
Companion Animals as Potential Reservoirs of Antibiotic Resistant Diarrheagenic Escherichia coli in Shandong, China


Abstract
:1. Introduction
2. Results
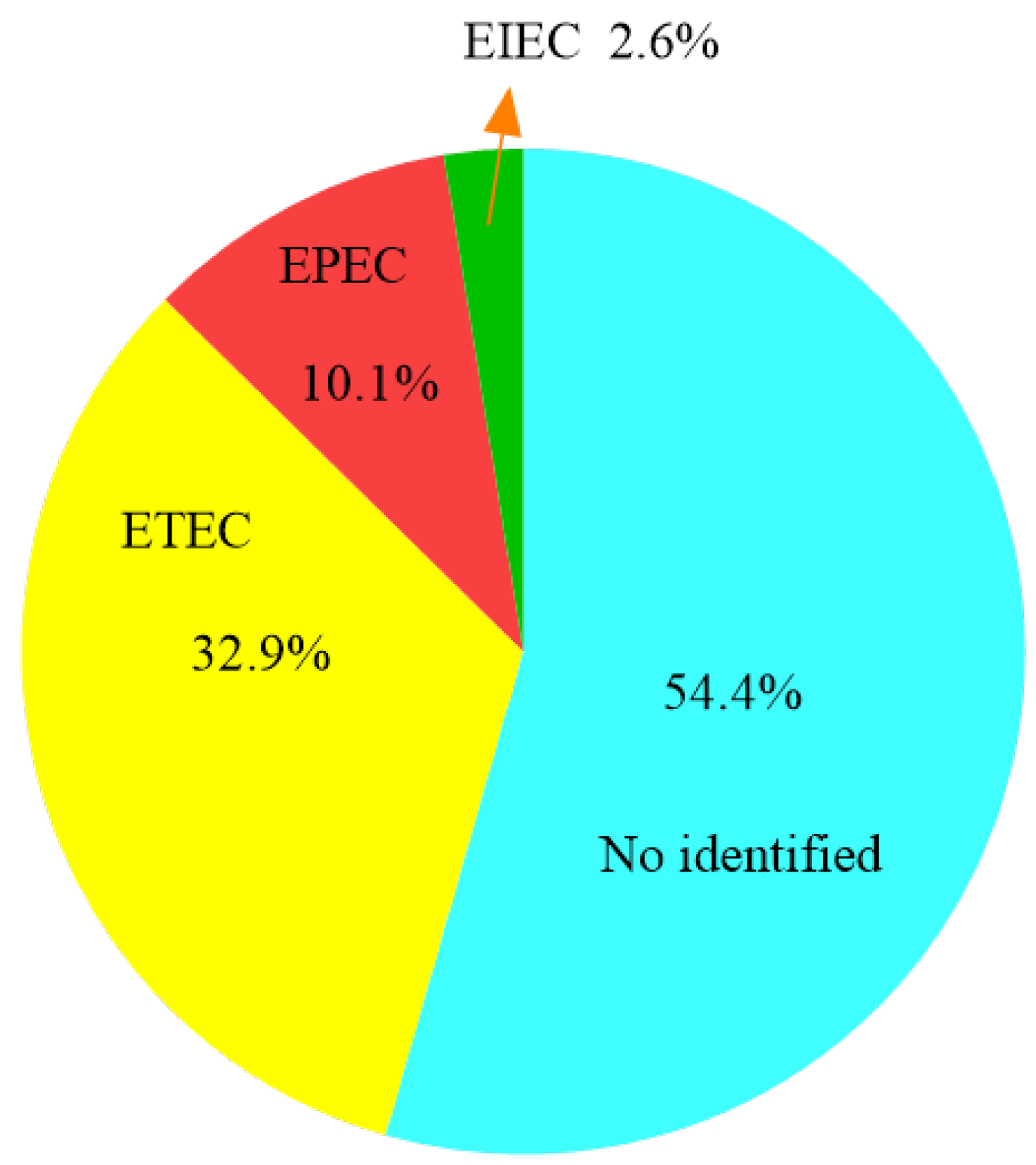
2.1. Isolation and Serologic Characterization of E. coli Strains
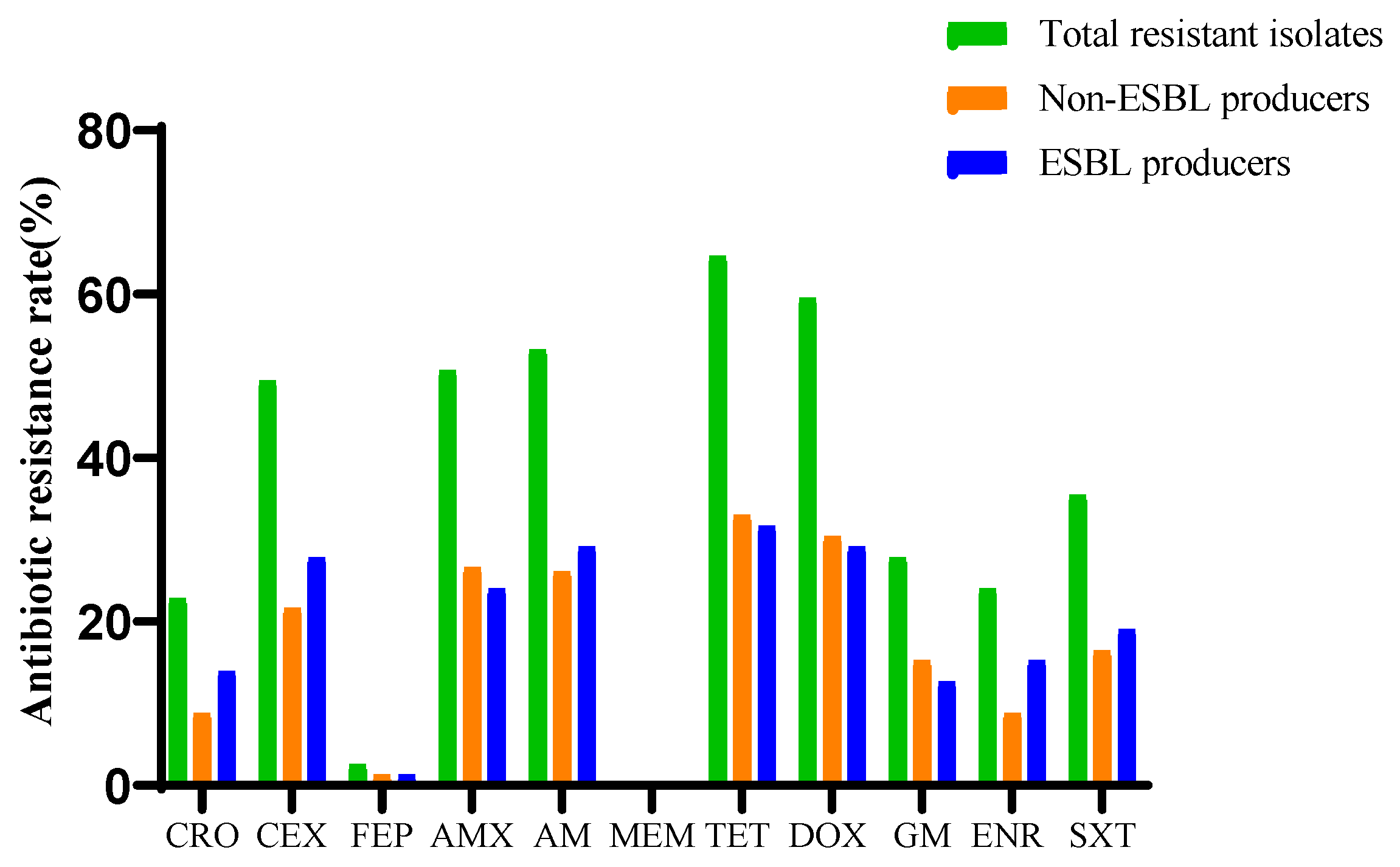
2.2. Resistance Profiles of E. coli Strains
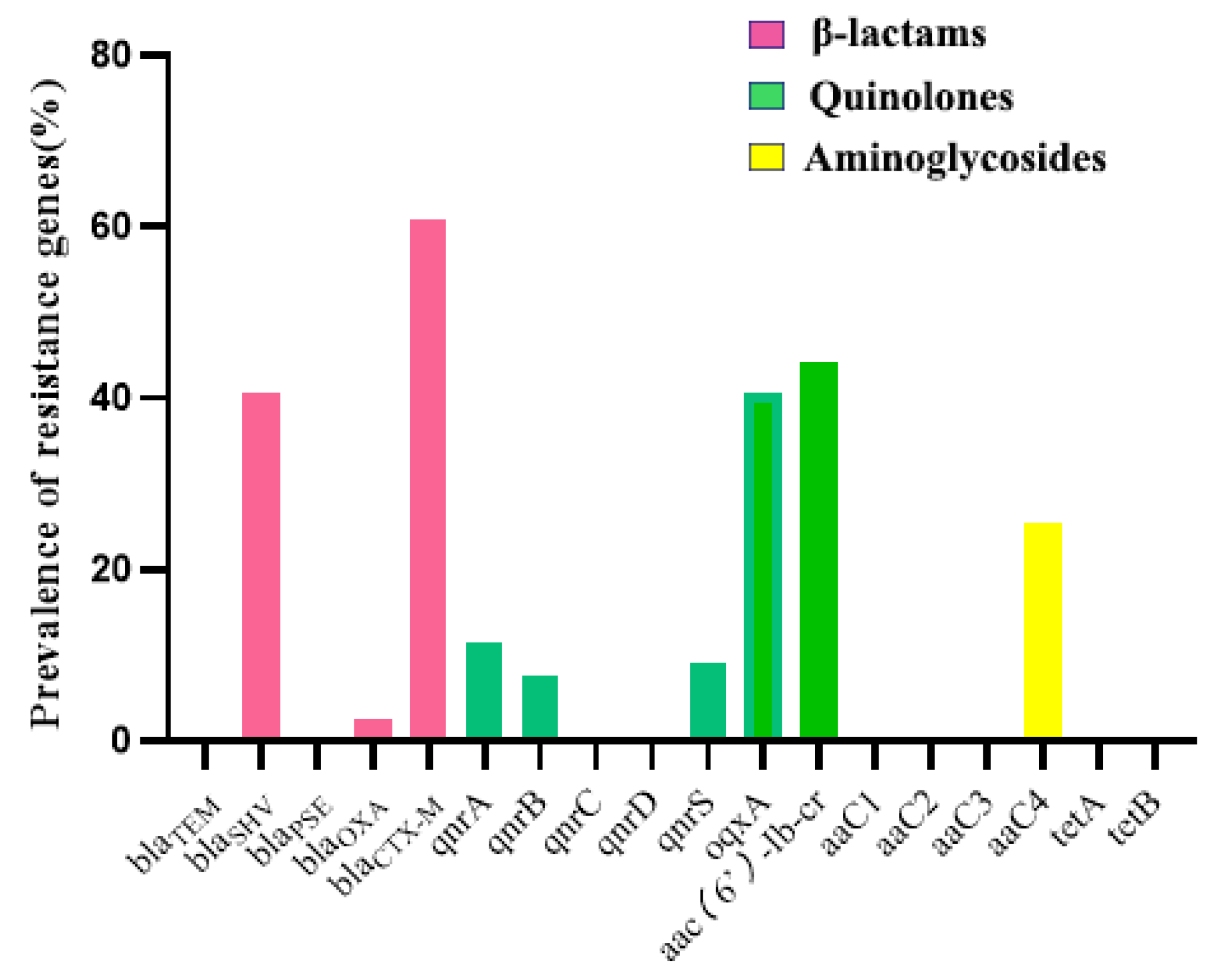
2.3. Detection of Antimicrobial Resistance Genes
2.4. Concordance of Genotypic-Phenotypic Antimicrobial Resistance
2.5. Distribution of Virulence Genes in E. coli Isolates
3. Discussion
4. Materials and Methods
4.1. Sampling Size Determination
4.2. Bacterial Isolates
4.3. Determination of Serotypes, and Genomic DNA Extraction
4.4. Antimicrobial Susceptibility Testing and Detection of ESBL-EC
4.5. Detection of Resistance Genes
4.6. Detection of Virulence Genes
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Sequence (5′ → 3′) | Product Size (bp) | Reference |
|---|---|---|---|
| β-Lactamases | |||
| blaTEM | F: ATAAAATTCTTGAAGACGAAA | 643 | [69] |
| R: GACAGTTACCAATGCTTAATC | |||
| blaSHV | F: TTATCTCCCTGTTAGCCACC | 860 | [69] |
| R: GATTTGCTGATTTCGCTCGG | |||
| blaPSE | F: TAGGTGTTTCCGTTCTTG | 150 | [70] |
| R: TCATTTCGCTCTTCCATT | |||
| blaOXA | F TCAACTTTCAAGATCGCA | 591 | [69] |
| R: GTGTGTTTAGAATGGTGA | |||
| blaCTX-M | F: CGCTTTGCGATGTGCAG | 550 | [69] |
| R: ACCGCGATATCGTTGGT | |||
| quinolones | |||
| qnrA | F: ATTTCTCACGCCAGGATTTG | 519 | [69] |
| R: GATCGGCAAAGGTCAGGTCA | |||
| qnrB | F: GATCGTGAAAGCCAGAAAGG | 513 | [69] |
| R: ACGATGCCTGGTAGTTGTCC | |||
| qnrC | F: GGTTGTACATTTATTGAATC | 666 | [71] |
| R: TCCACTTTACGAGGTTCT | |||
| qnrD | F: AGATCAATTTACGGGGAATA | 984 | [71] |
| R: AACAAGCTGAAGCGCCTG | |||
| qnrS | F: ACGACATTCGTCAACTGCAA | 417 | [69] |
| R: TAAATTGGCACCCTGTAGGC | |||
| qqxA | F: GATCAGTCAGTGGGATAGTTT | 670 | [71] |
| R: TACTCGGCGTTAACTGATTA | |||
| Aac(6’)-Ib-cr | F: TTGCGATGCTCTATGAGTGGCTA | 482 | [69] |
| R: CTCGAATGCCTGGCGTGTTT | |||
| aminoglycosides | |||
| aaC1 | F: ACCTACTCCCAACATCAGCC | 528 | [72] |
| R: ATATAGATCTCACTACGCGC | |||
| aaC2 | F: ACTGTGATGGGATACGCGTC | 482 | [72] |
| R: CTCCGTCAGCGTTTCAGCTA | |||
| aaC3 | F: CACAAGAACGTGGTCCGCTA | 185 | [72] |
| R: AACAGGTAAGCATCCGCATC | |||
| aaC4 | F: CTTCAGGATGGCAAGTTGGT | 286 | [72] |
| R: TCATCTCGTTCTCCGCTCAT | |||
| tetracyclines | |||
| tetA | F: GCGCCTTTCCTTTGGGTTCT | 211 | [72] |
| R: CCACCCGTTCCACGTTGTTA | |||
| tetB | F: CATTAATAGGCGCATCGCTG | 391 | [72] |
| R: TGAAGGTCATCGATAGCAGG | |||
| R: AAGCAGACTTGACCTGA | |||
| Virulence Factors | Sequence (5′-3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|
| K88 | F: GATGAAAAAGACTCTGATTGCA | 841 | [73] |
| R: GATTGCTACGTTCAGCGGAGCG | |||
| K99 | F: CTGAAAAAAACACTGCTAGCTATT | 543 | [73] |
| R: CATATAAGTGACTAAGAAGGATGC | |||
| F17 | F: GCAGAAAATTCAATTTATCCTTGG | 537 | [74] |
| R: CTGATAAGCGATGGTGTAATTAAC | |||
| F18 | F: ATGAAAAGACTAGTGTTTATTTCTT | 520 | [73] |
| R: TTACTTGTAAGTAACCGCGTAAGCC | |||
| F41 | F: GATGAAAAAGACTCTGATTGCA | 682 | [73] |
| R: TCTGAGGTCATCCCAATTGTGG | |||
| 987p | F: GTTACTGCCAGTCTATGCCAAGTG | 463 | [73] |
| R: TCGGTGTACCTGCTGAACGAATAG | |||
| CS31A | F: GGGCGCTCTCTCCTTCAAC | 402 | [74] |
| R: CGCCCTAATTGCTGGCGAC | |||
| bfpA | F: GGTCTGTCTTTGATTGAATC | 485 | [75] |
| R: TTTACATGCAGTTGCCGCTT | |||
| Stx1 | F: ATTCGCTGAATGTCATTCGCT | 664 | [73] |
