
Antimicrobial Activity of Catechol-Containing Biopolymer Poly[3-(3,4-dihydroxyphenyl)glyceric Acid] from Different Medicinal Plants of Boraginaceae Family

 , , ,
, , ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
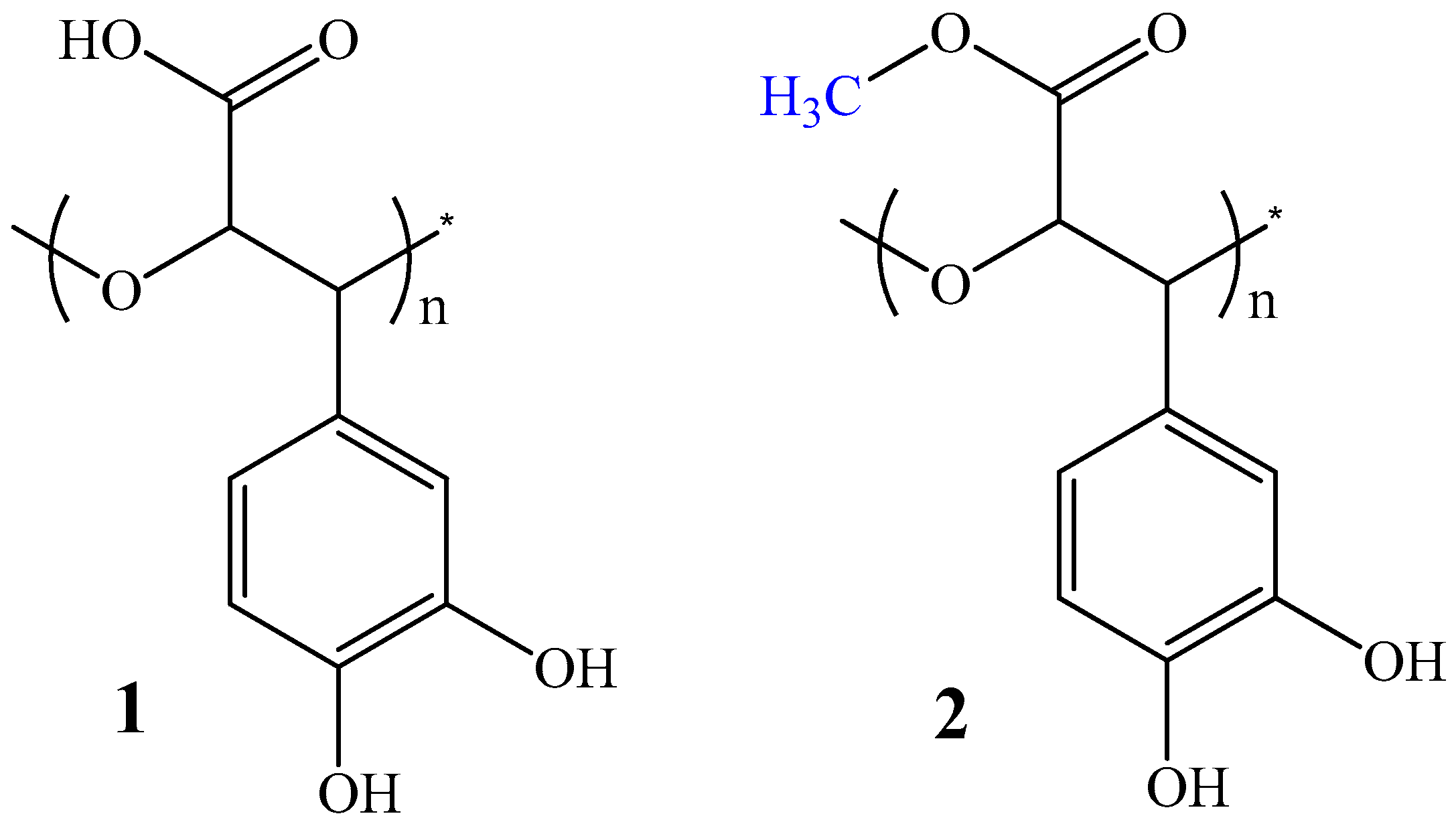
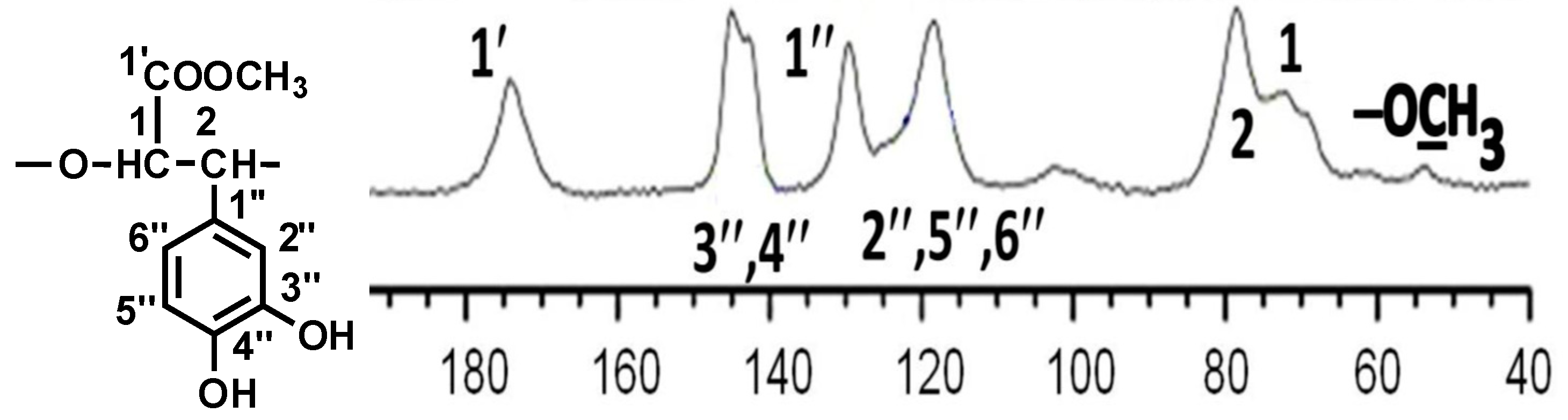
2.1. Chemistry
2.2. Biological Evaluation
2.2.1. Antibacterial Activity Evaluation
2.2.2. Antifungal Activity Evaluation
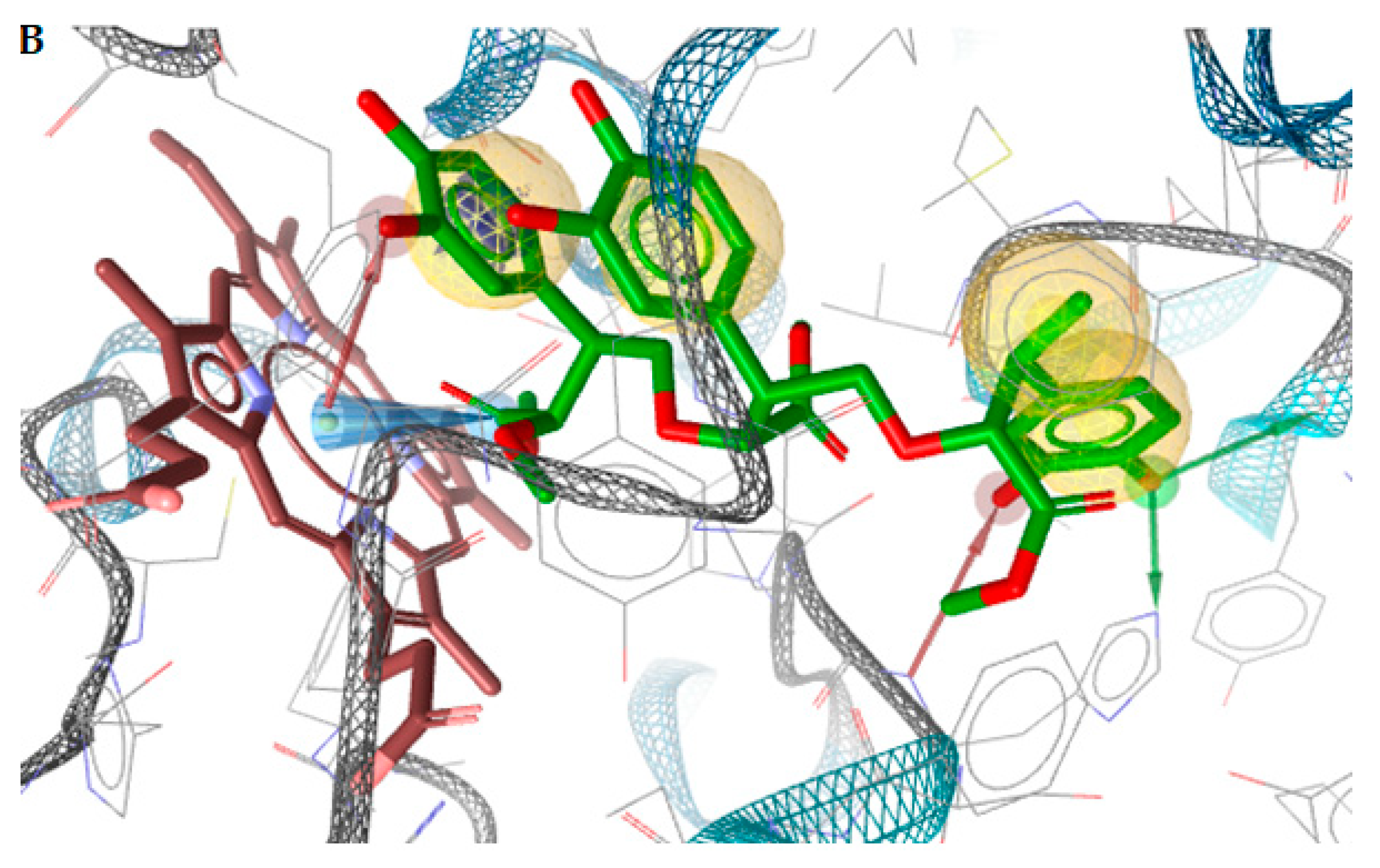
2.3. Docking Studies
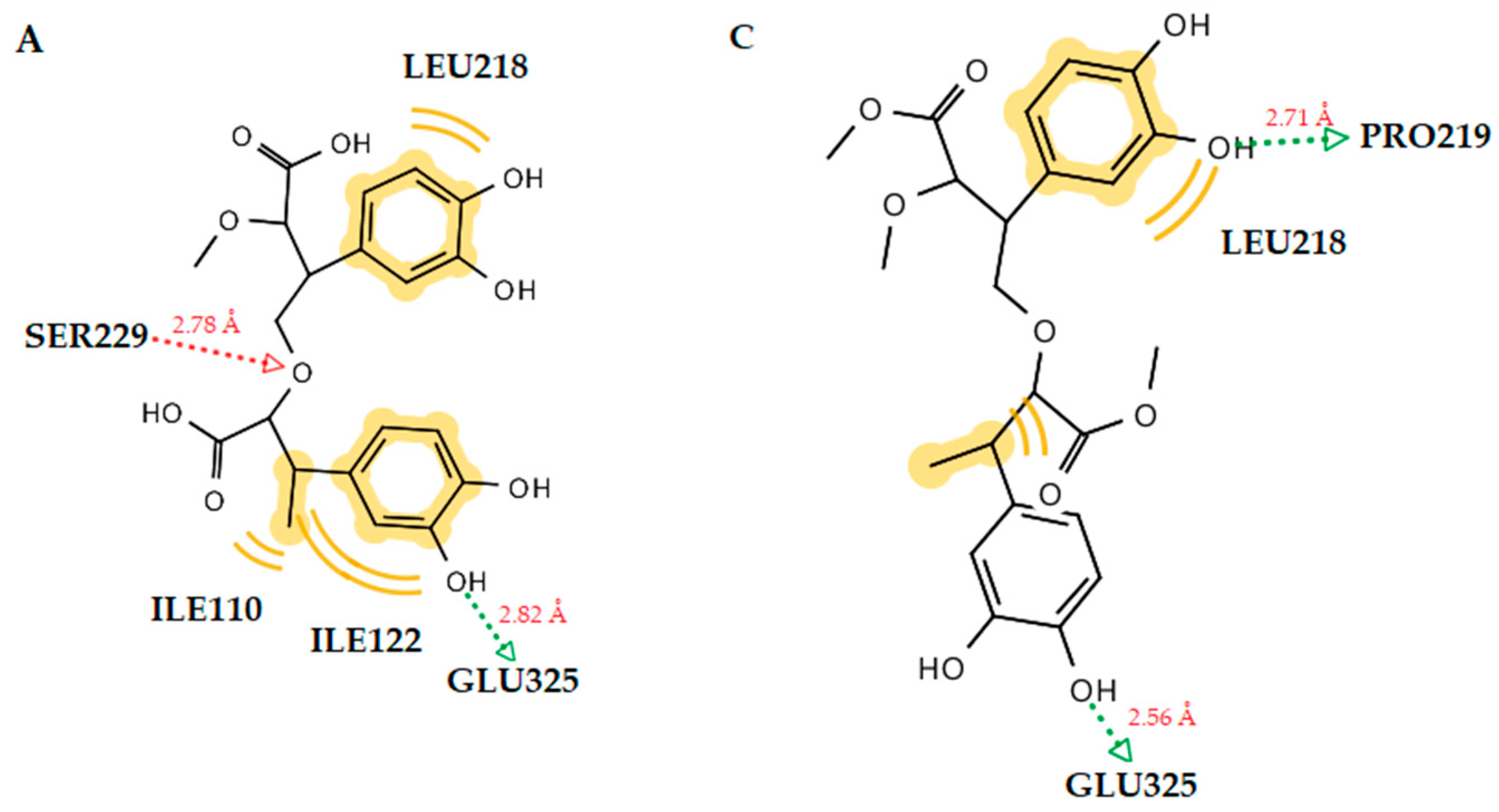
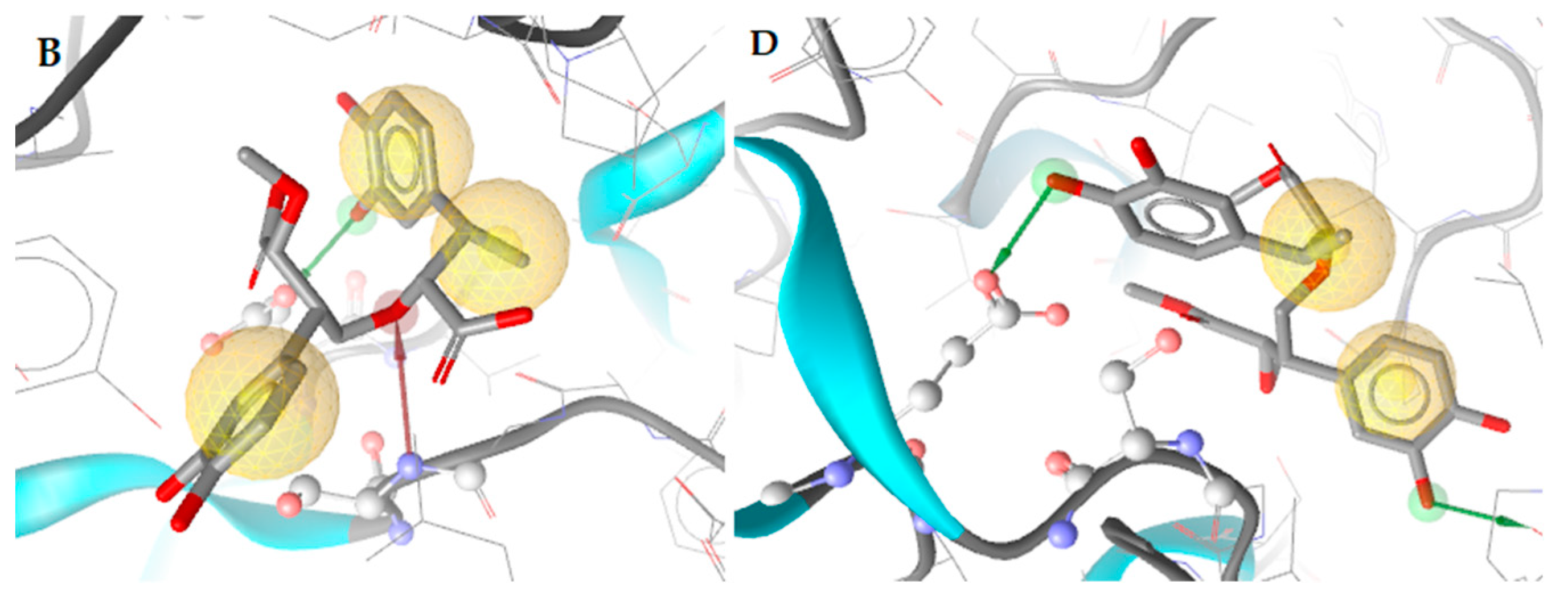
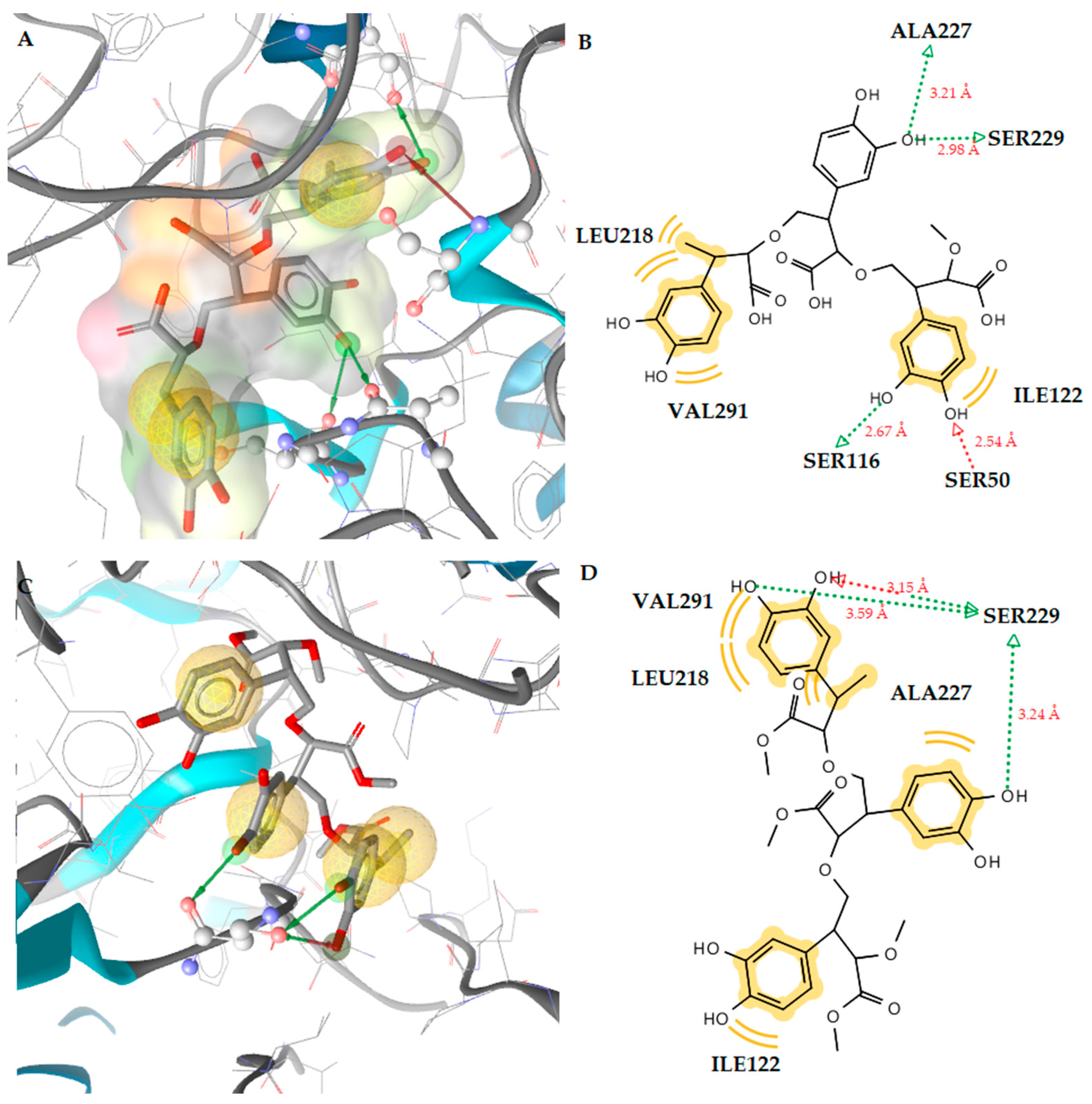
2.3.1. Docking to Antibacterial Targets
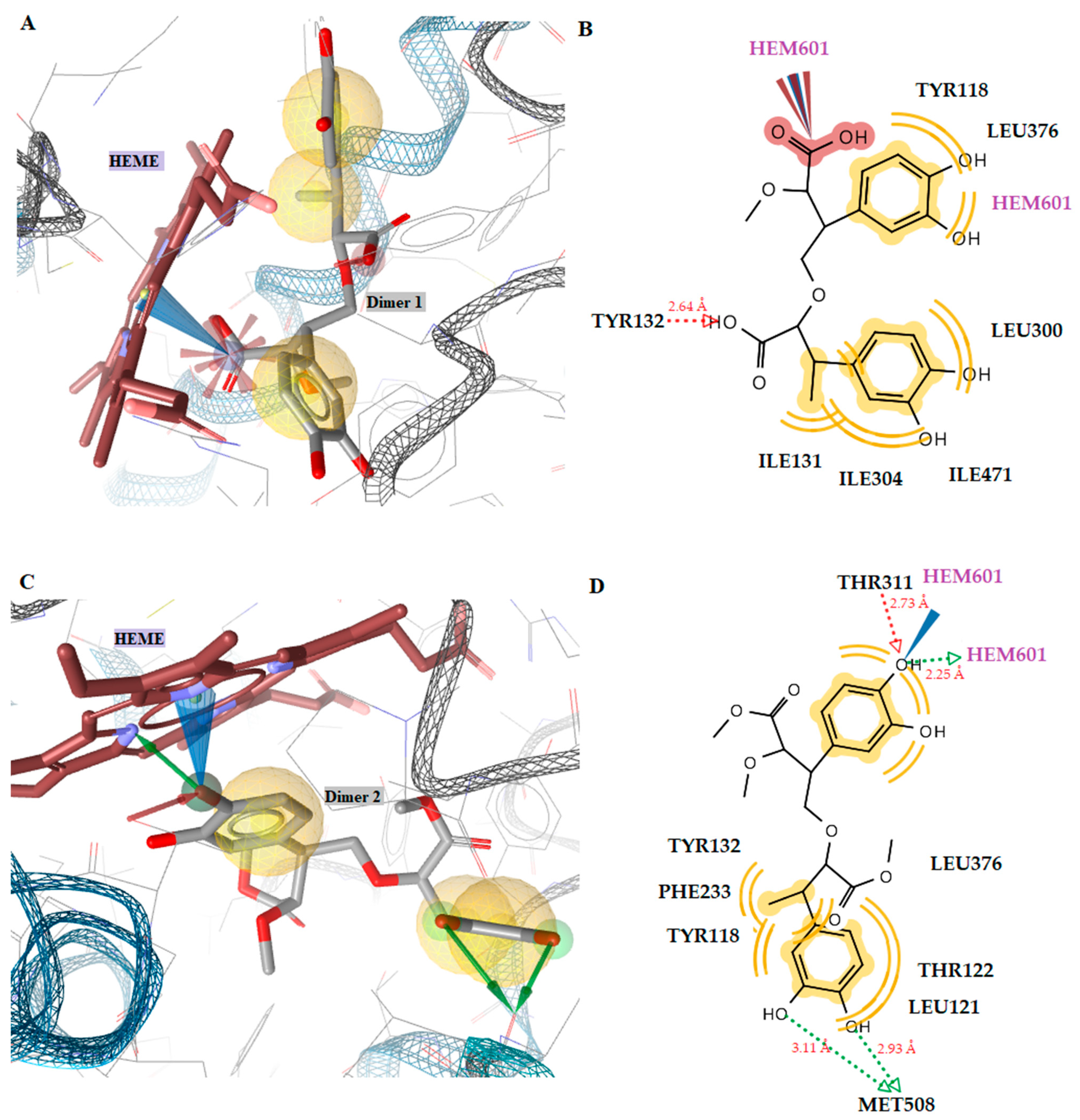
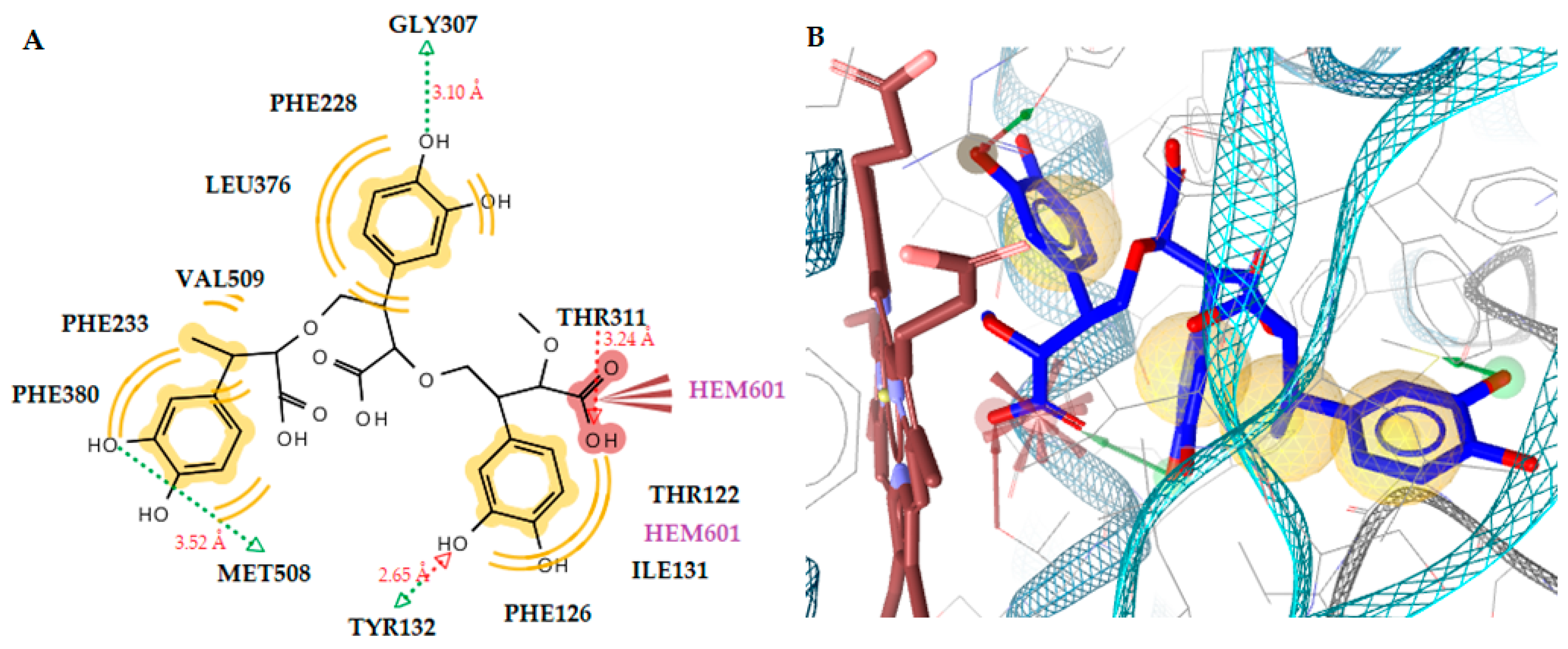
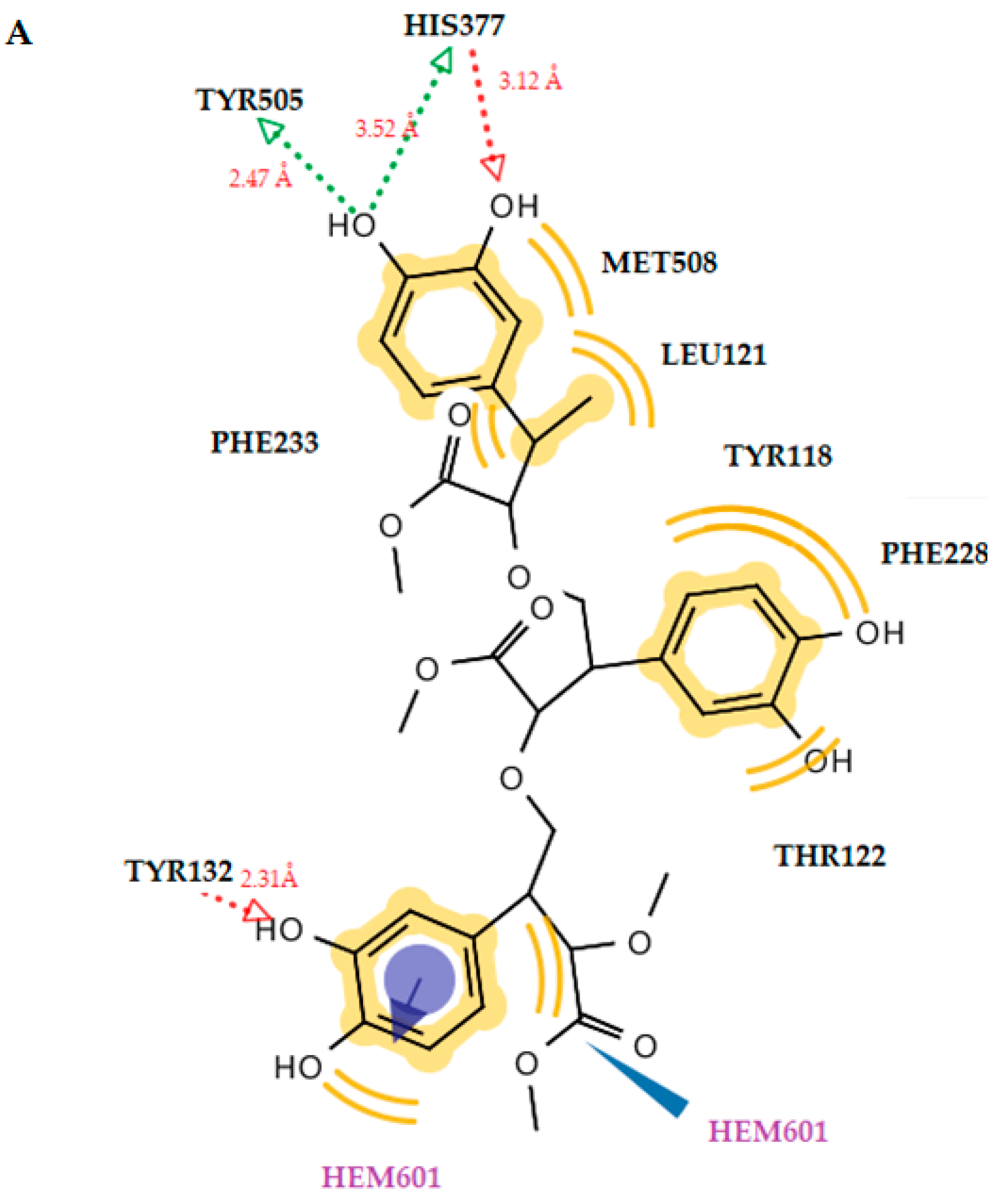
2.3.2. Docking to Antifungal Targets
2.4. Prediction of Toxicity
- Class I: fatal if swallowed (LD50 ≤ 5)
- Class II: fatal if swallowed (5 < LD50 ≤ 50)
- Class III: toxic if swallowed (50 < LD50 ≤ 300)
- Class IV: harmful if swallowed (300 < LD50 ≤ 2000)
- Class V: may be harmful if swallowed (2000 < LD50 ≤ 5000)
- Class VI: non-toxic (LD50 > 5000)
3. Materials and Methods
3.1. Plant Material
3.2. Methods
3.3. Biological Evaluation
3.3.1. Evaluation of Antimicrobial Activity
3.3.2. Evaluation of Antifungal Activity
3.4. Docking Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bibire, T.; Yilmaz, O.; Ghiciuc, C.M.; Bibire, N.; Danila, R. Biopolymers for Surgical Applications. Coatings 2022, 12, 211. [Google Scholar] [CrossRef]
