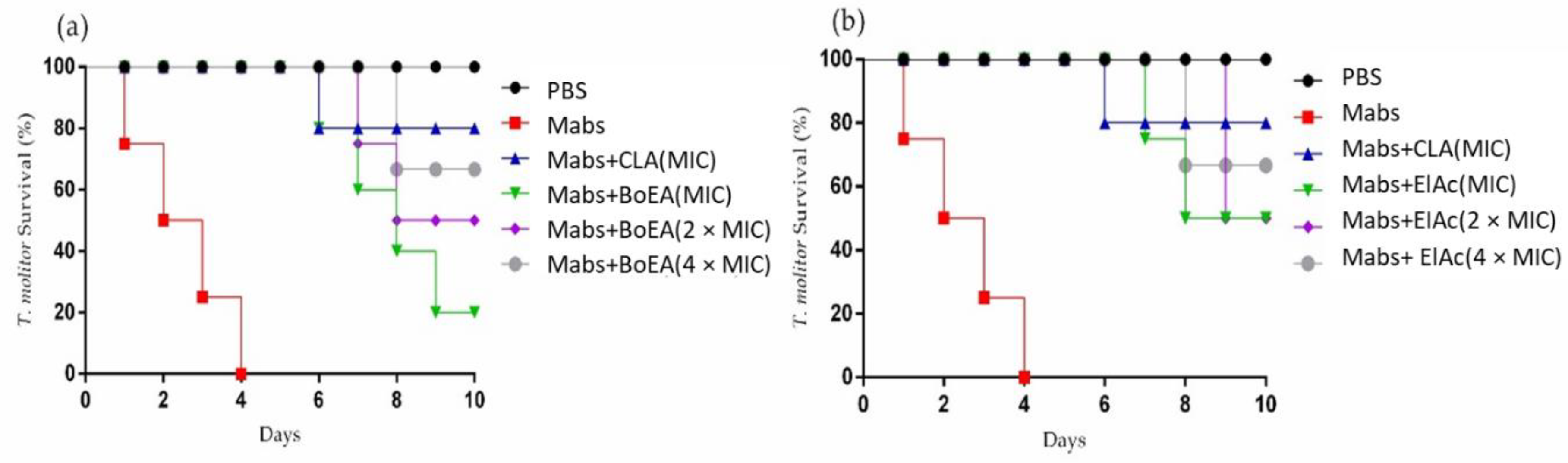
2.6. BoEA and ElAc Increase Survival of Tenebrio molitor Infected with Mabs
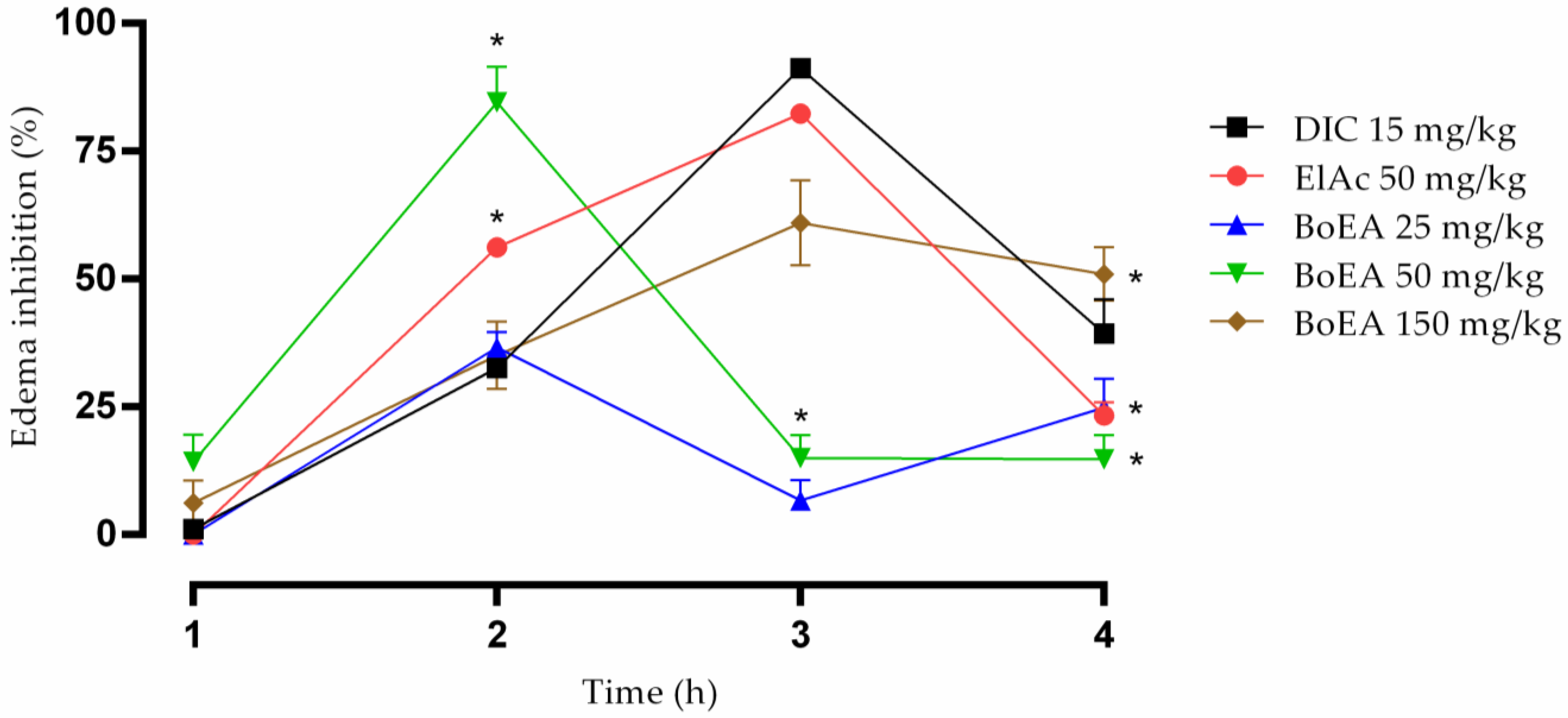
2.7. Anti-Inflammatory Activity of BoEA and ElAc
3. Discussion
4. Materials and Methods
4.1. Drugs, Chemicals, and Reagents
4.2. Preparation of BoEA and Isolation/Identification of ElAc
4.2.1. Collection of B. orellana Leaves and Separation of Fractions
4.2.2. Isolation and Characterization of Compounds in BoEA
4.3. In Silico Analysis
4.3.1. Prediction of Biological Activities of ElAc
4.3.2. Prediction of Pharmacokinetic Characteristics and Toxic Effects of ElAc
4.3.3. Molecular Docking Study
4.4. In Vitro Analysis
4.4.1. Strain
4.4.2. Determining MIC and MBC
4.4.3. Time–Kill Assay
4.5. In Vivo Analysis
4.5.1. T. molitor Survival Assay
4.5.2. Carrageenan-Induced Paw Edema Model
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Degiacomi, G.; Sammartino, J.C.; Chiarelli, L.R.; Riabova, O.; Makarov, V.; Pasca, M.R. Mycobacterium Abscessus, an Emerging and Worrisome Pathogen among Cystic Fibrosis Patients. Int. J. Mol. Sci. 2019, 20, 5868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudehen, Y.M.; Kremer, L. Mycobacterium Abscessus. Trends Microbiol. 2021, 29, 951–952. [Google Scholar] [CrossRef] [PubMed]
- Johansen, M.D.; Herrmann, J.L.; Kremer, L. Non-Tuberculous Mycobacteria and the Rise of Mycobacterium Abscessus. Nat. Rev. Microbiol. 2020, 18, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Strnad, L.; Winthrop, K.L. Treatment of Mycobacterium Abscessus Complex. Semin. Respir. Crit. Care Med. 2018, 39, 362–376. [Google Scholar] [CrossRef]
- Nasiri, M.J.; Haeili, M.; Ghazi, M.; Goudarzi, H.; Pormohammad, A.; Fooladi, A.A.I.; Feizabadi, M.M. New Insights in to the Intrinsic and Acquired Drug Resistance Mechanisms in Mycobacteria. Front. Microbiol. 2017, 8, 681. [Google Scholar] [CrossRef] [Green Version]
- Meir, M.; Barkan, D. Alternative and Experimental Therapies of Mycobacterium Abscessus Infections. Int. J. Mol. Sci. 2020, 21, 6793. [Google Scholar] [CrossRef]
- Victoria, L.; Gupta, A.; Gómez, J.L.; Robledo, J. Mycobacterium Abscessus Complex: A Review of Recent Developments in an Emerging Pathogen. Front. Cell. Infect. Microbiol. 2021, 11, 659997. [Google Scholar] [CrossRef]
- Liu, Q.; Meng, X.; Li, Y.; Zhao, C.N.; Tang, G.Y.; Li, H. Bin Antibacterial and Antifungal Activities of Spices. Int. J. Mol. Sci. 2017, 18, 1283. [Google Scholar] [CrossRef] [Green Version]
- Tran, N.; Pham, B.; Le, L. Bioactive Compounds in Anti-Diabetic Plants: From Herbal Medicine to Modern Drug Discovery. Biology 2020, 9, 252. [Google Scholar] [CrossRef]
- Santos, D.C.; Barboza, A.S.; Ribeiro, J.S.; Rodrigues Junior, S.A.; Campos, Â.D.; Lund, R.G. Bixa Orellana L. (Achiote, Annatto) as an Antimicrobial Agent: A Scoping Review of Its Efficiency and Technological Prospecting. J. Ethnopharmacol. 2022, 287, 114961. [Google Scholar] [CrossRef]
- Sousa-Andrade, L.M.; Oliveira, A.B.M.; Leal, A.L.A.B.; Oliveira, F.A.A.; Portela, A.L.; Lima-Neto, J.S.; Siqueira-Júnior, J.P.; Kaatz, G.W.; Rocha, C.Q.; Barreto, H.M. Antimicrobial Activity and Inhibition of the NorA Efflux Pump of Staphylococcus Aureus by Extract and Isolated Compounds from Arrabidaea Brachypoda. Microb. Pathog. 2020, 140, 103935. [Google Scholar] [CrossRef] [PubMed]