| R: ACGCTTCCCAGAATTGCATTA | |||
| Stx2 | F: GAATGAAGAAGATGTTTATAGCGG | 281 | [73] |
| R: GGTTATGCCTCAGTCATTATTAA | |||
| hlyA | F: GCATCATCAAGCGTACGTTCC | 533 | [73] |
| R: AATGAGCCAAGCTGGTTAAGCT | |||
| LT | F: CCGAATTCTGTTATATATGTC | 696 | [75] |
| R: GGCGACAGATTATACCGTGC | |||
| Sta | F: GGGTTGGCAATTTTTATTTCTGTA | 183 | [73] |
| R: ATTACAACAAAGTTCACAGCAGTA | |||
| EAST1 | F: ATGCCATCAACACAGTATATC | 117 | [73] |
| R: TCAGGTCGCGAGTGACGG | |||
| irp2 | F: AAGGATTCGCTGTTACCGGAC | 287 | [73] |
| R: TCGTCGGGCAGCGTTTCTTCT | |||
| eaeA | F: CATTGATCAGGATTTTTCTGGT | 510 | [75] |
References
- Gomes, T.A.; Elias, W.P.; Scaletsky, I.C.; Guth, B.E.; Rodrigues, J.F.; Piazza, R.M.; Ferreira, L.C.; Martinez, M.B. Diarrheagenic Escherichia coli. Braz. J. Microbiol. 2016, 47 (Suppl. 1), 3–30. [Google Scholar] [CrossRef] [Green Version]
- Puno-Sarmiento, J.; Medeiros, L.; Chiconi, C.; Martins, F.; Pelayo, J.; Rocha, S.; Blanco, J.; Blanco, M.; Zanutto, M.; Kobayashi, R.; et al. Detection of Diarrheagenic Escherichia coli Strains Isolated from Dogs and Cats in Brazil. Vet. Microbiol. 2013, 166, 676–680. [Google Scholar] [CrossRef]
- Prah, I.; Ayibieke, A.; Mahazu, S.; Sassa, C.T.; Hayashi, T.; Yamaoka, S.; Suzuki, T.; Iwanaga, S.; Ablordey, A.; Saito, R. Emergence of Oxacillinase-181 Carbapenemase-Producing Diarrheagenic Escherichia coli in Ghana. Emerg. Microbes Infect. 2021, 10, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Beutin, L. Escherichia coli as A Pathogen in Dogs and Cats. Vet. Res. 1999, 30, 285–298. [Google Scholar] [PubMed]
- Krause, G.; Zimmermann, S.; Beutin, L. Investigation of Domestic Animals and Pets as A Reservoir for Intimin- (eae) Gene Positive Escherichia coli Types. Vet. Microbiol. 2005, 106, 87–95. [Google Scholar] [CrossRef]
- Vega-Manriquez, X.D.; Ubiarco-Lopez, A.; Verdugo-Rodriguez, A.; Hernandez-Chinas, U.; Navarro-Ocana, A.; Ahumada-Cota, R.E.; Ramirez-Badillo, D.; Hernandez-Diaz de Leon, N.; Eslava, C.A. Pet Dogs Potential Transmitters of Pathogenic Escherichia coli with Resistance to Antimicrobials. Arch. Microbiol. 2020, 202, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Escher, M.; Scavia, G.; Morabito, S.; Tozzoli, R.; Maugliani, A.; Cantoni, S.; Fracchia, S.; Bettati, A.; Casa, R.; Gesu, G.P.; et al. A Severe Foodborne Outbreak of Diarrhoea Linked to A Canteen in Italy Caused by Enteroinvasive Escherichia coli, An Uncommon Agent. Epidemiol. Infect. 2014, 142, 2559–2566. [Google Scholar] [CrossRef]
- Newitt, S.; MacGregor, V.; Robbins, V.; Bayliss, L.; Chattaway, M.A.; Dallman, T.; Ready, D.; Aird, H.; Puleston, R.; Hawker, J. Two Linked Enteroinvasive Escherichia coli Outbreaks, Nottingham, UK, June 2014. Emerg. Infect. Dis. 2016, 22, 1178–1184. [Google Scholar] [CrossRef] [Green Version]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.M.; Wertheim, H.F.L.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic Resistance—The Need for Global Solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.; Clift, C.; Kai, S.; Sagan, A.; Mossialos, E. Averting the AMR Crisis. What are the Avenues for Policy Action for Countries in Europe? European Observatory on Health Systems and Policies: Copenhagen, Denmark, 2019. [Google Scholar]
