
New Weapons to Fight against Staphylococcus aureus Skin Infections




Abstract
:1. Introduction
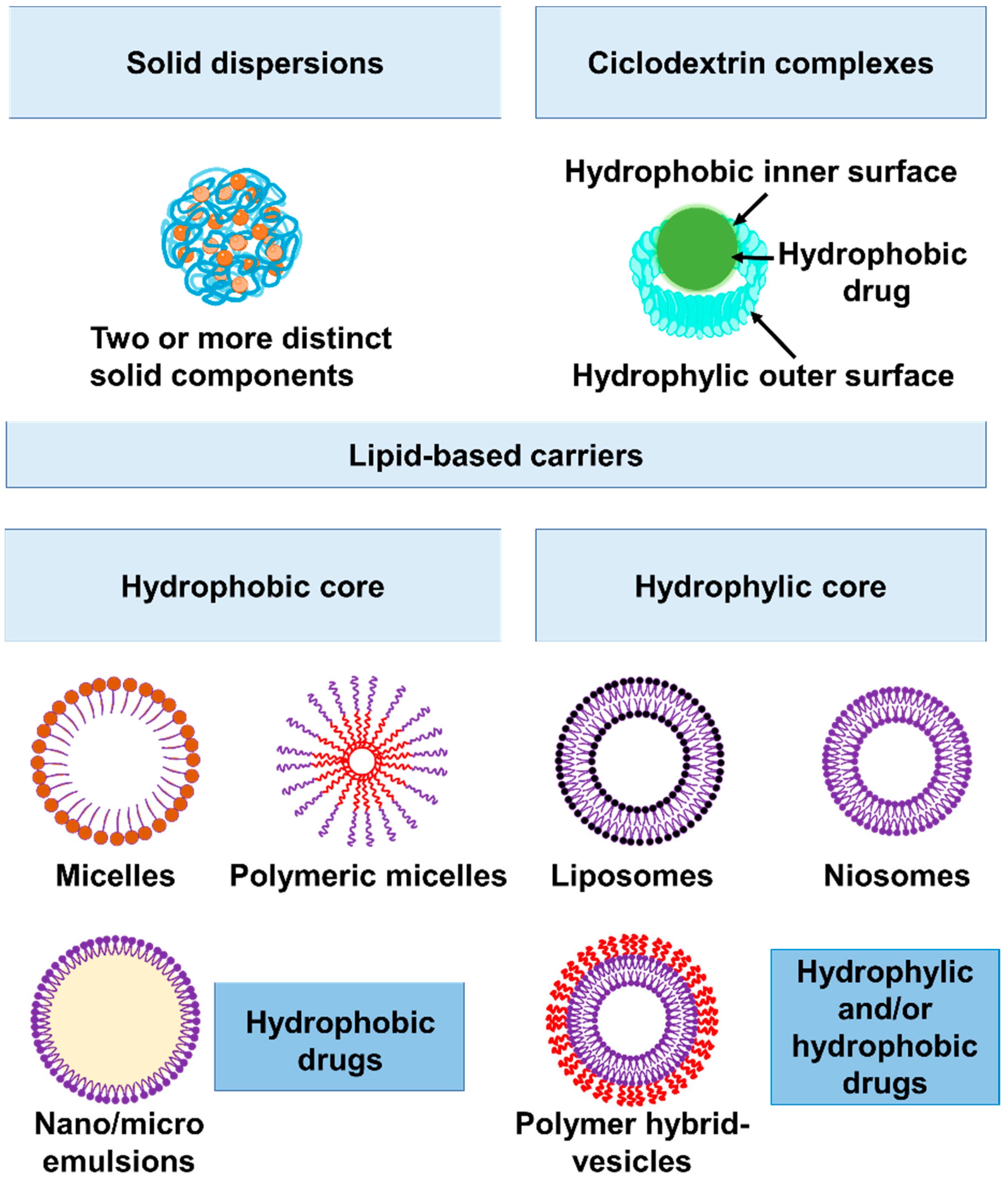
2. Novel Strategies to Increase Sensitivity to Currently Available Antibiotics as well as Their Bioavailability, Stability, and Tissue Penetration
2.1. Restoring Sensitivity to β-Lactam Antibiotics
2.2. Increasing Bioavailability of Hydrophobic Antibiotics: Macrolide and Fluoroquinolones
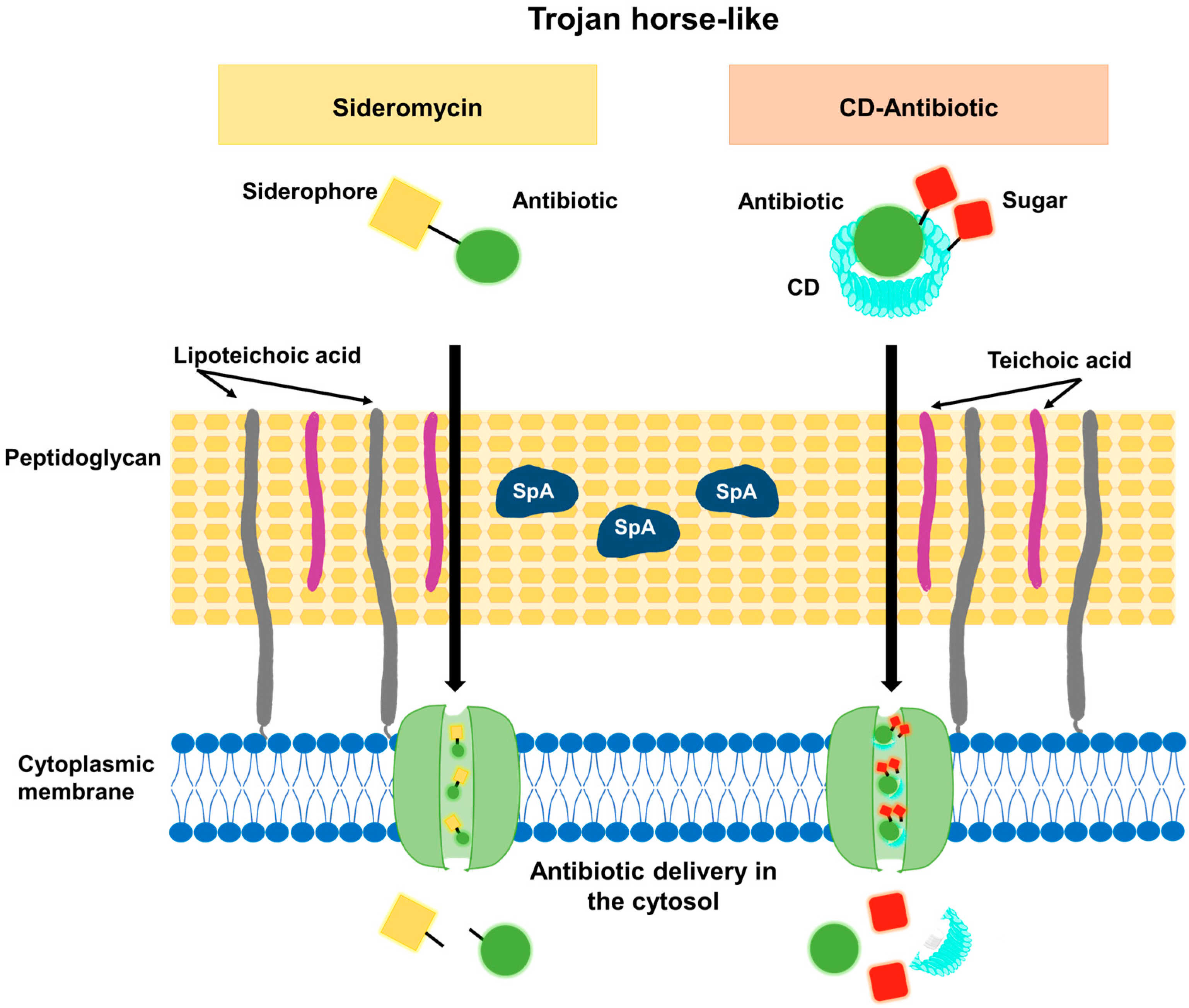
2.3. Increasing Bacterial Uptake of Antimicrobials: The Trojan-Horse-like Strategy
2.4. Improving Local Delivery of Antibiotics during Skin Infections
3. Novel Non-Antibiotic-Based Strategies for the Local Treatment of SSTIs
3.1. Local Delivery of Antimicrobial Peptides
3.2. Local Administration or Production of Nitric Oxide
3.3. Phototherapy
3.3.1. Photodynamic Therapy
3.3.2. UV Phototherapy
3.3.3. Photothermal Therapy
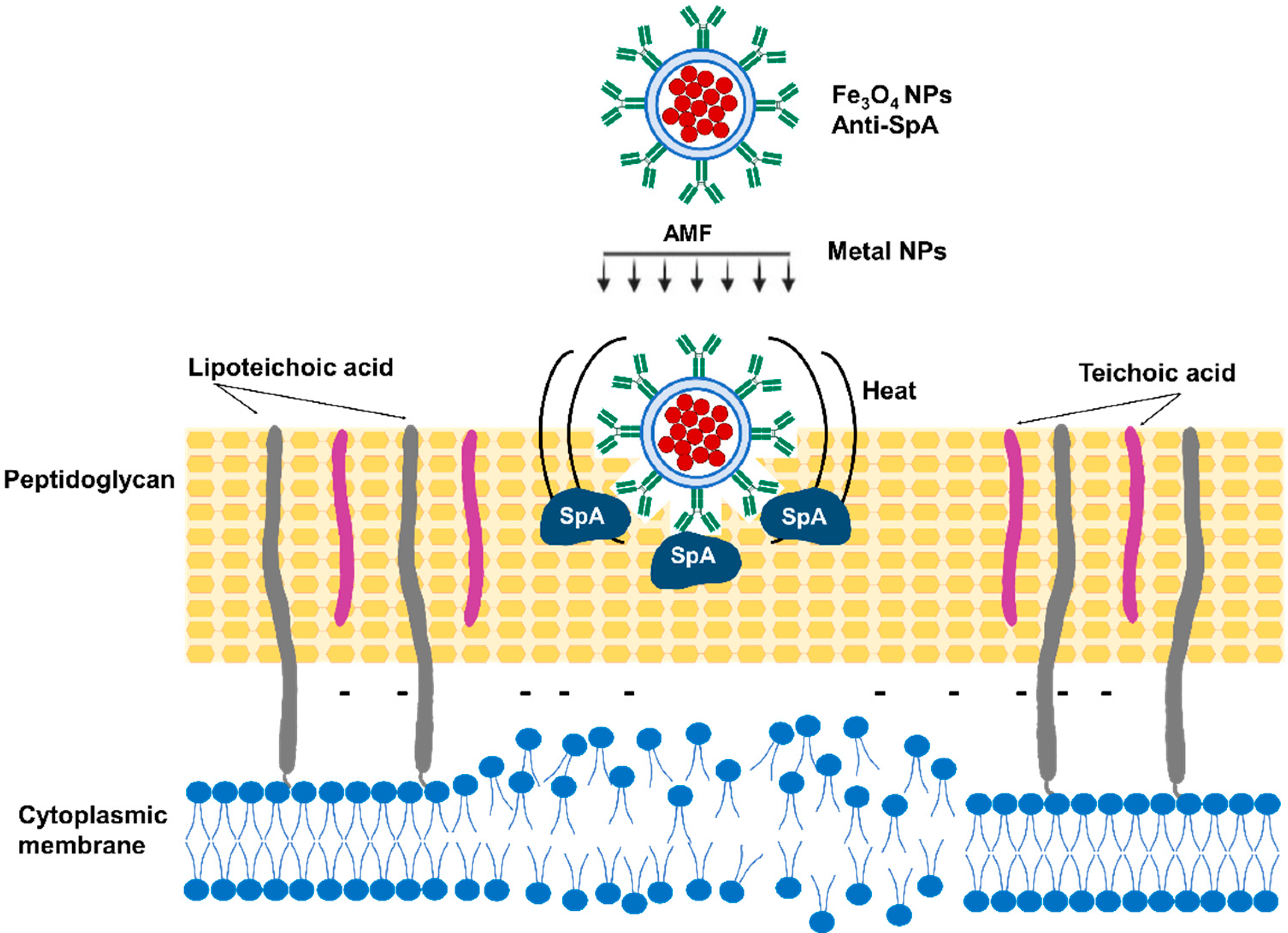
3.4. Magnetic Nanoparticle Hyperthermia
3.5. Mercaptophenylboronic Acid-Activated Gold Nanoparticles
3.6. Dialdehyde Nanocrystalline Cellulose
3.7. Bioactive Glass Nanoparticles
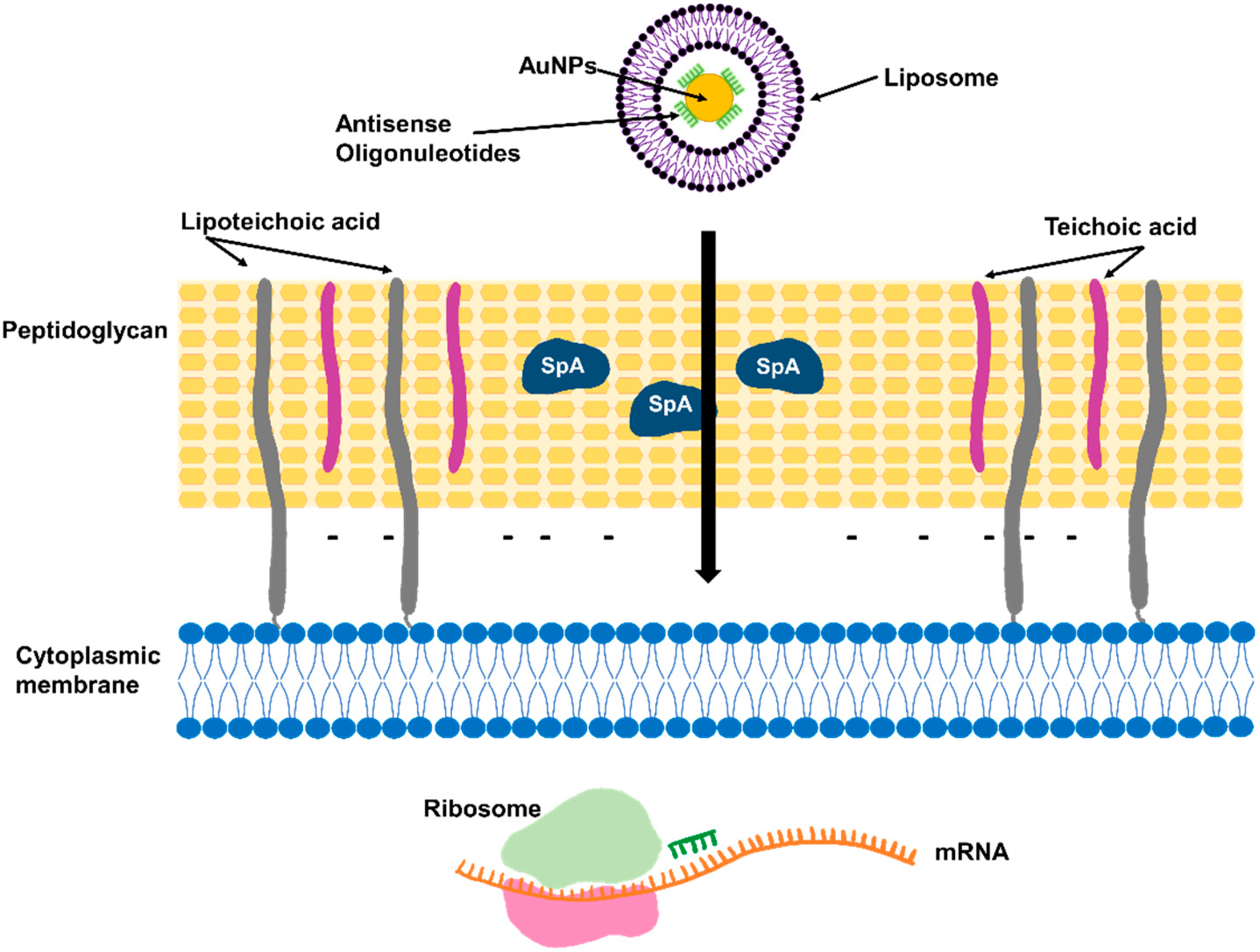
3.8. Antisense Oligonucleotides Targeting Essential Genes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hatlen, T.J.; Miller, L.G. Staphylococcal Skin and Soft Tissue Infections. Infect. Dis. Clin. N. Am. 2021, 35, 81–105. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.S.; Cho, J.S. Immunity against Staphylococcus aureus Cutaneous Infections. Nat. Rev. Immunol. 2011, 11, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Klevens, R.M.; Morrison, M.A.; Nadle, J.; Petit, S.; Gershman, K.; Ray, S.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; Townes, J.M.; et al. Invasive Methicillin-Resistant Staphylococcus aureus Infections in the United States. JAMA 2007, 298, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Dryden, M.S. Complicated Skin and Soft Tissue Infection. J. Antimicrob. Chemother. 2010, 65, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, J.D. Bioavailability of Antibiotics. J. Fr. Ophtalmol. 2000, 23, 510–513. [Google Scholar]
- Lanzerstorfer, P.; Sandner, G.; Pitsch, J.; Mascher, B.; Aumiller, T.; Weghuber, J. Acute, Reproductive, and Developmental Toxicity of Essential Oils Assessed with Alternative in vitro and in vivo Systems. Arch. Toxicol. 2021, 95, 673–691. [Google Scholar] [CrossRef]
- Moola, S.; Orchard, A.; van Vuuren, S. The Antimicrobial and Toxicity Influence of Six Carrier Oils on Essential Oil Compounds. Molecules 2023, 28, 30. [Google Scholar] [CrossRef]
- Sinha, S.; Jothiramajayam, M.; Ghosh, M.; Mukherjee, A. Evaluation of Toxicity of Essential Oils Palmarosa, Citronella, Lemongrass and Vetiver in Human Lymphocytes. Food Chem. Toxicol. 2014, 68, 71–77. [Google Scholar] [CrossRef]
- Turchi, B.; Mancini, S.; Pistelli, L.; Najar, B.; Fratini, F. Sub-Inhibitory Concentration of Essential Oils Induces Antibiotic Resistance in Staphylococcus aureus. Nat. Prod. Res. 2019, 33, 1509–1513. [Google Scholar] [CrossRef]
- Plumet, L.; Ahmad-Mansour, N.; Dunyach-Remy, C.; Kissa, K.; Sotto, A.; Lavigne, J.P.; Costechareyre, D.; Molle, V. Bacteriophage Therapy for Staphylococcus aureus Infections: A Review of Animal Models, Treatments, and Clinical Trials. Front. Cell. Infect. Microbiol. 2022, 12, 1–16. [Google Scholar] [CrossRef]
- Liu, X.; Ma, L.; Chen, F.; Liu, J.; Yang, H.; Lu, Z. Synergistic Antibacterial Mechanism of Bi2Te3 Nanoparticles Combined with the Ineffective β-Lactam Antibiotic Cefotaxime against Methicillin-Resistant Staphylococcus aureus. J. Inorg. Biochem. 2019, 196, 110687. [Google Scholar] [CrossRef] [PubMed]