- Vilar, D.D.A.; Vilar, M.S.D.A.; Moura, T.F.A.D.L.E.; Raffin, F.N.; Oliveira, M.R.D.; Franco, C.F.D.O.; De Athayde-Filho, P.F.; Diniz, M.D.F.F.M.; Barbosa-Filho, J.M. Traditional Uses, Chemical Constituents, and Biological Activities of Bixa Orellana L.: A Review. Sci. World J. 2014, 2014, 857292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-Flores, D.; Ulloa-Urizar, G.; Camere-Colarossi, R.; Caballero-García, S.; Mayta-Tovalino, F.; del Valle-Mendoza, J. Antibacterial Activity of Bixa Orellana L. (Achiote) against Streptococcus Mutans and Streptococcus Sanguinis. Asian Pac. J. Trop. Biomed. 2016, 6, 400–403. [Google Scholar] [CrossRef] [Green Version]
- Lima-Viana, J.; Zagmignan, A.; Lobato, L.F.L.; Abreu, A.G.; Nascimento da Silva, L.C.; Cortez-Sá, J.; Monteiro, C.A.; Lago, J.H.G.; Gonçalves, L.M.; Carvalho, R.C.; et al. Hydroalcoholic Extract and Ethyl Acetate Fraction of Bixa Orellana Leaves Decrease the Inflammatory Response to Mycobacterium Abscessus Subsp. Massiliense. Evid. -Based Complement. Altern. Med. 2018, 2018, 6091934. [Google Scholar] [CrossRef] [Green Version]
- Moraes-Neto, R.N.; Coutinho, G.G.; Oliveira Rezende, A.; Pontes, D.B.; Ferreira, R.L.P.S.; Morais, D.A.; Albuquerque, R.P.; Lima-Neto, L.G.; Nascimento-Silva, L.C.; Rocha, C.Q.; et al. Compounds Isolated from Bixa Orellana: Evidence-Based Advances to Treat Infectious Diseases. Rev. Colomb. Cienc. Quím. Farm 2020, 49, 581–601. [Google Scholar] [CrossRef]
- Alfei, S.; Turrini, F.; Catena, S.; Zunin, P.; Grilli, M.; Pittaluga, A.M.; Boggia, R. Ellagic Acid a Multi-Target Bioactive Compound for Drug Discovery in CNS? A Narrative Review. Eur. J. Med. Chem. 2019, 183, 111724. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Castillo, C.M.S.; Caroca, R.; Lazo-Vélez, M.A.; Antonyak, H.; Polishchuk, A.; Lysiuk, R.; Oliinyk, P.; De Masi, L.; et al. Ellagic Acid: A Review on Its Natural Sources, Chemical Stability, and Therapeutic Potential. Oxid. Med. Cell. Longev. 2022, 2022, 1–24. [Google Scholar] [CrossRef]
- Zagminan, A.; Viana, J.; Dutra, R.; Pinheiro, A.; Carvalho, L.; Paschoal, M.A.; Falcai, A.; Lima-Neto, L.; Gonçalves, L.; Sousa, E. Effect of Ethyl Acetate Fraction of Urucum Extract on Cell Viability of Peripheral Blood Mononuclear Cells and Antimicrobial Activity in Human Pathogenic Bacteria. In Proceedings of the 11th Congress of the Latin American Association of Immunology, Medellin, Colombia, 13–16 October 2015; Volume 6. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Zhao, L.; Mehmood, A.; Soliman, M.M.; Iftikhar, A.; Iftikhar, M.; Aboelenin, S.M.; Wang, C. Protective Effects of Ellagic Acid Against Alcoholic Liver Disease in Mice. Front. Nutr. 2021, 8, 744520. [Google Scholar] [CrossRef]
- Ramos, R.S.; Borges, R.S.; de Souza, J.S.N.; Araujo, I.F.; Chaves, M.H.; Santos, C.B.R. Identification of Potential Antiviral Inhibitors from Hydroxychloroquine and 1,2,4,5-Tetraoxanes Analogues and Investigation of the Mechanism of Action in SARS-CoV-2. Int. J. Mol. Sci. 2022, 23, 1781. [Google Scholar] [CrossRef]