- Hong, J.S.; Song, W.; Park, H.M.; Oh, J.Y.; Chae, J.C.; Jeong, S.; Jeong, S.H. Molecular Characterization of Fecal Extended-Spectrum Beta-Lactamase- and AmpC Beta-Lactamase-Producing Escherichia coli from Healthy Companion Animals and Cohabiting Humans in South Korea. Front. Microbiol. 2020, 11, 674. [Google Scholar] [CrossRef] [Green Version]
- Kock, R.; Daniels-Haardt, I.; Becker, K.; Mellmann, A.; Friedrich, A.W.; Mevius, D.; Schwarz, S.; Jurke, A. Carbapenem-Resistant Enterobacteriaceae in Wildlife, Food-Producing, and Companion Animals: A Systematic Review. Clin. Microbiol. Infect. 2018, 24, 1241–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, L.; Wang, Y.; He, J.; Cai, C.; Liu, Q.; Yang, D.; Zou, Z.; Shi, L.; Jia, J.; Wang, Y.; et al. Prevalence and Risk Analysis of Mobile Colistin Resistance and Extended-Spectrum Beta-Lactamase Genes Carriage in Pet Dogs and Their Owners: A Population Based Cross-Sectional Study. Emerg. Microbes Infect. 2021, 10, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.M.; Cowling, A. The Distribution and Genetic Structure of Escherichia coli in Australian Vertebrates: Host and Geographic Effects. Microbiology 2003, 149, 3575–3586. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Madec, J.Y.; Lupo, A.; Schink, A.K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectrum 2018, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- WHO Organisation. WHO Advisory Group on Integrated Surveillance of Antimicrobial Resistance (AGISAR). In Proceedings of the Report of The 7th Meeting, Raleigh, NC, USA, 17–20 October 2016; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215, S28–S36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skov, R.L.; Hammerum, A.M.; Larsen, J.; Aarestrup, F.M.; Lester, C.H.; Skytte, T.; Mordhorst, H.; Olsen, S.S.; Andersen, V.D.; Hansen, F. Characterization of Extended-Spectrum Beta-Lactamase (ESBL)-Producing Escherichia coli Obtained from Danish Pigs, Pig Farmers and Their Families from Farms with High or No Consumption of Third- or Fourth-Generation Cephalosporins. J. Antimicrob. Chemother. 2014, 69, 2650–2657. [Google Scholar]
- Wedley, A.L.; Dawson, S.; Maddox, T.W.; Coyne, K.P.; Pinchbeck, G.L.; Clegg, P.; Nuttall, T.; Kirchner, M.; Williams, N.J. Carriage of Antimicrobial Resistant Escherichia coli in Dogs: Prevalence, Associated Risk Factors and Molecular Characteristics. Vet. Microbiol. 2017, 199, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.J.; Kim, S.S.; Mollenkopf, D.F.; Mathys, D.A.; Schuenemann, G.M.; Daniels, J.B.; Wittum, T.E. Antimicrobial-Resistant Enterobacteriaceae Recovered from Companion Animal and Livestock Environments. Zoonoses Public Health 2018, 65, 519–527. [Google Scholar] [CrossRef]
- Ho, P.L.; Chow, K.H.; Lai, E.L.; Lo, W.U.; Yeung, M.K.; Chan, J.; Chan, P.Y.; Yuen, K.Y. Extensive Dissemination of CTX-M-Producing Escherichia coli with Multidrug Resistance to ‘Critically Important’ Antibiotics among Food Animals in Hong Kong, 2008–2010. J. Antimicrob. Chemother. 2011, 66, 765–768. [Google Scholar] [CrossRef] [Green Version]
- Rubin, J.E.; Pitout, J.D. Extended-Spectrum Beta-Lactamase, Carbapenemase and AmpC Producing Enterobacteriaceae in Companion Animals. Vet. Microbiol. 2014, 170, 10–18. [Google Scholar] [CrossRef]
