
Antimicrobial Resistance and Biofilms Underlying Catheter-Related Bloodstream Coinfection by Enterobacter cloacae Complex and Candida parapsilosis

 , ,
, ,  , ,
, ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. CRBSI Aetiological Agents
2.2. Antimicrobial Agent Resistance
2.3. Central Venous Catheter Colonization and CRBSI
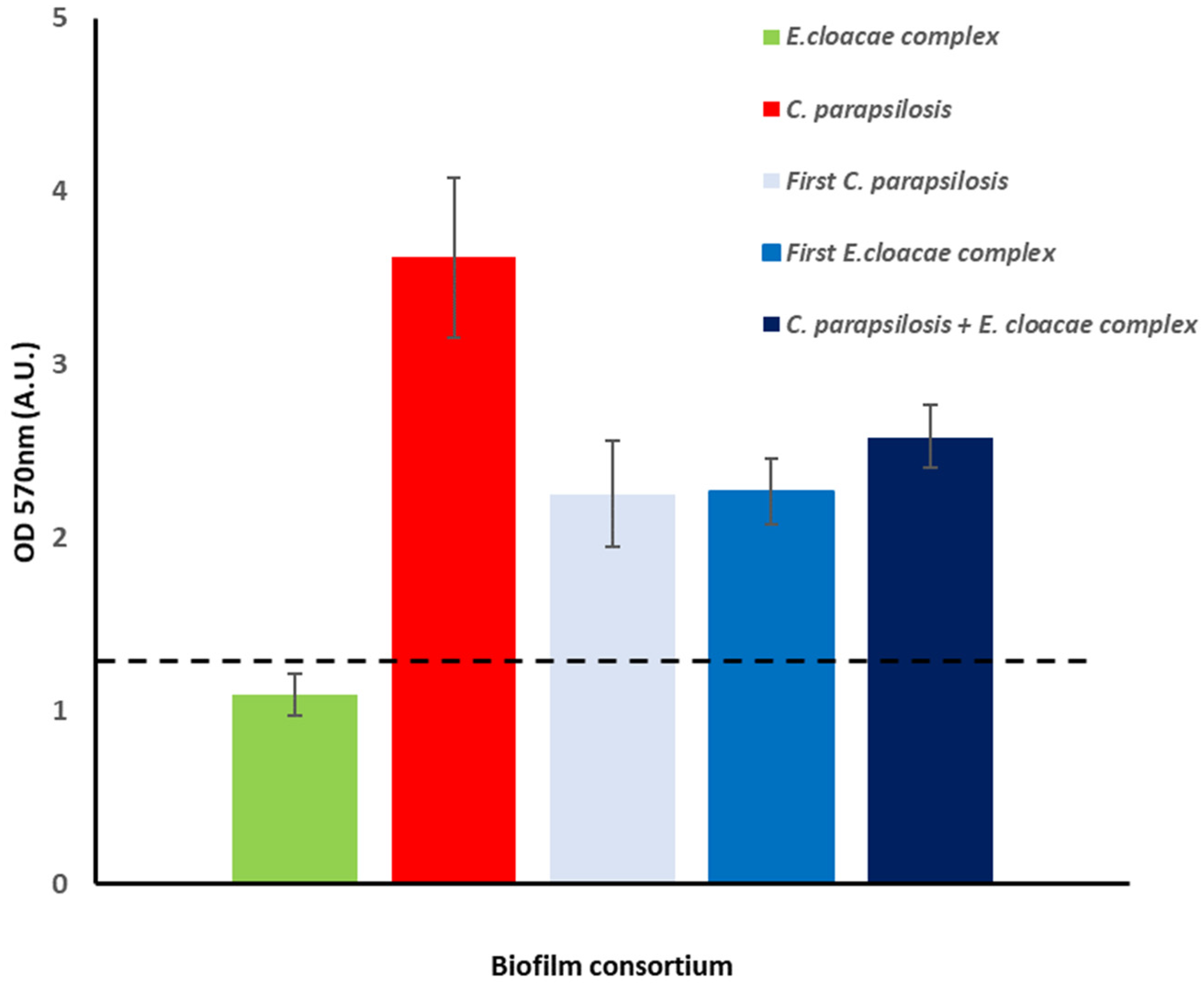
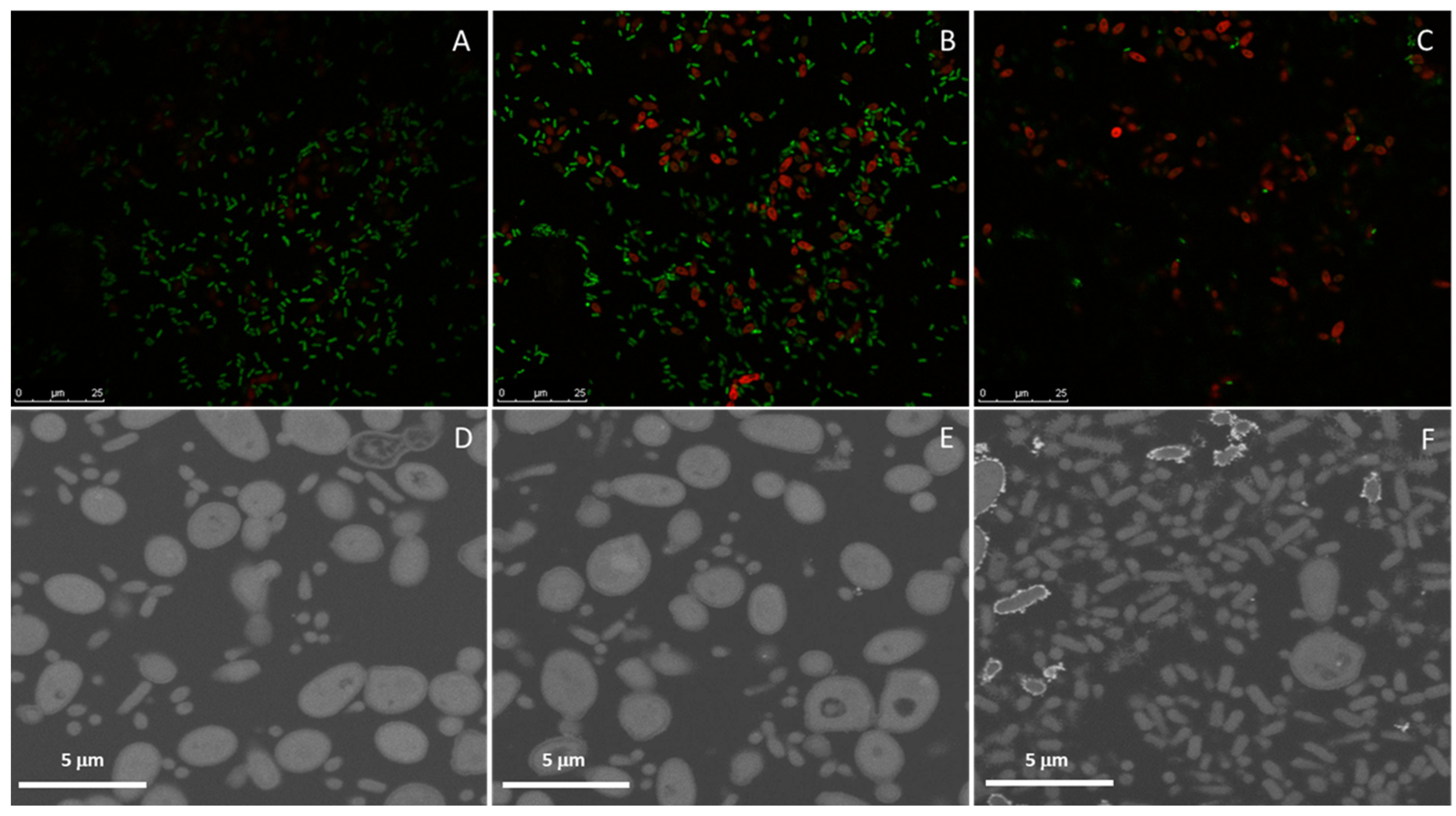
2.4. Dual Biofilms of E. cloacae Complex (E. bugandensis) and C. parapsilosis
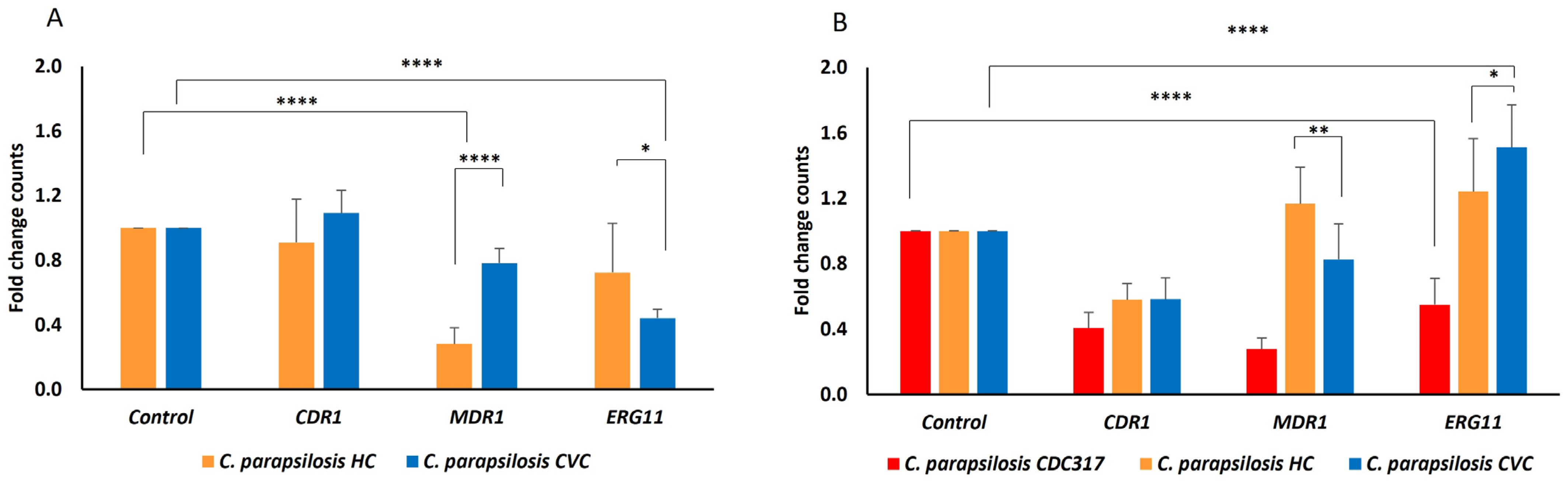
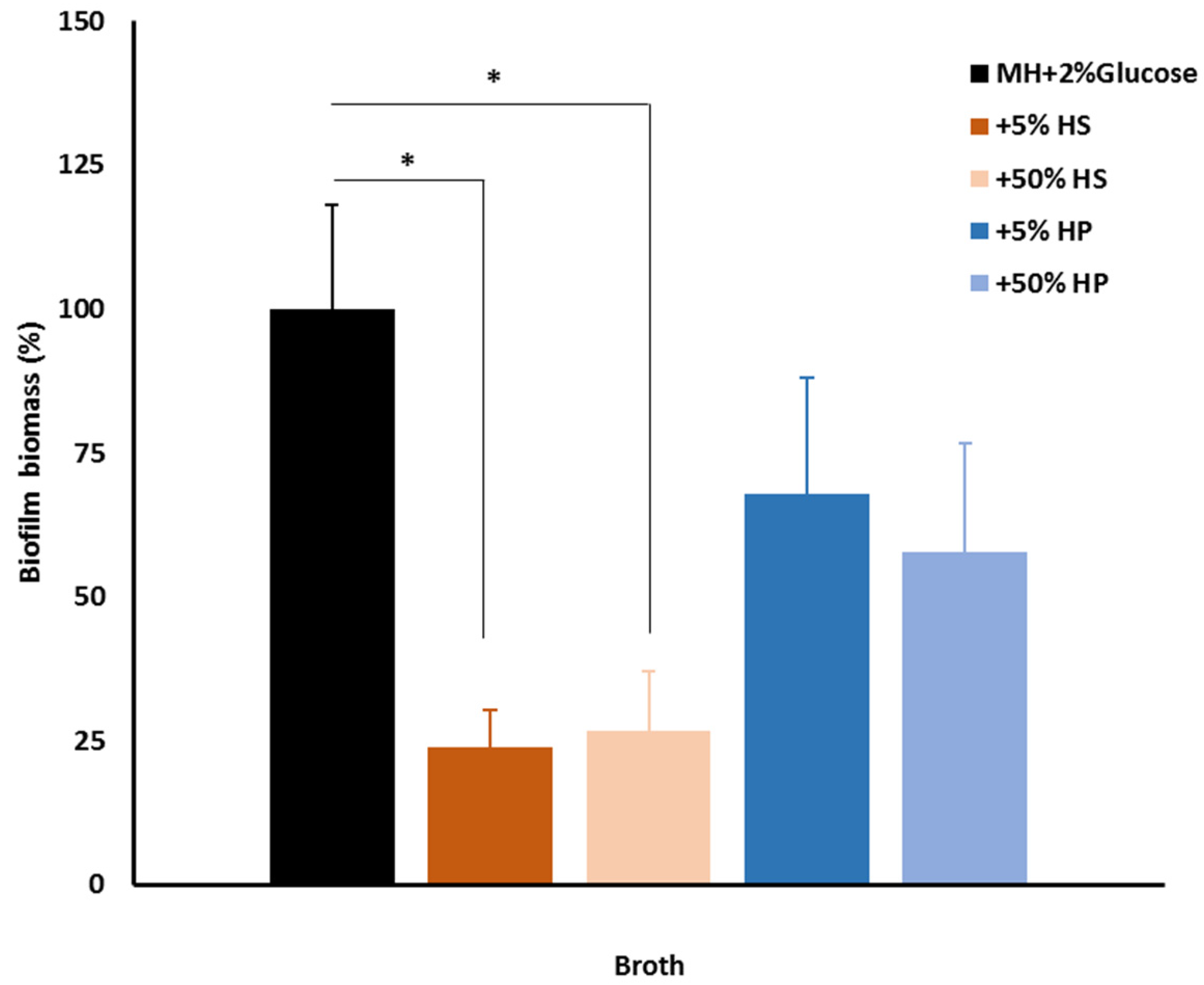
2.5. Host Factors and Biofilm
3. Discussion
4. Materials and Methods