- BIOVIA Discovery Studio-BIOVIA-Dassault Systèmes®. Available online: https://www.3ds.com/products-services/biovia/products/molecular-modeling-simulation/biovia-discovery-studio/ (accessed on 11 June 2022).
- Rowlinson, S.W.; Kiefer, J.R.; Prusakiewicz, J.J.; Pawlitz, J.L.; Kozak, K.R.; Kalgutkar, A.S.; Stallings, W.C.; Kurumbail, R.G.; Marnett, L.J. A Novel Mechanism of Cyclooxygenase-2 Inhibition Involving Interactions with Ser-530 and Tyr-385. J. Biol. Chem. 2003, 278, 45763–45769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlando, B.J.; Malkowski, M.G. Substrate-Selective Inhibition of Cyclooxygeanse-2 by Fenamic Acid Derivatives Is Dependent on Peroxide Tone. J. Biol. Chem. 2016, 291, 15069–15081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RCSB PDB—7K6C: Crystal Structure of Dihydrofolate Reductase (DHFR) from Mycobacterium Abscessus ATCC 19977/DSM 44196 with NADP and Inhibitor P218. Available online: https://www.rcsb.org/structure/7K6C (accessed on 11 June 2022).
- Charoensutthivarakul, S.; Thomas, S.E.; Curran, A.; Brown, K.P.; Belardinelli, J.M.; Whitehouse, A.J.; Acebrón-García-de-Eulate, M.; Sangan, J.; Gramani, S.G.; Jackson, M.; et al. Development of Inhibitors of SAICAR Synthetase (PurC) from Mycobacterium Abscessus Using a Fragment-Based Approach. ACS Infect. Dis. 2022, 8, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, A.J.; Thomas, S.E.; Brown, K.P.; Fanourakis, A.; Chan, D.S.-H.; Libardo, M.D.J.; Mendes, V.; Boshoff, H.I.M.; Floto, R.A.; Abell, C.; et al. Development of Inhibitors against Mycobacterium Abscessus TRNA (m 1 G37) Methyltransferase (TrmD) Using Fragment-Based Approaches. J. Med. Chem. 2019, 62, 7210–7232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, I.A.; Barreto, A.M.S.; Marinho, D.; Krempser, E.; Kuenemann, M.A.; Sperandio, O.; Dardenne, L.E.; Miteva, M.A. New Machine Learning and Physics-Based Scoring Functions for Drug Discovery. Sci. Rep. 2021, 11, 3198. [Google Scholar] [CrossRef]
- Djedjibegovic, J.; Marjanovic, A.; Panieri, E.; Saso, L. Ellagic Acid-Derived Urolithins as Modulators of Oxidative Stress. Oxid. Med. Cell. Longev. 2020, 2020, 5194508. [Google Scholar] [CrossRef]
- Aslan, A.; Hussein, Y.T.; Gok, O.; Beyaz, S.; Erman, O.; Baspinar, S. Ellagic Acid Ameliorates Lung Damage in Rats via Modulating Antioxidant Activities, Inhibitory Effects on Inflammatory Mediators and Apoptosis-Inducing Activities. Environ. Sci. Pollut. Res. 2020, 27, 7526–7537. [Google Scholar] [CrossRef]
- Kiela, P.R.; Ghishan, F.K. Physiology of Intestinal Absorption and Secretion. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 145–159. [Google Scholar] [CrossRef] [Green Version]
- Baby, J.; Devan, A.R.; Kumar, A.R.; Gorantla, J.N.; Nair, B.; Aishwarya, T.S.; Nath, L.R. Cogent Role of Flavonoids as Key Orchestrators of Chemoprevention of Hepatocellular Carcinoma: A Review. J. Food Biochem. 2021, 45, e13761. [Google Scholar] [CrossRef]
- Montenegro, C.; Gonçalves, G.; Oliveira Filho, A.; Lira, A.; Cassiano, T.; Lima, N.; Barbosa-Filho, J.; Diniz, M.; Pessôa, H. In Silico Study and Bioprospection of the Antibacterial and Antioxidant Effects of Flavone and Its Hydroxylated Derivatives. Molecules 2017, 22, 869. [Google Scholar] [CrossRef] [Green Version]