- Varma, K.; Gopi, S. Biopolymers and their role in medicinal and pharmaceutical applications. In Biopolymers and Their Industrial Applications from Plant, Animal, and Marine Sources, to Functional Products; Elsevier: Amsterdam, The Netherlands, 2021; Chapter 7; pp. 175–191. [Google Scholar]
- Maghchiche, A. A Review: Application of Biopolymers in the Pharmaceutical Formulation. J. Adv. Bio-Pharm. Pharmacovigil. 2019, 1, 15–25. [Google Scholar]
- Fonseca, C.; Lima, M.; Sousa, A.F.; Silvestre, A.; Coelho, J.; Serra, A.C. Cinnamic acid derivatives as promising building blocks for advanced polymers: Synthesis, properties and applications. Polym. Chem. 2019, 10, 1696–1723. [Google Scholar] [CrossRef]
- Kaneko, D.; Kinugawa, S.; Matsumoto, K.; Kaneko, T. Terminally-catecholized hyper-branched polymers with high performance adhesive characteristics. Plant Biotechnol. 2010, 27, 293–296. [Google Scholar] [CrossRef]
- Kaneko, T.; Kaneko, D.; Wang, S. High-performance lignin-mimetic polyesters. Plant Biotechnol. 2010, 27, 243–250. [Google Scholar] [CrossRef]
- Razaviamri, S.; Wang, K.; Liu, B.; Lee, B.P. Catechol-Based Antimicrobial Polymers. Molecules 2021, 26, 559. [Google Scholar] [CrossRef]
- Chibane, L.B.; Degraeve, P.; Ferhout, H.; Bouajila, J.; Oulahal, N. Plant antimicrobial polyphenols as potential natural food preservatives. J. Sci. Food Agric. 2019, 99, 1457–1474. [Google Scholar] [CrossRef]