- Chavan, C.; Kamble, S.; Murthy, A.; Kale, S.N. Ampicillin-Mediated Functionalized Gold Nanoparticles against Ampicillin-Resistant Bacteria: Strategy, Preparation and Interaction Studies To. Nanotechnology 2020, 31, 215604. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.M.; Deming, M.A.; Sarkar, A. β-Lactamase Suppression as a Strategy to Target Methicillin-Resistant Staphylococcus aureus: Proof of Concept. ACS Omega 2022, 7, 46213–46221. [Google Scholar] [CrossRef] [PubMed]
- Moreillon, P. Amoxycillin-Clavulanate versus Methicillin or Isoxazolyl Penicillins for Treatment of Staphylococcus aureus Infections. J. Antimicrob. Chemother. 1995, 35, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, D.; Peters, B.M.; Li, L.; Li, B.; Xu, Z.; Shirliff, M.E. Staphylococcal Chromosomal Cassettes Mec (SCCmec): A Mobile Genetic Element in Methicillin-Resistant Staphylococcus aureus. Microb. Pathog. 2016, 101, 56–67. [Google Scholar] [CrossRef]
- Meng, J.; Wang, H.; Hou, Z.; Chen, T.; Fu, J.; Ma, X.; He, G.; Xue, X.; Jia, M.; Luo, X. Novel Anion Liposome-Encapsulated Antisense Oligonucleotide Restores Susceptibility of Methicillin-Resistant Staphylococcus aureus and Rescues Mice from Lethal Sepsis by Targeting MecA. Antimicrob. Agents Chemother. 2009, 53, 2871–2878. [Google Scholar] [CrossRef]
- Beha, M.J.; Ryu, J.S.; Kim, Y.S.; Chung, H.J. Delivery of Antisense Oligonucleotides Using Multi-Layer Coated Gold Nanoparticles to Methicillin-Resistant S. aureus for Combinatorial Treatment. Mater. Sci. Eng. C 2021, 126, 112167. [Google Scholar] [CrossRef]
- Sharaf, M.H.; El-Sherbiny, G.M.; Moghannem, S.A.; Abdelmonem, M.; Elsehemy, I.A.; Metwaly, A.M.; Kalaba, M.H. New Combination Approaches to Combat Methicillin-Resistant Staphylococcus aureus (MRSA). Sci. Rep. 2021, 11, 4240. [Google Scholar] [CrossRef]
- Vemula, V.R.; Lagishetty, V.; Lingala, S. Solubility Enhancement Techniques. Int. J. Pharm. Sci. Rev. Res. 2010, 5, 41–51. [Google Scholar]
- Edward, K.H.; Li, D. Drug-Like Properties: Concept, Structure Design and Methods, From ADME to Toxicity Optimization. In Drug-like Properties: Concepts, Structure Design and Methods; Academic Press: Cambridge, MA, USA, 2008; pp. 6–16. [Google Scholar] [CrossRef]
- Khalikov, S.S.; Dushkin, A.V. Strategies for Solubility Enhancement of Anthelmintics (Review). Pharm. Chem. J. 2020, 54, 504–508. [Google Scholar] [CrossRef]
- Williams, H.D.; Trevaskis, N.L.; Charman, S.A.; Shanker, R.M.; Charman, W.N.; Pouton, C.W.; Porter, C.J.H. Strategies to Address Low Drug Solubility in Discovery and Development. Pharmacol. Rev. 2013, 65, 315–499. [Google Scholar] [CrossRef] [PubMed]
- Naskar, A.; Lee, S.; Lee, Y.; Kim, S.; Kim, K.S. A New Nano-Platform of Erythromycin Combined with Ag Nano-Particle ZnO Nano-Structure against Methicillin-Resistant Staphylococcus aureus. Pharmaceutics 2020, 12, 841. [Google Scholar] [CrossRef] [PubMed]
- Slavin, Y.N.; Asnis, J.; Häfeli, U.O.; Bach, H. Metal Nanoparticles: Understanding the Mechanisms behind Antibacterial Activity. J. Nanobiotechnol. 2017, 15, 65. [Google Scholar] [CrossRef] [PubMed]
- Ibraheem, D.R.; Hussein, N.N.; Sulaiman, G.M.; Mohammed, H.A.; Khan, R.A.; Al Rugaie, O. Ciprofloxacin-Loaded Silver Nanoparticles as Potent Nano-Antibiotics against Resistant Pathogenic Bacteria. Nanomaterials 2022, 12, 2808. [Google Scholar] [CrossRef] [PubMed]
- Steckiewicz, K.P.; Cieciórski, P.; Barcińska, E.; Jaśkiewicz, M.; Narajczyk, M.; Bauer, M.; Kamysz, W.; Megiel, E.; Inkielewicz-Stepniak, I. Silver Nanoparticles as Chlorhexidine and Metronidazole Drug Delivery Platforms: Their Potential Use in Treating Periodontitis. Int. J. Nanomed. 2022, 17, 495–517. [Google Scholar] [CrossRef] [PubMed]
- Mondal Roy, S.; Sharma, B.K.; Roy, D.R. Synthesis of Ciprofloxacin Drug Capped Silver Nanoparticles and Their Antimicrobial Activity: A Joint Spectrophotometric and Density Functional Investigation. J. Clust. Sci. 2021, 32, 1575–1584. [Google Scholar] [CrossRef]
- Mohsen, E.; El-Borady, O.M.; Mohamed, M.B.; Fahim, I.S. Synthesis and Characterization of Ciprofloxacin Loaded Silver Nanoparticles and Investigation of Their Antibacterial Effect. J. Radiat. Res. Appl. Sci. 2020, 13, 416–425. [Google Scholar] [CrossRef]