- Morato, E.P.; Leomil, L.; Beutin, L.; Krause, G.; Moura, R.A.; Pestana de Castro, A.F. Domestic Cats Constitute a Natural Reservoir of Human Enteropathogenic Escherichia coli Types. Zoonoses Public Health 2009, 56, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Fischer, J.; Helmuth, R. An Emerging Public Health Problem: Acquired Carbapenemase-Producing Microorganisms are Present in Food-Producing Animals, Their Environment, Companion Animals and Wild Birds. Vet. Microbiol. 2014, 171, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Huber, H.; Zweifel, C.; Wittenbrink, M.M.; Stephan, R. ESBL-Producing Uropathogenic Escherichia coli Isolated from Dogs and Cats in Switzerland. Vet. Microbiol. 2013, 162, 992–996. [Google Scholar] [CrossRef] [PubMed]
- So, J.H.; Kim, J.; Bae, I.K.; Jeong, S.H.; Kim, S.H.; Lim, S.K.; Park, Y.H.; Lee, K. Dissemination of Multidrug-Resistant Escherichia coli in Korean Veterinary Hospitals. Diagn. Microbiol. Infect. Dis. 2012, 73, 195–199. [Google Scholar] [CrossRef]
- Edney, A.T. Companion Animals and Human Health. Vet. Rec. 1992, 130, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.M.D.; Menezes, J.; Marques, C.; Pomba, C.F. Companion Animals-an Overlooked and Misdiagnosed Reservoir of Carbapenem Resistance. Antibiotics 2022, 11, 533. [Google Scholar] [CrossRef]
- Gronthal, T.; Osterblad, M.; Eklund, M.; Jalava, J.; Nykasenoja, S.; Pekkanen, K.; Rantala, M. Sharing More than Friendship-Transmission of NDM-5 ST167 and CTX-M-9 ST69 Escherichia coli between Dogs and Humans in a Family, Finland, 2015. Eurosurveill 2018, 23, 1700497. [Google Scholar] [CrossRef]
- Arais, L.R.; Barbosa, A.V.; Andrade, J.R.C.; Gomes, T.A.T.; Asensi, M.D.; Aires, C.A.M.; Cerqueira, A.M.F. Zoonotic Potential of a typical Enteropathogenic Escherichia coli (aEPEC) Isolated from Puppies with Diarrhoea in Brazil. Vet. Microbiol. 2018, 227, 45–51. [Google Scholar] [CrossRef]
- Askari, A.; Ghanbarpour, R.; Akhtardanesh, B.; Aflatoonian, M.R.; Sharifi, H.; Jajarmi, M.; Molaei, R. Detection of Zoonotic Diarrheagenic Pathotypes of Escherichia coli in Healthy Household Dogs. Iran. J. Microbiol. 2020, 12, 522–530. [Google Scholar] [CrossRef]
- Zahraei Salehi, T.; Askari Badouei, M.; Mehdizadeh Gohari, I. Molecular Detection and Antibacterial Susceptibility of Enteropathogenic Escherichia coli (EPEC) and Shigatoxigenic Escherichia coli (STEC) Strains Isolated from Healthy and Diarrhoeic Dogs. Comp. Clin. Path. 2010, 20, 585–589. [Google Scholar] [CrossRef]
- Reygaert, W.C. An Overview of the Antimicrobial Resistance Mechanisms of Bacteria. AIMS Microbiol. 2018, 4, 482–501. [Google Scholar] [CrossRef] [PubMed]
- Beceiro, A.; Tomas, M.; Bou, G. Antimicrobial Resistance and Virulence: A Successful or Deleterious Association in the Bacterial World? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guardabassi, L.; Schwarz, S.; Lloyd, D.H. Pet Animals as Reservoirs of Antimicrobial-Resistant Bacteria. J. Antimicrob. Chemoth. 2004, 54, 321–332. [Google Scholar] [CrossRef]
- Costa, D.; Poeta, P.; Saenz, Y.; Coelho, A.C.; Matos, M.; Vinue, L.; Rodrigues, J.; Torres, C. Prevalence of Antimicrobial Resistance and Resistance Genes in Faecal Escherichia coli Isolates Recovered from Healthy Pets. Vet. Microbiol. 2008, 127, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, L.; Buldain, D.; Gortari Castillo, L.; Buchamer, A.; Chirino-Trejo, M.; Mestorino, N. Pet and Stray Dogs as Reservoirs of Antimicrobial-Resistant Escherichia coli. Int. J. Microbiol. 2021, 2021, 6664557. [Google Scholar] [CrossRef]
