
Dinactin: A New Antitumor Antibiotic with Cell Cycle Progression and Cancer Stemness Inhibiting Activities in Lung Cancer

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
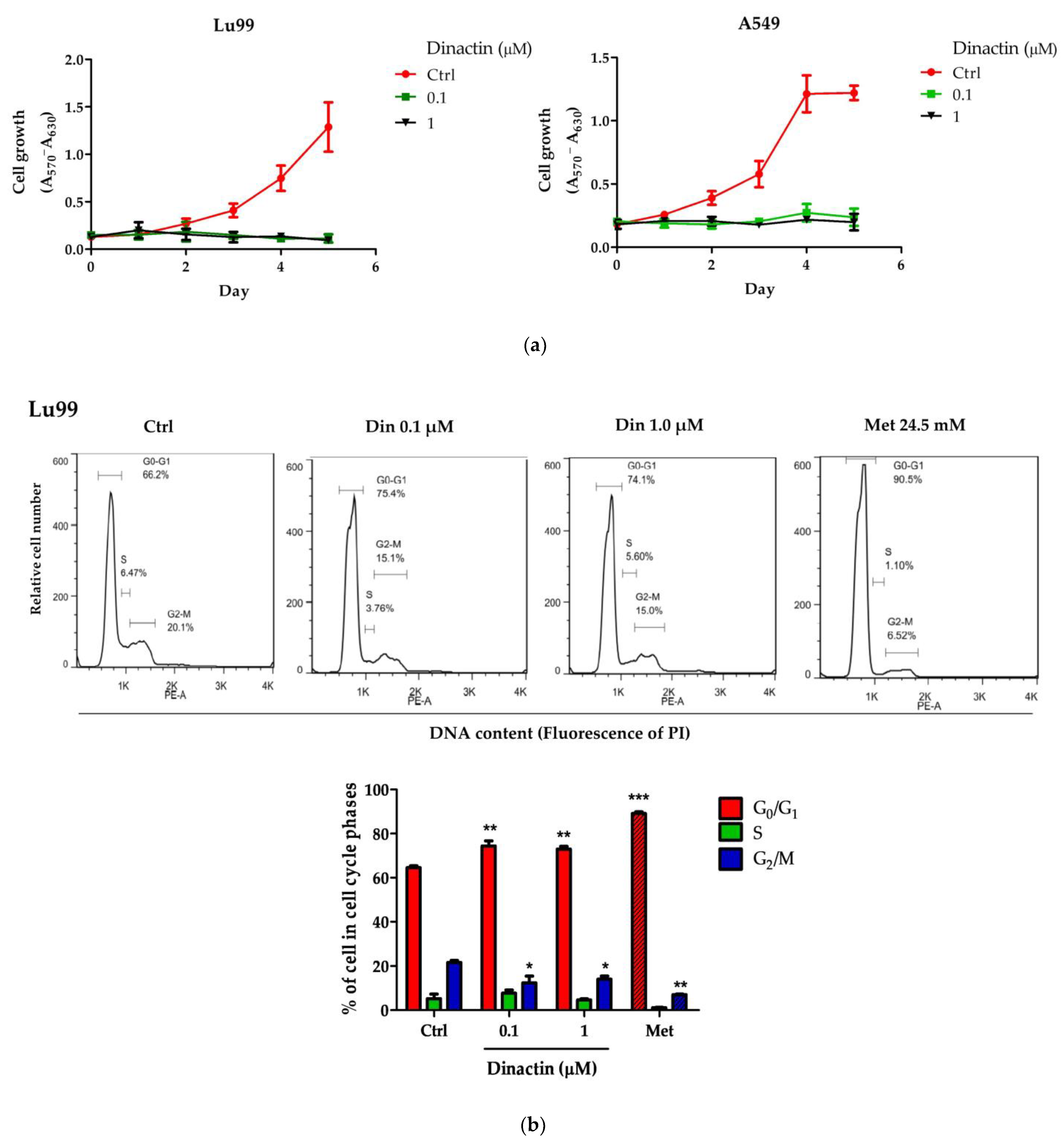
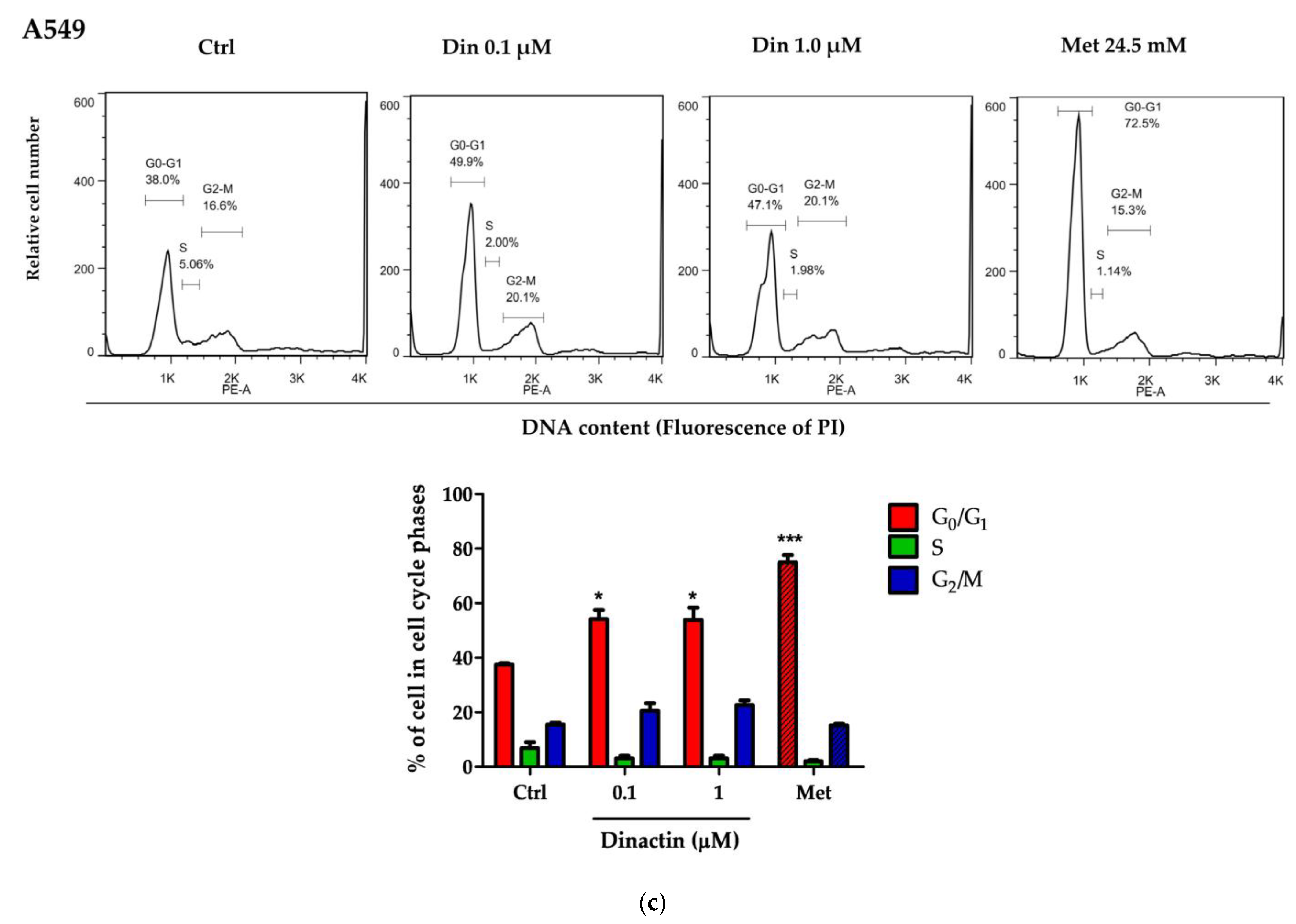
2.1. Dinactin Inhibited Cancer Cell Growth by Inducing G0/G1 Arrest
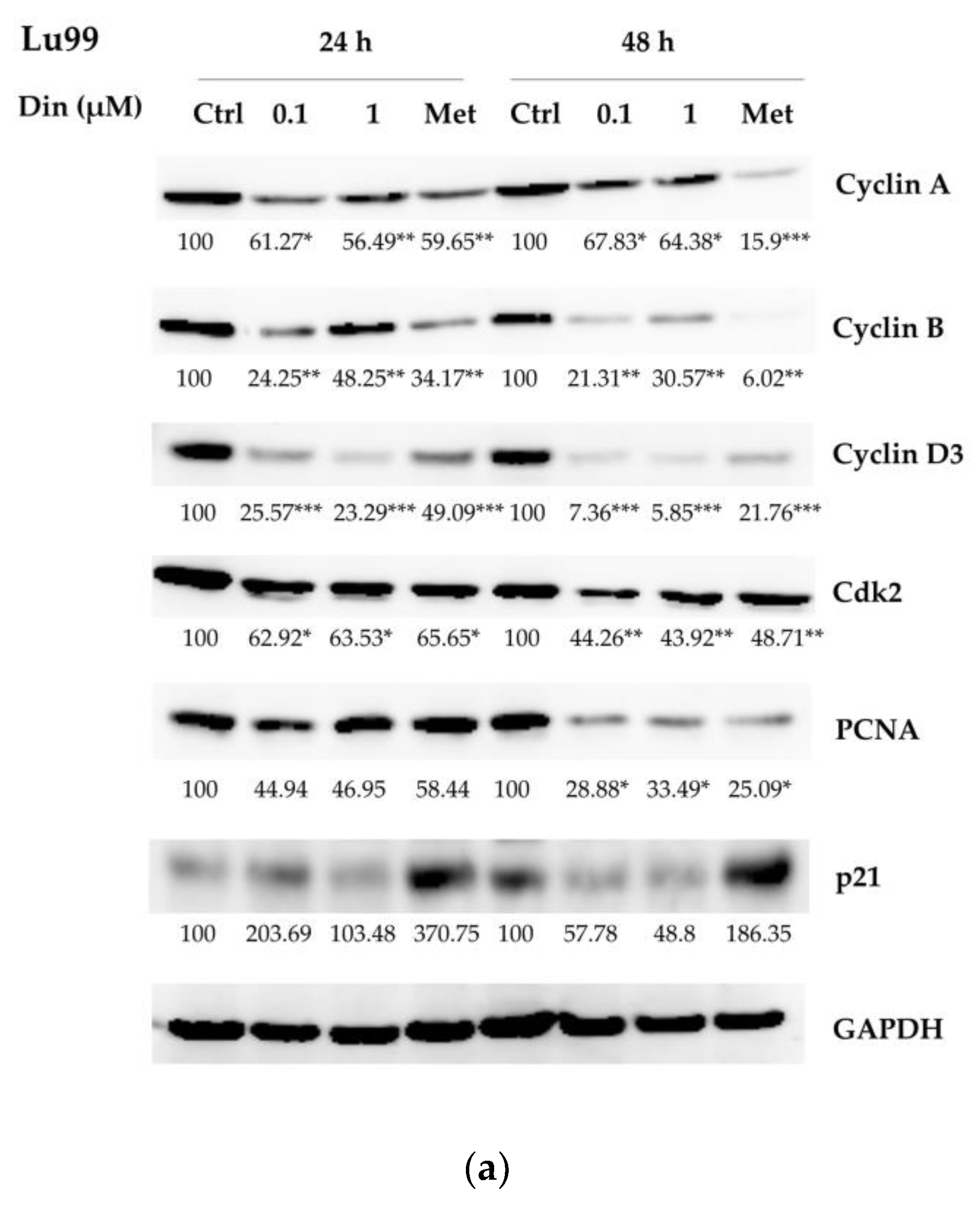
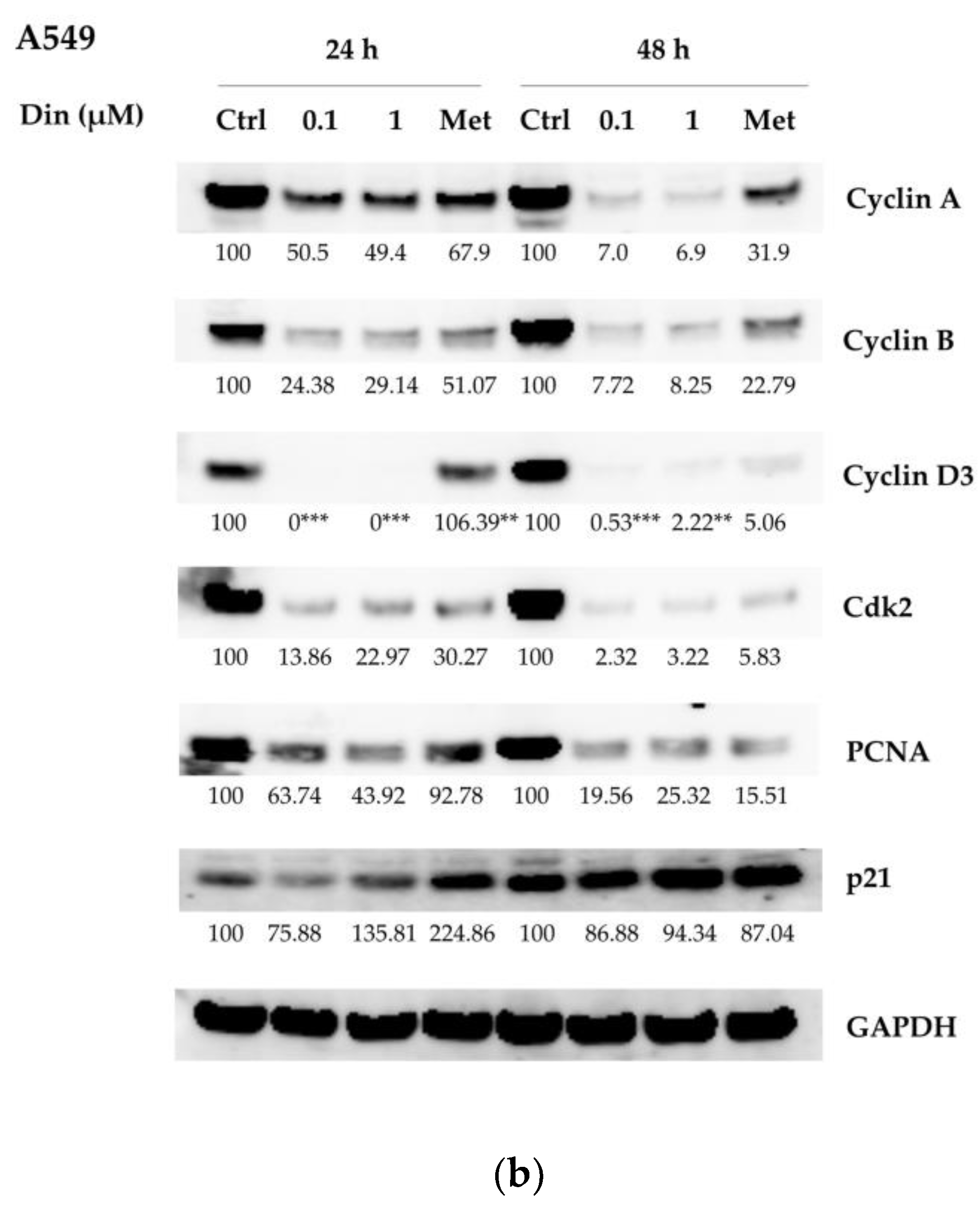
2.2. Dinactin Down-Regulated Cyclins A, B, and D3, cdk2 and PCNA in Lu99 and A549 Cells
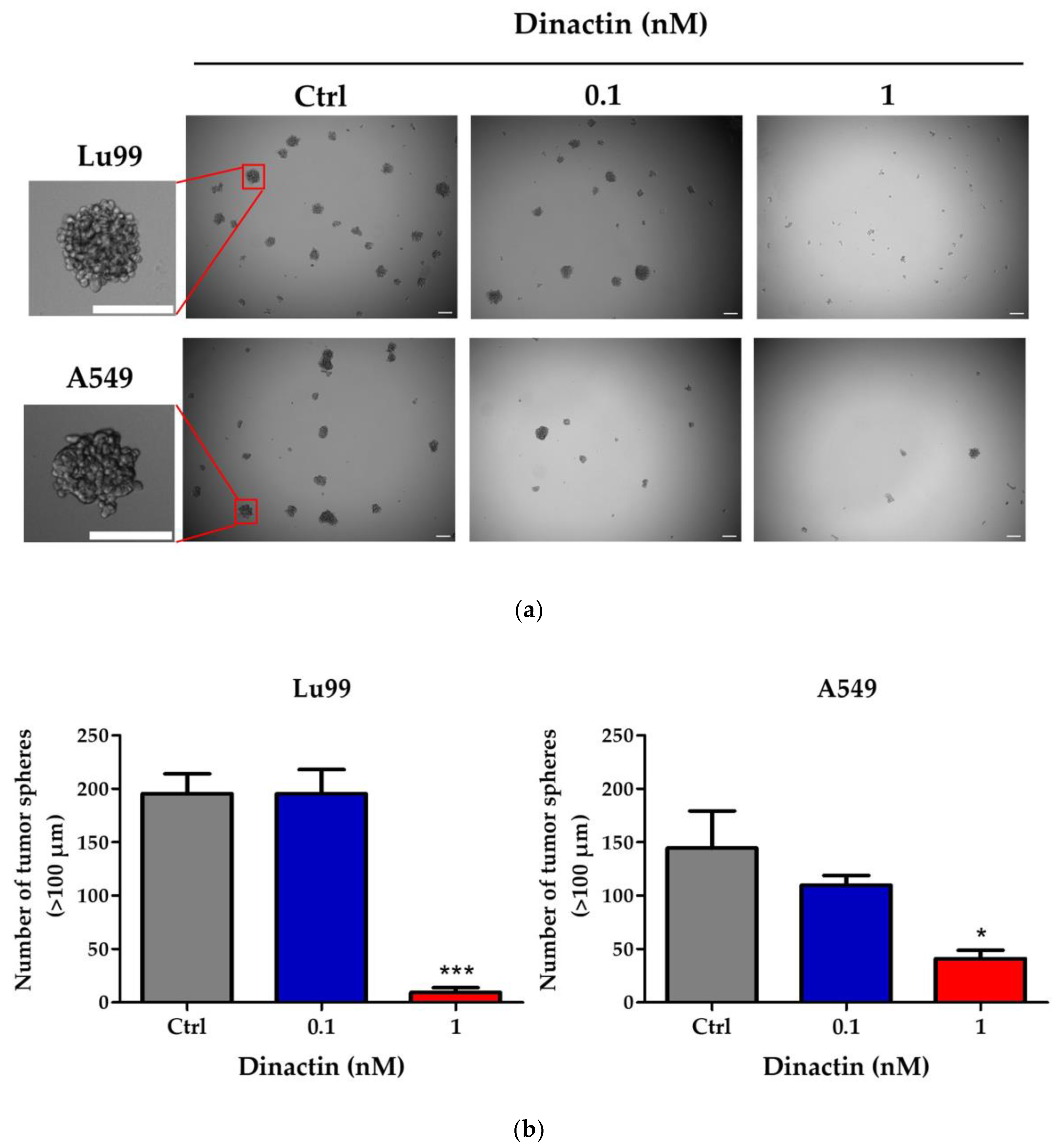
2.3. Dinactin Inhibited Tumor-Sphere Formation in Lu99 and A549 Cells
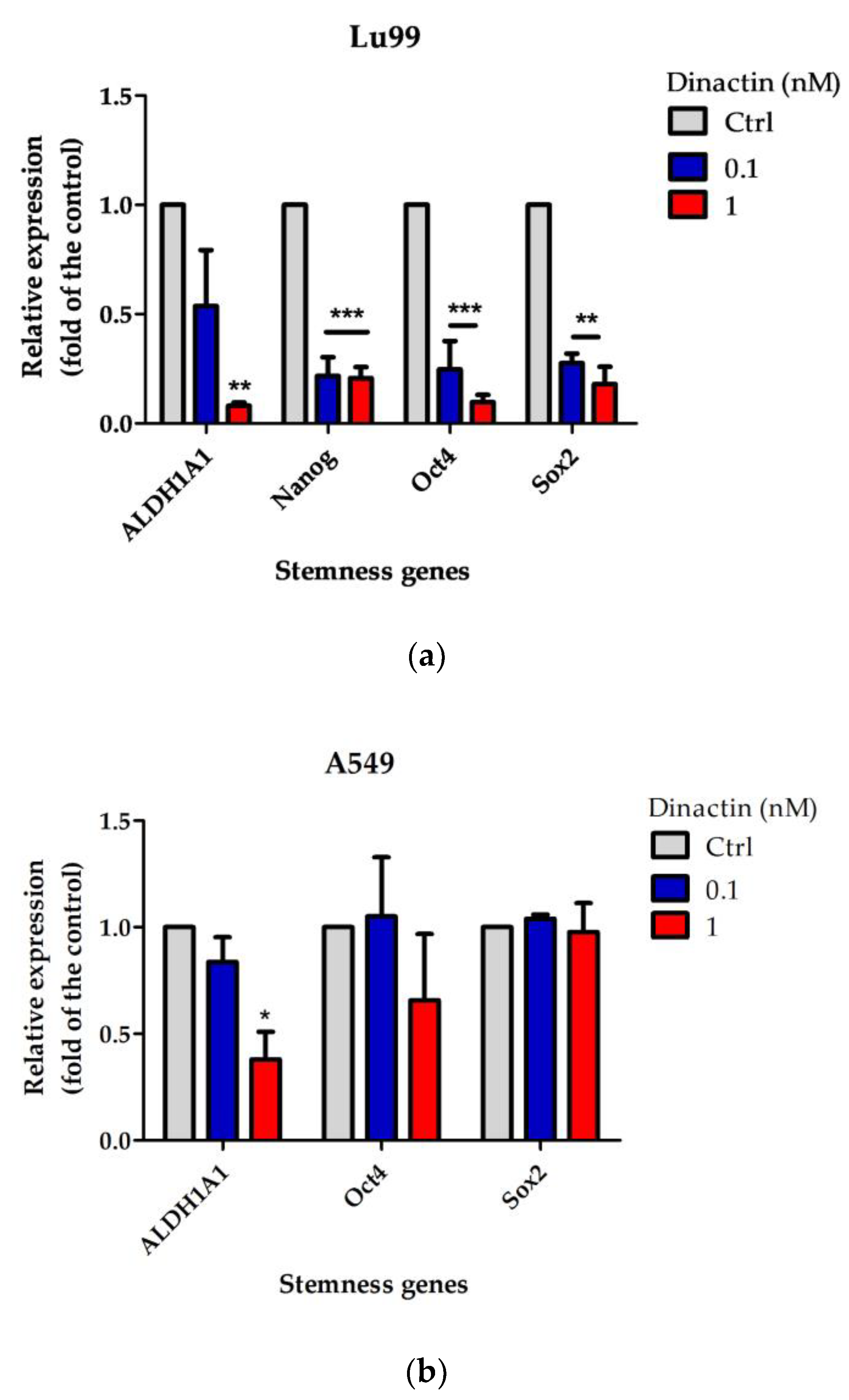
2.4. Effects of Dinactin on Stemness Related Gene Expressions in Lu99 and A549 Tumor-Spheres
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Reagents
4.2. Cell Proliferation and Viability
4.3. Cell Cycle Analysis
4.4. Western Blot Analysis
4.5. Tumor-Sphere Formation
4.6. Quantitative Real-Time RT-PCR (qRT-PCR)
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Duma, N.; Santana-Davila, R.; Molina, J.R. Non-Small Cell Lung Cancer: Epidemiology, Screening, Diagnosis, and Treatment. Mayo Clin. Proc. 2019, 94, 1623–1640. [Google Scholar] [CrossRef] [PubMed]
- Mithoowani, H.; Febbraro, M. Non-Small-Cell Lung Cancer in 2022: A Review for General Practitioners in Oncology. Curr. Oncol. 2022, 29, 1828–1839. [Google Scholar] [CrossRef] [PubMed]
- Reungwetwattana, T.; Oranratnachai, S.; Puataweepong, P.; Tangsujaritvijit, V.; Cherntanomwong, P. Lung Cancer in Thailand. J. Thorac. Oncol. 2020, 15, 1714–1721. [Google Scholar] [CrossRef] [PubMed]
- Rodak, O.; Peris-Díaz, M.D.; Olbromski, M.; Podhorska-Okołów, M.; Dzięgiel, P. Current Landscape of Non-Small Cell Lung Cancer: Epidemiology, Histological Classification, Targeted Therapies, and Immunotherapy. Cancers 2021, 13, 4705. [Google Scholar] [CrossRef]
- Ding, H.; Xin, W.; Tong, Y.; Sun, J.; Xu, G.; Ye, Z.; Rao, Y. Cost effectiveness of immune checkpoint inhibitors for treatment of non-small cell lung cancer: A systematic review. PLoS ONE 2020, 15, e0238536. [Google Scholar] [CrossRef]