4.1. Microorganism Isolation and Culture
4.2. Antimicrobial Susceptibility Tests
4.3. Biofilm Assay
4.4. Biofilms and Host Factors
4.5. Analysis of Biofilm Assembled In Vitro on Different Surfaces
- (i)
- Scanning electron microscopy (SEM). Samples were washed with PBS and fixed with 4% paraformaldehyde (Merck, Darmstadt, Germany) in PBS for 30 min at room temperature (RT) protected from light. The fixative was removed, and the samples were washed twice in PBS for 10 min and post-fixed with 1% osmium tetroxide (EMS, Hatfield, PA, USA) in the same buffer for 90 min on ice protected from the light. Then, the samples were washed twice for 10 min at RT with PBS and twice with deionized water. Dehydration was performed at RT using serial dilutions of ethanol as follows: once in 50%, 70%, 80% and 95% ethanol for 30 min and twice in 100% ethanol for 30 min each. Samples were then trimmed (CVC was cut longitudinally), allowed to dry at RT, mounted on top of double-sided carbon tape (CVC was mounted in such a way that both inner and outer surfaces were visible), coated with 20 nm thick gold-palladium film using a sputter coater QISOT ES (Quorum Technologies, Laughton, UK) and analyzed under a scanning electron microscope, JSM-7100F (JEOL, Tokyo, Japan) using secondary electron detector.