- Gowthaman, U.; Jayakanthan, M.; Sundar, D. Molecular Docking Studies of Dithionitrobenzoic Acid and Its Related Compounds to Protein Disulfide Isomerase: Computational Screening of Inhibitors to HIV-1 Entry. BMC Bioinform. 2008, 9, S14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hevener, K.E.; Zhao, W.; Ball, D.M.; Babaoglu, K.; Qi, J.; White, S.W.; Lee, R.E. Validation of Molecular Docking Programs for Virtual Screening against Dihydropteroate Synthase. J. Chem. Inf. Model. 2009, 49, 444–460. [Google Scholar] [CrossRef] [PubMed]
- Raju, A.; Kulkarni, S.; Ray, M.K.; Rajan, M.G.R.; Degani, M.S. E84G Mutation in Dihydrofolate Reductase from Drug Resistant Strains of Mycobacterium Tuberculosis (Mumbai, India) Leads to Increased Interaction with Trimethoprim. Int. J. Mycobacteriol. 2015, 4, 97–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alim, S.; Bairagi, N.; Shahriyar, S.; Masnoon Kabir, M.; Habibur Rahman, M.; Sayeed Shahriyar, C. In Vitro Antibacterial Potential of Bixa Orellana L. against Some Pathogenic Bacteria and Comparative Investigation on Some Standard Antibiotics. J. Pharmacogn. Phytochem. 2016, 5, 178–181. [Google Scholar]
- Poma-Castillo, L.; Espinoza-Poma, M.; Mauricio, F.; Mauricio-Vilchez, C.; Alvítez-Temoche, D.; Mayta-Tovalino, F. Antifungal Activity of Ethanol-Extracted Bixa Orellana (L) (Achiote) on Candida Albicans, at Six Different Concentrations. J. Contemp. Dent. Pract. 2019, 20, 1159–1163. [Google Scholar] [CrossRef] [PubMed]
- Machín, L.; Tamargo, B.; Piñón, A.; Atíes, R.C.; Scull, R.; Setzer, W.N.; Monzote, L. Bixa Orellana L. (Bixaceae) and Dysphania Ambrosioides (L.) Mosyakin & Clemants (Amaranthaceae) Essential Oils Formulated in Nanocochleates against Leishmania Amazonensis. Molecules 2019, 24, 4222. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Calienes Valdés, A.; Mendiola Martínez, J.; Acuña Rodríguez, D.; Scull Lizama, R.; Gutiérrez Gaitén, Y. Antimalarial Activity of Hydroalcoholic Extract from Bixa Orellana L. Rev. Cubana Med. Trop. 2011, 63, 181–185. [Google Scholar]
- Obakiro, S.B.; Kiprop, A.; K’owino, I.; Andima, M.; Owor, R.O.; Chacha, R.; Kigondu, E. Phytochemical, Cytotoxicity, and Antimycobacterial Activity Evaluation of Extracts and Compounds from the Stem Bark of Albizia Coriaria Welw Ex. Oliver. Evid.-Based Complement. Altern. Med. 2022, 2022, 7148511. [Google Scholar] [CrossRef]
- Dey, D.; Ray, R.; Hazra, B. Antimicrobial Activity of Pomegranate Fruit Constituents against Drug-Resistant Mycobacterium Tuberculosis and β-Lactamase Producing Klebsiella Pneumoniae. Pharm. Biol. 2015, 53, 1474–1480. [Google Scholar] [CrossRef] [Green Version]
- Sridevi, D.; Sudhakar, K.U.; Ananthathatmula, R.; Nankar, R.P.; Doble, M. Mutation at G103 of MtbFtsZ Altered Their Sensitivity to Coumarins. Front. Microbiol. 2017, 8, 578. [Google Scholar] [CrossRef]
- Salih, E.Y.A.; Julkunen-Tiitto, R.; Luukkanen, O.; Fahmi, M.K.M.; Fyhrquist, P. Hydrolyzable Tannins (Ellagitannins), Flavonoids, Pentacyclic Triterpenes and Their Glycosides in Antimycobacterial Extracts of the Ethnopharmacologically Selected Sudanese Medicinal Plant Combretum Hartmannianum Schweinf. Biomed. Pharmacother. 2021, 144, 112264. [Google Scholar] [CrossRef] [PubMed]