- Andrews, A. Treating with Checkpoint Inhibitors-Figure $1 Million per Patient. Am. Health Drug Benefits 2015, 8, 9. [Google Scholar]
- Iqbal, H.; Menaa, F.; Khan, N.U.; Razzaq, A.; Khan, Z.U.; Ullah, K.; Kamal, R.; Sohail, M.; Thiripuranathar, G.; Uzair, B.; et al. Two Promising Anti-Cancer Compounds, 2-Hydroxycinnaldehyde and 2- Benzoyloxycinnamaldehyde: Where do we stand? Comb. Chem. High Throughput Screen. 2022, 25, 808–818. [Google Scholar] [CrossRef]
- Ullah, A.; Ullah, N.; Nawaz, T.; Aziz, T. Molecular mechanisms of Sanguinarine in cancer prevention and treatment. Anti-Cancer Agents Med. Chem. 2022, 22. Published online ahead of print. [Google Scholar] [CrossRef]
- MacDonagh, L.; Gray, S.G.; Breen, E.; Cuffe, S.; Finn, S.P.; O’Byrne, K.J.; Barr, M.P. Lung cancer stem cells: The root of resistance. Cancer Lett. 2016, 372, 147–156. [Google Scholar] [CrossRef]
- Phi, L.T.H.; Sari, I.N.; Yang, Y.G.; Lee, S.H.; Jun, N.; Kim, K.S.; Lee, Y.K.; Kwon, H.Y. Cancer Stem Cells (CSCs) in Drug Resistance and their Therapeutic Implications in Cancer Treatment. Stem Cells Int. 2018, 2018, 5416923. [Google Scholar] [CrossRef] [PubMed]
- Zizka, Z. Biological effects of macrotetrolide antibiotics and nonactic acids. Folia Microbiol. 1998, 43, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Jakubiec-Krzesniak, K.; Rajnisz-Mateusiak, A.; Guspiel, A.; Ziemska, J.; Solecka, J. Secondary Metabolites of Actinomycetes and their Antibacterial, Antifungal and Antiviral Properties. Pol. J. Microbiol. 2018, 67, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jizba, J.; Sedmera, P.; Zima, J.; Beran, M.; Blumauerová, M.; Kandybin, N.V.; Samoukina, G.V. Macrotetrolide antibiotics produced byStreptomyces globisporus. Folia Microbiol. 1991, 36, 437–443. [Google Scholar] [CrossRef]