- (ii)
- Focused ion beam scanning electron microscopy (FIB-SEM) tomography. Samples were prepared as described previously for SEM until the second incubation with 100% ethanol. Then, Epoxy 812 resin (EMS) was added with the samples left uncapped overnight in a chemical chamber. The resin was then replaced by a new batch and further incubated for 3 h. Then, the sample and one drop of resin were compressed between 2 sheets of ACLAR film (EMS) allowing the sample to be encased in a thin resin layer. Samples were allowed to polymerize at 65 °C. FIB-SEM tomography was performed with an FEI HELIOS G4 instrument (Thermo Fisher Scientific, Waltham, MA, USA). Slicing was performed with a Ga+ ion beam accelerated at 30 kV and a current of 1 nA, while imaging was performed with an electron beam accelerated at 5 kV, a current of 1.6 nA and using the backscattered electron detector. A slice thickness of 30 nm was used. The acquisition operation was controlled through the Auto Slice & View 4.0 software package and reconstruction was carried out with Avizo Fire software package. Individual fungi and bacteria were manually labelled for intensity thresholding and 3D volume reconstruction.
- (iii)
- Fluorescence in situ hybridization (FISH)/laser scanning confocal microscopy. For biofilms assembled on glass coverslips, FISH was used to assess the distribution of fungi (C. parapsilosis-probe PF2) and bacteria (E. cloacae complex-probe EUB) using 5‘-labeled oligonucleotide probe fluorochromes FITC and Cy3, respectively (Biomers.net, Ulm, Germany). Non-sense probes labelled with the same fluorochromes were used as control. The probe sequences were previously described [56,57]. Samples were fixed as described for SEM, washed with PBS, permeabilized with 200 U/mL of lyticase (Sigma-Aldrich) followed by 10 mg/mL of lysozyme in hybridization buffer pH 8 (20 mM Tris-HCl pH8, 0.9 M sodium chloride, 20% formamide, 0.01% sodium dodecyl sulfate all from Sigma-Aldrich) for 15 min at 30 °C and incubated in a humidified chamber with 1 µM PF2 probe in the hybridization buffer for 3 h at 45 °C. After the first hybridization step, the unbound probe was washed off with a 45 °C pre-warmed washing buffer (20 mM Tris-HCl pH8, 0.212 M sodium chloride, 5 mM EDTA, 0.01% SDS) and water for 10 min each. Hybridization with 1 µM with EUB probe was performed for 2 h at 45 °C; the unbound probe was removed as previously. Coverslips were mounted by inversion on a drop of fluorescence-mounting medium previously placed on a glass slide. Samples were stored at 4 °C, protected from light, until visualization through a confocal microscope (Leica, SP2, Wetzlar, Germany) under an immersion objective of ×63 and an ocular objective of ×10.
4.6. Analysis of CVC Colonization
4.7. DNA Extraction and WGS
4.8. Genome Characterization
4.9. Isolation of RNA from C. parapsilosis
4.10. Evaluation of Gene Expression Related to FLU Resistance in C. parapsilosis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Centre for Disease Prevention and Control. Healthcare-Associated Infections Acquired in Intensive Care Units. In ECDC. Annual Epidemiological Report for 2016; ECDC Stockholm: Stockholm, Sweden, 2018. [Google Scholar]
- Selby, L.M.; Rupp, M.E.; Cawcutt, K.A. Prevention of Central-Line Associated Bloodstream Infections: 2021 Update. Infect. Dis. Clin. N. Am. 2021, 35, 841–856. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Granda, M.J.; Carrillo, C.S.; Rabadán, P.M.; Valerio, M.; Olmedo, M.; Muñoz, P.; Bouza, E. Increase in the Frequency of Catheter-Related Bloodstream Infections during the COVID-19 Pandemic: A Plea for Control. J. Hosp. Infect. 2022, 119, 149–154. [Google Scholar] [CrossRef]
- Qian, Z.; Zhang, S.; Li, N.; Ma, W.; Zhang, K.; Song, F.; Zheng, C.; Zhong, L.; Wang, Y.; Cai, J.; et al. Risk Factors for and Clinical Outcomes of Polymicrobial Acinetobacter baumannii Bloodstream Infections. BioMed Res. Int. 2022, 2022, 5122085. [Google Scholar] [CrossRef]
- Li, S.; Duan, W.; Lei, Y.; Wang, Z.; Fu, C.; He, L.; Shen, Z.; Li, M.; Chen, Y.; Huang, Y. Effects of Lipid Emulsions on the Formation of Escherichia coli–Candida albicans Mixed-Species Biofilms on PVC. Sci. Rep. 2021, 11, 16929. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.; Borges, V.; Nascimento, M.; Martins, F.; Pessanha, M.A.; Faria, I.; Rodrigues, J.; Matias, R.; Gomes, J.P.; Jordao, L. Insights on Catheter-Related Bloodstream Infections: A Prospective Observational Study on the Catheter Colonization and Multidrug Resistance. J. Hosp. Infect. 2022, 123, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Gominet, M.; Compain, F.; Beloin, C.; Lebeaux, D. Central Venous Catheters and Biofilms: Where Do We Stand in 2017? APMIS 2017, 125, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Resistance Surveillance in Europe 2022–2020 Data. Available online: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-surveillance-europe-2022-2020-data (accessed on 4 August 2022).