- Mendes, C.; Meirelles, G.C.; Barp, C.G.; Assreuy, J.; Silva, M.A.S.; Ponchel, G. Cyclodextrin Based Nanosponge of Norfloxacin: Intestinal Permeation Enhancement and Improved Antibacterial Activity. Carbohydr. Polym. 2018, 195, 586–592. [Google Scholar] [CrossRef]
- Jansook, P.; Ogawa, N.; Loftsson, T. Cyclodextrins: Structure, Physicochemical Properties and Pharmaceutical Applications. Int. J. Pharm. 2018, 535, 272–284. [Google Scholar] [CrossRef]
- Szabó, Z.I.; Deme, R.; Mucsi, Z.; Rusu, A.; Mare, A.D.; Fiser, B.; Toma, F.; Sipos, E.; Tóth, G. Equilibrium, Structural and Antibacterial Characterization of Moxifloxacin-β-Cyclodextrin Complex. J. Mol. Struct. 2018, 1166, 228–236. [Google Scholar] [CrossRef]
- Tănase, M.A.; Raducan, A.; Oancea, P.; Diţu, L.M.; Stan, M.; Petcu, C.; Scomoroşcenco, C.; Ninciuleanu, C.M.; Nistor, C.L.; Cinteza, L.O. Mixed Pluronic—Cremophor Polymeric Micelles as Nanocarriers for Poorly Soluble Antibiotics—The Influence on the Antibacterial Activity. Pharmaceutics 2021, 13, 435. [Google Scholar] [CrossRef] [PubMed]
- Akbar, N.; Gul, J.; Siddiqui, R.; Shah, M.R.; Khan, N.A. Moxifloxacin and Sulfamethoxazole-Based Nanocarriers Exhibit Potent Antibacterial Activities. Antibiotics 2021, 10, 964. [Google Scholar] [CrossRef] [PubMed]
- Albelda-Berenguer, M.; Monachon, M.; Joseph, E. Siderophores: From Natural Roles to Potential Applications. Adv. Appl. Microbiol. 2019, 106, 193–225. [Google Scholar] [CrossRef] [PubMed]
- Braun, V.; Pramanik, A.; Gwinner, T.; Köberle, M.; Bohn, E. Sideromycins: Tools and Antibiotics. BioMetals 2009, 22, 3–13. [Google Scholar] [CrossRef]
- Wencewicz, T.A.; Long, T.E.; Möllmann, U.; Miller, M.J. Trihydroxamate Siderophore-Fluoroquinolone Conjugates Are Selective Sideromycin Antibiotics That Target Staphylococcus aureus. Bioconjug. Chem. 2013, 24, 473–486. [Google Scholar] [CrossRef]
- Pramanik, A.; Stroeher, U.H.; Krejci, J.; Standish, A.J.; Bohn, E.; Paton, J.C.; Autenrieth, I.B.; Braun, V. Albomycin Is an Effective Antibiotic, as Exemplified with Yersinia Enterocolitica and Streptococcus Pneumoniae. Int. J. Med. Microbiol. 2007, 297, 459–469. [Google Scholar] [CrossRef]
- Pramanik, A.; Braun, V. Albomycin Uptake via a Ferric Hydroxamate Transport System of Streptococcus Pneumoniae R6. J. Bacteriol. 2006, 188, 3878–3886. [Google Scholar] [CrossRef]
- Stefanska, A.L.; Fulston, M.; Houge-Frydrych, C.S.V.; Jones, J.J.; Warr, S.R. A Potent Seryl TRNA Synthetase Inhibitor SB-217452 Isolated from a Streptomyces Species. J. Antibiot. 2000, 53, 1346–1353. [Google Scholar] [CrossRef]
- Lin, Z.; Xu, X.; Zhao, S.; Yang, X.; Guo, J.; Zhang, Q.; Jing, C.; Chen, S.; He, Y. Total Synthesis and Antimicrobial Evaluation of Natural Albomycins against Clinical Pathogens. Nat. Commun. 2018, 9, 3445. [Google Scholar] [CrossRef]
- Gause, G.F.; Biol, D. Recent Studies on Albomycin, a New Antiriotic. Br. Med. J. 1955, 2, 1177–1179. [Google Scholar] [CrossRef]
- Ushimaru, R.; Chen, Z.; Zhao, H.; Fan, P.; Liu, H. The Enzymes Mediating Maturation of the Seryl-TRNA Synthetase Inhibitor SB-217452 during Biosynthesis of Albomycins. Angew. Chem. Int. Ed. Engl. 2020, 59, 3558–3562. [Google Scholar] [CrossRef]
- Li, M.; Neoh, K.G.; Xu, L.; Yuan, L.; Leong, D.T.; Kang, E.T.; Chua, K.L.; Hsu, L.Y. Sugar-Grafted Cyclodextrin Nanocarrier as a “Trojan Horse” for Potentiating Antibiotic Activity. Pharm. Res. 2016, 33, 1161–1174. [Google Scholar] [CrossRef] [PubMed]
- Jeckelmann, J.M.; Erni, B. Transporters of Glucose and Other Carbohydrates in Bacteria. Pflug. Arch. Eur. J. Physiol. 2020, 472, 1129–1153. [Google Scholar] [CrossRef] [PubMed]
- Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 5.0; The European Committee on Antimicrobial Susceptibility Testing; European Committee: Luxembourg, 2015; pp. 1–77.