- Silva, L.J.; Crevelin, E.J.; Souza, W.R.; Moraes, L.A.; Melo, I.S.; Zucchi, T.D. Streptomyces araujoniae Produces a Multiantibiotic Complex with Ionophoric Properties to Control Botrytis cinerea. Phytopathology 2014, 104, 1298–1305. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Dar, M.S.; Bano, N.; Hossain, M.M.; Basit, R.; Bhat, A.Q.; Aga, M.A.; Ali, S.; Hassan, Q.P.; Dar, M.J. Identification of dinactin, a macrolide antibiotic, as a natural product-based small molecule targeting Wnt/β-catenin signaling pathway in cancer cells. Cancer Chemother. Pharmacol. 2019, 84, 551–559. [Google Scholar] [CrossRef]
- Hussain, A.; Rather, M.A.; Dar, M.S.; Dangroo, N.A.; Aga, M.A.; Qayum, A.; Shah, A.M.; Ahmad, Z.; Dar, M.J.; Hassan, Q.P. Streptomyces puniceus strain AS13., Production, characterization and evaluation of bioactive metabolites: A new face of dinactin as an antitumor antibiotic. Microbiol. Res. 2018, 207, 196–202. [Google Scholar] [CrossRef]
- Kaaniche, F.; Hamed, A.; Elleuch, L.; Chakchouk-Mtibaa, A.; Slim, S.; Karray-Rebai, I.; Koubaa, I.; Arcile, G.; Allouche, N.; Mellouli, L. Purification and characterization of seven bioactive compounds from the newly isolated Streptomyces cavourensis TN638 strain via solid-state fermentation. Microb. Pathog. 2020, 142, 104106. [Google Scholar] [CrossRef]
- Zhang, K.; Gu, L.; Zhang, Y.; Liu, Z.; Li, X. Dinactin from a new producer, Streptomyces badius gz-8, and its antifungal activity against the rubber anthracnose fungus Colletotrichum gloeosporioides. Microbiol. Res. 2020, 240, 126548. [Google Scholar] [CrossRef]
- Zhou, J.; He, H.; Wang, X.; Lu, J.; Zhou, X.; Cai, M.; Zhang, Y. Optimization of nutrients for dinactin production by a marine Streptomyces sp. from the high latitude Arctic. Biotechnol. Bioprocess Eng. 2015, 20, 725–732. [Google Scholar] [CrossRef]
- Harinantenaina Rakotondraibe, L.; Rasolomampianina, R.; Park, H.Y.; Li, J.; Slebodnik, C.; Brodie, P.J.; Blasiak, L.C.; Hill, R.; TenDyke, K.; Shen, Y.; et al. Antiproliferative and antiplasmodial compounds from selected Streptomyces species. Bioorg. Med. Chem. Lett. 2015, 25, 5646–5649. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.; Ozsvari, B.; Lisanti, C.L.; Tanowitz, H.B.; Howell, A.; Martinez-Outschoorn, U.E.; Sotgia, F.; Lisanti, M.P. Antibiotics that target mitochondria effectively eradicate cancer stem cells, across multiple tumor types: Treating cancer like an infectious disease. Oncotarget 2015, 6, 4569–4584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karp, I.; Lyakhovich, A. Targeting cancer stem cells with antibiotics inducing mitochondrial dysfunction as an alternative anticancer therapy. Biochem. Pharmacol. 2022, 198, 114966. [Google Scholar] [CrossRef] [PubMed]
- Procópio, R.E.; Silva, I.R.; Martins, M.K.; Azevedo, J.L.; Araújo, J.M. Antibiotics produced by Streptomyces. Braz. J. Infect. Dis. 2012, 16, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Shang, Q.; Li, W.; Guo, W.; Stojadinovic, A.; Mannion, C.; Man, Y.G.; Chen, T. Antibiotics for cancer treatment: A double-edged sword. J. Cancer 2020, 11, 5135–5149. [Google Scholar] [CrossRef]
- Karpiński, T.M.; Adamczak, A. Anticancer Activity of Bacterial Proteins and Peptides. Pharmaceutics 2018, 10, 54. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, B.; Mukherjee, S. Cancer Therapy Using Antibiotics. J. Cancer Ther. 2015, 06, 849–858. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wu, X.; Zhang, Z.; Ma, C.; Wu, T.; Tang, S.; Zeng, Z.; Huang, S.; Gong, C.; Yuan, C.; et al. Monensin inhibits cell proliferation and tumor growth of chemo-resistant pancreatic cancer cells by targeting the EGFR signaling pathway. Sci. Rep. 2018, 8, 17914. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Sperl, B.; Ullrich, A.; Knyazev, P. Metformin and salinomycin as the best combination for the eradication of NSCLC monolayer cells and their alveospheres (cancer stem cells) irrespective of EGFR, KRAS, EML4/ALK and LKB1 status. Oncotarget 2014, 5, 12877–12890. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.G.; Shin, S.J.; Chung, H.W.; Kwon, S.H.; Cha, S.D.; Lee, J.E.; Cho, C.H. Salinomycin reduces stemness and induces apoptosis on human ovarian cancer stem cell. J. Gynecol. Oncol. 2017, 28, e14. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Luo, Q.; Liu, L.; Yang, X.; Zhu, S.; Song, G. Salinomycin attenuates liver cancer stem cell motility by enhancing cell stiffness and increasing F-actin formation via the FAK-ERK1/2 signalling pathway. Toxicology 2017, 384, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Wang, X.; Shang, Y.; Li, Y.; Chen, S.Z. Azithromycin enhances anticancer activity of TRAIL by inhibiting autophagy and up-regulating the protein levels of DR4/5 in colon cancer cells in vitro and in vivo. Cancer Commun. 2018, 38, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parajuli, B.; Shin, S.J.; Kwon, S.H.; Cha, S.D.; Chung, R.; Park, W.J.; Lee, H.G.; Cho, C.H. Salinomycin induces apoptosis via death receptor-5 up-regulation in cisplatin-resistant ovarian cancer cells. Anticancer Res. 2013, 33, 1457–1462. [Google Scholar] [PubMed]
- Dijk, S.N.; Protasoni, M.; Elpidorou, M.; Kroon, A.M.; Taanman, J.-W. Mitochondria as target to inhibit proliferation and induce apoptosis of cancer cells: The effects of doxycycline and gemcitabine. Sci. Rep. 2020, 10, 4363. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.; Ha, J.; Kang, E.; Cho, S. The role of epithelial-mesenchymal transition-regulating transcription factors in anti-cancer drug resistance. Arch. Pharm. Res. 2021, 44, 281–292. [Google Scholar] [CrossRef]
- Shapiro, G.I.; Harper, J.W. Anticancer drug targets: Cell cycle and checkpoint control. J. Clin. Investig. 1999, 104, 1645–1653. [Google Scholar] [CrossRef] [Green Version]
- Tan, Q.; Yan, X.; Song, L.; Yi, H.; Li, P.; Sun, G.; Yu, D.; Li, L.; Zeng, Z.; Guo, Z. Induction of Mitochondrial Dysfunction and Oxidative Damage by Antibiotic Drug Doxycycline Enhances the Responsiveness of Glioblastoma to Chemotherapy. Med. Sci. Monit. 2017, 23, 4117–4125. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, H.; Yoshida, T.; Horinaka, M.; Yasuda, T.; Goda, A.E.; Konishi, M.; Wakada, M.; Kataoka, K.; Yoshikawa, T.; Sakai, T. Baicalein overcomes tumor necrosis factor-related apoptosis-inducing ligand resistance via two different cell-specific pathways in cancer cells but not in normal cells. Cancer Res. 2008, 68, 8918–8927. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Dai, J.; Huang, W.; Jiao, Y.; Liu, L.; Wu, M.; Tan, D. hMTERF4 knockdown in HeLa cells results in sub-G1 cell accumulation and cell death. Acta Biochim. Biophys. Sin. 2011, 43, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Dutta, A. p21 in cancer: Intricate networks and multiple activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef]
- Iida, K.; Sakai, R.; Yokoyama, S.; Kobayashi, N.; Togo, S.; Yoshikawa, H.Y.; Rawangkan, A.; Namiki, K.; Suganuma, M. Cell softening in malignant progression of human lung cancer cells by activation of receptor tyrosine kinase AXL. Sci. Rep. 2017, 7, 17770. [Google Scholar] [CrossRef] [PubMed]
- Meng, E.; Mitra, A.; Tripathi, K.; Finan, M.A.; Scalici, J.; McClellan, S.; Madeira da Silva, L.; Reed, E.; Shevde, L.A.; Palle, K.; et al. ALDH1A1 maintains ovarian cancer stem cell-like properties by altered regulation of cell cycle checkpoint and DNA repair network signaling. PLoS ONE 2014, 9, e107142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabakov, A.; Yakimova, A.; Matchuk, O. Molecular Chaperones in Cancer Stem Cells: Determinants of Stemness and Potential Targets for Antitumor Therapy. Cells 2020, 9, 892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaksch, M.; Múnera, J.; Bajpai, R.; Terskikh, A.; Oshima, R.G. Cell cycle-dependent variation of a CD133 epitope in human embryonic stem cell, colon cancer, and melanoma cell lines. Cancer Res. 2008, 68, 7882–7886. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Gong, P.; Li, J.; Fu, Y.; Zhou, Z.; Liu, L. Role of CD133 in human embryonic stem cell proliferation and teratoma formation. Stem Cell Res. Ther. 2020, 11, 208. [Google Scholar] [CrossRef]
- Zhang, X.; Neganova, I.; Przyborski, S.; Yang, C.; Cooke, M.; Atkinson, S.P.; Anyfantis, G.; Fenyk, S.; Keith, W.N.; Hoare, S.F.; et al. A role for NANOG in G1 to S transition in human embryonic stem cells through direct binding of CDK6 and CDC25A. J. Cell Biol. 2009, 184, 67–82. [Google Scholar] [CrossRef]