- Salih, E.Y.A.; Julkunen-Tiitto, R.; Luukkanen, O.; Sipi, M.; Fahmi, M.K.M.; Fyhrquist, P.J. Potential Anti-Tuberculosis Activity of the Extracts and Their Active Components of Anogeissus Leiocarpa (DC.) Guill. and Perr. with Special Emphasis on Polyphenols. Antibiotics 2020, 9, 364. [Google Scholar] [CrossRef] [PubMed]
- Luthra, S.; Rominski, A.; Sander, P. The Role of Antibiotic-Target-Modifying and Antibiotic-Modifying Enzymes in Mycobacterium Abscessus Drug Resistance. Front. Microbiol. 2018, 9, 364. [Google Scholar] [CrossRef]
- Da Fonseca Amorim, E.F.; Castro, E.J.M.; da Silva Souza, V.; Alves, M.S.; Dias, L.R.L.; Melo, M.H.F.; da Silva, I.M.A.; Villis, P.C.M.; Bonfim, M.R.Q.; Falcai, A.; et al. Antimicrobial Potential of Streptomyces Ansochromogenes (PB3) Isolated From a Plant Native to the Amazon Against Pseudomonas Aeruginosa. Front. Microbiol. 2020, 11, 574693. [Google Scholar] [CrossRef]
- Silva, T.F.; Filho, J.R.N.C.; Fonsêca, M.M.L.B.; Dos Santos, N.M.; da Silva, A.C.B.; Zagmignan, A.; Abreu, A.G.; da Silva, A.P.S.; de Menezes Lima, V.L.; da Silva, N.H.; et al. Products Derived from Buchenavia Tetraphylla Leaves Have in Vitro Antioxidant Activity and Protect Tenebrio Molitor Larvae against Escherichia Coli-induced Injury. Pharmaceuticals 2020, 13, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, Y.K.; Sulaiman, N.; Hakim, M.N.; Lian, G.E.C.; Zakaria, Z.A.; Othman, F.; Ahmad, Z. Suppressions of Serotonin-Induced Increased Vascular Permeability and Leukocyte Infiltration by Bixa Orellana Leaf Extract. Biomed Res. Int. 2013, 2013, 463145. [Google Scholar] [CrossRef] [Green Version]
- Zouari Bouassida, K.; Makni, S.; Tounsi, A.; Jlaiel, L.; Trigui, M.; Tounsi, S. Effects of Juniperus Phoenicea Hydroalcoholic Extract on Inflammatory Mediators and Oxidative Stress Markers in Carrageenan-Induced Paw Oedema in Mice. Biomed Res. Int. 2018, 2018, 3785487. [Google Scholar] [CrossRef] [Green Version]
- Mayouf, N.; Charef, N.; Saoudi, S.; Baghiani, A.; Khennouf, S.; Arrar, L. Antioxidant and Anti-Inflammatory Effect of Asphodelus Microcarpus Methanolic Extracts. J. Ethnopharmacol. 2019, 239, 111914. [Google Scholar] [CrossRef]
- Yilmazer, N.; Coskun, C.; Gurel-Gurevin, E.; Yaylim, I.; Eraltan, E.H.; Ikitimur-Armutak, E.I. Antioxidant and Anti-Inflammatory Activities of a Commercial Noni Juice Revealed by Carrageenan-Induced Paw Edema. Pol. J. Vet. Sci. 2016, 19, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Laavola, M.; Leppänen, T.; Eräsalo, H.; Hämäläinen, M.; Nieminen, R.; Moilanen, E. Anti-Inflammatory Effects of Nortrachelogenin in Murine J774 Macrophages and in Carrageenan-Induced Paw Edema Model in the Mouse. Planta Med. 2017, 83, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Ghorbanzadeh, B.; Mansouri, M.; Hemmati, A.; Naghizadeh, B.; Mard, S.; Rezaie, A. A Study of the Mechanisms Underlying the Anti-Inflammatory Effect of Ellagic Acid in Carrageenan-Induced Paw Edema in Rats. Indian J. Pharmacol. 2015, 47, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, A.M.; Martins de Sousa, E.; Viana-Niero, C.; Bonfim de Bortoli, F.; Pereira das Neves, Z.C.; Leão, S.C.; Junqueira-Kipnis, A.P.; Kipnis, A. Emergence of Nosocomial Mycobacterium Massiliense Infection in Goiás, Brazil. Microbes Infect. 