- Giandalia, G.; De Caro, V.; Cordone, L.; Giannola, L.I. Trehalose-Hydroxyethylcellulose Microspheres Containing Vancomycin for Topical Drug Delivery. Eur. J. Pharm. Biopharm. 2001, 52, 83–89. [Google Scholar] [CrossRef]
- Mawhinney, W.M.; Adair, C.G.; Gorman, S.P.; McClurg, B. Stability of Vancomycin Hydrochloride in Peritoneal Dialysis Solution. Am. J. Hosp. Pharm. 1992, 49, 137–139. [Google Scholar] [CrossRef]
- Raverdy, V.; Ampe, E.; Hecq, J.D.; Tulkens, P.M. Stability and Compatibility of Vancomycin for Administration by Continuous Infusion. J. Antimicrob. Chemother. 2013, 68, 1179–1182. [Google Scholar] [CrossRef]
- Vandecasteele, S.J.; De Vriese, A.S.; Tacconelli, E. The Pharmacokinetics and Pharmacodynamics of Vancomycin in Clinical Practice: Evidence and Uncertainties. J. Antimicrob. Chemother. 2013, 68, 743–748. [Google Scholar] [CrossRef]
- Walvekar, P.; Gannimani, R.; Salih, M.; Makhathini, S.; Mocktar, C.; Govender, T. Self-Assembled Oleylamine Grafted Hyaluronic Acid Polymersomes for Delivery of Vancomycin against Methicillin Resistant Staphylococcus aureus (MRSA). Colloids Surf. B Biointerfaces 2019, 182, 110388. [Google Scholar] [CrossRef]
- Hassan, D.; Omolo, C.A.; Fasiku, V.O.; Mocktar, C.; Govender, T. Novel Chitosan-Based PH-Responsive Lipid-Polymer Hybrid Nanovesicles (OLA-LPHVs) for Delivery of Vancomycin against Methicillin-Resistant Staphylococcus aureus Infections. Int. J. Biol. Macromol. 2020, 147, 385–398. [Google Scholar] [CrossRef]
- López-León, T.; Carvalho, E.L.S.; Seijo, B.; Ortega-Vinuesa, J.L.; Bastos-González, D. Physicochemical Characterization of Chitosan Nanoparticles: Electrokinetic and Stability Behavior. J. Colloid Interface Sci. 2005, 283, 344–351. [Google Scholar] [CrossRef]
- Park, D.; Ryu, H.; Kim, H.S.; Kim, Y.S.; Choi, K.S.; Park, H.; Seo, J. Sonophoresis Using Ultrasound Contrast Agents for Transdermal Drug Delivery: An in vivo Experimental Study. Ultrasound Med. Biol. 2012, 38, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Argenziano, M.; Banche, G.; Luganini, A.; Finesso, N.; Allizond, V.; Gulino, G.R.; Khadjavi, A.; Spagnolo, R.; Tullio, V.; Giribaldi, G.; et al. Vancomycin-Loaded Nanobubbles: A New Platform for Controlled Antibiotic Delivery against Methicillin-Resistant Staphylococcus Aureus Infections; Elsevier B.V.: Amsterdam, The Netherlands, 2017; Volume 523, ISBN 3901167056. [Google Scholar]
- Dhanalakshmi, V.; Nimal, T.R.; Sabitha, M.; Biswas, R.; Jayakumar, R. Skin and Muscle Permeating Antibacterial Nanoparticles for Treating Staphylococcus aureus Infected Wounds. J. Biomed. Mater. Res.—Part B Appl. Biomater. 2016, 104, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Sonvico, F.; Cagnani, A.; Rossi, A.; Motta, S.; Di Bari, M.T.; Cavatorta, F.; Alonso, M.J.; Deriu, A.; Colombo, P. Formation of Self-Organized Nanoparticles by Lecithin/Chitosan Ionic Interaction. Int. J. Pharm. 2006, 324, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Schäfer-Korting, M.; Mehnert, W.; Korting, H.C. Lipid Nanoparticles for Improved Topical Application of Drugs for Skin Diseases. Adv. Drug Deliv. Rev. 2007, 59, 427–443. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Lin, Y.; Lu, H.; Li, Q.; He, J.; Chen, B.; Wu, C.; Xu, Y. Percutaneous Delivery of Econazole Using Microemulsion as Vehicle: Formulation, Evaluation and Vesicle-Skin Interaction. Int. J. Pharm. 2014, 465, 120–131. [Google Scholar] [CrossRef]
- Abruzzo, A.; Parolin, C.; Bigucci, F.; Rossi, M.; Vitali, B.; Cappadone, C.; Bigucci, F. Development and Characterization of Azithromycin-Loaded Microemulsions: A Promising Tool for the Treatment of Bacterial Skin Infections. Antibiotics 2022, 11, 1040. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Sahl, H.G. Antimicrobial and Host-Defense Peptides as New Anti-Infective Therapeutic Strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 1–21. [Google Scholar] [CrossRef]
- Wei, J.; Cao, X.; Qian, J.; Liu, Z.; Wang, X.; Su, Q.; Wang, Y.; Xie, R.; Li, X. Evaluation of Antimicrobial Peptide LL-37 for Treatment of Staphylococcus Aureus Biofilm on Titanium Plate. Medicine 2021, 100, e27426. [Google Scholar] [CrossRef]
- Sadeghi, S.; Bakhshandeh, H.; Cohan, R.A.; Peirovi, A.; Ehsani, P.; Norouzian, D. Synergistic Anti-Staphylococcal Activity of Niosomal Recombinant Lysostaphin-LL-37. Int. J. Nanomed. 2019, 14, 9777–9792. [Google Scholar] [CrossRef]
- Desbois, A.P.; Gemmell, C.G.; Coote, P.J. In Vivo Efficacy of the Antimicrobial Peptide Ranalexin in Combination with the Endopeptidase Lysostaphin against Wound and Systemic Meticillin-Resistant Staphylococcus aureus (MRSA) Infections. Int. J. Antimicrob. Agents 2010, 35, 559–565. [Google Scholar] [CrossRef]
- Polak, J.; Latta, P.D.; Blackburn, P. In Vitro Activity of Recombinant Lysostaphin-Antibiotic Combinations toward Methicillin-Resistant Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 1993, 17, 265–270. [Google Scholar] [CrossRef]
- Were, L.M.; Bruce, B.; Davidson, P.M.; Weiss, J. Encapsulation of Nisin and Lysozyme in Liposomes Enhances Efficacy against Listeria Monocytogenes. J. Food Prot. 2004, 67, 922–927. [Google Scholar] [CrossRef]
- Somwongin, S.; Chantawannakul, P.; Chaiyana, W. Antioxidant Activity and Irritation Property of Venoms from Apis Species. Toxicon 2018, 145, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Aufschnaiter, A.; Kohler, V.; Khalifa, S.; El-Wahed, A.A.; Du, M.; El-Seedi, H.; Büttner, S. Apitoxin and Its Components against Cancer, Neurodegeneration and Rheumatoid Arthritis: Limitations and Possibilities. Toxins 2020, 12, 66. [Google Scholar] [CrossRef] [PubMed]
- Sangboonruang, S.; Semakul, N.; Obeid, M.A.; Ruano, M.; Kitidee, K.; Anukool, U.; Pringproa, K.; Chantawannakul, P.; Ferro, V.A.; Tragoolpua, Y.; et al. Potentiality of Melittin-Loaded Niosomal Vesicles against Vancomycin-Intermediate Staphylococcus aureus and Staphylococcal Skin Infection. Int. J. Nanomed. 2021, 16, 7639–7661. [Google Scholar] [CrossRef]
- Choi, J.H.; Jang, A.Y.; Lin, S.; Lim, S.; Kim, D.; Park, K.; Han, S.M.; Yeo, J.H.; Seo, H.S. Melittin, a Honeybee Venom-Derived Antimicrobial Peptide, May Target Methicillin-Resistant Staphylococcus aureus. Mol. Med. Rep. 2015, 12, 6483–6490. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Bae, H. Anti-Inflammatory Applications of Melittin, a Major Component of Bee Venom: Detailed Mechanism of Action and Adverse Effects. Molecules 2016, 21, 616. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, A.W.; Schoenfisch, M.H. Nitric Oxide Release: Part II. Therapeutic Applications. Chem. Soc. Rev. 2012, 41, 3742–3752. [Google Scholar] [CrossRef]
- Jones, M.L.; Ganopolsky, J.G.; Labbé, A.; Wahl, C.; Prakash, S. Antimicrobial Properties of Nitric Oxide and Its Application in Antimicrobial Formulations and Medical Devices. Appl. Microbiol. Biotechnol. 2010, 88, 401–407. [Google Scholar] [CrossRef]