- del Pozo, J.L. Biofilm-Related Disease. Expert Rev. Anti-Infect. Ther. 2018, 16, 51–65. [Google Scholar] [CrossRef]
- Yaita, K.; Gotoh, K.; Nakano, R.; Iwahashi, J.; Sakai, Y.; Horita, R.; Yano, H.; Watanabe, H. Biofilm-Forming by Carbapenem Resistant Enterobacteriaceae May Contribute to the Blood Stream Infection. Int. J. Mol. Sci. 2019, 20, 5954. [Google Scholar] [CrossRef]
- van der Zwet, W.C.; Nijsen, I.E.J.; Jamin, C.; van Alphen, L.B.; von Wintersdorff, C.J.H.; Demandt, A.M.P.; Savelkoul, P.H.M. Role of the Environment in Transmission of Gram-Negative Bacteria in Two Consecutive Outbreaks in a Haematology-Oncology Department. Infect. Prev. Pract. 2022, 4, 100209. [Google Scholar] [CrossRef]
- Hendrickx, A.P.A.; Debast, S.; Pérez-Vázquez, M.; Schoffelen, A.F.; Notermans, D.W.; Landman, F.; Wielders, C.C.H.; Cañada Garcia, J.E.; Flipse, J.; de Haan, A.; et al. A Genetic Cluster of MDR Enterobacter cloacae Complex ST78 Harbouring a Plasmid Containing Bla VIM-1 and Mcr-9 in the Netherlands. Jac-Antimicrob. Resist. 2021, 3, dlab046. [Google Scholar] [CrossRef]
- Zhu, X.; Li, P.; Qian, C.; Liu, H.; Lin, H.; Zhang, X.; Li, Q.; Lu, J.; Lin, X.; Xu, T.; et al. Prevalence of Aminoglycoside Resistance Genes and Molecular Characterization of a Novel Gene, Aac (3)IIg, among Clinical Isolates of the Enterobacter cloacae Complex from a Chinese Teaching Hospital. Antimicrob. Agents Chemother. 2020, 64, e00852-20. [Google Scholar] [CrossRef] [PubMed]
- Matic, T.; Novak, M.; Braovac, D.; Vinkovic, M.; Cicak, A.M.; Milosevic, M.; Galic, S.; Cvitkovic, M.; Rubic, F.; Ille, V.; et al. Characteristics, Risk Factors and Predictors for Candidemia in the Pediatric Intensive Care Unit at the University Hospital Centre Zagreb in Croatia: A 9-Year Retrospective Study. Pediatr. Infect. Dis. J. 2021, 40, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Riera, F.O.; Caeiro, J.P.; Angiolini, S.C.; Vigezzi, C.; Rodriguez, E.; Icely, P.A.; Sotomayor, C.E. Invasive Candidiasis: Update and Current Challenges in the Management of This Mycosis in South America. Antibiotics 2022, 11, 877. [Google Scholar] [CrossRef] [PubMed]
- Miyake, A.; Gotoh, K.; Iwahashi, J.; Togo, A.; Horita, R.; Miura, M.; Kinoshita, M.; Ohta, K.; Yamashita, Y.; Watanabe, H. Characteristics of Biofilms Formed by C. parapsilosis Causing an Outbreak in a Neonatal Intensive Care Unit. J. Fungi 2022, 8, 700. [Google Scholar] [CrossRef]
- Healthcare-Associated Infections in Intensive Care Units-Annual Epidemiological Report for 2017. Available online: https://www.ecdc.europa.eu/en/publications-data/healthcare-associated-infections-intensive-care-units-annual-epidemiological-1 (accessed on 4 August 2022).
- Tavanti, A.; Davidson, A.D.; Gow, N.A.R.; Maiden, M.C.J.; Odds, F.C. Candida orthopsilosis and Candida metapsilosis Spp. Nov. to Replace Candida parapsilosis Groups II and III. J. Clin. Microbiol. 2005, 43, 284–292. [Google Scholar] [CrossRef]
- Doijad, S.; Imirzalioglu, C.; Yao, Y.; Pati, N.B.; Falgenhauer, L.; Hain, T.; Foesel, B.U.; Abt, B.; Overmann, J.; Mirambo, M.M.; et al. Enterobacter bugandensis sp. Nov., Isolated from Neonatal Blood. Int. J. Syst. Evol. Microbiol. 2016, 66, 968–974. [Google Scholar] [CrossRef]
- Singh, N.K.; Bezdan, D.; Checinska Sielaff, A.; Wheeler, K.; Mason, C.E.; Venkateswaran, K. Multi-Drug Resistant Enterobacter bugandensis Species Isolated from the International Space Station and Comparative Genomic Analyses with Human Pathogenic Strains. BMC Microbiol. 2018, 18, 175. [Google Scholar] [CrossRef]
- Pati, N.B.; Doijad, S.P.; Schultze, T.; Mannala, G.K.; Yao, Y.; Jaiswal, S.; Ryan, D.; Suar, M.; Gwozdzinski, K.; Bunk, B.; et al. Enterobacter bugandensis: A Novel Enterobacterial Species Associated with Severe Clinical Infection. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Nishida, S.; Matsunaga, N.; Kamimura, Y.; Ishigaki, S.; Furukawa, T.; Ono, Y. Emergence of Enterobacter Cloacae Complex Co-Producing IMP-10 and CTX-M, and Klebsiella pneumoniae Producing VIM-1 in Clinical Isolates in Japan. Microorganisms 2020, 8, 1816. [Google Scholar] [CrossRef]