- Zogg, A.L.; Zurfluh, K.; Schmitt, S.; Nuesch-Inderbinen, M.; Stephan, R. Antimicrobial Resistance, Multilocus Sequence Types and Virulence Profiles of ESBL Producing and Non-ESBL Producing Uropathogenic Escherichia coli Isolated from Cats and Dogs in Switzerland. Vet. Microbiol. 2018, 216, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Saputra, S.; Jordan, D.; Mitchell, T.; Wong, H.S.; Abraham, R.J.; Kidsley, A.; Turnidge, J.; Trott, D.J.; Abraham, S. Antimicrobial Resistance in Clinical Escherichia coli Isolated From Companion Animals in Australia. Vet. Microbiol. 2017, 211, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Weese, J.S.; Blondeau, J.; Boothe, D.; Guardabassi, L.G.; Gumley, N.; Papich, M.; Jessen, L.R.; Lappin, M.; Rankin, S.; Westropp, J.L.; et al. International Society for Companion Animal Infectious Diseases (ISCAID) Guidelines for the Diagnosis and Management of Bacterial Urinary Tract Infections in Dogs and Cats. Vet. J. 2019, 247, 8–25. [Google Scholar] [CrossRef]
- Weese, J.S.; Blondeau, J.M.; Boothe, D.; Breitschwerdt, E.B.; Guardabassi, L.; Hillier, A.; Lloyd, D.H.; Papich, M.G.; Rankin, S.C.; Turnidge, J.D.; et al. Antimicrobial Use Guidelines for Treatment of Urinary Tract Disease in Dogs and Cats: Antimicrobial Guidelines Working Group of the International Society for Companion Animal Infectious Diseases. Vet. Med. Int. 2011, 2011, 263768. [Google Scholar] [CrossRef] [Green Version]
- Rzewuska, M.; Czopowicz, M.; Kizerwetter-Swida, M.; Chrobak, D.; Blaszczak, B.; Binek, M. Multidrug Resistance in Escherichia coli Strains Isolated from Infections in Dogs and Cats in Poland (2007–2013). Sci. World J. 2015, 2015, 408205. [Google Scholar] [CrossRef] [Green Version]
- Thungrat, K.; Price, S.B.; Carpenter, D.M.; Boothe, D.M. Antimicrobial Susceptibility Patterns of Clinical Escherichia coli Isolates from Dogs and Cats in the United States: January 2008 through January 2013. Vet. Microbiol. 2015, 179, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.; Reid-Smith, R.J.; Prescott, J.F.; Bonnett, B.N.; Mcewen, S.A. Occurrence of Antimicrobial Resistant Bacteria in Healthy Dogs and Cats Presented to Private Veterinary Hospitals in Southern Ontario: A Preliminary Study. Can. Vet. J. 2009, 50, 1047. [Google Scholar] [PubMed]
- Surgers, L.; Boyd, A.; Girard, P.M.; Arlet, G.; Decré, D. Biofilm Formation by ESBL-Producing Strains of Escherichia coli and Klebsiella pneumoniae. Int. J. Med. Microbiol. 2018, 309, 13–18. [Google Scholar] [CrossRef]
- Melo, L.C.; Oresco, C.; Leigue, L.; Netto, H.M.; Melville, P.A.; Benites, N.R.; Saras, E.; Haenni, M.; Lincopan, N.; Madec, J.Y. Prevalence and Molecular Features of ESBL/pAmpC-Producing Enterobacteriaceae in Healthy and Diseased Companion Animals in Brazil. Vet. Microbiol. 2018, 221, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Maeyama, Y.; Taniguchi, Y.; Hayashi, W.; Ohsaki, Y.; Osaka, S.; Koide, S.; Tamai, K.; Nagano, Y.; Arakawa, Y.; Nagano, N. Prevalence of ESBL/AmpC Genes and Specific Clones among the Third-Generation Cephalosporin-Resistant Enterobacteriaceae from Canine and Feline Clinical Specimens in Japan. Vet. Microbiol. 