- Burgner, D.; Rockett, K.; Kwiatkowski, D. Nitric Oxide and Infectious Diseases. Arch. Dis. Child. 1999, 81, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Schairer, D.O.; Chouake, J.S.; Nosanchuk, J.D.; Friedman, A.J. The Potential of Nitric Oxide Releasing Therapies as Antimicrobial Agents. Virulence 2012, 3, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Privett, B.J.; Broadnax, A.D.; Bauman, S.J.; Riccio, D.A.; Schoenfisch, M.H. Examination of Bacterial Resistance to Exogenous Nitric Oxide. Nitric Oxide—Biol. Chem. 2012, 26, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric Oxide and the Immune Response—Nature Immunology. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef]
- Schäffer, M.R.; Tantry, U.; Gross, S.S.; Wasserkrug, H.L.; Barbul, A. Nitric Oxide Regulates Wound Healing. J. Surg. Res. 1996, 63, 237–240. [Google Scholar] [CrossRef]
- Wallace, J.L. Nitric Oxide as a Regulator of Inflammatory Processes. Mem. Inst. Oswaldo Cruz 2005, 100, 5–9. [Google Scholar] [CrossRef]
- Smith, A.W. Biofilms and Antibiotic Therapy: Is There a Role for Combating Bacterial Resistance by the Use of Novel Drug Delivery Systems? Adv. Drug Deliv. Rev. 2005, 57, 1539–1550. [Google Scholar] [CrossRef]
- Malone-Povolny, M.J.; Maloney, S.E.; Schoenfisch, M.H. Nitric Oxide Therapy for Diabetic Wound Healing. Adv. Healthc. Mater. 2019, 8, 1801210. [Google Scholar] [CrossRef]
- Barone, M.; Sciortino, M.T.; Zaccaria, D.; Mazzaglia, A.; Sortino, S. Nitric Oxide Photocaging Platinum Nanoparticles with Anticancer Potential. J. Mater. Chem. 2008, 18, 5531–5536. [Google Scholar] [CrossRef]
- Lu, Y.; Slomberg, D.L.; Sun, B.; Schoenfisch, M.H. Shape- and Nitric Oxide Flux-Dependent Bactericidal Activity of Nitric Oxide-Releasing Silica Nanorods. Small 2013, 9, 2189–2198. [Google Scholar] [CrossRef]
- Slomberg, D.L.; Lu, Y.; Broadnax, A.D.; Hunter, R.A.; Carpenter, A.W.; Schoenfisch, M.H. Role of Size and Shape on Biofilm Eradication for Nitric Oxide-Releasing Silica Nanoparticles. ACS Appl. Mater. Interfaces 2013, 5, 9322–9329. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Slomberg, D.L.; Shah, A.; Schoenfisch, M.H. Nitric Oxide-Releasing Amphiphilic Poly(Amidoamine) (PAMAM) Dendrimers as Antibacterial Agents. Biomacromolecules 2013, 14, 3589–3598. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Slomberg, D.L.; Schoenfisch, M.H. Nitric Oxide-Releasing Chitosan Oligosaccharides as Antibacterial Agents. Biomaterials 2014, 35, 1716–1724. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.L.; Kee, P.H.; Kim, H.; Moody, M.R.; Chrzanowski, S.M.; MacDonald, R.C.; McPherson, D.D. Nitric Oxide-Loaded Echogenic Liposomes for Nitric Oxide Delivery and Inhibition of Intimal Hyperplasia. J. Am. Coll. Cardiol. 2009, 54, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Nablo, B.J.; Rothrock, A.R.; Schoenfisch, M.H. Nitric Oxide-Releasing Sol-Gels as Antibacterial Coatings for Orthopedic Implants. Biomaterials 2005, 26, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Nurhasni, H.; Cao, J.; Choi, M.; Kim, I.; Lee, B.L.; Jung, Y.; Yoo, J.W. Nitric Oxide-Releasing Poly(Lactic-Co-Glycolic Acid)-Polyethylenimine Nanoparticles for Prolonged Nitric Oxide Release, Antibacterial Efficacy, and in vivo Wound Healing Activity. Int. J. Nanomedicine 2015, 10, 3065–3080. [Google Scholar] [CrossRef]
- Hasan, N.; Cao, J.; Lee, J.; Naeem, M.; Hlaing, S.P.; Kim, J.; Jung, Y.; Lee, B.L.; Yoo, J.W. PEI/NONOates-Doped PLGA Nanoparticles for Eradicating Methicillin-Resistant Staphylococcus aureus Biofilm in Diabetic Wounds via Binding to the Biofilm Matrix. Mater. Sci. Eng. C 2019, 103, 109741. [Google Scholar] [CrossRef]
- Hetrick, E.M.; Shin, J.H.; Paul, H.S.; Schoenfisch, M.H. Anti-Biofilm Efficacy of Nitric Oxide-Releasing Silica Nanoparticles. Biomaterials 2009, 30, 2782–2789. [Google Scholar] [CrossRef]
- Kafshgari, M.H.; Cavallaro, A.; Delalat, B.; Harding, F.J.; McInnes, S.J.P.; Mäkilä, E.; Salonen, J.; Vasilev, K.; Voelcker, N.H. Nitric Oxide-Releasing Porous Silicon Nanoparticles. Nanoscale Res. Lett. 2014, 9, 333. [Google Scholar] [CrossRef]
- Ma, X.; Cheng, Y.; Jian, H.; Feng, Y.; Chang, Y.; Zheng, R.; Wu, X.; Wang, L.; Li, X.; Zhang, H. Hollow, Rough, and Nitric Oxide-Releasing Cerium Oxide Nanoparticles for Promoting Multiple Stages of Wound Healing. Adv. Healthc. Mater. 2019, 8, 1900256. [Google Scholar] [CrossRef]
- Pieretti, J.C.; Seabra, A.B. Nitric Oxide-Releasing Nanomaterials and Skin Infections. In Nanotechnology in Skin, Soft Tissue, and Bone Infections; Springer: Berlin/Heidelberg, Germany, 2020; pp. 3–23. [Google Scholar] [CrossRef]
- Niska, K.; Zielinska, E.; Radomski, M.W.; Inkielewicz-Stepniak, I. Metal Nanoparticles in Dermatology and Cosmetology: Interactions with Human Skin Cells. Chem. Biol. Interact. 2018, 295, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Jura, J.; Szmyd, R.; Goralczyk, A.G.; Skalniak, L.; Cierniak, A.; Lipert, B.; Filon, F.L.; Crosera, M.; Borowczyk, J.; Laczna, E.; et al. Effect of Silver Nanoparticles on Human Primary Keratinocytes. Biol. Chem. 2013, 394, 113–123. [Google Scholar] [CrossRef]
- Lewinski, N.; Colvin, V.; Drezek, R. Cytotoxicity of Nanopartides. Small 2008, 4, 26–49. [Google Scholar] [CrossRef]
- Champeau, M.; Póvoa, V.; Militão, L.; Cabrini, F.M.; Picheth, G.F.; Meneau, F.; Jara, C.P.; de Araujo, E.P.; de Oliveira, M.G. Supramolecular Poly(Acrylic Acid)/F127 Hydrogel with Hydration-Controlled Nitric Oxide Release for Enhancing Wound Healing. Acta Biomater. 2018, 74, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Amadeu, T.P.; Seabra, A.B.; de Oliveira, M.G.; Costa, A.M.A. S-Nitrosoglutathione-Containing Hydrogel Accelerates Rat Cutaneous Wound Repair. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 629–637. [Google Scholar] [CrossRef]
- Kim, J.O.; Noh, J.K.; Thapa, R.K.; Hasan, N.; Choi, M.; Kim, J.H.; Lee, J.H.; Ku, S.K.; Yoo, J.W. Nitric Oxide-Releasing Chitosan Film for Enhanced Antibacterial and in vivo Wound-Healing Efficacy. Int. J. Biol. Macromol. 2015, 79, 217–225. [Google Scholar] [CrossRef]
- Blevins, J.S.; Beenken, K.E.; Elasri, M.O.; Hurlburt, B.K.; Smeltzer, M.S. Strain-Dependent Differences in the Regulatory Roles of SarA and Agr in Staphylococcus aureus. Infect. Immun. 2002, 70, 470–480. [Google Scholar] [CrossRef]
- Lee, J.; Hlaing, S.P.; Cao, J.; Hasan, N.; Ahn, H.J.; Song, K.W.; Yoo, J.W. In Situ Hydrogel-Forming/Nitric Oxide-Releasing Wound Dressing for Enhanced Antibacterial Activity and Healing in Mice with Infected Wounds. Pharmaceutics 2019, 11, 496. [Google Scholar] [CrossRef]
- Broniowska, K.A.; Diers, A.R.; Hogg, N. S-Nitrosoglutathione. Biochim. Biophys. Acta—Gen. Subj. 2013, 1830, 3173–3181. [Google Scholar] [CrossRef]
- Forman, H.J. Glutathione—From Antioxidant to Post-Translational Modifier. Arch. Biochem. Biophys. 2016, 595, 64–67. [Google Scholar] [CrossRef]