- Dresler, S.; Szymczak, G.; Wojcik, M. Comparison of some secondary metabolite content in the seventeen species of the Boraginaceae family. Pharm. Biol. 2017, 55, 691–695. [Google Scholar] [CrossRef]
- El-Shazly, A.; Wink, M. Diversity of Pyrrolizidine Alkaloids in the Boraginaceae Structures, Distribution, and Biological Properties. Diversity 2014, 6, 188–282. [Google Scholar] [CrossRef]
- Raphaelli, C.O.; Dannenberg, G.; Dalmazo, G.O.; Pereira, E.S.; Radünz, M.; Vizzotto, M.; Fiorentini, A.M.; Gandra, E.A.; Nora, L. Antibacterial and antioxidant properties of phenolic-rich extracts from apple (Malus domestica cv. Gala). Int. Food Res. J. 2019, 26, 1133–1142. [Google Scholar]
- Hussain, M.I.; Semreen, M.H.; Shanableh, A.; Khattak, M.N.K.; Ismail Saadoun, I.; Ahmady, I.M.; Muath Mousa, M.; Darwish, N.; Wameed Radeef, W.; Soliman, S.S.M. Phenolic Composition and Antimicrobial Activity of Different Emirati Date (Phoenix dactylifera L.) Pits: A Comparative Study. Plants 2019, 8, 497. [Google Scholar] [CrossRef] [PubMed]
- Dikpınar, T.; Süzgeç-Selçuk, S. Antimicrobial Activities of Medicinal Plants Containing Phenolic Compounds. J. Nat. Prod. 2020, 10, 514–534. [Google Scholar] [CrossRef]
- Salazar-Aranda, R.; Pérez-Lopez, L.A.; Lopez-Arroyo, J.; Alanic-Garza, B.A.; de Torres, N.W. Antimicrobial and Antioxidant Activities of Plants from Northeast of Mexico. Evid.-Based Complement. Altern. Med. 2011, 2011, 1–6. [Google Scholar]
- Gyawali, R.; Ibrahim, S.A. Natural products as antimicrobial agents. Food Control 2014, 46, 412–429. [Google Scholar] [CrossRef]