2018, 216, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogaerts, P.; Huang, T.D.; Bouchahrouf, W.; Bauraing, C.; Berhin, C.; El Garch, F.; Glupczynski, Y.; ComPath Study, G. Characterization of ESBL- and AmpC-Producing Enterobacteriaceae from Diseased Companion Animals in Europe. Microb. Drug Resist 2015, 21, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, B.W.; Nayak, R.; Foley, S.L.; Kweon, O.; Deck, J.; Park, M.; Rafii, F.; Boothe, D.M. Molecular Characterization of Resistance to Extended-Spectrum Cephalosporins in Clinical Escherichia coli Isolates from Companion Animals in the United States. Antimicrob. Agents Chemother. 2011, 55, 5666–5675. [Google Scholar] [CrossRef] [Green Version]
- Abraham, S.; Jordan, D.; Wong, H.S.; Johnson, J.R.; Toleman, M.A.; Wakeham, D.L.; Gordon, D.M.; Turnidge, J.D.; Mollinger, J.L.; Gibson, J.S.; et al. First Detection of Extended-Spectrum Cephalosporin- and Fluoroquinolone-Resistant Escherichia coli in Australian Food-Producing Animals. J.Glob. Antimicrob. Resist. 2015, 3, 273–277. [Google Scholar] [CrossRef]
- Sidjabat, H.E.; Seah, K.Y.; Coleman, L.; Sartor, A.; Derrington, P.; Heney, C.; Faoagali, J.; Nimmo, G.R.; Paterson, D.L. Expansive Spread of IncI1 Plasmids Carrying blaCMY-2 amongst Escherichia coli. Int. J. Antimicrob. Agents 2014, 44, 203–208. [Google Scholar] [CrossRef]
- Abbas, G.; Khan, I.; Mohsin, M.; Sajjad Ur, R.; Younas, T.; Ali, S. High Rates of CTX-M Group-1 Extended-Spectrum Beta-Lactamases Producing Escherichia coli from Pets and Their Owners in Faisalabad, Pakistan. Infect. Drug Resist. 2019, 12, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Liu, H.; Li, Y.; Hao, C. High Prevalence of Beta-Lactamase and Plasmid-Mediated Quinolone Resistance Genes in Extended-Spectrum Cephalosporin-Resistant Escherichia coli from Dogs in Shanxi, China. Front. Microbiol. 2016, 7, 1843. [Google Scholar] [CrossRef] [Green Version]
- Rocha-Gracia, R.C.; Cortes-Cortes, G.; Lozano-Zarain, P.; Bello, F.; Martinez-Laguna, Y.; Torres, C. Faecal Escherichia coli Isolates from Healthy Dogs Harbour CTX-M-15 and CMY-2 Beta-Lactamases. Vet. J. 2015, 203, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Igbinosa, I.H. Prevalence and Detection of Antibiotic-Resistant Determinant in Salmonella Isolated from Food-Producing Animals. Trop. Anim. Health Prod. 2015, 47, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; He, T.; Han, J.; Wang, J.; Foley, S.L.; Yang, G.; Wan, S.; Shen, J.; Wu, C. Prevalence of ESBLs and PMQR Genes in Fecal Escherichia coli Isolated from the Non-Human Primates in Six Zoos in China. Vet. Microbiol. 2012, 159, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J.S.; Cobbold, R.N.; Heisig, P.; Sidjabat, H.E.; Kyaw-Tanner, M.T.; Trott, D.J. Identification of qnr and aac(6’)-1b-cr Plasmid-Mediated Fluoroquinolone Resistance Determinants in Multidrug-Resistant Enterobacter spp. Isolated from Extraintestinal Infections in Companion Animals. Vet. Microbiol. 2010, 143, 329–336. [Google Scholar] [CrossRef]
- Wang, J.; Zhi, C.P.; Chen, X.J.; Guo, Z.W.; Liu, W.L.; Luo, J.; Huang, X.Y.; Zeng, L.; Huang, J.W.; Xia, Y.B.; et al. Characterization of oqxAB in Escherichia coli Isolates from Animals, Retail Meat, and Human Patients in Guangzhou, China. Front. Microbiol. 2017, 8, 1982. [Google Scholar] [CrossRef] [Green Version]
- Elmi, S.Y.K.; Ashour, M.S.; Alsewy, F.Z.; Abd El Moez Azzam, N.F. Phenotypic and Genotypic Detection of Extended Spectrum Beta-Lactamases among Escherichia coli and Klebsiella pneumoniae Isolates from Type 2 Diabetic Patients with Urinary Tract Infections. Afr. Health Sci. 2021, 21, 497–504. [Google Scholar] [CrossRef]