- Duong, H.T.T.; Kamarudin, Z.M.; Erlich, R.B.; Li, Y.; Jones, M.W.; Kavallaris, M.; Boyer, C.; Davis, T.P. Intracellular Nitric Oxide Delivery from Stable No-Polymeric Nanoparticle Carriers. Chem. Commun. 2013, 49, 4190–4192. [Google Scholar] [CrossRef] [PubMed]
- Marcato, P.D.; Adami, L.F.; de Melo Barbosa, R.; Melo, P.S.; Ferreira, I.R.; de Paula, L.; Duran, N.; Seabra, A.B. Development of a Sustained-Release System for Nitric Oxide Delivery Using Alginate/Chitosan Nanoparticles. Curr. Nanosci. 2013, 9, 1–7. [Google Scholar] [CrossRef]
- Wu, W.; Gaucher, C.; Diab, R.; Fries, I.; Xiao, Y.L.; Hu, X.M.; Maincent, P.; Sapin-Minet, A. Time Lasting S-Nitrosoglutathione Polymeric Nanoparticles Delay Cellular Protein S-Nitrosation. Eur. J. Pharm. Biopharm. 2015, 89, 1–8. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, D.E.; Park, D.J.; Choi, G.H.; Yang, D.N.; Heo, J.S.; Lee, S.C. PH-Responsive Mineralized Nanoparticles as Stable Nanocarriers for Intracellular Nitric Oxide Delivery. Colloids Surf. B Biointerfaces 2016, 146, 1–8. [Google Scholar] [CrossRef]
- Lee, J.; Kwak, D.; Kim, H.; Kim, J.; Hlaing, S.P.; Hasan, N.; Cao, J.; Yoo, J.W. Nitric Oxide-Releasing s-Nitrosoglutathione-Conjugated Poly(Lactic-Co-Glycolic Acid) Nanoparticles for the Treatment of MRSA-Infected Cutaneous Wounds. Pharmaceutics 2020, 12, 618. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, X.; Lai, X.; Dzuvor, C.K.O.; Lyu, L.; Chow, S.H.; He, L.; Yu, L.; Wang, Y.; Song, J.; et al. Coassembled Nitric Oxide-Releasing Nanoparticles with Potent Antimicrobial Efficacy against Methicillin-Resistant Staphylococcus aureus (MRSA) Strains. ACS Appl. Mater. Interfaces 2022, 14, 37369–37379. [Google Scholar] [CrossRef] [PubMed]
- Hockberger, P.E. The Discovery of the Damaging Effect of Sunlight on Bacteria. J. Photochem. Photobiol. B Biol. 2000, 58, 185–191. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B.; Pathak, M.A. Historical Aspects of Methoxsalen and Other Furocoumarins. J. Investig. Dermatol. 1959, 32, 229–231. [Google Scholar] [CrossRef]
- Roelandts, R. The History of Phototherapy: Something New under the Sun? J. Am. Acad. Dermatol. 2002, 46, 926–930. [Google Scholar] [CrossRef]
- Gates, F.L. Study of the Bactericidal Action of Ultra Violet Light: I. The Reaction to Monochromatic Radiations. J. Gen. Physiol. 1929, 13, 231–248. [Google Scholar] [CrossRef]
- Gates, F.L. Study of the Bactericidal Action of Ultra Violet Light: II. The Effect of Various Environmental Factors and Conditions. J. Gen. Physiol. 1929, 13, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Gates, F.L. A Study of the Bactericidal Action of Ultra Violet Light: III. the Absorption of Ultra Violet Light by Bacteria. J. Gen. Physiol. 1930, 14, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Cabral, J.; Rodrigues, A. Blue Light Disinfection in Hospital Infection Control: Advantages, Drawbacks, and Pitfalls. Antibiotics 2019, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, J.; Rahban, D.; Aghamiri, S.; Teymouri, A.; Bahador, A. Photosensitizers in Antibacterial Photodynamic Therapy: An Overview. Laser Ther. 2018, 27, 293–302. [Google Scholar] [CrossRef]
- Wei, G.; Yang, G.; Wang, Y.; Jiang, H.; Fu, Y.; Yue, G.; Ju, R. Phototherapy-Based Combination Strategies for Bacterial Infection Treatment. Theranostics 2020, 10, 12241–12262. [Google Scholar] [CrossRef] [PubMed]
- Cieplik, F.; Deng, D.; Crielaard, W.; Buchalla, W.; Hellwig, E.; Al-Ahmad, A.; Maisch, T. Antimicrobial Photodynamic Therapy–What We Know and What We Don’t. Crit. Rev. Microbiol. 2018, 44, 571–589. [Google Scholar] [CrossRef]
- Pérez, C.; Zúñiga, T.; Palavecino, C.E. Photodynamic Therapy for Treatment of Staphylococcus aureus Infections. Photodiagnosis Photodyn. Ther. 2021, 34, 102285. [Google Scholar] [CrossRef]
- Grinholc, M.; Nakonieczna, J.; Fila, G.; Taraszkiewicz, A.; Kawiak, A.; Szewczyk, G.; Sarna, T.; Lilge, L.; Bielawski, K.P. Antimicrobial Photodynamic Therapy with Fulleropyrrolidine: Photoinactivation Mechanism of Staphylococcus aureus, in vitro and in vivo Studies. Appl. Microbiol. Biotechnol. 2015, 99, 4031–4043. [Google Scholar] [CrossRef]
- Torabi, S.; Joharchi, K.; Kalhori, K.A.M.; Sohrabi, M.; Fekrazad, R. Evaluation of Antimicrobial Photodynamic Therapy on Wounds Infected by Staphylococcus aureus in Animal Models. Photodiagnosis Photodyn. Ther. 2021, 33, 33212266. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Wang, Y.; Murray, C.K.; Hamblin, M.R.; Hooper, D.C.; Dai, T. Antimicrobial Blue Light Inactivation of Pathogenic Microbes: State of the Art. Drug Resist. Updat. 2017, 33–35, 1–22. [Google Scholar] [CrossRef]
- Pelgrift, R.Y.; Friedman, A.J. Nanotechnology as a Therapeutic Tool to Combat Microbial Resistance. Adv. Drug Deliv. Rev. 2013, 65, 1803–1815. [Google Scholar] [CrossRef] [PubMed]
- Halstead, F.D.; Thwaite, J.E.; Burt, R.; Laws, T.R.; Raguse, M.; Moeller, R.; Webber, M.A.; Oppenheim, B.A. Antibacterial Activity of Blue Light against Nosocomial Wound Pathogens Growing Planktonically and as Mature Biofilms. Appl. Environ. Microbiol. 2016, 82, 4006–4016. [Google Scholar] [CrossRef]
- Dai, T.; Gupta, A.; Huang, Y.Y.; Sherwood, M.E.; Murray, C.K.; Vrahas, M.S.; Kielian, T.; Hamblin, M.R. Blue Light Eliminates Community-Acquired Methicillin-Resistant Staphylococcus aureus in Infected Mouse Skin Abrasions. Photomed. Laser Surg. 2013, 31, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Makdoumi, K.; Goodrich, R.; Backman, A. Photochemical Eradication of Methicillin-resistant Staphylococcus aureus by Blue Light Activation of Riboflavin. Acta Ophthalmol. 2017, 95, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Dogru, E.; Demirbas, A.; Altinsoy, B.; Duman, F.; Ocsoy, I. Formation of Matricaria Chamomilla Extract-Incorporated Ag Nanoparticles and Size-Dependent Enhanced Antimicrobial Property. J. Photochem. Photobiol. B Biol. 2017, 174, 78–83. [Google Scholar] [CrossRef]
- Yang, P.; Wang, N.; Wang, C.; Yao, Y.; Fu, X.; Yu, W.; Cai, R.; Yao, M. 460 Nm Visible Light Irradiation Eradicates MRSA via Inducing Prophage Activation. J. Photochem. Photobiol. B Biol. 2017, 166, 311–322. [Google Scholar] [CrossRef]
- Akhtar, F.; Khan, A.U.; Misba, L.; Akhtar, K.; Ali, A. Antimicrobial and Antibiofilm Photodynamic Therapy against Vancomycin Resistant Staphylococcus aureus (VRSA) Induced Infection in vitro and in vivo. Eur. J. Pharm. Biopharm. 2021, 160, 65–76. [Google Scholar] [CrossRef]
- Almeida, P.P.; Pereira, Í.S.; Rodrigues, K.B.; Leal, L.S.; Marques, A.S.; Rosa, L.P.; da Silva, F.C.; da Silva, R.A.A. Photodynamic Therapy Controls of Staphylococcus aureus Intradermal Infection in Mice. Lasers Med. Sci. 2017, 32, 1337–1342. [Google Scholar] [CrossRef]