- Zambrano, C.; Kerekes, E.B.; Kotogán, A.; Papp, T.; Vágvölgyi, C.; Krisch, J.; Takó, M. Antimicrobial activity of grape, apple and pitahaya residue extracts after carbohydrase treatment against food-related bacteria. LWT Food Sci. Technol. 2019, 100, 416–425. [Google Scholar] [CrossRef]
- Takó, M.; Kerekes, E.B.; Zambrano, C.; Kotogán, A.; Papp, T.; Krisch, I.; Vágvölgyi, C. Plant Phenolics and Phenolic-Enriched Extracts as Antimicrobial Agents against Food-Contaminating Microorganisms. Antioxidants 2020, 9, 165. [Google Scholar] [CrossRef]
- Kabir, F.; Sultana, M.S.; Kurnianta, H. Antimicrobial activities of grape (Vitis vinifera L.) pomace polyphenols as a source of naturally occurring bioactive components. Afr. J. Biotechnol. 2015, 14, 2157–2161. [Google Scholar]
- Silván, J.M.; Mingo, E.; Hidalgo, M.; De Pascual-Teresa, S.; Carrascosa, A.V.; Martinez-Rodriguez, A.J. Antibacterial activity of a grape seed extract and its fractions against Campylobacter spp. Food Control 2013, 29, 25–31. [Google Scholar] [CrossRef]
- Levy, J.; Boyer, R.R.; Neilson, A.P.; O’Keefe, S.F.; Chu, H.S.S.; Williams, R.C.; Dorenkott, M.R.; Goodrich, K.M. Evaluation of peanut skin and grape seed extracts to inhibit growth of foodborne pathogens. Food Sci. Nutr. 2017, 5, 1130–1138. [Google Scholar] [CrossRef]
- Tenore, G.C.; Novellino, E.; Basile, A. Nutraceutical potential and antioxidant benefits of red pitaya (Hylocereus polyrhizus) extracts. J. Funct. Foods 2012, 4, 129–136. [Google Scholar] [CrossRef]