2008, 10, 1552–1557. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, A.M.S.; de Andrade Monteiro, C.; Moraes-Neto, R.N.; Monteiro, A.S.; Mondego-Oliveira, R.; Nascimento, C.E.C.; da Silva, L.C.N.; Lima-Neto, L.G.; Carvalho, R.C.; de Sousa, E.M. Ethyl Acetate Fraction of Punica Granatum and Its Galloyl-HHDP-Glucose Compound, Alone or in Combination with Fluconazole, Have Antifungal and Antivirulence Properties against Candida Spp. Antibiotics 2022, 11, 265. [Google Scholar] [CrossRef]
- Shukla, A.; Sharma, P.; Prakash, O.; Singh, M.; Kalani, K.; Khan, F.; Bawankule, D.U.; Luqman, S.; Srivastava, S.K. QSAR and Docking Studies on Capsazepine Derivatives for Immunomodulatory and Anti-Inflammatory Activity. PLoS ONE 2014, 9, e100797. [Google Scholar] [CrossRef] [Green Version]
- Woods, G.L. Susceptibility Testing of Mycobacteria, Nocardiae and Other Aerobic Actinomycetes: Approved Standard; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011; ISBN 1562387464. [Google Scholar]
- Garcia de Carvalho, N.F.; Sato, D.N.; Pavan, F.R.; Ferrazoli, L.; Chimara, E. Resazurin Microtiter Assay for Clarithromycin Susceptibility Testing of Clinical Isolates of Mycobacterium Abscessus Group. J. Clin. Lab. Anal. 2016, 30, 751–755. [Google Scholar] [CrossRef] [Green Version]
- Neves, R.C.; Andrade, R.G.; Carollo, C.A.; Boaretto, A.G.; Kipnis, A.; Junqueira-Kipnis, A.P. Hyptis Brevipes and Paullinia Pinnata Extracts and Their Fractions Presenting Activity against Mycobacterium Abscessus SubSP. Massiliense. J. Trop. Pathol. 2019, 48, 148–160. [Google Scholar] [CrossRef]
- Lozoya-Pérez, N.E.; García-Carnero, L.C.; Martínez-Álvarez, J.A.; Martínez-Duncker, I.; Mora-Montes, H.M. Tenebrio Molitor as an Alternative Model to Analyze the Sporothrix Species Virulence. Infect. Drug Resist. 2021, 14, 2059–2072. [Google Scholar] [CrossRef]
- Santos, A.L.E.; Júnior, C.P.S.; Neto, R.N.M.; Santos, M.H.C.; Santos, V.F.; Rocha, B.A.M.; Sousa, E.M.; Carvalho, R.C.; Menezes, I.R.A.; Oliveira, M.R.C.; et al. Machaerium Acutifolium Lectin Inhibits Inflammatory Responses through Cytokine Modulation. Process Biochem. 2020, 97, 149–157. [Google Scholar] [CrossRef]
- Meshram, G.G.; Kumar, A.; Rizvi, W.; Tripathi, C.D.; Khan, R.A. Evaluation of the Anti-Inflammatory Activity of the Aqueous and Ethanolic Extracts of the Leaves of Albizzia Lebbeck in Rats. J. Tradit. Complement. Med. 2016, 6, 172–175. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | UV Max (nm) |
|---|---|
| G1 (mixture) | 300; 325; 278 |
| G2 (mixture) | 300; 325; 278 |
| G3 (mixture) | 300; 325; 278 |
| G4 (mixture) | 300; 325; 278 |
| G5 (mixture) | 255; 267; 349 |
| G6 (mixture) | 253; 267; 345 |
| G7 (mixture) | 253; 267; 345 |
| G8 (mixture) | 253; 267; 345 |
| G9 (mixture) | 300; 325; 278 |
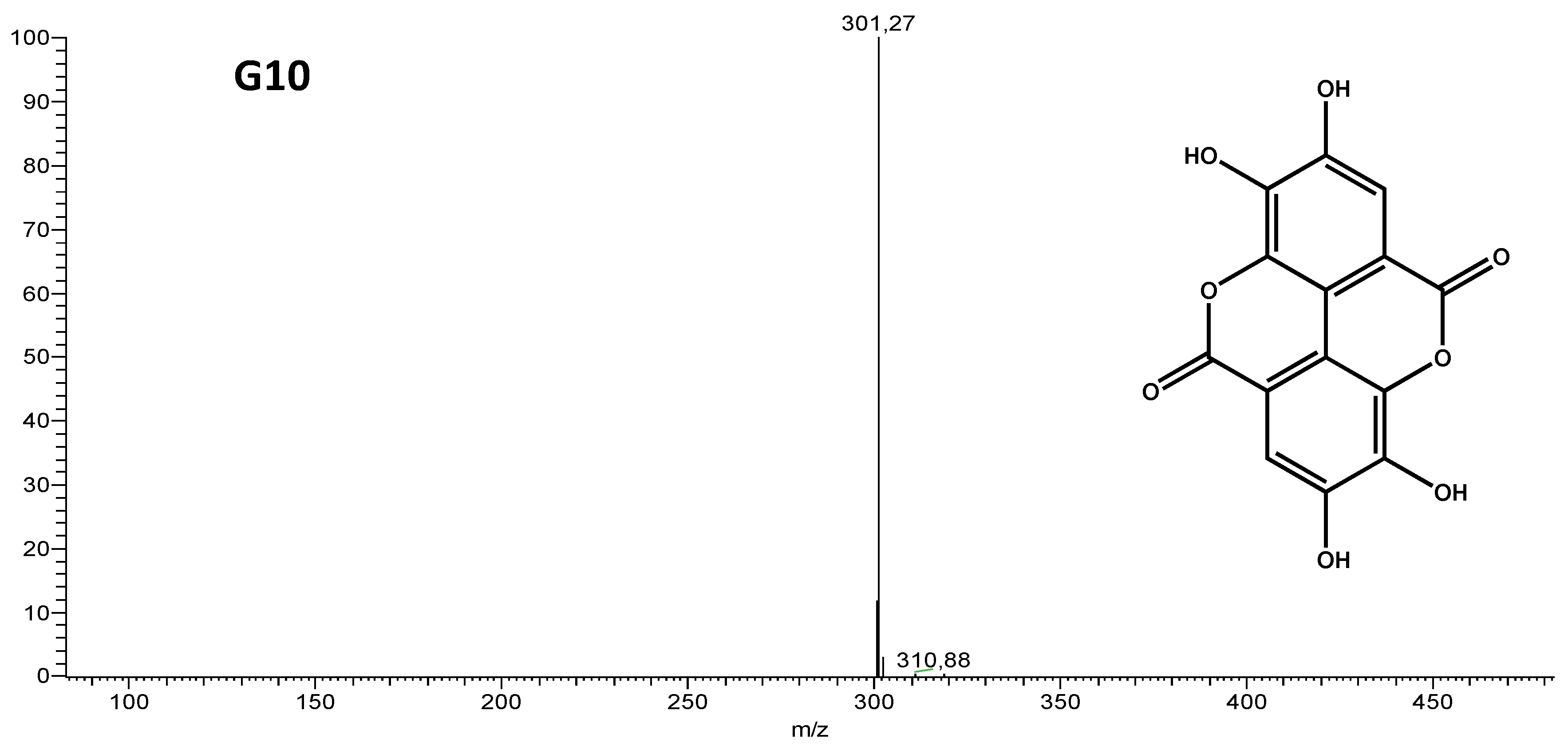
| G10 (pure compound) | 254; 300; 366 |
| [M-H]- | MSnions | Identification |
|---|---|---|
| 301 | 257 [M-44-H]- | Ellagic Acid |
| 229 [M-44-28-H]- |
| Activities | PASS Predictions of Ellagic Acid | |
|---|---|---|
| Pa | Pi | |
| Anti-inflammatory | 0.749 | 0.010 |
| Antioxidant | 0.699 | 0.004 |