- Leanse, L.G.; Zeng, X.; Dai, T. Potentiated Antimicrobial Blue Light Killing of Methicillin Resistant Staphylococcus aureus by Pyocyanin. J. Photochem. Photobiol. B Biol. 2021, 215, 112109. [Google Scholar] [CrossRef]
- Lei, X.; Qiu, L.; Lan, M.; Du, X.; Zhou, S.; Cui, P.; Zheng, R.; Jiang, P.; Wang, J.; Xia, J. Antibacterial Photodynamic Peptides for Staphylococcal Skin Infection. Biomater. Sci. 2020, 8, 6695–6702. [Google Scholar] [CrossRef]
- Cadet, J.; Mouret, S.; Ravanat, J.L.; Douki, T. Photoinduced Damage to Cellular DNA: Direct and Photosensitized Reactions. Photochem. Photobiol. 2012, 88, 1048–1065. [Google Scholar] [CrossRef]
- Yin, R.; Dai, T.; Avci, P.; Jorge, A.E.S.; De Melo, W.C.M.A.; Vecchio, D.; Huang, Y.Y.; Gupta, A.; Hamblin, M.R. Light Based Anti-Infectives: Ultraviolet C Irradiation, Photodynamic Therapy, Blue Light, and Beyond. Curr. Opin. Pharmacol. 2013, 13, 731–762. [Google Scholar] [CrossRef]
- Song, C.; Wen, R.; Zhou, J.; Zeng, X.; Kou, Z.; Li, Y.; Yun, F.; Wu, R. UV C Light from a Light-Emitting Diode at 275 Nanometers Shortens Wound Healing Time in Bacterium- and Fungus-Infected Skin in Mice. Microbiol. Spectr. 2022, 10, e0342422. [Google Scholar] [CrossRef]
- Chen, Y.; Gao, Y.; Chen, Y.; Liu, L.; Mo, A.; Peng, Q. Nanomaterials-Based Photothermal Therapy and Its Potentials in Antibacterial Treatment. J. Control. Release 2020, 328, 251–262. [Google Scholar] [CrossRef]
- Naskar, A.; Lee, S.; Kim, K. sun Au-Zno Conjugated Black Phosphorus as a near-Infrared Light-Triggering and Recurrence-Suppressing Nanoantibiotic Platform against Staphylococcus aureus. Pharmaceutics 2021, 13, 52. [Google Scholar] [CrossRef]
- Kim, M.-H.; Yamayoshi, I.; Mathew, S.; Liln, H.; Nayfach, J.; Simon, S.I. Magnetic Nanoparticle Targeted Hyperthermia of Cutaneous Staphylococcus aureus Infection. Ann. Biomed. Eng. 2013, 41, 598–609. [Google Scholar] [CrossRef]
- Bárcena, C.; Sra, A.K.; Gao, J. Applications of Magnetic Nanoparticles in Biomedicine. Nanoscale Magn. Mater. Appl. 2009, 167, 591–626. [Google Scholar] [CrossRef]
- Dudeck, O.; Bogusiewicz, K.; Pinkernelle, J.; Gaffke, G.; Pech, M.; Wieners, G.; Bruhn, H.; Jordan, A.; Ricke, J. Local Arterial Infusion of Superparamagnetic Iron Oxide Particles in Hepatocellular Carcinoma: A Feasibility and 3.0 T MRI Study. Investig. Radiol. 2006, 41, 527–535. [Google Scholar] [CrossRef]
- Jordan, A.; Scholz, R.; Maier-Hauff, K.; van Landeghem, F.K.H.; Waldoefner, N.; Teichgraeber, U.; Pinkernelle, J.; Bruhn, H.; Neumann, F.; Thiesen, B.; et al. The Effect of Thermotherapy Using Magnetic Nanoparticles on Rat Malignant Glioma. J. Neurooncol. 2006, 78, 7–14. [Google Scholar] [CrossRef]
- Maier-Hauff, K.; Rothe, R.; Scholz, R.; Gneveckow, U.; Wust, P.; Thiesen, B.; Feussner, A.; Deimling, A.; Waldoefner, N.; Felix, R.; et al. Intracranial Thermotherapy Using Magnetic Nanoparticles Combined with External Beam Radiotherapy: Results of a Feasibility Study on Patients with Glioblastoma Multiforme. J. Neurooncol. 2007, 81, 53–60. [Google Scholar] [CrossRef]
- Miller, L.S.; O’Connell, R.M.; Gutierrez, M.A.; Pietras, E.M.; Shahangian, A.; Gross, C.E.; Thirumala, A.; Cheung, A.L.; Cheng, G.; Modlin, R.L. MyD88 Mediates Neutrophil Recruitment Initiated by IL-1R but Not TLR2 Activation in Immunity against Staphylococcus aureus. Immunity 2006, 24, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Bernthal, N.M.; Stavrakis, A.I.; Billi, F.; Cho, J.S.; Kremen, T.J.; Simon, S.I.; Cheung, A.L.; Finerman, G.A.; Lieberman, J.R.; Adams, J.S.; et al. A Mouse Model of Post-Arthroplasty Staphylococcus aureus Joint Infection to Evaluate in vivo the Efficacy of Antimicrobial Implant Coatings. PLoS ONE 2010, 5, e12580. [Google Scholar] [CrossRef] [PubMed]
- Yuan, K.; Mei, Q.; Guo, X.; Xu, Y.; Yang, D.; Sánchez, B.J.; Sheng, B.; Liu, C.; Hu, Z.; Yu, G.; et al. Antimicrobial Peptide Based Magnetic Recognition Elements and Au@Ag-GO SERS Tags with Stable Internal Standards: A Three in One Biosensor for Isolation, Discrimination and Killing of Multiple Bacteria in Whole Blood. Chem. Sci. 2018, 9, 8781–8795. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Pang, S.; Pearson, B.; Chujo, Y.; McLandsborough, L.; Fan, M.; He, L. Rapid Concentration Detection and Differentiation of Bacteria in Skimmed Milk Using Surface Enhanced Raman Scattering Mapping on 4-Mercaptophenylboronic Acid Functionalized Silver Dendrites. Anal. Bioanal. Chem. 2017, 409, 2229–2238. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, J.; Yang, X.; Hou, Q.; Liu, S.; Zheng, W.; Long, Y.; Jiang, X. Mercaptophenylboronic Acid-Activated Gold Nanoparticles as Nanoantibiotics against Multidrug-Resistant Bacteria. ACS Appl. Mater. Interfaces 2020, 12, 51148–51159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, Y.; Cha, R.; Long, K.; Li, J.; Jiang, X. Manufacture of Hydrophobic Nanocomposite Films with High Printability. ACS Sustain. Chem. Eng. 2019, 7, 15404–15412. [Google Scholar] [CrossRef]
- Li, Z.; Wang, J.; Dai, L.; Sun, X.; An, M.; Duan, C.; Li, J.; Ni, Y. Asymmetrically Patterned Cellulose Nanofibers/Graphene Oxide Composite Film for Humidity Sensing and Moist-Induced Electricity Generation. ACS Appl. Mater. Interfaces 2020, 12, 55205–55214. [Google Scholar] [CrossRef]
- Chen, N.; Wang, H.; Ling, C.; Vermerris, W.; Wang, B.; Tong, Z. Cellulose-Based Injectable Hydrogel Composite for PH-Responsive and Controllable Drug Delivery; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; Volume 225, ISBN 0135239249. [Google Scholar]
- Luo, H.; Cha, R.; Li, J.; Hao, W.; Zhang, Y.; Zhou, F. Advances in Tissue Engineering of Nanocellulose-Based Scaffolds: A Review. Carbohydr. Polym. 2019, 224, 115144. [Google Scholar] [CrossRef]
- Luo, H.; Lan, H.; Cha, R.; Yu, X.; Gao, P.; Zhang, P.; Zhang, C.; Han, L.; Jiang, X. Dialdehyde Nanocrystalline Cellulose as Antibiotic Substitutes against Multidrug-Resistant Bacteria. ACS Appl. Mater. Interfaces 2021, 13, 33802–33811. [Google Scholar] [CrossRef]
- Bardhan, S.; Pal, K.; Roy, S.; Das, S.; Chakraborty, A.; Karmakar, P.; Basu, R.; Das, S. Nanoparticle Size-Dependent Antibacterial Activities in Natural Minerals. J. Nanosci. Nanotechnol. 2019, 19, 7112–7122. [Google Scholar] [CrossRef]
- Mcdonnell, G.; Russell, A.D. Antiseptics and Disinfectants: Activity, Action, and Resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef] [PubMed]
- Spangler, D.; Rothenburger, S.; Nguyen, K.; Jampani, H.; Weiss, S.; Bhende, S. In Vitro Antimicrobial Activity of Oxidized Regenerated Cellulose Against Antibiotic-Resistant Microorganisms. Surg. Infect. 2003, 4, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Ge, L.; Lyu, Y.; Zi, Y.; Li, X.; Li, D.; Mu, C. Preparation, Characterization and Antibacterial Activity of Oxidized κ-Carrageenan. Carbohydr. Polym. 2017, 174, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhao, L.; Chen, Z.; Zhang, L.; Li, L.; Zhao, M.; Yan, L.; Liao, L.; Zhang, C.; Wu, Z. Macrophage-Targeting Bioactive Glass Nanoparticles for the Treatment of Intracellular Infection and Subcutaneous Abscess. Biomater. Sci. 2022, 10, 6535–6548. [Google Scholar] [CrossRef]