- Yong, Y.Y.; Dykes, G.; Lee, S.M.; Choo, W.S. Comparative study of betacyanin profile and antimicrobial activity of red pitahaya (Hylocereus polyrhizus) and red spinach (Amaranthus dubius). Plant Food. Hum. Nutr. 2017, 72, 41–47. [Google Scholar] [CrossRef]
- Merlani, M.; Barbakadze, V.; Amiranashvili, L.; Gogilashvili, L.; Poroikov, V.; Petrou, A.; Geronikaki, A.; Ćirić, A.; Glamočlija, J.; Soković, M. New Caffeic Acid Derivatives as Antimicrobial Agents: Design, Synthesis, Evaluation and Docking. Curr. Top. Med. Chem. 2019, 19, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Merlani, M.; Barbakadze, V.; Amiranashvili, L.; Gogilashvili, L.; Petrou, A.; Geronikaki, A.; Ćirić, A.; Glamočlija, J.; Soković, M. Caffeic and 3-(3,4-dihydroxyphenyl)glyceric acid derivatives as antimicrobial agent: Biological evaluation and molecular docking studies. SAR QSAR Env. Res. 2022, 33, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Kepa, M.; MiklasiNska-Majdanik, M.; Wojtyczka, R.D.; Idzik, D.; Korzeniowski, K.; Smolen-Dzirba, I.; Wdsik, T.J. Antimicrobial Potential of Caffeic Acid against Staphylococcus aureus Clinical Strains. BioMed Res. Inter. 2018, 2018, 7413504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbakadze, V.; Gogilashvili, L.; Amiranashvili, L.; Merlani, M.; Getia, M.; Gogilashvili, A.; Salgado, A.; Chankvetadze, B. Biologically active sugar-based biopolyether poly [3-(3,4-dihydroxyphenyl)glyceric acid] from the stems and roots of Paracynoglossum imeretinum (Kusn.) M.Pop. Bull. Georg. Natl. Acad. Sci. 2022, 16, 110–115. [Google Scholar]