- Kananizadeh, P.; Oshiro, S.; Watanabe, S.; Iwata, S.; Kuwahara-Arai, K.; Shimojima, M.; Ogawa, M.; Tada, T.; Kirikae, T. Emergence of Carbapenem-Resistant and Colistin-Susceptible Enterobacter cloacae Complex Co-Harboring Bla IMP-1 and Mcr-9 in Japan. BMC Infect. Dis. 2020, 20, 282. [Google Scholar] [CrossRef] [Green Version]
- Bolourchi, N.; Giske, C.G.; Nematzadeh, S.; Mirzaie, A.; Abhari, S.S.; Solgi, H.; Badmasti, F. Comparative Resistome and Virulome Analysis of Clinical NDM-1-Producing Carbapenem-Resistant Enterobacter cloacae Complex. J. Glob. Antimicrob. Resist. 2022, 28, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Notley-McRobb, L.; King, T.; Ferenci, T. rpoS Mutations and Loss of General Stress Resistance in Escherichia coli Populations as a Consequence of Conflict between Competing Stress Responses. J. Bacteriol. 2002, 184, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Nazir, F.; Ibrahim, M.; Zaman, G.; Hussain, A.; Yar, A.M.; Bo, Z. Genetic Diversity and Functional Analysis of Sigma Factors in Enterobacter cloacae Complex Resourced from Various Niche. Evol. Bioinform. Online 2018, 14, 1176934318754878. [Google Scholar] [CrossRef] [PubMed]
- Tóth, R.; Nosek, J.; Mora-Montes, H.M.; Gabaldon, T.; Bliss, J.M.; Nosanchuk, J.D.; Turner, S.A.; Butler, G.; Vágvölgyi, C.; Gácser, A. Candida parapsilosis: From Genes to the Bedside. Clin. Microbiol. Rev. 2019, 32, e00111-18. [Google Scholar] [CrossRef] [PubMed]
- Doorley, L.A.; Rybak, J.M.; Berkow, E.L.; Zhang, Q.; Morschhäuser, J.; Rogers, P.D. Candida parapsilosis Mdr1B and Cdr1B Are Drivers of Mrr1-Mediated Clinical Fluconazole Resistance. Antimicrob. Agents Chemother. 2022, 66, e00289-22. [Google Scholar] [CrossRef]
- Díaz-García, J.; Gómez, A.; Alcalá, L.; Reigadas, E.; Sánchez-Carrillo, C.; Pérez-Ayala, A.; Gómez-García de la Pedrosa, E.; González-Romo, F.; Merino-Amador, P.; Cuétara, M.S.; et al. Evidence of Fluconazole-Resistant Candida parapsilosis Genotypes Spreading across Hospitals Located in Madrid, Spain and Harboring the Y132F ERG11p Substitution. Antimicrob. Agents Chemother. 2022, 66, e00710-22. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, Y.J.; Yong, D.; Byun, J.H.; Kim, T.S.; Chang, Y.S.; Choi, M.J.; Byeon, S.A.; Won, E.J.; Kim, S.H.; et al. Fluconazole-Resistant Candida parapsilosis Bloodstream Isolates with Y132F Mutation in ERG11 Gene, South Korea. Emerg. Infect. Dis. 2018, 24, 1768–1770. [Google Scholar] [CrossRef]
- Martini, C.; Torelli, R.; de Groot, T.; de Carolis, E.; Morandotti, G.A.; de Angelis, G.; Posteraro, B.; Meis, J.F.; Sanguinetti, M. Prevalence and Clonal Distribution of Azole-Resistant Candida parapsilosis Isolates Causing Bloodstream Infections in a Large Italian Hospital. Front. Cell. Infect. Microbiol. 2020, 10, 232. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; di Bonaventura, G.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of Biofilm in Microtiter Plates: Overview of Testing Conditions and Practical Recommendations for Assessment of Biofilm Production by Staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Ramage, G.; VandeWalle, K.; López-Ribot, J.L.; Wickes, B.L. The Filamentation Pathway Controlled by the Efg1 Regulator Protein Is Required for Normal Biofilm Formation and Development in Candida albicans. FEMS Microbiol. Lett. 2002, 214, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Metwalli, K.H.; Khan, S.A.; Krom, B.P.; Jabra-Rizk, M.A. Streptococcus mutans, Candida albicans, and the Human Mouth: A Sticky Situation. PLoS Pathog. 2013, 9, e1003616. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Hu, D.; He, J.; Zou, L.; Chen, Z.; Li, M. Effect of LongZhang Gargle on Dual-Species Biofilm of Candida albicans and Streptococcus mutans. BioMed Res. Int. 2021, 2021, 6654793. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.M.; Jabra-Rizk, M.A.; Scheper, M.A.; Leid, J.G.; Costerton, J.W.; Shirtliff, M.E. Microbial Interactions and Differential Protein Expression in Staphylococcus aureus -Candida albicans Dual-Species Biofilms. FEMS Immunol. Med. Microbiol. 2010, 59, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.P.; Cowley, E.S.; Nobile, C.J.; Hartooni, N.; Newman, D.K.; Johnson, A.D. Anaerobic Bacteria Grow within Candida albicans Biofilms and Induce Biofilm Formation in Suspension Cultures. Curr. Biol. 2014, 24, 2411–2416. [Google Scholar] [CrossRef] [PubMed]