- Wang, H.; Sun, Y.B.; Chen, Q.W.; Yu, Y.F.; Cheng, K. Synthesis of Carbon-Encapsulated Superparamagnetic Colloidal Nanoparticles with Magnetic-Responsive Photonic Crystal Property. Dalt. Trans. 2010, 39, 9565–9569. [Google Scholar] [CrossRef]
- Yin, I.X.; Zhang, J.; Zhao, I.S.; Mei, M.L.; Li, Q.; Chu, C.H. The Antibacterial Mechanism of Silver Nanoparticles and Its Application in Dentistry. Int. J. Nanomed. 2020, 15, 2555–2562. [Google Scholar] [CrossRef]
- Abtin, A.; Jain, R.; Mitchell, A.J.; Roediger, B.; Brzoska, A.J.; Tikoo, S.; Cheng, Q.; Guan Ng, L.; Cavanagh, L.L.; von Andrian, U.H.; et al. Perivascular Macrophages Mediate Neutrophil Recruitment during Bacterial Skin Infection. Nat. Inmunol. 2014, 15, 45–53. [Google Scholar] [CrossRef]
- Nestle, F.O.; Di Meglio, P.; Qin, J.-Z.; Nickoloff, B.J. Skin Immune Sentinels in Health and Disease. Nat. Rev. Immunol. 2009, 9, 679–691. [Google Scholar] [CrossRef]
- Hassanshahi, A.; Moradzad, M.; Ghalamkari, S.; Fadaei, M.; Cowin, A.J.; Hassanshahi, M. Macrophage-Mediated Inflammation in Skin Wound Healing. Cells 2022, 11, 2953. [Google Scholar] [CrossRef]
- Vergunst, A.C.; Carranza, N.L.; Zhang, L.; Gomes, M.C.; Tasrini, Y.; Meijer, A.H.; Callaghan, D.O. Macrophages as Drivers of an Opportunistic Infection. Microb. Cell 2017, 4, 362–364. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Ren, Y.; Su, L.; Li, A.; An, Y.; Rotello, V.; Zhang, Z.; Wang, Y.; Liu, Y.; et al. Coating of a Novel Antimicrobial Nanoparticle with a Macrophage Membrane for the Selective Entry into Infected Macrophages and Killing of Intracellular Staphylococci. Adv. Funct. Mater. 2020, 30, 2004942. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, B.H.; Huh, B.K.; Yoo, Y.C.; Heo, C.Y.; Choy, Y.B.; Park, J.H. Surgical Suture Releasing Macrophage-Targeted Drug-Loaded Nanoparticles for an Enhanced Anti-Inflammatory Effect. Biomater. Sci. 2017, 5, 1670–1677. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, H.; Ji, B.; Qian, W.; Xia, S.; Wang, L.; Xu, Y.; Chen, J.; Yang, L.; Mao, H. Targeted Imaging of CD206 Expressing Tumor-Associated M2-like Macrophages Using Mannose-Conjugated Antibiofouling Magnetic Iron Oxide Nanoparticles. ACS Appl. Bio Mater. 2020, 3, 4335–4347. [Google Scholar] [CrossRef] [PubMed]
- Ruan, G.X.; Chen, Y.Z.; Yao, X.L.; Du, A.; Tang, G.P.; Shen, Y.Q.; Tabata, Y.; Gao, J.Q. Macrophage Mannose Receptor-Specific Gene Delivery Vehicle for Macrophage Engineering. Acta Biomater. 2014, 10, 1847–1855. [Google Scholar] [CrossRef]
- Liang, S.; He, Y.; Xia, Y.; Wang, H.; Wang, L.; Gao, R.; Zhang, M. Inhibiting the Growth of Methicillin-Resistant Staphylococcus aureus in vitro with Antisense Peptide Nucleic Acid Conjugates Targeting the FtsZ Gene. Int. J. Infect. Dis. 2015, 30, e1–e6. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, W.; Zhu, Y.; Shi, S.; Li, Q.; Mao, C.; Zhao, D.; Zhan, Y.; Shi, J.; Li, W.; et al. Inhibiting Methicillin-Resistant Staphylococcus aureus by Tetrahedral DNA Nanostructure-Enabled Antisense Peptide Nucleic Acid Delivery. Nano Lett. 2018, 18, 5652–5659. [Google Scholar] [CrossRef]
- Long, Q.; Jia, B.; Shi, Y.; Wang, Q.; Yu, H.; Li, Z. DNA Nanodevice as a Co-Delivery Vehicle of Antisense Oligonucleotide and Silver Ions for Selective Inhibition of Bacteria Growth. ACS Appl. Mater. Interfaces 2021, 13, 47987–47995. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic | Mechanism of Action | Therapeutical Problem | Alternative Strategies | Ref. |
|---|---|---|---|---|
| β-Lactam (penicillins, cephalosporins, and carbapenems) | Interference with the synthesis of peptidoglycan | Antibiotic resistance: 1. β- Lactamases, blaZ | (1a) Combination with metal nanoparticles. (1b) Chemical enzyme inhibitors. (1c) Inhibition of blaZ expresion. | (1a) [11,12] (1b) [14] (1c) [13] |
| 2. mecA product PBP2a reduced affinity for PBP | (2a) Antisense technology to block PDP2a expression. (2b) Combination with ZA-S. (2c) Non-antibiotic-based treatments. | (2a) [16,17] (2b) [18] (2c) See Section 2. | ||
| Vancomycin (Vm) | Blocking of peptidoglycan crosslink formation through binding to (D-Ala-D-Ala) | 1. Elevated toxicity and stability problems | (1a) Enhance antimicrobial activity with lipid-based nanoformulations. Vm-loaded polymersomes and OLA-LPHNVs1-Vm nanovesicles. | (1a) [50,51] |
| (1b) Local treatments to enhance tissue penetration and skin accumulation of ATB. Vm-loaded nanobubbles and US and Vm-loaded chitosan nanoparticles coated with lecithin (CLNPs). | (1b) [54,55] | |||
| 2. Antibiotic resistance. | 2. Non-antibiotic-based treatments Blue-light-mediated VISA and VRSA killing or in combination with PS. | 2. [131,132,133] | ||
| Quinolones | Inhibition of bacterial replication | High toxicity and low solubility | Nanoformulations that increase biodisponibility and reduce antibiotic dosis: (1a) Solid dispersions (SD); AgNPs-PEG-Cip (1b) Lipidbased formulations; Pluronic F127/Cremophor EL-Nfx and niosomal vesicles (MNV) as Mox nanocarriers. (1c) Complex formations with cyclodextrins; CD-Nfx, CD-Mox | (1a) [25] (1b) [32,33] (1c) [30,31] |
| Macrolides | Inhibition of protein synthesis 50S inhibition | High toxicity and solubility problems | 1. Nanoformulations that increase bioavailability and reduce antibiotic dosage: (1a) Solid dispersions (SD); AZO-Em. (1b) Complex formations with modified cyclodextrins; CD-MAN-Em, CD-GLU-Em. | (1a) [23] (1b) [43] |
| 2. Local treatments that enhance tissue penetration and skin accumulation of ATB; Azt - Microemulsions (MEs). | 2. [59] | |||
| Tetracyclines | Inhibition of protein synthesis 30S inhibition | High toxicity and solubility problems | 1. Local treatments that enhance tissue penetration and skin accumulation of ATB. Tig-loaded chitosan nanoparticles coated with lecithin (CLNPs). | 1. [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cela, E.M.; Urquiza, D.; Gómez, M.I.; Gonzalez, C.D. New Weapons to Fight against Staphylococcus aureus Skin Infections. Antibiotics 2023, 12, 1477. https://doi.org/10.3390/antibiotics12101477
Cela EM, Urquiza D, Gómez MI, Gonzalez CD. New Weapons to Fight against Staphylococcus aureus Skin Infections. Antibiotics. 2023; 12(10):1477. https://doi.org/10.3390/antibiotics12101477
Chicago/Turabian StyleCela, Eliana M., Dolores Urquiza, Marisa I. Gómez, and Cintia D. Gonzalez. 2023. "New Weapons to Fight against Staphylococcus aureus Skin Infections" Antibiotics 12, no. 10: 1477. https://doi.org/10.3390/antibiotics12101477
APA StyleCela, E. M., Urquiza, D., Gómez, M. I., & Gonzalez, C. D. (2023). New Weapons to Fight against Staphylococcus aureus Skin Infections. Antibiotics, 12(10), 1477. https://doi.org/10.3390/antibiotics12101477