- Merlani, M.; Scheibel, D.M.; Barbakadze, V.; Gogilashvili, L.; Amiranashvili, L.; Geronikaki, A.; Catania, V.; Schillaci, D.; Gallo, G.; Gitsov, I. Enzymatic synthesis and antimicrobial activity of oligomer analogues of medicinal biopolymers from comfrey and their species of the Boraginaceae Family. Pharmaceutics 2022, 14, 115. [Google Scholar] [CrossRef]
- Barbakadze, V.; Gogilashvili, L.; Amiranashvili, L.; Merlani, M.; Li, S.P.; Chankvetadze, B. Fractionation of Biologically Active Poly[3-(3,4-Dihydroxyphenyl)Glyceric Acid] Preparation from Symphytum asperum, Simultaneous Determination of Molecular Weights and Contents of its Fractions Using HPSEC-MALLS-RID. Bull. Georg. Natl. Acad. Sci. 2021, 15, 69–75. [Google Scholar]
- Liu, B.; Zhou, C.; Zhang, Z.; Roland, J.D.; Lee, B.P. Antimicrobial property of halogenated catechols. Chem. Eng. J. 2021, 403, 126340. [Google Scholar] [CrossRef]
- Benson, T.E.; Walsh, C.T.; Massey, V. Kinetic characterization of wild-type and S229A mutant MurB: Evidence for the role of Ser 229 as a general acid. Biochemistry 1997, 36, 796–805. [Google Scholar] [CrossRef]
- Banerjee, P.; Eckert, A.O.; Schrey, A.K.; Preissner, R. ProTox-II: A webserver for the prediction of toxicity of chemicals. Nucleic Acids Res. 2018, 46, W257–W263. [Google Scholar] [CrossRef]
- GHS-UNECE. Available online: http://www.unece.org/trans/danger/publi/ghs/ghs_welcome_e.html (accessed on 10 November 2022).