| Antibacterial | 0.380 | 0.035 |
| Antimycobacterial | 0.301 | 0.080 |
| Hepatoprotective | 0.599 | 0.012 |
| DNA ligase (ATP) inhibitor | 0.466 | 0.009 |
| Superoxide dismutase inhibitor | 0.423 | 0.058 |
| DNA-3-methyladenine glycosylase I inhibitor | 0.379 | 0.029 |
| RNA directed DNA polymerase inhibitor | 0.323 | 0.026 |
| DNA polymerase I inhibitor | 0.330 | 0.041 |
| Nitric oxide scavenger | 0.282 | 0.011 |
| DNA synthesis inhibitor | 0.261 | 0.074 |
| Antibiotic | 0.158 | 0.044 |
| Antibacterial Ophthalmic | 0.154 | 0.051 |
| Cell wall synthesis inhibitor | 0.094 | 0.072 |
| tRNA-pseudouridine synthase I inhibitor | 0.617 | 0.014 |
| Compound | PPB | Hepatotoxic | Solubility | BBBP | HIA | Lipinski Violations (Max 4) |
|---|---|---|---|---|---|---|
| Clarithromycin | false | True | 2 | 4 | 3 | 3 |
| Diclofenac | true | True | 2 | 1 | 0 | 0 |
| Ellagic Acid | true | True | 3 | 4 | 1 | 0 |
| Compound | Mouse Female | Rat Female | Ames Mutagenicity | Skin Irritancy | Skin Sensitization | Ocular Irritancy |
|---|---|---|---|---|---|---|
| Clarithromycin | NC | NC | NM | Mild | None | Mild |
| Diclofenac | NC | NC | NM | None | Strong | Mild |
| Ellagic Acid | NC | NC | NM | None | Strong | Mild |
| Compound | Rat Oral LD50 (g/kg Body Weight) | Rat Chronic LOAEL (g/kg Body Weight) | Fathead Minnow LC50 (g/L) | Daphnia EC50 (mg/L) |
|---|---|---|---|---|
| Clarithromycin | 0.787 | 0.001 | 0.004 | 0.278 |
| Diclofenac | 0.564 | 0.034 | 0.002 | 9.665 |
| Ellagic Acid | 0.428 | 0.302 | 0.049 | 16.407 |
| Compound | Rat TD50 (mg/kg Body Weight/Day) | RMTD—Feed (mg/kg Body Weight/Day) |
|---|---|---|
| Clarithromycin | 0.002 | 9.995 |
| Diclofenac | 51.448 | 742.520 |
| Ellagic Acid | 5.838 | 721.906 |
| Enzyme | Ligands Ligand ID/(Synonyms) | Coordinates of Grid Center |
|---|---|---|
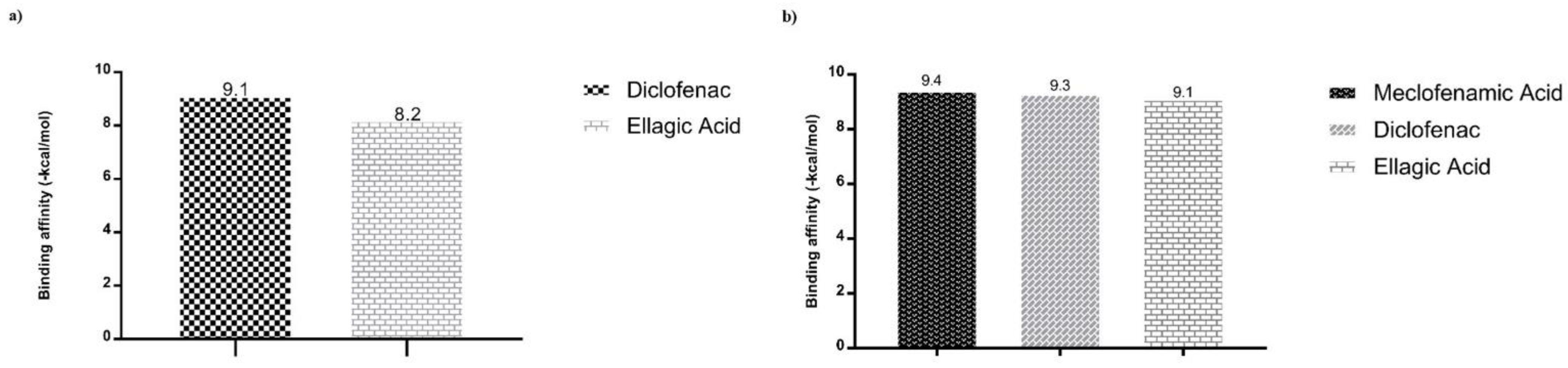
| COX2 (Prostaglandin G/H synthase 2) (Mus musculus) PDB ID: 1PXX Resolution: 2.90 Å Reference: [23] | 2-[2-[(2,6-dichlorophenyl)amino]phenyl]ethanoic acid DIF Diclofenac | X = 27,115 Y = 24,090 Z = 14,936 |
| COX2 Prostaglandin G/H synthase 2 (Homo sapiens) PDB ID: 5IKQ Resolution: 2.41 Å Reference: [24] | 2-[(2,6-dichloro-3-methyl-phenyl)amino]benzoic acid 2- / JMS Meclofenamic Acid | X = 21,597 Y = 51,876 Z = 17,696 |
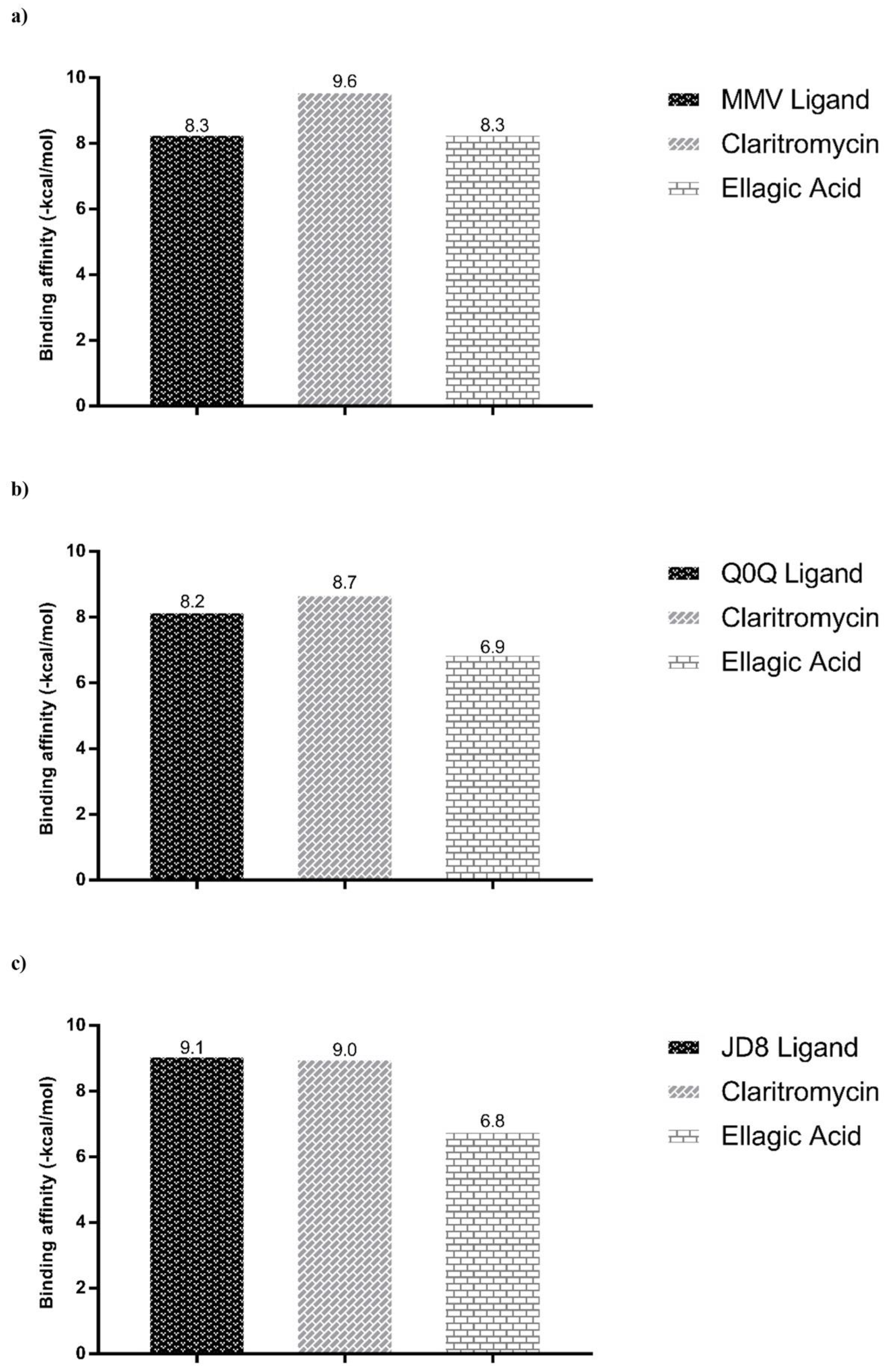
| Dihydrofolate reductase (Mycobacteroides abscessus ATCC 19977) PDB ID: 7K6C Resolution: 2.00 Å Reference: [25] | 3-(2-{3-[(2,4-diamino-6-ethylpyrimidin-5-yl)oxy]propoxy}phenyl)propanoic acid/ MMV | X = -33,882 Y = −7502 Z = 56,281 |
| Phosphoribosylaminoimidazole-succinocarboxamide synthase (Mycobacteroides abscessus ATCC 19977) PDB ID: 6YYB Resolution: 1.51 Å Reference: [26] | 4-azanyl-6-[1-[(1~{R})-1-phenylethyl]pyrazol-4-yl]pyrimidine-5-carbonitrile Q0Q | X = 21,1853 Y = 14,726 Z = 34,7921 |
| tRNA (guanine-N(1)-)-methyltransferase (Mycobacteroides abscessus) PDB ID: 6QR4 Resolution: 1.52 Å Reference: [27] | zanyl-3-[1-[[(2~{R})-1-methylpiperidin-2-yl]methyl]indol-6-yl]-1~{H}-pyrazole-4-carbonitrile JD8 | X = -12,718 Y = 7688 Z = −27,062 |
| Time (h) | Control Group | Antibacterial Agents | ||||||
|---|---|---|---|---|---|---|---|---|