- Yamin, D.H.; Husin, A.; Harun, A. Risk Factors of Candida parapsilosis Catheter-Related Bloodstream Infection. Front. Public Health 2021, 9, 631865. [Google Scholar] [CrossRef] [PubMed]
- Meher, M.K.; Poluri, K.M. Bifunctional Dalteparin/Enoxaparin Coated Nanosilver Formulation to Prevent Bloodstream Infections during Hemodialysis. Carbohydr. Polym. 2022, 291, 119546. [Google Scholar] [CrossRef]
- Sancho, S.; Artero, A.; Zaragoza, R.; Camarena, J.J.; González, R.; Nogueira, J.M. Impact of Nosocomial Polymicrobial Bloodstream Infections on the Outcome in Critically Ill Patients. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1791–1796. [Google Scholar] [CrossRef]
- Fukushima, S.; Hagiya, H.; Fujita, K.; Kamiyama, S.; Yamada, H.; Kishida, M.; Otsuka, F. Clinical and Microbiological Characteristics of Polymicrobial Bacteremia: A Retrospective, Multicenter Study. Infection 2022, 1–10. [Google Scholar] [CrossRef]
- Kvich, L.; Burmølle, M.; Bjarnsholt, T.; Lichtenberg, M. Do Mixed-Species Biofilms Dominate in Chronic Infections?–Need for in Situ Visualization of Bacterial Organization. Front. Cell. Infect. Microbiol. 2020, 10, 396. [Google Scholar] [CrossRef]
- Girlich, D.; Ouzani, S.; Emeraud, C.; Gauthier, L.; Bonnin, R.A.; le Sache, N.; Mokhtari, M.; Langlois, I.; Begasse, C.; Arangia, N.; et al. Uncovering the Novel Enterobacter cloacae Complex Species Responsible for Septic Shock Deaths in Newborns: A Cohort Study. Lancet Microbe 2021, 2, e536–e544. [Google Scholar] [CrossRef]
- Subtil, J.; Bajanka-Lavado, M.P.; Rodrigues, J.; Duarte, A.; Reis, L.; Nogueira, I.; Jordao, M.L. Cross-Sectional Study of Adenoidal Biofilms in a Paediatric Population and Its Clinical Implications. Otolaryngol. Pol. 2018, 73, 1–5. [Google Scholar] [CrossRef]
- Guembe, M.; Guinea, J.; Marcos-Zambrano, L.; Fernández-Cruz, A.; Peláez, T.; Muñoz, P.; Bouza, E. Is Biofilm Production a Predictor of Catheter-Related Candidemia? Med. Mycol. 2014, 52, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Branchini, M.L.; Pfaller, M.A.; Rhine-Chalberg, J.; Frempong, T.; Isenberg, H.D. Genotypic Variation and Slime Production among Blood and Catheter Isolates of Candida. parapsilosis. J. Clin. Microbiol. 1994, 32, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Thomaz, D.Y.; Melhem, M.D.S.C.; de Almeida Júnior, J.N.; Benard, G.; del Negro, G.M.B. Lack of Efficacy of Echinocandins against High Metabolic Activity Biofilms of Candida parapsilosis Clinical Isolates. Braz. J. Microbiol. 2020, 51, 1129–1133. [Google Scholar] [CrossRef] [PubMed]
- Rickard, A.H.; Gilbert, P.; High, N.J.; Kolenbrander, P.E.; Handley, P.S. Bacterial Coaggregation: An Integral Process in the Development of Multi-Species Biofilms. Trends Microbiol. 2003, 11, 94–100. [Google Scholar] [CrossRef]
- Higgins, M.; Zhang, L.; Ford, R.; Brownlie, J.; Kleidon, T.; Rickard, C.M.; Ullman, A. The Microbial Biofilm Composition on Peripherally Inserted Central Catheters: A Comparison of Polyurethane and Hydrophobic Catheters Collected from Paediatric Patients. J. Vasc. Access 2021, 22, 388–393. [Google Scholar] [CrossRef]
- Pathak, R.; Bierman, S.F.; D’arnaud, P. Inhibition of Bacterial Attachment and Biofilm Formation by a Novel Intravenous Catheter Material Using an in Vitro Percutaneous Catheter Insertion Model. Med. Devices Evid. Res. 2018, 11, 427–432. [Google Scholar] [CrossRef]
- Maki, D.G.; Weise, C.E.; Sarafin, H.W. A Semiquantitative Culture Method for Identifying Intravenous-Catheter-Related Infection. N. Engl. J. Med. 1977, 296, 1305–1309. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 31 January 2022).
- The European Committee on Antimicrobial Susceptibility Testing Breakpoint Tables for Interpretation of MICs for Antifungal Agents. Version 10.0. 2020, pp. 1–8. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Clinical_breakpoints/AFST_BP_v10.0_200204_updatd_links_200924 (accessed on 28 February 2020).