- Barthomeuf, C.M.; Debiton, E.; Barbakadze, V.V.; Kemertelidze, E.P. Evaluation of the dietetic and therapeutic potential of a high molecular weight hydroxicinnamate-derived polymer from Symphytum asperum Lepech. regarding its antioxidant, antilipoperoxidant, antiinflammatory, and cytotoxic properties. J. Agric. Food Chem. 2001, 49, 3942–3946. [Google Scholar] [CrossRef]
- Kartsev, V.; Lichitsky, B.; Geronikaki, A.; Petrou, A.; Smiljkovic, M.; Kostic, M.; Radanovic, O.; Soković, M. Design, synthesis and antimicrobial activity of usnic acid derivatives. Med. Chem. Comm. 2018, 9, 870–882. [Google Scholar] [CrossRef]
- Fesatidou, M.; Zagaliotis, P.; Camoutsis, C.; Petrou, A.; Eleftheriou, P.; Tratrat, C.; Haroun, M.; Geronikaki, A.; Ciric, A.; Sokovic, M. 5-Adamantan thiadiazole-based thiazolidinones as antimicrobial agents. Design, synthesis, molecular docking and evaluation. Bioorg. Med. Chem. 2018, 26, 4664–4676. [Google Scholar] [PubMed]
- Zubenko, A.; Kartsev, V.; Petrou, A.; Geronikaki, A.; Ivanov, M.; Glamočlija, J.; Sokovic, M.; Divaeva, L.; Morkovnik, A.; Klimenko, A. Experimental and In Silico Evaluation of New Heteroaryl Benzothiazole Derivatives as Antimicrobial Agents. Antibiotics 2022, 11, 1654. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| С Atom No. | 13C Chemical Shift, δC, ppm |
|---|---|
| 1 | 72 |
| 2 | 78 |
| 1′ | 174 |
| (−OCH3) | 54 |
| 1″ | 130 |
| 2″, 5″, 6″ | 118 |
| 3″, 4″ | 143 |
| Compounds | S. a. | MRSA | E. coli | Rez. E. coli | P. a. | Rez. P. a. | |
|---|---|---|---|---|---|---|---|
| PDHPGA and PMDHPO of BO (stems) | MIC | 1.50 | 1.50 | 1.00 | 1.50 | 1.00 | 0.75 |
| MBC | 3.00 | 3.00 | 1.50 | 3.00 | 1.50 | 1.50 | |
| PDHPGA and PMDHPO of AI (roots) | MIC | - | - | 3.00 | - | - | 3.00 |
| MBC | - | - | 6.00 | - | - | 6.00 | |
| PDHPGA of CO (stems) | MIC | - | - | - | - | - | 3.00 |
| MBC | - | - | - | - | - | 6.00 | |
| PDHPGA of SA (roots) | MIC | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 | 3.00 |
| MBC | 9.00 | 9.00 | 9.00 | 9.00 | 12.00 | 6.00 | |
| PDHPGA of SC (roots) | MIC | - | - | - | - | - | 3.00 |
| MBC | - | - | - | - | - | 6.00 | |
| PDHPGA and PMDHPO of SG (stems) | MIC | 0.75 | 1.00 | 0.75 | 1.50 | 1.00 | 0.75 |
| MBC | 1.50 | 1.50 | 1.50 | 3.00 | 1.50 | 1.50 | |
| Ampicillin | MIC | 0.10 | - | 0.15 | 0.20 | 0.30 | 0.20 |
| MBC | 0.15 | - | 0.20 | - | 0.50 | - | |
| Streptomycin | MIC | 0.10 | 0.10 | 0.10 | 0.05 | 0.10 | 0.10 |
| MBC | 0.20 | - | 0.20 | 0.10 | 0.20 | 0.20 | |
| Compounds | A. fu. | A. n. | T. v. | P. f. | P. v. c. | C. a. | |
|---|---|---|---|---|---|---|---|
| PDHPGA and PMDHPO of BO (stems) | MIC | 0.75 | 1.00 | 0.37 | 0.37 | 0.37 | 0.37 |
| MFC | 1.50 | 1.50 | 0.75 | 0.75 | 0.75 | 0.75 | |
| PDHPGA and PMDHPO of AI (roots) | MIC | 1.50 | 2.25 | 0.37 | 0.75 | 1.10 | 0.19 |
| MFC | 3.00 | 3.00 | 0.75 | 1.50 | 3.00 | 0.37 | |
| PDHPGA of CO (stems) | MIC | 0.75 | 0.56 | 0.19 | 0.37 | 0.75 | 0.37 |
| MFC | 1.50 | 0.75 | 0.37 | 0.75 | 1.50 | 0.75 | |
| PDHPGA of SA (roots) | MIC | 3.00 | - | 0.75 | 2.25 | 2.25 | 0.09 |
| MFC | 4.50 | - | 1.50 | 4.50 | 4.50 | 0.19 | |
| PDHPGA of SC (roots) | MIC | 0.37 | 0.37 | 0.19 | 0.75 | 0.75 | 0.19 |
| MFC | 0.75 | 0.75 | 0.37 | 1.50 | 1.50 | 0.37 | |
| PDHPGA and PMDHPO of SG (stems) | MIC | 0.37 | 0.56 | 0.09 | 0.56 | 0.75 | 0.14 |
| MFC | 0.75 | 0.75 | 0.19 | 0.75 | 1.50 | 0.19 | |
| Ketoconazole | MIC | 0.20 | 0.20 | 1.00 | 0.20 | 0.20 | 1.00 |
| MFC | 0.50 | 0.50 | 1.50 | 0.50 | 0.30 | 2.00 | |
| Bifonazole | MIC | 0.15 | 0.15 | 0.15 | 0.20 | 0.10 | 0.20 |
| MFC | 0.20 | 0.20 | 0.20 | 0.25 | 0.20 | 0.30 | |
| No | Est. Binding Energy (kcal/mol) | Residues Involved in Hydrogen Bonds | Residues Involved in Hydrophobic Interactions | |||
|---|---|---|---|---|---|---|
| E. coli Gyrase 1KZN | S. aureus Thymidylate Kinase 4QGG | E. coli Primase 1DDE | E. coli MurB 2Q85 | |||
| 1 | −4.55 | −2.56 | −2.41 | −5.16 | Ser229 (O···H, 2.65Å) | Leu218, Ala227, Val291 |
| 1 (dimer) | −4.31 | −2.67 | - | −7.80 | Ser229 (O···H, 2.78Å), Glu325 (O···H, 2.82Å) | Ile110, Leu218, Ile122 |