- Carniel, E. The Yersinia High-Pathogenicity Island: An Iron-Uptake Island. Microbes Infect. 2001, 3, 561–569. [Google Scholar] [CrossRef]
- Cespedes, S.; Saitz, W.; Del Canto, F.; De la Fuente, M.; Quera, R.; Hermoso, M.; Munoz, R.; Ginard, D.; Khorrami, S.; Giron, J.; et al. Genetic Diversity and Virulence Determinants of Escherichia coli Strains Isolated from Patients with Crohn’s Disease in Spain and Chile. Front. Microbiol. 2017, 8, 639. [Google Scholar] [CrossRef]
- Schubert, S.; Cuenca, S.; Fischer, D.; Heesemann, J. High-Pathogenicity Island of Yersinia Pestis in Enterobacteriaceae Isolated from Blood Cultures and Urine Samples: Prevalence and Functional Expression. J. Infect. Dis. 2000, 182, 1268–1271. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, R.; Sharma, M.C.; Singh, B.P. Buffalo Calf Health Care in Commercial Dairy Farms: A Field Study in Uttar Pradesh (India). Livest. Res. Rural. Dev. 2007, 19, 8. [Google Scholar]
- Persoons, D.; Bollaerts, K.; Smet, A.; Herman, L.; Heyndrickx, M.; Martel, A.; Butaye, P.; Catry, B.; Haesebrouck, F.; Dewulf, J. The Importance of Sample Size in the Determination of a Flock-Level Antimicrobial Resistance Profile for Escherichia coli in Broilers. Microb. Drug Resist. 2011, 17, 513–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, J.A.; Chong, W.L.; Gordon, D.M. Genetic Structure, Antimicrobial Resistance and Frequency of Human Associated Escherichia coli Sequence Types among Faecal Isolates from Healthy Dogs and Cats Living in Canberra, Australia. PLoS ONE 2019, 14, e0212867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-second Informational Supplement; CLSI Document M100-S22; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.M.; Motoi, Y.; Sato, M.; Maruyama, A.; Watanabe, H.; Fukumoto, Y.; Shimamoto, T. Zoo Animals as Reservoirs of Gram-Negative Bacteria Harboring Integrons and Antimicrobial Resistance Genes. Appl. Environ. Microbiol. 2007, 73, 6686–6690. [Google Scholar] [CrossRef] [Green Version]
- Bert, F.; Branger, C.; Lambert-Zechovsky, N. Identification of PSE and OXA Beta-Lactamase Genes in Pseudomonas Aeruginosa Using PCR-restriction Fragment Length Polymorphism. J. Antimicrob. Chemother. 2002, 50, 11–18. [Google Scholar] [CrossRef]
- Liao, X.P.; Xia, J.; Yang, L.; Li, L.; Sun, J.; Liu, Y.H.; Jiang, H.X. Characterization of CTX-M-14-Producing Escherichia coli from Food-Producing Animals. Front. Microbiol. 2015, 6, 1136. [Google Scholar] [CrossRef] [Green Version]
- Saenz, Y.; Brinas, L.; Dominguez, E.; Ruiz, J.; Zarazaga, M.; Vila, J.; Torres, C. Mechanisms of Resistance in Multiple-Antibiotic-resistant Escherichia coli Strains of Human, Animal, and Food Origins. Antimicrob. Agents Chemother. 2004, 48, 3996–4001. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, L.; Zhou, Y.; Miao, Z. Prevalence and Characterization of Virulence Genes in Escherichia coli Isolated from Piglets Suffering Post-Weaning Diarrhoea in Shandong Province, China. Vet. Med. Sci. 2020, 6, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.B.; Zanardo, L.G.; Galvao, N.N.; Carvalho, I.A.; Nero, L.A.; Moreira, M.A. Escherichia coli from Clinical Mastitis: Serotypes and Virulence Factors. J. Vet. Diagn. Invest 2011, 23, 1146–1152. [Google Scholar] [CrossRef] [Green Version]