- Lu, Y.; Qu, H.; Qi, D.; Xu, W.; Liu, S.; Jin, X.; Song, P.; Guo, Y.; Jia, Y.; Wang, X.; et al. OCT4 maintains self-renewal and reverses senescence in human hair follicle mesenchymal stem cells through the downregulation of p21 by DNA methyltransferases. Stem Cell Res. Ther. 2019, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Świstowska, M.; Gil-Kulik, P.; Krzyzanowski, A.; Bielecki, T.; Czop, M.; Kwasniewska, A.; Kocki, J. Potential Effect of SOX2 on the Cell Cycle of Wharton’s Jelly Stem Cells (WJSCs). Oxidative Med. Cell. Longev. 2019, 2019, 5084689. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, H.; Zhu, Y.; Wu, Z.; Cui, C.; Cai, F. Anticancer Mechanisms of Salinomycin in Breast Cancer and Its Clinical Applications. Front. Oncol. 2021, 11, 2748. [Google Scholar] [CrossRef]
- Qi, D.; Liu, Y.; Li, J.; Huang, J.H.; Hu, X.; Wu, E. Salinomycin as a potent anticancer stem cell agent: State of the art and future directions. Med. Res. Rev. 2022, 42, 1037–1063. [Google Scholar] [CrossRef]
- Rawangkan, A.; Wongsirisin, P.; Namiki, K.; Iida, K.; Kobayashi, Y.; Shimizu, Y.; Fujiki, H.; Suganuma, M. Green Tea Catechin Is an Alternative Immune Checkpoint Inhibitor that Inhibits PD-L1 Expression and Lung Tumor Growth. Molecules 2018, 23, 2071. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Hirano, K.; Takahashi, A.; Yamaguchi, K.; Beppu, M.; Fujiki, H.; Suganuma, M. Nucleolin on the cell surface as a new molecular target for gastric cancer treatment. Biol. Pharm. Bull. 2010, 33, 796–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okabe, S.; Suganuma, M.; Hayashi, M.; Sueoka, E.; Komori, A.; Fujiki, H. Mechanisms of growth inhibition of human lung cancer cell line, PC-9, by tea polyphenols. Jpn. J. Cancer Res. 1997, 88, 639–643. [Google Scholar] [CrossRef]
- Namiki, K.; Wongsirisin, P.; Yokoyama, S.; Sato, M.; Rawangkan, A.; Sakai, R.; Iida, K.; Suganuma, M. (−)-Epigallocatechin gallate inhibits stemness and tumourigenicity stimulated by AXL receptor tyrosine kinase in human lung cancer cells. Sci. Rep. 2020, 10, 2444. [Google Scholar] [CrossRef] [Green Version]