| 1 (trimer) | −2.16 | − | − | −9.81 | Ser50 (O···H, 2.54Å), Ser116 (O···H, 2.67Å), Ser229 (O···H, 2.98Å), Ala227 (O···H, 3.21Å) | Leu218, Ile122, Val291 |
| 2 | −4.12 | −2.46 | −2.57 | −5.89 | Ser229 (O···H, 2.55Å) | Leu218, Ile122 |
| 2 (dimer) | −3.57 | −1.28 | −1.06 | −6.73 | Pro219 (O···H, 2.71Å), Glu325 (O···H, 2.56Å) | Leu218 |
| 2 (trimer) | −1.67 | - | - | −9.16 | Ser229 (O···H, 3.15Å), Ser229 (O···H, 3.59Å), Ser229 (O···H, 3.24Å) | Leu218, Ile122, Ala227, Val291 |
| N/N | Est. Binding Energy (kcal/mol) | Residues Involved in Hydrogen Bonds | Residues Involved in Hydrophobic Interactions | Interactions with HEM601 | |
|---|---|---|---|---|---|
| DNA TopoIV 1S16 | CYP51 of C. albicans 5V5Z | ||||
| 1 | −1.52 | −7.96 | Tyr132 (O···H, 2.64Å) | Tyr118, Ile131, Tyr132, Leu300, Leu376 | Negative ionizable, Hydrophobic |
| 1 (dimer) | - | −9.55 | Tyr132 (O···H, 2.64Å), | Tyr118, Ile131, Tyr132, Leu300, Leu304, Leu376, Ile407 | Negative ionizable, Hydrophobic, Fe-binding |
| 1 (trimer) | - | −9.13 | Tyr132 (O···H, 2.65Å), Gly307 (O···H, 3.10Å), Thr311 (O···H, 3.24Å), Met508 (O···H, 3.52Å) | Ile131, Thr122, Phe126, Phe380, Phe233, Phe228, Leu376, Val509, Met508 | Negative ionizable, Hydrophobic |
| 2 | −1.78 | −9.26 | Tyr132 (O···H, 2.67Å) | Tyr118, Leu121, Tyr122, Tyr132, Phe233, Thr311, Leu376 | Negative ionizable, Hydrophobic |
| 2 (dimer) | −1.78 | −11.35 | Thr311 (O···H, 2.73Å), Met508 (O···H, 3.93Å), Met508 (O···H, 3.11Å), Hem601 (O···H, 2.25Å) | Tyr118, Leu121, Tyr122, Tyr132, Phe233, Thr311, Leu376 | Hydrophobic, H-Bond, Fe-binding |
| 2 (trimer) | - | −11.10 | Tyr132 (O···H, 2.31Å), His377 (O···H, 3.12Å), His377 (O···H, 3.52Å), Tyr505 (O···H, 2.47Å) | Met508, Leu121, Tyr118, Phe228, Thr122, Phe233 | Hydrophobic, aromatic, Fe-binding |
| ketoconazole | - | −8.23 | Tyr64 (O···H, 2.51Å) | Tyr118, Ile131, Tyr132, Leu300, Ile304, Leu376, Met508 | Hydrophobic, aromatic |
| No | Predicted LD50 (mg/kg) | Predicted Toxicity Class | Hepatotoxicity | Carcinogenicity | Immunotoxicity | Mutagenicity | Cytotoxicity |
|---|---|---|---|---|---|---|---|
| 1 | 2000 | IV | Inactive (0.6) | Inactive (0.56) | Inactive (0.54) | Inactive (0.94) | Inactive (0.95) |
| 2 | 2000 | IV | Inactive (0.57) | Inactive (0.56) | Inactive (0.62) | Inactive (0.91) | Inactive (0.92) |
| 1 (dimer) | 3500 | V | Inactive (0.79) | Inactive (0.73) | Inactive (0.58) | Inactive (0.77) | Inactive (0.86) |
| 2 (dimer) | 3500 | V | Inactive (0.80) | Inactive (0.74) | Inactive (0.50) | Inactive (0.78) | Inactive (0.84) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbakadze, V.; Merlani, M.; Gogilashvili, L.; Amiranashvili, L.; Petrou, A.; Geronikaki, A.; Ćirić, A.; Glamočlija, J.; Soković, M. Antimicrobial Activity of Catechol-Containing Biopolymer Poly[3-(3,4-dihydroxyphenyl)glyceric Acid] from Different Medicinal Plants of Boraginaceae Family. Antibiotics 2023, 12, 285. https://doi.org/10.3390/antibiotics12020285
Barbakadze V, Merlani M, Gogilashvili L, Amiranashvili L, Petrou A, Geronikaki A, Ćirić A, Glamočlija J, Soković M. Antimicrobial Activity of Catechol-Containing Biopolymer Poly[3-(3,4-dihydroxyphenyl)glyceric Acid] from Different Medicinal Plants of Boraginaceae Family. Antibiotics. 2023; 12(2):285. https://doi.org/10.3390/antibiotics12020285
Chicago/Turabian StyleBarbakadze, Vakhtang, Maia Merlani, Lali Gogilashvili, Lela Amiranashvili, Anthi Petrou, Athina Geronikaki, Ana Ćirić, Jasmina Glamočlija, and Marina Soković. 2023. "Antimicrobial Activity of Catechol-Containing Biopolymer Poly[3-(3,4-dihydroxyphenyl)glyceric Acid] from Different Medicinal Plants of Boraginaceae Family" Antibiotics 12, no. 2: 285. https://doi.org/10.3390/antibiotics12020285
APA StyleBarbakadze, V., Merlani, M., Gogilashvili, L., Amiranashvili, L., Petrou, A., Geronikaki, A., Ćirić, A., Glamočlija, J., & Soković, M. (2023). Antimicrobial Activity of Catechol-Containing Biopolymer Poly[3-(3,4-dihydroxyphenyl)glyceric Acid] from Different Medicinal Plants of Boraginaceae Family. Antibiotics, 12(2), 285. https://doi.org/10.3390/antibiotics12020285