- Gioia-Di Chiacchio, R.M.; Cunha, M.P.V.; de Sa, L.R.M.; Davies, Y.M.; Pereira, C.B.P.; Martins, F.H.; Munhoz, D.D.; Abe, C.M.; Franzolin, M.R.; Dos Santos, L.F.; et al. Novel Hybrid of Typical Enteropathogenic Escherichia coli and Shiga-Toxin-Producing E. coli (tEPEC/STEC) Emerging From Pet Birds. Front. Microbiol. 2018, 9, 2975. [Google Scholar] [CrossRef] [PubMed]
- Picco, N.Y.; Alustiza, F.E.; Bellingeri, R.V.; Grosso, M.C.; Motta, C.E.; Larriestra, A.J.; Vissio, C.; Tiranti, K.I.; Terzolo, H.R.; Moreira, A.R.; et al. Molecular Screening of Pathogenic Escherichia coli Strains Isolated from Dairy Neonatal Calves in Cordoba Province, Argentina. Rev. Argent Microbiol. 2015, 47, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]




| Clinical Status | ETEC (n = 26) | EPEC (n = 8) | EIEC (n = 2) |
|---|---|---|---|
| healthy | O6:K15(4), O78:K80(2), O25:K19(3), O8:K40(2), O9:K9(1), O20:K17(1), O7:K1(1) | O114:K90(1), O128:K67(1), O125:K70(2), | O29:K?(1), O124:K7(1) |
| diarrhea | O6:K15(4), O78:K80(2), O25:K19(2), O8:K40(3), O15:K?(1) | O114:K90(2), O128:K67(1), O142:K86(1) | - |
| Antibiotic Classes | Number (%) of Resistant Isolates | |
|---|---|---|
| ESBL (+) | ESBL (−) | |
| (n = 13) | (n = 23) | |
| β-lactames | 8 (61.5) | 10 (43.5) |
| Tetracyclines | 12 (92.3) | 17 (73.9) |
| Quinolones | 4 (3.1) | 2 (8.7) |
| Penicillins | 10 (76.9) | 12 (52.2) |
| Aminoglycosides | 4 (30.8) | 8 (34.8) |
| Carbapenems | 0 | 0 |
| Antibiotics Resistance Phenotype | Characteristics of Strains | p Value 5 | ||||
|---|---|---|---|---|---|---|
| n-Pr 1 | Resistance Genes | n-Gp 2 | P+/G− 3 | P−/G+ 4 | ||
| CRO | 18 | blaCTX-M | 48 | 1 | 31 | 0.010 * |
| blaSHV | 32 | 11 | 25 | 0.166 | ||
| blaOXA | 2 | 1 | 1 | 0.260 | ||
| CEX | 39 | blaCTX-M | 48 | 3 | 12 | 0.128 |
| blaSHV | 32 | 25 | 18 | 0.738 | ||
| blaOXA | 2 | 0 | 2 | 0.007 ** | ||
| FEP | 2 | blaCTX-M | 48 | 0 | 46 | 0.496 |
| blaSHV | 32 | 1 | 31 | 1.000 | ||
| blaOXA | 2 | 1 | 1 | 1.000 | ||
| GM | 22 | aaC4 | 20 | 13 | 11 | 0.889 |
| 19 | qnrA | 9 | 5 | 4 | 0.786 | |
| qnrB | 6 | 2 | 4 | 0.067 | ||
| ENR | qnrS | 15 | 7 | 3 | 0.665 | |
| oqxA | 32 | 9 | 22 | 0.446 | ||
| aac (6′)-Ib-cr | 35 | 8 | 24 | 0.325 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, L.; Zhao, X.; Li, R.; Han, Y.; Hao, G.; Wang, G.; Sun, S. Companion Animals as Potential Reservoirs of Antibiotic Resistant Diarrheagenic Escherichia coli in Shandong, China. Antibiotics 2022, 11, 828. https://doi.org/10.3390/antibiotics11060828
Cui L, Zhao X, Li R, Han Y, Hao G, Wang G, Sun S. Companion Animals as Potential Reservoirs of Antibiotic Resistant Diarrheagenic Escherichia coli in Shandong, China. Antibiotics. 2022; 11(6):828. https://doi.org/10.3390/antibiotics11060828
Chicago/Turabian StyleCui, Lulu, Xiaonan Zhao, Ruibo Li, Yu Han, Guijuan Hao, Guisheng Wang, and Shuhong Sun. 2022. "Companion Animals as Potential Reservoirs of Antibiotic Resistant Diarrheagenic Escherichia coli in Shandong, China" Antibiotics 11, no. 6: 828. https://doi.org/10.3390/antibiotics11060828
APA StyleCui, L., Zhao, X., Li, R., Han, Y., Hao, G., Wang, G., & Sun, S. (2022). Companion Animals as Potential Reservoirs of Antibiotic Resistant Diarrheagenic Escherichia coli in Shandong, China. Antibiotics, 11(6), 828. https://doi.org/10.3390/antibiotics11060828