- Tozuka, K.; Wongsirisin, P.; Nagai, S.E.; Kobayashi, Y.; Kanno, M.; Kubo, K.; Takai, K.; Inoue, K.; Matsumoto, H.; Shimizu, Y.; et al. Deregulation of protein phosphatase 2A inhibitor SET is associated with malignant progression in breast cancer. Sci. Rep. 2021, 11, 14238. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dinactin (nM) | Average Size of Tumor Spheres (µm) | |
|---|---|---|
| Lu99 | A549 | |
| Control | 175.54 ± 55.78 | 158.72 ± 47.40 |
| 0.1 | 181.04 ± 46.43 | 159.28 ± 43.98 |
| 1 | 128.82 ± 20.07 *** | 134.87 ± 25.71 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rawangkan, A.; Wongsirisin, P.; Pook-In, G.; Siriphap, A.; Yosboonruang, A.; Kiddee, A.; Chuerduangphui, J.; Reukngam, N.; Duangjai, A.; Saokaew, S.; et al. Dinactin: A New Antitumor Antibiotic with Cell Cycle Progression and Cancer Stemness Inhibiting Activities in Lung Cancer. Antibiotics 2022, 11, 1845. https://doi.org/10.3390/antibiotics11121845
Rawangkan A, Wongsirisin P, Pook-In G, Siriphap A, Yosboonruang A, Kiddee A, Chuerduangphui J, Reukngam N, Duangjai A, Saokaew S, et al. Dinactin: A New Antitumor Antibiotic with Cell Cycle Progression and Cancer Stemness Inhibiting Activities in Lung Cancer. Antibiotics. 2022; 11(12):1845. https://doi.org/10.3390/antibiotics11121845
Chicago/Turabian StyleRawangkan, Anchalee, Pattama Wongsirisin, Grissana Pook-In, Achiraya Siriphap, Atchariya Yosboonruang, Anong Kiddee, Jureeporn Chuerduangphui, Nanthawan Reukngam, Acharaporn Duangjai, Surasak Saokaew, and et al. 2022. "Dinactin: A New Antitumor Antibiotic with Cell Cycle Progression and Cancer Stemness Inhibiting Activities in Lung Cancer" Antibiotics 11, no. 12: 1845. https://doi.org/10.3390/antibiotics11121845
APA StyleRawangkan, A., Wongsirisin, P., Pook-In, G., Siriphap, A., Yosboonruang, A., Kiddee, A., Chuerduangphui, J., Reukngam, N., Duangjai, A., Saokaew, S., & Praphasawat, R. (2022). Dinactin: A New Antitumor Antibiotic with Cell Cycle Progression and Cancer Stemness Inhibiting Activities in Lung Cancer. Antibiotics, 11(12), 1845. https://doi.org/10.3390/antibiotics11121845