- Harriott, M.M.; Noverr, M.C. Importance of Candida–Bacterial Polymicrobial Biofilms in Disease. Trends Microbiol. 2011, 19, 557–563. [Google Scholar] [CrossRef]
- Bandeira, M.; Borges, V.; Gomes, J.P.; Duarte, A.; Jordao, L. Insights on Klebsiella pneumoniae Biofilms Assembled on Different Surfaces Using Phenotypic and Genotypic Approaches. Microorganisms 2017, 5, 16. [Google Scholar] [CrossRef]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S RRNA-Targeted Oligonucleotide Probes with Flow Cytometry for Analyzing Mixed Microbial Populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [CrossRef]
- Kempf, V.A.J.; Trebesius, K.; Autenrieth, I.B. Fluorescent In Situ Hybridization Allows Rapid Identification of Microorganisms in Blood Cultures. J. Clin. Microbiol. 2000, 38, 830. [Google Scholar] [CrossRef]
- Llarena, A.-K.; Ribeiro-Gonçalves, B.F.; Nuno Silva, D.; Halkilahti, J.; Machado, M.P.; Santos, M.; Silva, D.; Jaakkonen, A.; Isidro, J.; Hämäläinen, C.; et al. INNUENDO: A Cross-Sectoral Platform for the Integration of Genomics in the Surveillance of Food-Borne Pathogens. EFSA Supporting Publ. 2018, 15, 1498E. [Google Scholar] [CrossRef]
- Alcoceba, E.; Gómez, A.; Lara-Esbrí, P.; Oliver, A.; Beltrán, A.F.; Ayestarán, I.; Muñoz, P.; Escribano, P.; Guinea, J. Fluconazole-Resistant Candida parapsilosis Clonally Related Genotypes: First Report Proving the Presence of Endemic Isolates Harbouring the Y132F ERG11 Gene Substitution in Spain. Clin. Microbiol. Infect. 2022, 28, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Branco, J.; Fonseca, E.; Gomes, N.C.; Martins-Cruz, C.; Silva, A.P.; Silva-Dias, A.; Pina-Vaz, C.; Rodrigues, A.G.; Miranda, I.M.; Erraught, C.; et al. Impact of ERG3 Mutations and Expression of Ergosterol Genes Controlled by UPC2 and NDT80 in Candida parapsilosis Azole Resistance. Clin. Microbiol. Infect. 2017, 23, 575.e1–575.e8. [Google Scholar] [CrossRef] [PubMed]
- Martí-Carrizosa, M.; Sánchez-Reus, F.; March, F.; Cantón, E.; Coll, P. Implication of Candida parapsilosis FKS1 and FKS2 Mutations in Reduced Echinocandin Susceptibility. Antimicrob. Agents Chemother. 2015, 59, 3570–3573. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A Reference Database for Bacterial Virulence Factors. Nucleic Acids Res. 2005, 33 (Suppl. S1), D325–D328. [Google Scholar] [CrossRef]
- Neji, S.; Hadrich, I.; Trabelsi, H.; Abbes, S.; Cheikhrouhou, F.; Sellami, H.; Makni, F.; Ayadi, A. Virulence Factors, Antifungal Susceptibility and Molecular Mechanisms of Azole Resistance among Candida parapsilosis Complex Isolates Recovered from Clinical Specimens. J. Biomed. Sci. 2017, 24, 1–16. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Central Venous Catheter | Hemoculture | ||
|---|---|---|---|---|
| Antibiotic | MIC (mg/L) | Phenotype 1 | MIC (mg/L) | Phenotype |
| Ticarcillin | ≥128 | R | ≥128 | R |
| Piperacillin/Tazobactam | ≥128 | R | ≥128 | R |
| Ceftazidime | ≥64 | R | ≥64 | R |
| Ceftolozane/Tazobactam | 2 | R | 1 | S |
| Cefepime | 0.25 | S | ≤0.12 | S |
| Aztreonam | 16 | R | 16 | R |
| Imipenem | ≤0.25 | S | ≤0.25 | S |
| Meropenem | ≤0.25 | S | ≤0.25 | S |
| Amikacin | 2 | S | ≤1 | S |
| Gentamicin | ≤1 | S | ≤1 | S |
| Tobramycin | ≤1 | S | ≤1 | S |
| Ciprofloxacin | ≤0.06 | S | ≤0.06 | S |
| Levofloxacin | ≤0.12 | S | ≤0.12 | S |
| Tigecyclin | ≤0.5 | S | ≤0.5 | S |
| Trimethoprim/Sulfamethoxazole | ≤20 | S | ≤20 | S |
| Colistin | >8 | R | >8 | R |
| Source | Central Venous Catheter | Hemoculture | ||
|---|---|---|---|---|
| Antimicrobial Agents | MIC (mg/L) | Phenotype 1 | MIC (mg/L) | Phenotype |
| Fluconazole | >256 | R | >256 | R |
| Amphotericin B | 0.032 | S | 0.125 | S |
| Anidulafungin | 0.50 | S | 0.50 | S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štefánek, M.; Wenner, S.; Borges, V.; Pinto, M.; Gomes, J.P.; Rodrigues, J.; Faria, I.; Pessanha, M.A.; Martins, F.; Sabino, R.; et al. Antimicrobial Resistance and Biofilms Underlying Catheter-Related Bloodstream Coinfection by Enterobacter cloacae Complex and Candida parapsilosis. Antibiotics 2022, 11, 1245. https://doi.org/10.3390/antibiotics11091245
Štefánek M, Wenner S, Borges V, Pinto M, Gomes JP, Rodrigues J, Faria I, Pessanha MA, Martins F, Sabino R, et al. Antimicrobial Resistance and Biofilms Underlying Catheter-Related Bloodstream Coinfection by Enterobacter cloacae Complex and Candida parapsilosis. Antibiotics. 2022; 11(9):1245. https://doi.org/10.3390/antibiotics11091245
Chicago/Turabian StyleŠtefánek, Matúš, Sigurd Wenner, Vítor Borges, Miguel Pinto, João Paulo Gomes, João Rodrigues, Isabel Faria, Maria Ana Pessanha, Filomena Martins, Raquel Sabino, and et al. 2022. "Antimicrobial Resistance and Biofilms Underlying Catheter-Related Bloodstream Coinfection by Enterobacter cloacae Complex and Candida parapsilosis" Antibiotics 11, no. 9: 1245. https://doi.org/10.3390/antibiotics11091245
APA StyleŠtefánek, M., Wenner, S., Borges, V., Pinto, M., Gomes, J. P., Rodrigues, J., Faria, I., Pessanha, M. A., Martins, F., Sabino, R., Veríssimo, C., Nogueira, I. D., Carvalho, P. A., Bujdáková, H., & Jordao, L. (2022). Antimicrobial Resistance and Biofilms Underlying Catheter-Related Bloodstream Coinfection by Enterobacter cloacae Complex and Candida parapsilosis. Antibiotics, 11(9), 1245. https://doi.org/10.3390/antibiotics11091245