| CLA | BoEA MIC | BoEA 2 × MIC | BoEA 4 × MIC | ElAc MIC | ElAc 2 × MIC | ElAc 4 × MIC | ||
| 0 | 5.9 ± 0.08 | 5.9 ± 0.08 | 5.9 ± 0.08 ** | 5.9 ± 0.08 ** | 5.9 ± 0.08 ** | 5.9 ± 0.08 ** | 5.9 ± 0.08 ** | 5.9 ± 0.08 ** |
| 12 | 7.03 ± 0.12 | 6.76 ± 0.16 | 6.83 ± 0.12 ** | 6.83 ± 0.12 ** | 6.9 ± 0.08 ** | 6.83 ± 0.12 ** | 6.66 ± 0.23 ** | 5.66 ± 0.12 ** |
| 24 | 8.9 ± 0.08 | 6.73 ± 0.16 | 7.23 ± 0.16 ** | 7.23 ± 0.16 ** | 6.53 ± 0.12 ** | 6.3 ± 0.08 ** | 6.06 ± 0.09 ** | 5.76 ± 0.26 ** |
| 36 | 9.1 ± 0.16 | 6.9 ± 0.08 | 7.4 ± 0.37 ** | 7.1 ± 0.08 ** | 6.33 ± 0.12 ** | 6.53 ± 0.12 ** | 5.83 ± 0.16 ** | 5.73 ± 0.20 ** |
| 48 | 9.76 ± 0.04 | 7.03 ± 0.12 | 8.5 ± 0.40 * | 7.86 ± 0.04 * | 6.66 ± 0.12 * | 6.8 ± 0.08 ** | 5.76 ± 0.04 ** | 5.30 ± 0.08 ** |
| 72 | 10.26 ± 0.44 | 7.5 ± 0.21 | 8.5 ± 0.37 * | 7.23 ± 0.30 * | 6.66 ± 0.12 * | 7.0 ± 0.08 ** | 6.23 ± 0.16 ** | 5.36 ± 0.26 ** |
| 96 | 10.7 ± 0.16 | 8.1 ± 0.14 | 9.7 ± 0.24 * | 8.46 ± 0.41 * | 7.16 ± 0,12 * | 7.36 ± 0.09 ** | 6.23 ± 0.30 ** | 5.33 ± 0.20 ** |
| 120 | 9.7 ± 0.16 | 8.33 ± 0.12 | 9.9 ± 0.08 * | 9.1 ± 0.08 * | 8.33 ± 0.23 * | 6.86 ± 0.12 ** | 7.03 ± 0.12 ** | 6.26 ± 0.18 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moraes-Neto, R.N.; Coutinho, G.G.; Ataíde, A.C.S.; de Oliveira Rezende, A.; Nascimento, C.E.C.; de Albuquerque, R.P.; da Rocha, C.Q.; Rêgo, A.S.; de Sousa Cartágenes, M.d.S.; Abreu-Silva, A.L.; et al. Ethyl Acetate Fraction of Bixa orellana and Its Component Ellagic Acid Exert Antibacterial and Anti-Inflammatory Properties against Mycobacterium abscessus subsp. massiliense. Antibiotics 2022, 11, 817. https://doi.org/10.3390/antibiotics11060817
Moraes-Neto RN, Coutinho GG, Ataíde ACS, de Oliveira Rezende A, Nascimento CEC, de Albuquerque RP, da Rocha CQ, Rêgo AS, de Sousa Cartágenes MdS, Abreu-Silva AL, et al. Ethyl Acetate Fraction of Bixa orellana and Its Component Ellagic Acid Exert Antibacterial and Anti-Inflammatory Properties against Mycobacterium abscessus subsp. massiliense. Antibiotics. 2022; 11(6):817. https://doi.org/10.3390/antibiotics11060817
Chicago/Turabian StyleMoraes-Neto, Roberval Nascimento, Gabrielle Guedes Coutinho, Ana Caroline Santos Ataíde, Aline de Oliveira Rezende, Camila Evangelista Carnib Nascimento, Rafaela Pontes de Albuquerque, Cláudia Quintino da Rocha, Adriana Sousa Rêgo, Maria do Socorro de Sousa Cartágenes, Ana Lúcia Abreu-Silva, and et al. 2022. "Ethyl Acetate Fraction of Bixa orellana and Its Component Ellagic Acid Exert Antibacterial and Anti-Inflammatory Properties against Mycobacterium abscessus subsp. massiliense" Antibiotics 11, no. 6: 817. https://doi.org/10.3390/antibiotics11060817
APA StyleMoraes-Neto, R. N., Coutinho, G. G., Ataíde, A. C. S., de Oliveira Rezende, A., Nascimento, C. E. C., de Albuquerque, R. P., da Rocha, C. Q., Rêgo, A. S., de Sousa Cartágenes, M. d. S., Abreu-Silva, A. L., dos Santos, I. V. F., dos Santos, C. B. R., Guerra, R. N. M., Ribeiro, R. M., Monteiro-Neto, V., de Sousa, E. M., & Carvalho, R. C. (2022). Ethyl Acetate Fraction of Bixa orellana and Its Component Ellagic Acid Exert Antibacterial and Anti-Inflammatory Properties against Mycobacterium abscessus subsp. massiliense. Antibiotics, 11(6), 817. https://doi.org/10.3390/antibiotics11060817